Peer review process
Revised: This Reviewed Preprint has been revised by the authors in response to the previous round of peer review; the eLife assessment and the public reviews have been updated where necessary by the editors and peer reviewers.
Read more about eLife’s peer review process.Editors
- Reviewing EditorHugo BellenBaylor College of Medicine, Houston, United States of America
- Senior EditorDavid RonUniversity of Cambridge, Cambridge, United Kingdom
Reviewer #1 (Public review):
Summary:
In this study, the authors investigate whether glycogen phosphorylase represents a molecular target of benzoylphenylurea insecticides and evaluate the physiological consequences of suppressing glycogen phosphorylase activity in the diamondback moth Plutella xylostella. The authors combine recombinant protein biochemistry, enzyme inhibition assays, RNA interference, structural modelling, metabolite profiling, gene expression analyses, and physiological measurements to determine whether diflubenzuron directly inhibits glycogen phosphorylase and whether suppression of this enzyme is sufficient to impair insect development. Based on these experiments, the authors conclude that diflubenzuron does not directly inhibit glycogen phosphorylase and that insects tolerate substantial suppression of this enzyme through compensatory metabolic responses.
Strengths:
This study addresses an important question in insect toxicology by systematically evaluating glycogen phosphorylase as a potential insecticidal target. The authors combine complementary biochemical, molecular, physiological, and structural approaches, including recombinant enzyme characterization, inhibitor assays, RNA interference, metabolite profiling, structural modelling, and measurements of fitness-related traits. This integrative approach provides a comprehensive evaluation of the biological consequences of glycogen phosphorylase suppression. In particular, the biochemical evidence that diflubenzuron does not inhibit glycogen phosphorylase, together with the observation that strong suppression of glycogen phosphorylase produces only transient physiological effects without measurable impacts on development or reproduction, provides strong support for the conclusion that glycogen phosphorylase is unlikely to represent an effective standalone insecticidal target.
Weaknesses:
The main limitation concerns the proposed mechanism underlying metabolic compensation. The observed increases in gluconeogenic gene expression, changes in metabolite abundance, and reductions in total protein are consistent with activation of compensatory metabolism, but are insufficient to directly demonstrate increased gluconeogenic flux or establish that amino acid-derived carbon is incorporated into newly synthesized glucose. Similarly, although the analyses of glycogen-associated enzymes strengthen the discussion of alternative metabolic pathways, changes in gene expression alone do not demonstrate that these pathways contribute to glycogen utilization in vivo.
Some mechanistic interpretations therefore extend beyond the data presented. For example, decreases in total protein are interpreted as evidence of protein catabolism fuelling gluconeogenesis, yet they do not directly demonstrate amino acid mobilization or incorporation into glucose. Likewise, increased expression of gluconeogenic genes is interpreted as evidence of increased pathway activity, although transcriptional changes do not necessarily reflect metabolic flux. Finally, the absence of major developmental defects following glycogen phosphorylase suppression is attributed primarily to metabolic compensation, but an alternative explanation is not fully considered. Such explanation could be that glycogen phosphorylase is not rate-limiting for glucose homeostasis under the nutrient-rich experimental conditions, where dietary carbohydrates are continuously available. Consequently, the proposed compensatory mechanism remains plausible and well supported by indirect evidence, but several aspects would benefit from more cautious interpretation.
Overall, the authors successfully achieve their primary objective of evaluating glycogen phosphorylase as a candidate insecticidal target. The study provides useful biochemical and physiological evidence that this enzyme is unlikely to represent an effective target for insecticide development in P. xylostella, while highlighting the importance of metabolic plasticity when assessing metabolic targets. The experimental approaches and datasets presented here should be valuable to researchers studying insect metabolism, insecticide mode of action, and target validation, although the precise mechanisms underlying the proposed metabolic compensation remain an important subject for future investigation.
Author response:
The following is the authors’ response to the original reviews.
eLife Assessment
This study addresses the mechanism of action of benzoylurea insecticides and explores the metabolic consequences of inhibiting glycogen breakdown in insects. Both reviewers identify major flaws with the premise of the work. The strength of the provided evidence is inadequate as the data do not, or poorly, support several central claims. The significance of the findings is considered marginal.
