Peer review process
Revised: This Reviewed Preprint has been revised by the authors in response to the previous round of peer review; the eLife assessment and the public reviews have been updated where necessary by the editors and peer reviewers.
Read more about eLife’s peer review process.Editors
- Reviewing EditorAdrien PeyracheMcGill University, Montreal, Canada
- Senior EditorLaura ColginUniversity of Texas at Austin, Austin, United States of America
Reviewer #1 (Public review):
Summary:
The manuscript by Yang et al. investigates the relationship between multi-unit activity in the locus coeruleus, putatively noradrenergic locus coeruleus, hippocampus (HP) sharp-wave ripples (SWR) and spindles using multi-site electrophysiology in freely behaving male rats. The study focuses on SWR during quiet wake and non-REM sleep, and their relation to cortical states (identified using EEG recordings in frontal areas) and LC units.
The manuscript highlights differential modulation of LC units as a function of HP-cortical communication during wake and sleep. They establish that ripples and LC units are inversely correlated to levels of arousal: wake, i.e. higher arousal correlates with higher LC unit activity and lower ripple rates. The authors show that LC neuron activity is strongly inhibited just before SWR detected during wake. During non-REM sleep, they distinguish "isolated" ripples from SWR coupled to spindles and show that inhibition of LC neuron activity is absent before spindle-coupled ripples but not before isolated ripples, suggesting a mechanism where noradrenaline (NA) tone is modulated by HP-cortical coupling. This result has interesting implications for the roles of noradrenaline in the modulation of sleep-dependent memory consolidation, as ripple-spindle coupling is a mechanism favoring consolidation. The authors further show that NA neuronal activity is downregulated before spindles.
Strengths:
In continuity with previous work from the laboratory, this work expands our understanding of the activity of neuromodulatory systems in relation to vigilance states and brain oscillations, an area of research that is timely and impactful. The manuscript presents strong results suggesting that NA tone varies differentially depending on coupling of HP SWR with cortical spindles. The authors place their findings back in the context of identified roles of HP ripples and coupling to cortical oscillations for memory formation in a very interesting discussion. The distinction of LC neuron activity between awake, ripple-spindle coupled events and isolated ripples is an exciting result and its relation to arousal and memory opens fascinating lines of research.
Weaknesses:
I regretted that the paper fell short of trying to push this line of idea a bit further, for example by contrasting in the same rats the LC unit-HP ripple coupling during exploration of a highly familiar context (as seemingly was the case in their study) versus a novel context, which would increase arousal and trigger memory-related mechanisms. Any kind of manipulation of arousal levels and investigation of the impact on awake vs nonREM sleep LC-HP ripple coordination would considerably strengthen the scope of the study.
Comments on revised version.
The authors have added methodological details to the results section after the first round of reviews, improving the manuscript readability. Some points might still be improved, for example, the authors use a delta/gamma ratio to track cortical states for example, but there is no methods section corresponding to this metric. Authors write that higher SI corresponds to a lower arousal state that is associated with "more synchronized cortical population activity, higher ripple rate and reduced LC neurons firing" but there are no references or analysis to support this statement, only examples showing changes in SI over a few minutes.
Reviewer #2 (Public review):
Summary:
In this study, authors studied the synchrony between ripple events in Hippocampus, cortical spindles and Locus Coeruleus spiking. The results in this study together with the established literature on the relationship of hippocampal ripples with widespread thalamic and cortical waves, guided authors to propose a role for Locus Coeruleus spiking patterns in memory consolidation. The findings provided here, i.e. correlations between LC spiking activity and Hippocampal ripples, could provide basis for future studies probing the directional flow or the necessity of these correlations in the memory consolidation process. Hence, the paper provides enough scientific advance to highlight the elusive yet important role of Norepinephrine circuitry in the memory processes.
Strengths:
Authors were able to demonstrate correlations of Locus Coeruleus spikes with hippocampal ripples as well as with cortical spindles. Specific strength of the paper is in the demonstration that the spindles that activate with the ripples are comparatively different in their correlations with Locus Coeruleus than those which do not.
Weaknesses:
The claims regarding the roles of these specific interactions were mostly derived from the literature that these processes individually contribute to the memory process, without any evidence of these specific interactions being necessary for memory processes. There are also issues with the description of methods, validation of shuffling procedures and unclear presentation and the interpretation of the findings, which are described in points that follow. I believe addressing these weaknesses might improve and add to the strength of the findings.
Comments on revised version.
The authors addressed all of my major concerns during the revision. As a result, the study now provides convincing evidence as well as improved presentation of results, that makes this manuscript important to the broader field of neuroscience, beyond the specific sub-field.
Reviewer #3 (Public review):
This manuscript examines how locus coeruleus (LC) activity relates to hippocampal ripple events across behavioral states in freely moving rats. Using multi-site electrophysiological recordings, the authors report that LC activity is suppressed prior to ripple events, with the magnitude of suppression depending on ripple subtype. Suppression is stronger during wakefulness than during NREM sleep and least pronounced for ripples coupled to spindles.
The study is technically sound and addresses a timely and important question regarding how LC activity interacts with hippocampal and thalamocortical network events across vigilance states. While the findings are interesting, they remain observational in nature. Following revision, the manuscript has substantially improved in both presentation and interpretation of the results, and most concerns have been addressed satisfactorily. I therefore only have a few minor considerations that the authors may wish to explore further in the current study or in future work, as these directions could provide additional mechanistic insight and would likely be of considerable interest to the field.
