Abstract
Eukaryotes are often exposed to microbes and respond to their secreted metabolites, such as the microbiome in animals or commensal bacteria in roots. Little is known about the effects of long-term exposure to volatile chemicals emitted by microbes, or other volatiles that we are exposed to over a long duration. Using the model system Drosophila melanogaster, we evaluate a yeast emitted volatile, diacetyl, found in high levels around fermenting fruits where they spend long periods of time. We find that exposure to just the headspace containing the volatile molecules can alter gene expression in the antenna. Experiments showed that diacetyl and structurally related volatile compounds inhibited human histone-deacetylases (HDACs), increased histone-H3K9 acetylation in human cells, and caused wide changes in gene expression in both Drosophila and mice. Diacetyl crosses the blood-brain barrier and exposure causes modulation of gene expression in the brain, therefore has potential as a therapeutic. Using two separate disease models known to be responsive to HDAC-inhibitors, we evaluated physiological effects of volatile exposure. First, we find that the HDAC inhibitor also halts proliferation of a neuroblastoma cell line in culture as predicted. Next, exposure to vapors slows progression of neurodegeneration in a Drosophila model for Huntington’s disease. These changes strongly suggest that unbeknown to us, certain volatiles in the surroundings can have profound effects on histone acetylation, gene expression and physiology in animals.
Introduction
Multicellular organisms evolve in the presence of microbial volatiles and metabolites (1, 2), often for prolonged periods of time. Human-associated microbes have incredible metabolic diversity, encoding 100x as many genes as the human genome itself(1). It is estimated that about half of the thousands of metabolites found in mammalian blood are either produced or altered by microbes (3). Microbial metabolites are an important mechanism by which the microbiome influences host immunity amongst other things (2, 4). Many volatiles are generated from the metabolism of a variety of food components, including triglycerides, sugars and amino acids (5). Some chemicals are also produced by microorganisms such as yeasts and lactic bacteria during fermentation in many foods and beverages (5).
In the model system Drosophila melanogaster, prolonged exposure to CO2 showed an increase in volume of the CO2-sensing glomerulus in the antennal lobe, which can lead to adaptations in fly behavior (6, 7, 8). Carbon dioxide is a common byproduct of yeast fermentation in overripe fruits, the preferred food source. In order to study the physiological effects of long-term exposure to other volatiles emitted during fermentation, we exposed flies for 5 days to diacetyl, a buttery odorant made at high levels by yeast in fermenting fruits (1350 ppb in ripe banana), that is also found in many commonly consumed foods such as butter, yoghurt, wine, fruits, beer, popcorn (5, 9–11). Diacetyl was selected for two reasons: first, Drosophila melanogaster use fermenting fruits as their favored food source, oviposition site and spend long durations of time exposed to it. Second, diacetyl is known to be an inhibitory odorant of the CO2 receptor and therefore an opposite effect is expected on the V-glomerulus from the previous study (12). Follow up experiments revealed that diacetyl acts as an HDAC inhibitor in vitro. We found that volatiles structurally like diacetyl have similar HDAC-inhibitory properties as well.
Subsequent analyses with two different model systems: Drosophila melanogaster and Mus musculus showed gene expression changes in tissues caused by odor exposure from a distant source. We found expression of hundreds of genes to be modulated upon exposure to the volatile, as would be expected from an inhibitor of HDACs. HDACs are histone-modifying enzymes involved in the removal of acetyl groups from lysine residues and the remodeling of chromatin structure to regulate gene expression (13, 14). With large-scale roles in gene regulation, HDACs are promising targets in drug development for many diseases such as cancers and neurodegenerative disorders (15–17). Several classes of orally administered HDAC inhibitors have been found to attenuate the progression of certain cancers and neurodegenerative diseases including Alzheimer’s disease and Huntington’s disease (17). We determined that exposure to diacetyl volatiles substantially altered gene expression in the mouse brain, presumably by crossing the blood-brain barrier. Diacetyl exposure also slowed degeneration of photoreceptor cells in a Huntington’s disease model in Drosophila. Last, exposure to diacetyl caused dramatic reduction in proliferation of a neuroblastoma cell line in culture. Our discovery of a family of volatile odorants that also act as HDAC inhibitors highlights the need to understand how small molecules present in our environment interact with and alter eukaryotic gene expression.