The Assessment stated that “both reviewers identify major flaws with the premise of the work” and that “the strength of the provided evidence is inadequate.” We have addressed both dimensions:
(1) Premise: The Introduction has been substantially restructured to explicitly acknowledge the compelling CRISPR/Cas9 evidence establishing CHS as the primary site of BPU resistance (Reference 1). The study is now reframed as a systematic evaluation of GP as an independent insecticidal target and an investigation of metabolic compensation mechanisms — questions with scientific value independent of the BPU mechanism debate (see details in lines 47-54 of the revised manuscript).
(2) Evidence: Four new sets of experiments directly address the specific evidence gaps identified by the reviewers: (i) GP enzyme activity measurements in RNAi-treated larvae; (ii) expression analysis of alternative glycogen catabolic enzymes; (iii) molecular docking and MM/GBSA binding free energy analysis; (iv) comprehensive fitness cost assessment including feeding rate, larval weight, pupal weight, adult wing area, and female fecundity.
Public Reviews:
Reviewer #1 (Public review):
Summary:
In this study, the authors investigate whether glycogen phosphorylase is a potential molecular target of benzoylphenylurea insecticides and examine the physiological consequences of inhibiting glycogen breakdown in the diamondback moth Plutella xylostella. The authors express and characterize recombinant glycogen phosphorylase, test its inhibition by a mammalian glycogen phosphorylase inhibitor and by the insecticide diflubenzuron, and assess the physiological effects of glycogen phosphorylase inhibition through chemical exposure and RNA interference. Based on these experiments, the authors conclude that benzoylphenylurea insecticides do not target glycogen phosphorylase and propose that insects compensate for glycogen phosphorylase inhibition through activation of gluconeogenesis, allowing them to maintain glucose homeostasis and complete development despite strong suppression of the enzyme.
Strengths:
The study addresses an interesting and long-standing question in insect toxicology regarding the mechanism of action of benzoylphenylurea insecticides. The authors combine several complementary approaches, including recombinant enzyme characterization, inhibitor assays, RNA interference, gene expression analyses, and metabolite measurements. The biochemical characterization of the recombinant glycogen phosphorylase and the demonstration that the tested glycogen phosphorylase inhibitor can strongly inhibit enzyme activity represent important technical strengths. In addition, the study integrates biochemical and physiological observations to explore how insects might compensate for disruptions in central carbohydrate metabolism.
We are grateful that Reviewer 1 recognized the study's strengths, including the complementary multi-approach strategy, the biochemical characterization of recombinant PxGP, and the integration of biochemical and physiological observations.
Weaknesses:
(1) The proposed compensatory mechanism relies on indirect evidence; direct measurements of gluconeogenic flux are lacking.
We agree that isotopic tracer experiments would provide the most direct evidence for gluconeogenic flux. Such experiments are beyond the scope of the current revision, and we now explicitly acknowledge this as a key limitation and an important direction for future research (revised Discussion: Study limitations and future directions).
However, we note that the convergent evidence from multiple independent lines collectively supports gluconeogenic activation: (i) transcriptional upregulation of PEPCK and G-6-Pase; (ii) declining protein levels now independently confirmed by our new enzyme activity data showing a 30.78% decrease in total protein concentration at 24 h post-RNAi (new Figure 10A); (iii) altered amino acid profiles; and (iv) maintained trehalose levels. The revised manuscript presents this evidence more cautiously, framing it as “consistent with gluconeogenic compensation” rather than establishing metabolic flux.
Additionally, we now provide new data on GP enzyme activity (new Figure 10A, B; see response to Recommendation 4 below) and alternative glycogen catabolic enzyme expression (new Figure 10C, D; see response to Recommendation 2 below) that further strengthen the evidence chain.
(2) Alternative glycogen degradation pathways are proposed but not experimentally examined.
We have now directly addressed this concern. RT-qPCR analysis of glycogen branching enzyme (GBE) and α-amylase following PxGP knockdown reveals a striking and informative differential response (new Figure 10C, D):
GBE was significantly upregulated at 24 h (+29.24%, P < 0.05), 48 h (+16.78%, P < 0.05), and 96 h (+44.46%, P < 0.001), indicating transcriptional activation of an alternative glycogen-remodeling enzyme in response to GP suppression.