The authors demonstrate clearly that tonic LC firing rates preceding ripples differ significantly between wake-associated ripples (highest LC firing), isolated ripples during NREM sleep (lower LC firing), and spindle-coupled ripples (lowest LC firing). They also appropriately note that baseline firing differences will naturally influence the magnitude of LC suppression, which they also observe (highest LC reduction for wake ripples, then isolated ripples and last spindle-coupled ripples). However, this aspect could be explored further, as it may provide additional insight into the regulation of spindle-associated ripple events. Since LC activity appears to decline gradually prior to ripple occurrence (Suppl. Figure 2), it would be interesting to test whether this gradual reduction helps organize the emergence of isolated versus spindle-coupled ripples. For example, isolated ripples may occur during the initial phase of LC decline, whereas spindle-coupled ripples may preferentially emerge when LC activity reaches its lowest levels. Such a relationship could also be consistent with the stronger synchronization observed for spindle-ripple coupling.
Related to this point, it would also be informative to examine whether isolated spindles occur more randomly in time, whereas spindle-associated ripple events appear more temporally clustered. If a single isolated spindle occurs, the associated LC suppression might be more pronounced. In contrast, when multiple spindle-associated ripple events occur in succession, LC activity may already be reduced following the first event, resulting in smaller additional suppression preceding subsequent events. Exploring this possibility could help clarify how LC dynamics shape the temporal emergence of ripple-subtypes
Author response:
The following is the authors’ response to the original reviews.
Reviewer #1 (Public review):
Summary:
The manuscript by Yang et al. investigates the relationship between multi-unit activity in the locus coeruleus, putatively noradrenergic locus coeruleus, hippocampus (HP), sharp-wave ripples (SWR), and spindles using multi-site electrophysiology in freely behaving male rats. The study focuses on SWR during quiet wake and non-REM sleep, and their relation to cortical states (identified using EEG recordings in frontal areas) and LC units.
The manuscript highlights differential modulation of LC units as a function of HP-cortical communication during wake and sleep. They establish that ripples and LC units are inversely correlated to levels of arousal: wake, i.e., higher arousal correlates with higher LC unit activity and lower ripple rates. The authors show that LC neuron activity is strongly inhibited just before SWR is detected during wake. During non-REM sleep, they distinguish "isolated" ripples from SWR coupled to spindles and show that inhibition of LC neuron activity is absent before spindle-coupled ripples but not before isolated ripples, suggesting a mechanism where noradrenaline (NA) tone is modulated by HP-cortical coupling. This result has interesting implications for the roles of noradrenaline in the modulation of sleep-dependent memory consolidation, as ripple-spindle coupling is a mechanism favoring consolidation. The authors further show that NA neuronal activity is downregulated before spindles.
Strengths:
In continuity with previous work from the laboratory, this work expands our understanding of the activity of neuromodulatory systems in relation to vigilance states and brain oscillations, an area of research that is timely and impactful. The manuscript presents strong results suggesting that NA tone varies differentially depending on the coupling of HP SWR with cortical spindles. The authors place their findings back in the context of identified roles of HP ripples and coupling to cortical oscillations for memory formation in a very interesting discussion. The distinction of LC neuron activity between awake, ripple-spindle coupled events and isolated ripples is an exciting result, and its relation to arousal and memory opens fascinating lines of research.
Weaknesses:
I regretted that the paper fell short of trying to push this line of idea a bit further, for example, by contrasting in the same rats the LC unit-HP ripple coupling during exploration of a highly familiar context (as seemingly was the case in their study) versus a novel context, which would increase arousal and trigger memory-related mechanisms. Any kind of manipulation of arousal levels and investigation of the impact on awake vs non-REM sleep LC-HP ripple coordination would considerably strengthen the scope of the study.
We agree that conducting specific behavioral tests before electrophysiological recordings, as well as manipulating arousal during the recording session, would strengthen the study. These experiments are planned for future work, and we acknowledged this point in the discussion.
We added the following text in the Discussion: “Conducting behavioral assays prior to electrophysiological recordings, along with spatially and temporally precise modulation of LC activity during recording sessions, will be essential for achieving a mechanistic understanding of network dynamics and its functional role for memory consolidation in future investigations.”
The main result shows that LC units are not modulated during non-REM sleep around spindle-coupled ripples (named spRipples, 17.2% of detected ripples); they also show that LC units are modulated around ripple-coupled spindles (ripSpindles, proportion of detected spindles not specified, please add). These results seem in contradiction; this point should be addressed by the authors.
The detection of coupled events - spindle-coupled ripples (spRipple) and ripple-coupled spindles (ripSpindle) - was performed independently, although, some overlap cannot be excluded. We found that LC suppression was generally weak around both types of coupled events. Specifically, LC suppression around spRipples and ripSpindles reached significance (exceeding the 95% confidence interval) in 4 sessions (from 3 rats) and 3 sessions (from 2 rats), respectively, out of a total of 20 sessions (from 7 rats).
We revised the manuscript by providing additional information in the Results section and adding a Supplementary Figure 5 showing a significant correlation (Pearson r = 0.72, p = 0.0003) between the modulation index (MI) for spRipple and ripSpindle.