Results
Activity-dependent expression modulation of chemosensory receptors
Previous studies demonstrated that prolonged exposure of ∼5 days to elevated CO2, causes an increase in volume of the CO2-sensitive V-glomerulus in the antennal lobe, which can lead to adaptations in fly behavior (6, 7, 8). To assess the effect of CO2 response inhibitor diacetyl on the antennal lobe, flies were exposed to the odorant during the first days immediately following eclosion. The CO2-detecting ab1C neuron was labelled using the promoters of the CO2 receptors, Gr63a-Gal4; UAS-mcd8GFP and Gr21a-Gal4; UAS-mcd8GFP, and the flies were exposed to headspace above a 1 % (V/V) diacetyl solution in an air-tight container. The flies were then imaged for expression of GFP in the ab1C neurons of the antenna, as well as the V glomerulus, which receives axonal input from ab1C neurons in the antennal lobe (18). Surprisingly, the Gr63a promoter driven GFP signal in the V glomerulus was completely abolished after 5 days of exposure to the odor for all brains imaged (Figure 1A,1B). However, there was also a concomitant decrease in GFP signal in ab1C neurons in the antenna (Figure 1A). Similar results were seen for Gr21a driven expression, where expression of GFP was lost in both the V-glomerulus and ab1C neurons (Figure S1A, S1B). Interestingly, after five days of recovery in clean air the GFP expression was regained in the V-glomerulus as well as in ab1C neurons in the antenna, indicating that both Gr21a and Gr63a expression had been restored (Figure S2). Because olfactory neurons do not regenerate in the fruit fly (19, 20), these results suggest that changes in GFP levels are likely due to changes in the promoter expression and not due to neuronal cell death.

Volatiles found in microbes can inhibit HDACs in vitro
(A) Antennal and whole mount brain staining of Gr63a-Gal4 flies. Flies were exposed to diacetyl (headspace from 10-2 soln) or air for 2 to 5 days. Brains and antenna were dissected on the indicated days, fixed, and then stained for neuropil marker nc82 (red) and anti-GFP (green). (B) Mean of ab1C neurons expressing GFP after indicated days of odor exposure. d4on=2,3-butanedione. n=6, error bars=s.e.m. Schematic chemical structures of diacetyl, β-hydroxybutyrate and Sodium butyrate. (C) Dose-activity curves of class I HDACs: HDAC1, HDAC2, HDAC3, HDAC8 and class II HDAC6 treated with various concentrations of diacetyl. Percentage of HDAC activity is relative to the activity of each enzyme without diacetyl. IC50s are indicated in the chart areas. Error bars, S.E.M., n = 4-5. (E) Representative structures of odorants that inhibit HDACs, and (F) Average percentage inhibition of class I HDACs: HDAC1, HDAC3 and class II HDAC4, HDAC6 treated with 15mM of indicated volatiles. Error bar= S.D., each tested in duplicate.
In order to test whether expression of Gr63a and Gr21a is being downregulated by diacetyl we relied on quantitative-PCR along with a few members of the Odorant receptor (Or) family genes. Apart from Gr63a and Gr21a, we also evaluated receptors where diacetyl has no activity (Or47a, Or88a) and the broadly expressed co-receptor (Or83b). Wild-type flies that were exposed to diacetyl for 6 days showed dramatic downregulation of all genes tested. However, after a 5-day recovery period, expression of all but one (Or88a) of the genes was recovered, including Gr63a (Figure S1C). This form of plasticity in gene expression is unexpected and therefore mechanistically interesting to investigate.