α-Amylase showed no significant change at any time point, demonstrating that the compensatory response is pathway-specific rather than a generalized upregulation of all glycogen-degrading enzymes.
This differential pattern — GBE up, α-Amylase unchanged — provides the first evidence that P. xylostella selectively activates specific glycogen remodeling pathways when GP function is compromised. Upregulation of GBE, which increases glycogen branching and solubility, may facilitate glycogen mobilization through alternative routes even when GP-mediated phosphorolysis is impaired. These data are incorporated as new Figure 10C, D and discussed in the revised Results and Discussion.
(3) Physiological consequences (fitness costs) are not explored.
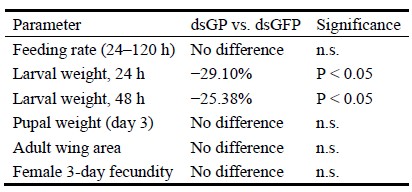
We have now conducted a comprehensive fitness cost assessment (new Figure 11). The results reveal a transient but significant fitness cost confined to the larval stage:
Feeding rate: no significant difference between dsGP and dsGFP groups at any time point (24–120 h; Figure 11B), confirming that the observed metabolic changes are not attributable to reduced food intake.
Larval weight: significantly reduced at 24 h (−29.10%, P < 0.05) and 48 h (−25.38%, P < 0.05; Figure 11C), demonstrating a measurable short-term cost of metabolic compensation.
Pupal weight: no significant difference (Figure 11D), indicating full recovery before the pupal transition.
Adult wing area: no significant difference (Figure 11E, Figures S5–S6), suggesting no impairment of flight capacity.
Female fecundity (3-day egg production): no significant difference (Figure 11F), demonstrating no reduction in reproductive output.
This pattern — transient larval weight loss with complete recovery of pupal weight, wing morphology, and reproductive performance — is consistent with our proposed model: GP suppression triggers protein catabolism to fuel gluconeogenesis (explaining the short-term weight loss), but the compensatory mechanism is sufficiently effective to restore metabolic homeostasis before pupation. These data strengthen the conclusion that GP is functionally non-essential for completing development and reproduction.
(4) Broader conclusions regarding BPU class may require testing additional compounds.
We agree. The revised manuscript now explicitly limits the biochemical conclusion to diflubenzuron: “DFB does not inhibit PxGP” rather than making broader claims about the BPU class as a whole. We discuss this as a limitation and note that testing additional BPU compounds would be needed before generalizing.
(5) Some biochemical and cell-based observations would benefit from confirmation in whole insects.
We have now provided whole-insect confirmation through: (i) GP enzyme activity measurements in RNAi-treated larvae (new Figure 10A, B); (ii) in vivo fitness assessment showing measurable physiological consequences of GP suppression (new Figure 11); and (iii) expression analysis of compensatory enzymes in intact larvae (new Figure 10C, D). These data bridge the gap between our cell-free biochemical observations and whole-organism biology.
Reviewer #2 (Public review):
(1) The central premise — that structural similarity among acylurea compounds implies shared targets — is not supported.
We agree that the original manuscript overstated the significance of the shared acylurea core as a predictor of common biological activity. The Introduction has been substantially restructured to:
Explicitly acknowledge the compelling genetic evidence from CRISPR/Cas9 experiments (Reference 5) establishing CHS as the primary site conferring BPU resistance.
Reframe the study's objective: rather than proposing to “resolve” the BPU target controversy, the revised manuscript focuses on the systematic evaluation of GP as an independent insecticidal target and the discovery of a gluconeogenic compensation mechanism — questions with scientific value independent of the BPU mechanism debate.
Remove the claim that the study “resolves the primary hypothesis.” The conclusion now states that our biochemical data demonstrate DFB does not inhibit PxGP, adding enzyme-level evidence to the existing genetic framework.
(2) Target selectivity is determined by side-chain composition, not the shared acylurea core.
We fully agree, and our new structural data now provide a molecular explanation for this principle at the atomic level. Molecular docking and MM/GBSA analysis (new Figure 12, new Table 1) reveal that both GPI and DFB anchor to PxGP through their common acylurea carbonyl groups (Arg193), but diverge dramatically in side-chain engagement:
GPI's methoxyphenyl-methylurea moiety establishes extensive contacts with seven residues across both subunits (Asn44 and Val45 from chain A; Trp67, Gln71, Tyr75, Arg193, and Asp227 from chain B), binding at the allosteric site at the dimer interface — consistent with the experimentally determined binding mode of acylurea inhibitors in mammalian GP (PDB: 2ATI).