Results are displayed per recording session, with 20 sessions total recorded from 7 rats (2 to 8 sessions per rat), which implies that one of the rats accounts for 40% of the dataset. Authors should provide controls and/or data displayed as average per rat to ensure that results are now skewed by the weight of that single rat in the results.
High-quality recordings from the LC in behaving rats are technically challenging and relatively rare; therefore, we included all valid datasets in analysis. The average modulation index (MI), calculated per animal and per session, fell within a consistent range (Supplementary Figure 3) despite variability in the number of recording sessions (2–8 sessions per rat).
In its current form, the manuscript presents a lack of methodological detail that needs to be addressed, as it clouds the understanding of the analysis and conclusions. For example, the method to account for the influence of cortical state on LC MUA is unclear, both for the exact methods (shuffling of the ripple or spindle onset times) and how this minimizes the influence of cortical states; this should be better described. If the authors wish to analyze unit modulation as a function of cortical state, could they also identify/sort based on cortical states and then look at unit modulation around ripple onset? For the first part of the paper, was an analysis performed on quiet wake, non-REM sleep, or both?
The LC activity around rippled was modulated at multiple temporal scales. First, we observed a relatively sharp drop in the LC firing rate ~ 2 s before the ripple onset. When computing peri-ripple LC activity over a longer time window ([–12, 12] sec), we observed a rather slow decrease in the LC firing rate beginning as early as 10 s before the ripple onset (Supplementary Figure 2).
Considering two temporal scales, we hypothesized that slow modulation of LC activity might be related to fluctuations of the global brain state. We quantified the ongoing cortical state using a synchronization index (SI), calculated as a power ratio (1–4 Hz/30–90 Hz) of the EEG within 4-s windows and computed the corresponding ripple and LC-MUA rates. Figure 3A (in the main manuscript) illustrates that a higher SI (more synchronized cortical population activity) corresponded to a lower arousal state and reduced LC tonic firing; this brain state was associated with a higher ripple activity. As shown in the new Figure 3B, the LC firing rate was negatively correlated with the SI and ripple rate. Thus, slow LC modulation was likely driven by cortical state transitions.
To correct for the influence of the global brain state on the peri-ripple LC activity, we generated surrogate events by jittering the times of detected ripples. First, we confirmed that triggering the hippocampal LFP on the surrogate events lacked the ripple-specific frequency component (main Figure 3C) and the SI state did not differ around ripples and surrogate events (main Figure 3D). Plotting the LC activity around surrogate evens captured its state-dependent dynamics (Figure 3 or Supplementary Figure 2, orange trace). To extract state-independent peri-ripple LC modulation, we subtracted the state-related LC activity (orange trace) from the ripple-triggered LC activity (blue trace). The resulting trace yielded a corrected estimate of ripple-associated LC activity that was largely free from the confounding influence of cortical state transitions (main Figure 3E).
In the Results subsection “LC-NE neuron spiking is suppressed around hippocampal ripples”, we reported LC modulation without accounting for the cortical state (main Figure 2). The state-dependent effects were instead examined in the subsequent Results subsection, “LC firing and ripple occurrence are state-dependent and inversely related” we report state-corrected LC modulation (main Figure 3). Finally, in the Results subsection “Peri-ripple LC modulation depends on the cortical–hippocampal interaction,” we characterized LC activity around ripples across different cortical states (quite awake and NREM sleep).
We revised Methods and Results to provide more methodological details and a rationale for each analysis, as requested.
Reviewer #2 (Public review):
Summary:
In this study, the authors studied the synchrony between ripple events in the Hippocampus, cortical spindles, and Locus Coeruleus spiking. The results in this study, together with the established literature on the relationship of hippocampal ripples with widespread thalamic and cortical waves, guided the authors to propose a role for Locus Coeruleus spiking patterns in memory consolidation. The findings provided here, i.e., correlations between LC spiking activity and Hippocampal ripples, could provide a basis for future studies probing the directional flow or the necessity of these correlations in the memory consolidation process. Hence, the paper provides enough scientific advances to highlight the elusive yet important role of Norepinephrine circuitry in the memory processes.
Strengths:
The authors were able to demonstrate correlations of Locus Coeruleus spikes with hippocampal ripples as well as with cortical spindles. A specific strength of the paper is in the demonstration that the spindles that activate with the ripples are comparatively different in their correlations with Locus Coeruleus than those that do not.
Weaknesses:
The claims regarding the roles of these specific interactions were mostly derived from the literature that these processes individually contribute to the memory process, without any evidence of these specific interactions being necessary for memory processes. There are also issues with the description of methods, validation of shuffling procedures, and unclear presentation and the interpretation of the findings, which are described in the points that follow. I believe addressing these weaknesses might improve and add to the strength of the findings.
We believe that our responses to the Reviewer 1 and Reviewer 2, corresponding revisions of the manuscript and new figures adequately addressed all issues raised by the Reviewer 2.
Reviewer #3 (Public review):
Summary:
This manuscript examines how locus coeruleus (LC) activity relates to hippocampal ripple events across behavioral states in freely moving rats. Using multi-site electrophysiological recordings, the authors report that LC activity is suppressed prior to ripple events, with the magnitude of suppression depending on the ripple subtype. Suppression is stronger during wakefulness than during NREM sleep and is least pronounced for ripples coupled to spindles.