Diacetyl and structurally related odorants act as HDAC inhibitors in vitro
Diacetyl is structurally related to the soluble metabolite secreted by the gut microbiome and liver, β-hydroxybutyrate, which is known to inhibit zinc-dependent HDACs (21). It is also structurally related to another known HDAC inhibitor Sodium butyrate. Diacetyl is present widely in nature as a pH-neutral fermentation product of microorganisms such as yeasts in over-ripe fruit, the favorite food for Drosophila, and also from lactic acid bacteria, and is found in many foods and beverages (5). In humans, diacetyl is also produced by microbes on the skin, in the oral cavity, and is found in breath (22). Interestingly, this volatile is known to traverse the cell membrane (23).
Soluble metabolites from the gut microbiome, such as short chain fatty acids, are known to affect histone modifying enzymes, gene expression, and physiology in various parts of the host (1, 24). It is presumed that these dissolved compounds absorbed in the blood circulate to various parts of the body. Microbes in our skin microbiome, mouth microbiome, and environment also emit a diverse repertoire of volatile metabolites that can traverse much greater distances away from the source (25). To examine if diacetyl can directly modulate HDACs like β-hydroxybutyrate, we performed in vitro acetylation assays with recombinant human HDACs. We found that indeed, diacetyl inhibited all four purified human Class I HDACs (HDAC1, 2, 3 and 8) and the one Class II HDAC (HDAC6) tested. The inhibition occurred in a dose-dependent manner. The IC50 values for HDAC1, 2, 3, 8 and 6 were 7.3 mM, 23.1 mM, 7.5 mM, 14.3 mM, and 24.5 mM respectively (Figure 1C). The levels of inhibition as observed from IC50 values for HDAC1 and HDAC3 were comparable to those of β-hydroxybutyrate whose IC50 values for HDAC1 and 3 were previously reported as 5.3 and 2.4 mM, respectively (21).
To test other odorants for their ability to inhibit HDACs, we identified a list of volatile odorants structurally similar to diacetyl using the similarity search feature on PubChem (Figure 1D) and are volatile and tested them on four purified human HDACs, from Class I (HDAC1 and HDAC3) and Class II (HDAC4 and HDAC6). Each compound was tested at 15 mM, the concentration at which diacetyl inhibits HDAC1, 3, and 6 with more than 50% efficacy (Figure 1C, D). All compounds tested reduced the activity of at least one of the HDACs (Figure 1D). Ethyl pyruvate, methyl pyruvate, 2,3-hexanedione, 2,3-heptandione, 2,3-pentandione inhibited HDAC1, 3 and 6 with more than 70% efficacy. Allyl butyrate also similarly inhibited those HDACs but with lower efficacy. 1-acetoxyacetone, and 2,3-butanediol inhibited HDAC6 more than 70% but inhibitory effects against HDAC1 and 4 were lower. Moreover, propyl formate strongly inhibited HDAC 6 but did not inhibit HDAC1, 3, and 4. Taken together, these results indicate that microbial volatiles and structurally related odorants can inhibit human HDACs, and that each compound has specific HDAC inhibitory properties.
Microbial volatile increases histone acetylation in human cell nuclei
In order to directly examine histone acetylation at the cellular level, we used human HEK293 cells, which offer a tractable system to prepare nuclear extracts, and picked the volatile diacetyl to test in more detail. We exposed the cells to different doses of diacetyl for 2 or 6 hours and monitored histone acetylation levels within nuclear extracts in Western Blot assays. Compared to the mock treatment, 10 mM diacetyl significantly increased H3K9 acetylation levels within 2 hours of treatment, whereas the acetylation levels of H3K14 and H4K5 were not affected (Figure 2A). This specificity for an increase of the H3K9 mark is consistent with previous observations for β-hydroxybutyrate. Preferential acetylation of H3K9 was also observed previously in HEK293 cells with treatment of the structurally related β-hydroxybutyrate (21). After 6 hours of treatment, the H3K9 acetylation induced by 10 mM diacetyl was further increased (Figure 2B). The increase in H3K9 acetylation with diacetyl treatment was dependent both on the duration of exposure and concentration of the odorant.