DFB contacts six residues primarily from subunit B, and its difluorobenzoyl moiety remains entirely solvent-exposed without productive protein contacts.
MM/GBSA analysis confirms GPI binds with substantially higher affinity (ΔG = −34.63 vs. −29.29 kcal/mol; ΔΔG = −5.34 kcal/mol), with van der Waals interactions as the dominant driver (ΔVDW = −11.49 kcal/mol), reflecting superior shape complementarity of GPI.
These structural data directly support Reviewer 2's important point and are now presented as new Figure 12 and Table 1.
(3) References 6–9 characterization.
We have replaced the original citations (former References 6–9) in the Introduction with references that directly demonstrate the absence of CHS inhibition by BPUs in cell-free preparations: Cohen & Casida (1980) showed DFB did not inhibit Tribolium gut chitin synthetase; Mayer et al. (1981) and Cohen (1985) systematically confirmed that BPU-type insect growth regulators do not inhibit chitin synthase in cell-free assays; and Zhang & Zhu (2013) reported only slight in vitro CHS inhibition by DFB in Anopheles gambiae with no in vivo effect. We also cite authoritative reviews by Matsumura (2010) and Merzendorfer (2013) that contextualize this evidence gap. We thank Reviewer 2 for identifying this important citation issue, which has led to a substantially more accurate and well-supported presentation of the literature.
(4) The term “dataology” is non-standard.
This term has been removed and replaced with “data.” In accordance with eLife's policy on AI tools and technology, we have added a statement in the Materials and Methods section declaring that AI-based language editing tools were used for English grammar and style refinement. All scientific content was generated entirely by the authors.
Recommendations for the authors:
Reviewer #1 (Recommendations for the authors):
(1) Direct assessment of gluconeogenic flux (e.g., metabolic tracer experiments).
As discussed above, isotopic tracer experiments are beyond the current scope. We acknowledge this as a key limitation in the revised Discussion (see details in lines 560-565 of the revised manuscript). However, we now provide additional supporting evidence: the 30.78% decline in total protein at 24 h post-RNAi (from our new enzyme activity data, Figure S3) provides independent biochemical confirmation of protein catabolism, consistent with amino acid mobilization for gluconeogenesis (see details in lines 357-359 of the revised manuscript). We have also added discussion of potential future approaches, including measurements of key enzymes in amino acid catabolism (e.g., aspartate aminotransferase, glutamate aminotransferase) and lipid content dynamics (see details in lines 565-569 of the revised manuscript).
(2) Expression or activity of alternative glycogen degradation enzymes (α-amylase, glycogen debranching enzymes).
We have measured the expression of GBE (glycogen branching enzyme) and α-amylase by RT-qPCR in RNAi-treated insects. We also attempted to measure glycogen debranching enzyme (GDE), but multiple primer pairs failed to yield amplification products, likely due to sequence annotation issues; this is noted as a limitation.
Results (new Figure 10C, D): GBE was significantly upregulated at 24 h (+29.24%), 48 h (+16.78%), and 96 h (+44.46%). α-Amylase was unchanged at all time points (see details in lines 377-381 of the revised manuscript). The selective upregulation of GBE but not α-amylase suggests a targeted compensatory response within the glycogen remodeling pathway.
The absence of glycogen accumulation following GP knockdown may reflect reduced flux into glycogen synthesis (potentially through feedback inhibition of glycogen synthase) rather than activation of alternative degradative routes. This possibility is discussed in the revised manuscript, with glycogen synthase expression identified as a key target for future investigation (see details in lines 501-505 of the revised manuscript).
(3) Fitness cost assessment (body size, flight capacity, reproductive performance).
Complete data are now provided (new Figure 11A–F, Figures S5–S6):
Author response table 1.
The transient larval weight reduction (24–48 h) with complete pupal and adult recovery demonstrates that metabolic compensation carries a short-term physiological cost but is ultimately effective in maintaining developmental trajectory and reproductive fitness (see details in lines 388-408 of the revised manuscript).
(4) Enzyme activity measurements in RNAi-treated insects.