The study is technically competent and addresses an important question regarding how LC activity interacts with hippocampal and thalamocortical network events across vigilance states.
Weaknesses:
The results are interesting, but entirely observational. Also, the study in its current form would benefit from optimization of figure labeling and presentation, and more detailed result descriptions to make the findings fully interpretable. Also, it would be beneficial if the authors could formulate the narrative and central hypothesis more clearly to ease the line of reasoning across sections.
We improved the presentation of results by incorporating additional figures and expanding the detail in the figure captions. In the main text, we clarified specific hypotheses and provided a rationale underlying each analysis.
Comments:
(1) Stronger evidence that recorded units represent noradrenergic LC neurons would reinforce the conclusions. While direct validation may not be possible, showing absolute firing rates (Hz) across quiet wake, active wake, NREM, and REM, and comparing them to published LC values, would help.
We added the requested data and a Supplementary Figure 1 in the revised manuscript: “The average firing rates of LC single units were 1.70 ± 0.21 Hz during wakefulness, 0.51 ± 0.07 Hz during NREM sleep, and 0.014 ± 0.01 Hz during REM sleep (Supplementary Figure 1). Firing rates differed significantly across arousal states, with the highest activity during wakefulness, reduced activity during NREM sleep, and minimal activity during REM sleep (one-way ANOVA: F(2,38) = 39.8, p < 0.0001). This firing pattern is characteristic of LC-NE neurons and is consistent with existing literature.”
(2) The analyses rely almost exclusively on z-scored LC firing and short baselines (~4-6 s), which limits biological interpretation. The authors should include absolute firing rates alongside normalized values for peri-ripple and peri-spindle analyses and extend pre-event windows to at least 20-30 s to assess tonic firing evolution. This would clarify whether differences across ripple subtypes arise from ceiling or floor effects in LC activity; if ripples require LC silence, the relative drop will appear larger during high-firing wake states. This limitation should be discussed and, if possible, results should be shown based on unnormalized firing rates.
We agree with the reviewer that a longer pre-event window provides a clearer estimate of baseline LC activity. However, given that both ripples and spindles are brief oscillatory events, we tested a range of time windows and found that a 12-s interval adequately captures baseline LC activity dynamics. Accordingly, we included plots with extended pre-event windows (−12 to 12 s), as requested.
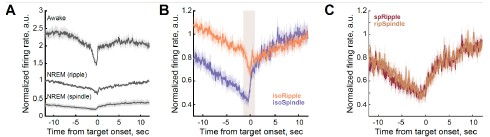
We added in the revised manuscript absolute firing rates for well-isolated LC single units. Because the number of neurons contributing to LC multi-unit activity (LC-MUA) is unknown, we avoided averaging absolute firing rates for this signal. For LC-MUA, we implemented a normalization approach in which firing rates (50-ms bins) around ripple or spindle are scaled to a baseline period preceding the trigger event (−12 to −10 s). Importantly, unlike z-scoring, this normalization method preserves baseline differences across behavioral states. As shown in Author response image 1A and new Figure 5 in the main manuscript, baseline LC firing rates were highest prior to awake ripples and lowest prior to sleep spindles. During ripples occurring in wakefulness, LC activity did not decrease to the levels observed during sleep. In contrast, during NREM sleep, LC activity was downregulated during both ripples and spindles, although it did not reach complete silence around either oscillatory event.
Author response image 1B illustrates a slow downward drift in the LC firing rate preceding either ripple or spindle. The slow LC dynamics likely reflected gradual transitions toward more synchronized brain state, which is optimal for ripple generation. In contrast, event-specific LC modulation had faster dynamics (Author response image 1B, highlighted interval) and was largely absent in cases where spRipples and ripSpindles were not associated with LC suppression (Author response image 1C).
To minimize the influence of global state fluctuations and emphasize event-related dynamics, we therefore presented the main results using state-corrected and z-scored PETHs.
Please also refer to our response to Reviewer 1 regarding the two temporal scales of LC modulation.
Author response image 1.
LC modulation around sleep oscillations. (A) Peri-event LC-MUA during awake and NREM sleep. LC activity and the range of peri-event LC modulation differed across behavioral states; it was overall higher preceding ripples occurring in wakefulness than in NREM sleep, and it was the lowest around sleep spindles. Despite the state-dependent differences in the firing rate, LC modulation was observed around all oscillatory events. During wakefulness, LC activity did not decrease to the levels observed during NREM sleep. During NREM sleep, LC activity was down-regulated around both ripples and spindles, and the LC firing did not completely cease around either oscillatory event. (B) Peri-event LC-MUA around isolated oscillatory events. LC activity exhibited fast peri-event dynamics (highlighted interval) superimposed on slower, state-dependent fluctuations. (C) Peri-event LC-MUA around coupled oscillatory events. Fast peri-event LC modulation was absent, while slow fluctuations were preserved around coupled oscillatory events. For all plots, LC-MUA firing rate was scaled to a pre-event baseline interval [-12 to -10 sec] to preserve baseline differences in LC activity across behavioral states. Bin size: 50 ms. isoRipple – isolated ripple, isoSpindle – isolated spindle, spRipple - spindle-coupled ripple, ripSpindle - ripple-coupled spindle.}
(3) Because spindles often occur in clusters, the timing of ripple occurrence within these clusters could influence LC suppression. Indicate whether this structure was considered or discuss how it might affect interpretation (e.g., first vs. subsequent ripples within a spindle cluster).