Diacetyl increases level of histone acetylation in HEK293 cells
(A and B) Representative images from Western blots showing acetylation levels of H3K9 (left), H3K14 (middle) and H4K5 (right) in HEK293 cells after 2 hours (A) and 6 hours (B) of diacetyl exposure. PCNA (Proliferating cell nuclear antigen) is a 29 kDa nuclear protein used as a loading control for nuclear protein extracts. (C) Western blots showing acetylation levels of H3K9 in HEK293 cells treated with 100 μM diacetyl for 72, 96, and120 hours. PCNA is used for a loading control. (D) Bar graph showing the relative intensities of acetylated H3K9 in HEK293 cells treated with 100 μM diacetyl for 72 -120 hours. n = 4 samples, * p < 0.05, and **** p < 0.0001.
Organisms are constantly exposed to volatiles commonly found in their food and environment for prolonged periods of time. In order to test the effect of a lower concentration of the volatiles, we selected a 5-day exposure time at a 100-fold lower concentration. When we treated HEK293 cells with this lower dose of diacetyl (100 μM), H3K9 acetylation level increased after 96 hours of exposure and reached significantly higher levels than control after 120 hours (Figure 2C,D). These results demonstrate that even prolonged exposure to low levels of diacetyl can greatly impact the epigenetic landscape inside the nucleus. More importantly, a 5-day exposure was sufficient to alter the epigenetic state of nuclei at concentrations that are present in some food sources. Taken together, these results demonstrate that diacetyl can act as an HDAC inhibitor, with the potential of causing broad modulations of gene expression and histone acetylation in cells.
Transcriptional response to odor exposure is conserved in invertebrates and vertebrates
We next performed in vivo experiments to determine whether eukaryotic cells alter gene expression in response to diacetyl exposure, as would be expected with HDAC-inhibition. Epigenetic changes occur slowly over days, and to test its effects on gene expression we used two model systems for eukaryotes: invertebrates (Drosophila melanogaster) and vertebrates (Mus musculus). We placed Drosophila adult males in vials and housed these vials within a closed container exposed to headspace from a 1% diacetyl solution (in paraffin oil) for 5 days, similar to a previous long-term odor-exposure study (7) (Figure 3A). The transcriptome of the primary olfactory organ, the antenna, was compared with that of the control group of age-matched flies that were exposed to the solvent paraffin oil (PO). The antennal transcriptional profile of diacetyl-exposed flies showed substantial changes in gene expression when compared to the solvent control (Figure 3B). We identified 1234 differentially expressed genes (DEGs) (false discovery rate, FDR < 0.05) in the antennal transcriptome of diacetyl-exposed flies compared to control animals. Of these, 645 genes were significantly up-regulated (log2 fold-change > 1; red dots in Figure 3B) and 589 genes were significantly down-regulated (log2 fold-change < -1; blue dots in Figure 3B). A broad range of genes was significantly altered, with several biological process GO terms significantly enriched in the up-regulated gene list including “response to biotic stimulus” (p < 3x10-7),“response to bacterium” (p < 3x10-7) and “defense response” (p < 3x10-7). In the down-regulated gene set the GO term most enriched was “sensory perception of chemical stimulus” (p< 6x10-72). The GO molecular functions that were most common in the upregulated sets included enzymes like hydrolases and oxidoreductases, as well as nucleic acid binding genes (Figure 3C). Down-regulated genes include receptors, hydrolases, and transporters (Figure 3C).

Remote exposure to HDACi volatile alters gene expression in an invertebrate, and vertebrate.
(A) Schematic of odor exposure protocol for transcriptome analysis from the antennae. (B) Plot highlighting up- and down-regulated genes in the diacetyl-exposed group. Red and blue dots represent up-regulated genes (false discovery rate (FDR) < 0.05, log2 fold change (LFC) > 1) and down-regulated genes (FDR < 0.05, LFC < 1), respectively. (C) Bar graphs denoting the protein classification of the genes up- and down-regulated after odor exposure. (D) Schematic of diacetyl exposure protocol for transcriptome analysis of mouse lung tissue. (E) Plot highlighting up- and down-regulated genes in the diacetyl-exposed groups. Red and blue dots represent up-regulated genes (FDR < 0.05, LFC > 1) and down-regulated genes (FDR < 0.05, LFC < 1), respectively in lungs. (F) Table showing pairwise tests of significance of overlap between gene sets. P-values from Fisher’s exact test, colored with associated odds ratio (strength of association). (G and H) Bar graphs denoting the protein classification of the genes up- and down-regulated in the lung after exposure to headspace above 0.1% (v/v) (G) and 1% (v/v) (H) of diacetyl solutions.