GP enzyme activity (GP-a) was measured in crude extracts from RNAi-treated larvae using a coupled-enzyme spectrophotometric assay kit (Solarbio BC3345) at 24, 48, 72, and 96 h post-injection (new Figure 10A, B).
Two normalization approaches were used: - Per-protein activity: significant reduction only at 48 h (−10.35%, P < 0.05; Figure 10A). The modest per-protein reduction reflects a concurrent 30.78% decline in total protein at 24 h, which inflates per-protein specific activity when the protein pool shrinks. - Per-larva activity: significant reduction at 24 h (−27.57%, P < 0.05) and 48 h (−29.28%, P < 0.01; Figure 10B), confirming that RNAi-mediated transcript suppression translates to reduced enzyme function in vivo.
The 30.78% decline in total protein at 24 h provides independent biochemical confirmation of protein catabolism — consistent with amino acid mobilization for gluconeogenesis (see details in lines 349-370 of the revised manuscript).
(5) Scope of BPU conclusion — clarify whether additional compounds should be tested.
The revised manuscript explicitly states that the biochemical conclusion applies to diflubenzuron specifically. We have added a Discussion paragraph noting that extending this conclusion to additional BPU compounds would require systematic testing, and that our structural analysis (Table 1) provides a framework for predicting which acyl urea side-chain architectures are compatible with GP binding (see details in lines 455-466 of the revised manuscript).
(6) RNAi transcript recovery at 96 h and implications for non-essentiality.
Our new enzyme activity data directly address this concern. GP activity (per-larva) showed partial recovery at 72 h and 96 h (Figure 10B), mirroring the transcript recovery pattern. However, the critical observation is that even during the period of maximum suppression (24–48 h), when per-larva GP activity was reduced by ~27–30%, larvae maintained glucose homeostasis and completed development. This confirms that GP is non-essential even during the period of strongest suppression. The revised Discussion addresses this point explicitly (see details in lines 479-488 of the revised manuscript).
(7) GPI concentrations in larval exposure experiments and pharmacokinetic considerations.
We have added a dedicated Discussion paragraph addressing this concern in detail. The GPI concentrations used (250–500 mg/L in diet) encompass a wide dose range; even at the highest concentration (500 mg/L, approximately 409,000-fold above the in vitro IC50 of 2.96 nM), no toxicity was observed. We discuss several pharmacokinetic factors that may contribute to this apparent discrepancy, including limited oral bioavailability, metabolic inactivation by detoxification enzymes, and sequestration by hemolymph binding proteins. However, we note that the metabolic phenotype observed (elevated trehalose, reduced protein, upregulated gluconeogenic enzymes) provides indirect evidence that GPI does reach its target. The most parsimonious interpretation is that GPI achieves sufficient target engagement to partially suppress GP activity in vivo, but metabolic compensation renders this suppression non-lethal — an interpretation reinforced by our RNAi data, in which direct genetic suppression of GP (bypassing all pharmacokinetic barriers) similarly fails to cause mortality (see details in lines 536-552 of the revised manuscript).
(8) Structural evidence that GPI binds PxGP comparably to its mammalian target.
This has been comprehensively addressed through molecular docking and MM/GBSA analysis (new Figure 12, new Table 1). The PxGP homodimer structure was modeled using SWISS-MODEL with the human liver GP–acyl urea co-crystal structure (PDB: 2ATI) as the template. Docking and binding free energy calculations were performed in Cresset Flare V11.
Key findings: GPI binds at the allosteric site at the dimer interface with ΔG = −34.63 kcal/mol, engaging seven residues across both subunits — a binding mode consistent with the experimentally determined site in mammalian GP. DFB binds with lower affinity (ΔG = −29.29 kcal/mol) and its difluorobenzoyl moiety is entirely solvent-exposed. Van der Waals interactions are the dominant driver of selectivity (ΔVDW = −11.49 kcal/mol). See new Figure 12 and Table 1 for complete data.
(9) Dietary carbohydrate compensation and feeding behavior.
Our new data directly address this concern. Feeding rate measurements show no significant difference between dsGP and dsGFP groups at any time point (24–120 h; Figure 11B), confirming that the metabolic changes are not attributable to altered food intake. A Discussion paragraph has been added acknowledging the potential contribution of dietary carbohydrates to glucose homeostasis and noting that starvation-challenge experiments would provide additional insight (see details in lines 570-581 of the revised manuscript).