We did not consider spindle clusters and classified the event as ripple-coupled spindle if the ripple occurred between the spindle on and offset.
(4) While the observational approach is appropriate here, causal tests (e.g., optogenetic or chemogenetic manipulation of LC around ripple events and in memory tasks) would considerably strengthen the mechanistic conclusions. At a minimum, a discussion of how such approaches could address current open questions would improve the manuscript.
We agree that conducting causal tests would strengthen the study. We added the following text in the Discussion: “Conducting behavioral assays prior to electrophysiological recordings, along with spatially and temporally precise modulation of LC activity during recording sessions, will be essential for achieving a mechanistic understanding of network dynamics and its functional role for memory consolidation in future investigations.”
(5) Please show how "Synchronization Index" (SI) differs quantitatively across behavioral states (wake, NREM, REM) and discuss whether it could serve as a state classifier. This would strengthen interpretations of the correlations between SI, ripple occurrence, and LC activity.
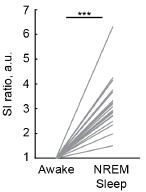
We plotted the awake state-normalized SIs for awake and NREM sleep. Due to small number of REM sleep episodes, SI for REM sleep is not shown. The average SI during NREM sleep was significantly higher than during awake state, consistent with the well-established dominance of low-frequency (1-4 Hz) oscillatory power and reduced high-frequency (30-90 Hz) power during NREM sleep.
Although SI could potentially serve as a behavioral state classifier, we have chosen not to address this point to maintain the focus in the discussion on new results.
Author response image 2.
Synchronization index differentiates behavioral states.
(6) The current use of SI to denote a delta/gamma power ratio is unconventional, as "SI" typically refers to phase-locking metrics. Consider adopting a more standard term, such as delta/gamma power ratio. Similarly, it would be easier to follow if you use common terminology (AUC) to describe the drop in LC-MUA rather than using "MI" and "sub-MI".
The ranges of delta and gamma bands might vary across studies; therefore, we prefer using SI, as defined here and in our previous publications (Novitskaya et al., 2016; Yang et al., 2019, 2021). We calculated the modulation index (MI) as the area under the curve of the peri-event time histogram within the 1 second preceding ripple onset. To avoid potential confusion with the AUC calculated over the entire signal window, we opted to use MI.
(7) The logic in Figure 3 is difficult to follow. The brain state (delta/gamma ratio) appears unchanged relative to surrogate events (3C), while LC activity that is supposedly negatively correlated to delta/gamma changes markedly (3D-E). Could this discrepancy reflect the low temporal resolution (4-s windows) used to calculate delta/gamma when the changes occur on a shorter time scale?
We appreciate the reviewer’s question. We revised the results and Figure 3 legend to clarify this point. The main Figures 3E and 3F show the 'state-corrected' peri-ripple LC activity. The purpose of generating ‘surrogate’ events was precisely to capture the component of LC activity dynamics that can be explained by cortical state fluctuations alone. As shown in Supplementary Figure 2, the orange trace represents LC activity aligned to surrogate events and, as the Reviewer noted, shows a clear decrease, yet at a slower time scale. We interpret this surrogate-aligned signal as the LC modulation attributable specifically to cortical state fluctuations. Importantly, shuffled events were associated with similar SIs (cortical state), but absent HPC LFP power increase in the ripple range (140-250 Hz), as shown in the main Figures 3C and 3D, respectively. To isolate the peri-event LC dynamics, we subtracted the state-related component (Figure 3, orange trace) from the ripple-triggered LC activity (blue trace). This correction yielded an estimate of ripple-associated LC activity that is largely independent of the confounding influence of ongoing cortical state.
Please, see our detailed response to the Reviewer 1 about multiple time scales of LC dynamics.
(8) There are apparent inconsistencies between Figures 4B and 4C-D. In B, it seems that the difference between the 10th and 90th percentile is mostly in higher frequencies, but in C and D, the only significant difference is in the delta band.
We repeated this analysis, clarified inconsistency, and revised Figure 4 legend.
(9) Because standard sleep scoring is based on EEG and EMG signals, please include an example of sleep scoring alongside the data used for state classification. It would also be relevant to include the delta/gamma power ratio in such an example plot.
We replaced ‘standard’ with ‘previously established” sleep scoring procedure and added a Supplementary Figure 4 showing representative NREM sleep and wake episodes with corresponding EEG and SI.
(10) Can variability in modulation index (subMI) across ripple subsets reflect differences in recording quality? Please report and compare mean LC firing rates across subsets to confirm this is not a confounding factor.
We agree that considering recording quality and unit stability over time as potential confounding factors is important. We therefore carefully evaluated each dataset to ensure the absence of significant drift in the LC firing rate. However, we find that comparing mean LC firing rates across subsets of ripples, as suggested by the Reviewer, is insufficient to control for recording stability, as LC activity varies substantially across behavioral states. At present, we are not aware of a robust method to fully eliminate variability related to recording quality and unit stability over time.
(11) Figure 6B: If the brown trace represents LC-MUA activity around random time points, why would there be a coinciding negative peak as relative to real sleep spindles? Or is it the subtracted trace?