Minor comments and suggestions
(1) Terminology.
“Gluconeogenolysis” has been replaced with “gluconeogenesis” throughout the manuscript.
(2) Typographical errors.
A thorough language revision has been performed. “Over over four decades” and other errors have been corrected. The term “dataology” has been removed.
(3) Metabolite normalization.
We now present GP enzyme activity using two normalization approaches (per-protein and per-larva; Figure 10A, B) and discuss the implications of protein level changes on per-protein normalization in the Results section. For metabolite data, we have added a note in the Methods explaining our normalization approach and discussing how declining protein levels may influence interpretation (see details in lines 908-920 of the revised manuscript).
(4) Clarity of pathway descriptions.
The description of metabolic pathways has been simplified and a revised schematic figure has been included (Figure 13).
(5) Figure clarity.
Figures have been added with clearer labeling and simplified schematics. To improve clarity, we used red arrows to show the blocked metabolic flow when GP is inhibited, and green arrows to depict the activated gluconeogenic pathway (Figure 13).
Cohen E, Casida JE. Inhibition of Tribolium gut chitin synthetase. Pestic Biochem Physiol. 1980;13(2):129-36. doi: 10.1016/0048-3575(80)90064-4.
Mayer RT, Chen AC, DeLoach JR. Chitin synthesis inhibiting insect growth regulators do not inhibit chitin synthase. Experientia. 1981;37(4):337-8. doi: 10.1007/BF01959848.
Cohen E. Chitin synthetase activity and inhibition in different insect microsomal preparations. Experientia. 1985;41(4):470-2. doi: 10.1007/BF01966152.
Zhang X, Yan Zhu K. Biochemical characterization of chitin synthase activity and inhibition in the African malaria mosquito, Anopheles gambiae. Insect Sci. 2013;20(2):158-66. doi: 10.1111/j.1744-7917.2012.01568.x.
Matsumura F. Studies on the action mechanism of benzoylurea insecticides to inhibit the process of chitin synthesis in insects: A review on the status of research activities in the past, the present and the future prospects. Pestic Biochem Physiol. 2010;97(2):133-9. doi: 10.1016/j.pestbp.2009.10.001.
Merzendorfer H. Chitin synthesis inhibitors: old molecules and new developments. Insect Sci. 2013;20(2):121-38. doi: 10.1111/j.1744-7917.2012.01535.x
Preiss J. Bacterial glycogen synthesis and its regulation. Annual review of microbiology. 1984;38:419-58. doi: 10.1146/annurev.mi.38.100184.002223.
Janeček Š, Svensson B, MacGregor EA. α-Amylase: an enzyme specificity found in various families of glycoside hydrolases. Cell Mol Life Sci. 2014;71(7):1149-70. doi: 10.1007/s00018-013-1388-z.
Waterhouse A, Bertoni M, Bienert S, Studer G, Tauriello G, Gumienny R, et al. SWISS-MODEL: homology modelling of protein structures and complexes. Nucleic Acids Res. 2018;46(W1):W296-W303. doi: 10.1093/nar/gky427.
İnak E, De Rouck S, Van Leeuwen T. Molecular mechanisms of pesticide selectivity: Insights from acaricide toxicology. Pestic Biochem Physiol. 2025;213:106537. doi: 10.1016/j.pestbp.2025.106537.
David MD. Insecticide ADME for support of early-phase discovery: combining classical and modern techniques. Pest Manage Sci. 2017;73(4):692-9. doi: 10.1002/ps.4345.
Haunerland NH, Bowers WS. Binding of insecticides to lipophorin and arylphorin, two hemolymph proteins of Heliothis zea. Arch Insect Biochem Physiol. 1986;3(1):87-96. doi: 10.1002/arch.940030110.
Other revisions
Correction of primer sequences. Upon re‑checking the primer sequences during revision, we noticed that the originally reported primers for dsRNA synthesis of the GP gene (Table S1) were inadvertently copied incorrectly. The correct sequences have now been substituted in the revised manuscript (dsPxGP-F, dsPxGP-R). This correction does not affect any of the experimental data, results, or conclusions of the study. We apologize for the oversight.