We have revised Figure 7 (original Figure 6) and its legend to improve clarity and readability.
(12) On page 8, lines 207-209, the authors write "Importantly, neither the LC-MUA rate nor SIs differed during a 2-sec time window preceding either group of spindles". It is unclear which data they refer to, but the statement seems to contradict Figure 6E as well as the following sentence: "Across sessions, MI values exceeded 95% CI in 17/20 datasets for isoSpindles and only 3/20 for ripSpindles". This should be clarified.
We have revised the corresponding text to improve clarity and readability.
(13) The results in Figures 5C and 6F do not align. It seems surprising that ripple-coupled spindles show a considerably higher LC modulation than spindle-coupled ripples, as these events should overlap. Could the discrepancy be due to Z-score normalization as mentioned above? Please include a discussion of this to help the interpretation of the results.
In the original manuscript, Figure 6F was mistakenly labelled for ripple-coupled (ripSpindles) and isolated (isoSpindles) spindles. Now it has been corrected.
Please, also see our response to the Reviewer 1 weaknesses.
(14) The text implies that 8 recordings came from one rat and two each from six others. This should be confirmed, and it should be explained how the recordings were balanced and analyzed across animals.
Since high-quality recordings from LC in behaving animals are challenging and rare, we used all valid sessions. We addressed the same point in our response to the Reviewer 1 weaknesses.
Recommendations for the authors:
Reviewer #1 (Recommendations for the authors):
Below are some suggestions for clarification/information that are needed to improve the paper's readability (and the understanding of the analysis and methods).
(1) The authors describe a consistently negative correlation between cortical EEG synchronization index and ripple rate or LC-MUA, show an example in Figure 3A, and report a range of r values in the text with a mention of p < 0.01. The reported p-value is presumably the highest p-value for the correlations - please specify. Visualization of the results might be improved by adding example correlations (also true for later correlations in Figure 6).
We revised the result description accordingly and included correlation plots in Figures 3 and 7.
(2) Description of statistical testing is missing for Figure 3C (nothing in the text or the figure legend); there is also no statistics section in the methods. For Figure 4, the statistics are reported for the Friedman test but not the post-hoc tests. Exact p-value and statistics should be reported for the comparison of LC-MUA rate and SI in the 2 s preceding spindles.
We have added the statistical results requested and revised figure legends by providing additional information. We added the Statistic Analysis section in the Methods.
Figure 3D (original Fig.3C): “Average Synchronization Index (SI) around ripples and shuffled events. The cortical state preceding shuffled events and ripples was comparable, as confirmed by the absence of significant differences in SI (Wilcoxon signed-rank test; shuffled: Z = -0.20, p = 0.84; ripples: Z = 0.14, p = 0.88). Cortical synchrony increased following both events (shuffled: Z = -3.50, p = 0.00044; ripples: Z = -3.66, p = 0.00026). Similar cortical state dynamics surrounding shuffled events and ripples indicate that the surrogate events adequately capture the cortical state associated with ripple occurrence.
Figure 6: Intra-ripple frequency (A) and peak amplitude (B) for different ripple types. Boxwhisker plots show the median, the 1st and 3rd quartiles, and min/max. Gray dots show data from individual rats. *** - p < 0.001 for post hoc pairwise comparisons (Wilcoxon signed-rank tests with Holm–Bonferroni correction for multiple comparisons).
We revised the Results accordingly: “The ripple subtypes differed in the intra-ripple frequency (Friedman test, chi2 = 35.62, p < 0.0001, post hoc pairwise comparisons were performed using Wilcoxon signed-rank tests with Holm–Bonferroni correction for multiple comparisons. awRipple vs isoRipple: p = 0.00003 awRipple vs spRipple: p = 0.00004 isoRipple vs spRipple: p = 0.0002}), with awRipples being the fastest and spRipples the slowest (Figure 6A).There was no difference in the ripple peak amplitude (Friedman test, $\chi$2 = 3.7, p = 0.16; Figure 6B).”
(3) The method description of ripple-spindle coupling detection is missing.
We have added the description of ripple-spindle coupling detection in the Methods.
(4) Based on Figure 6D, the authors report that ripple-coupled spindles are significantly shorter than isolated spindles. What are the measurements reported on lines 206-207, and how do they relate to the averaged spectrograms shown in Figure 6D?
Spindle duration was calculated as the time between spindle onset and offset (as described now in the Methods and Figure 7 legend). Ripple-coupled spindle was considered if at least one ripple occurred between the spindle onset and offset. The duration of ripple-coupled and uncoupled spindles was statistically compared (the stats is reported in text). In Figure 7E, the peri-event averaged EEG spectrograms are plotted for isolated and ripple-coupled spindles, highlighting the difference in the event duration.
(5) None of the color scales have legends (Figures 2A, B, C, Figure 3D, etc.).
We have added the color scales on all Figures.
(6) Description of what is represented in the box plots is missing.
We have added the description.
(7) Figure 4C, D, legend for the color code is missing.
We have added color scales legends.
(8) Figure 5A legend, assuming this should read intra-ripple frequency instead of inter-ripple.
We corrected the typo.
(9) Figure 5E, while LC units are not modulated before, it could still be informative to overlay the z-scored firing rate on the same graph for comparison.
Figure 6E (original Figure 5E) shows overlay for awRipples and isoRipples.
(10) The discussion states a 4s resolution for cortical state quantification (line 237), but the methods mention 2.5s (line 382).
We corrected this discrepancy.
(11) Results, p.5, line 138, Methods and materials, p.13, line 423: 30% in result text but 20% in method, please correct.
We corrected this discrepancy.
(12) The manuscript cites the biorxiv version of Osorio-Forero et al., but the paper has been published since then; please update.
We updated this reference.
(13) Results, p.2, line 70. The average duration of a session is presented in seconds. Minutes or hours would be more meaningful to the reader.
We consider this suggestion as optional.
(14) Figure 2C is not referenced.
We added the reference to Figure 2C.
(15) Reference missing line 406.
We added the reference.
(16) Lines 352-356: There seems to be an error in the sentence (an extra verb, or an "and" missing somewhere).
We have corrected this sentence.
(17) Figure 3C "synchronization".
We corrected this typo.
Reviewer #2 (Recommendations for the authors):
(1) Line 94 states that "A significant peri-ripple decrease in LC-SUA"; however, which test and how many samples were used are unclear.
We revised this text as follows: “A significant peri-ripple (± 6 s) decrease in LCSUA, detected by the firing suppression exceeding 2 SDs, was observed in 13 of 15 cases (n = 4 rats).”
(2) Line 96 states that "we calculated the modulation onset, duration, and magnitude". Please define modulation before presenting the comparisons.
We now illustrate the extraction of quantitative variables in Figure 2D.
(3) Line 119 states that "we generated surrogate time series for each session by shuffling ripple onset times" which gives the impression that ripple events were shuffled throughout the sleep; however, the method section states that it was jittered within a specific time window for each event. Please clarify the matter.
We have substantially revised this section to improve clarity and readability.
(4) Line 120 states that "Comparisons of SI values before and after ripples and surrogate events confirmed that surrogate events preserved the cortical states in which ripples occurred". Ripple power doesn't seem to be different in pre vs post in the shuffled data (Figure 3B). If ripple timing was randomized, please clarify the observation shown in Figure 3C that the shuffled events had higher SI after than before, as also seen in the real SI data? Please also elaborate what specific groups were significantly different in before vs after bars; data, shuffle, or both?
We have substantially revised this section to improve clarity and readability.
(5) Line 113 and Figure 3A: Because both LC activity and HPC ripples were correlated to SI, the direct relationship between LC and HPC independent of SI (a covariate) was not clear. The authors might be able to conduct a partial correlation analysis to show this effect.
We appreciate this suggestion and added the correlation plots in Figures 3 and 7. After careful consideration, we believe that the suggested partial correlation analysis does not contribute substantially beyond the main findings already presented.
(6) Figure 5A: Inter-ripple frequency needs definition, not provided in the paper nor in the reference paper. The value (180 Hz) suggests a time interval of around 5 ms, which I fail to understand.
We apologize for this typo. In Figure 6A (original Fig.5A), intra-ripple frequency is plotted. We have corrected this typo in the text and figure legend.
(7) Figure 5D: Comparison between aw and sp ripples should also be shown. Please explain the dashed line at 10 (y-axis) a.u.
Figure 6E (original Fig.5E) shows LC activity around awRipples and isoRipples.
(8) Figure 5E: Legend states aw and iso ripples, but the caption says NREM sleep. Please clarify this matter.
We have revised Figure 6 legend (original Figure 5).
(9) Figure 6B: If the spindle time is permuted randomly, why is LC activity in the permuted data still modulated by the spindle times? Can you test the significance of the modulation index of the shuffled data?
The LC modulation around shuffled time points was not significant. Figure 7C shows LC modulation dynamics around spindles; brown trace showing state-corrected LCMUA trace (after subtraction of LC-MUA around shuffled events).
(10) Line 203: Is the unit in Hz (events per second) correctly calculated or shown? ~15 events per second seems arbitrarily large.
We corrected the units for the event rate. We report the mean oscillatory frequency of spindles ~15 Hz, not events per second.
(11) Line 207 states that "neither the LC-MUA rate nor SIs differed during a 2-sec time window preceding either group of spindles"; however, from Figure 6E, the average trace and errors around them (errors need to be stated clearly, for e.g., SEM or SD) show that they are non-overlapping and different. I suspect tests such as the rank-sum test, which test the difference in the central tendencies (as opposed to the KS test, which tests the overall trend in the distribution of the continuous data), might reveal the difference between these values.
We compared the absolute (not normalized) LC-MUA rate and SI during 2 sec time window preceding spindle onset and did not find any statistical differences. In Figure 7F, the difference during ~ 2 sec before the spindle onset is due to the z-score normalization to their own baseline.
We revised the Result text to improve clarity.
(12) Line 209: Modulation seems to be greater in ripp-spindles as shown in fig 6E-F, yet, the text and the interpretation are the opposite i.e,. iso spindles had greater modulation. Hence, authors might have to provide further clarifications or analyses.
We corrected the labelling in all plots.
(13) Line 316: Claims of "suppression of noradrenergic system facilitating the generation of hippocampal ripples and sleep spindles by memory synchrony" are not fully supported by data, as the data seem to be correlational. Also, claims of "preserved LC activity during ripples coinciding with sleep spindles suggest a role for NE in facilitating cross-regional communication underlying memory-related information transfer" lack clarity and contradict the earlier mechanism. Both "suppression" as well as "preservation" of LC neurons are proposed to mechanistically support memory synchrony and/or consolidation in two different brain states (awake and sleep). The authors might need to clarify how both suppression as well as preservation (which I assume is not an activation or positive modulation) of LC neurons can help in memory synchrony or consolidation.
We revised this part of discussion by making it less speculative.
Reviewer #3 (Recommendations for the authors):
I would recommend that the authors optimize their figure and result presentation, as the current version of the manuscript is unclear in several places, limiting the interpretation of results.
We substantially revised the manuscript to improve the results presentation and readability.
(1) Multiple results are described but not shown quantitatively. Please plot quantifications and statistics (mean {plus minus} error and individual values) in relevant figures. For example, the results referenced on p. 4 (l. 113-116), p. 5 (l. 129-133, 143-147), p. 6 (l. 159161), p. 7 (l. 188-190), and p. 8 (l. 203-207) should be supported by explicit data plots.
We have revised the manuscript to ensure all results are supported by quantitative and statistical analyses. We revised figures and legends and added new plots showing individual datapoints.
(2) Improvements in figures and descriptions are needed. Below are some examples I found:
(a) All figures with color scales lack labeling of the color axis, i.e., measure and unit.
We have revised the figures accordingly.
(b) Use precise labeling of axes such as "ripple-band power" and "LC-MUA firing rate", rather than just "power" and "firing rate".
We have revised the figures accordingly.
(c) Figure 1: Indicate behavioral state (wake vs. sleep) in the example trace.
We have indicated the behavioral state (quiet awake) in the figure legend.
(d) Define "peri-ripple" windows explicitly (e.g., {plus minus}6 s or {plus minus}30 s).
We have revised the text and figure legends accordingly.
(e) Clarify how "modulation magnitude" is calculated (line 96).
We now illustrate the extraction of quantitative variables in Figure 2D
(f) Figure 2C: The white overlaid mean trace lacks Y-axis labeling.
We have added y-axis labeling.
(g) Figure 3A: The labeling of "amplitude" is confusing when referring to firing frequency.
We have corrected the figure labelling.
(h) Figure 4B: Is the X-axis time from ripple onset?
We have corrected the figure labelling.
(i) Figure 4C-D lacks an X-axis or color legend.
We have added x-axis and color legend.
(j) Figures 5-6: Include tonic firing rates and time scales.
We have added in the main text the time scales and average firing rates for LC single units and also show it in Supplementary Figure 1. Because the number of neurons contributing to LC multi-unit activity (LC-MUA) is unknown, we avoided averaging absolute firing rates for this signal. For LC-MUA, we implemented a normalization approach in which firing rates (50-ms bins) around ripple were scaled to a baseline period preceding the trigger event (−12 to −10 s). Importantly, unlike z-scoring, this normalization method preserved baseline differences across behavioral states, as shown in new Figure 5.
(k) Add tonic firing rate baselines where relevant.
We have added the Supplementary Figure 1 and new Figure 5 showing the difference in the LC baseline firing rate across behavioral states.
(3) Minor Comments to add more clarity
(a) Clarify "spike train" selection criteria (Methods, p. 4, line 93).
We revised the text as follows: “In six out of twenty LC-MUA recordings, we could reliably isolate spikes from a total of 15 single units (LC-SUA, n = 4 rats).”
(b) Define "EEG transients" (p. 4, line 109) and support with data.
We revised the text as follows: “Indeed, transient spectral changes in the prefrontal EEG coincided with the occurrence of hippocampal ripples (Figure 2B).”
(c) You refer to Figure 3E as a histogram (p. 5, line 128), but I believe it shows an average trace.
We have corrected this typo.
(d) Standard sleep scoring procedures normally involve EMG measurements (p. 6, line 154).
We have replaced ‘standard’ with “previously established”.
(e) Explain how surrogate shuffling preserves the distribution of behavioral states.
We revised the text as follows: “We first verified that hippocampal LFPs (140– 250 Hz) triggered on these surrogate events lacked the ripple-specific frequency component (Figure 3C), and that the SI state did not differ between real ripples and surrogate events (Figure 3D).”
(f) You refer to inter-ripple frequency (p. 6, line 168), which suggests time between ripples. Do you mean the "intra-ripple" or simply ripple frequency?
We have corrected this typo.
(g) Ensure all references cited in the text (e.g., p. 12, line 406) are included in the bibliography.
We have updated the bibliography.
(h) On p. 10, line 304-305 authors refer to observations related to offline memory consolidation. However, the present study does not contain any behavioral memory data.
We have revised the Discussion to make it less speculative about the role of describe LC dynamics for offline memory consolidation.
References
Novitskaya Y, Sara SJ, Logothetis NK, Eschenko O (2016) Ripple-triggered stimulation of the locus coeruleus during post-learning sleep disrupts ripple/spindle coupling and impairs memory consolidation. Learn Mem 23:238-248.
Yang M, Logothetis NK, Eschenko O (2019) Occurrence of Hippocampal Ripples is Associated with Activity Suppression in the Mediodorsal Thalamic Nucleus. J Neurosci 39:434-444.
Yang M, Logothetis NK, Eschenko O (2021) Phasic activation of the locus coeruleus attenuates the acoustic startle response by increasing cortical arousal. Sci Rep 11:1409.