The first tissue that would have contact with airborne volatiles in terrestrial vertebrates are the airways and lungs. We therefore performed transcriptome analyses on lung tissue of mice exposed to diacetyl headspace at different doses for a period of 5 days, as was done in Drosophila (Figure 3D). The dose of the volatile in the chamber was measured in parallel experiments by tracking volatile loss and airflow and determined to range between a peak 1920 ppm at the end of hour 2, tapering off to 28 ppm by hour 120 (Supplemental Figure S3B). Indeed, expression of a substantial number of genes was modulated in the diacetyl-exposed lungs compared to the control. The changes were dose-dependent and more pronounced in mice exposed to headspace from a 1% diacetyl source compared to those exposed similarly to 0.1% (v/v) diacetyl source (Figure 3E, Supplemental Figure S3C). Among these diverse sets of regulated genes in the lung tissue, a significant overlap was found between 1% (v/v) and 0.1% (v/v) headspace for both up-regulated genes (p=3x10-3) and down-regulated genes (p=6x10-3, Figure 4F), further supporting a dose-dependent effect of diacetyl on gene expression in the mouse lung.

Gene expression changes are partly reversible, and overlaps with known HDACi drugs
(A) Schematic of odor exposure and recovery protocol for transcriptome analysis from the antennae. (B) Plot highlighting up- and down-regulated genes in the recovery from diacetyl exposure group. Red and blue dots represent up-regulated genes (FDR < 0.05, LFC > 1) and down-regulated genes (FDR < 0.05, LFC < 1), respectively. (C) Table showing pairwise tests of significance of overlap between gene sets. P-values from Fisher’s exact test, colored with associated odds ratio (strength of association). (D and E) Plots showing enrichment of up- and down-regulated genes in sodium butyrate-(D) and valproic acid-treated (E) groups. Red and blue dots represent up-regulated genes (FDR < 0.05, LFC > 1) and down-regulated genes (FDR < 0.05, LFC < 1), respectively. (F) Left: Venn diagrams showing the overlaps of up-regulated genes among diacetyl-, sodium butyrate- and valproic acid-treated groups. Right: Table showing pairwise tests of significance of overlap between gene sets. P-values from Fisher’s exact test. (G and H) GO-enrichment analysis of the common Up-regulated and down-regulated genes across all 3 treatments in (F).
The GO enrichment analysis of the lung transcriptome from mice exposed to diacetyl revealed several interesting sets of genes that were significantly altered (Figure 3G,H). Among these genes is the SARS-CoV-2 entry receptor ortholog, ACE2. The level of this gene in the lungs of diacetyl exposed mice showed a reduction in expression levels (log2 fold-change= - 0.62, FDR= 2.02x10-5). The level of ACE2 in the nasal tissue, lungs and other organs play a critical role in infection by the SARS-CoV-2 virus, suggesting that epigenetic mechanisms may contribute to expression. While we do not know whether expression of ACE2 in humans is regulated similarly, these results highlight the importance of understanding volatile-induced modulation of gene expression in the lungs and other tissues.
HDAC family members are highly conserved across eukaryotes, spanning both animal and insects. Volatile microbial metabolites have been present throughout the evolution of animals, insects, and plants, and have potential as signals for multi-domain communication because they can travel wide ranges; plants detect root infections and time their immune response using volatiles (26), and microbial volatiles elicit olfactory behaviors in insects and nematodes. Our results indicate that a volatile compound, capable of directly inhibiting HDACs and altering acetylation levels, can cause significant changes in gene expression in a variety of organisms following exposure from a distance, through the air.
DEGs overlap those for known HDAC inhibitors and upregulation is partially reversible
One of the hallmarks of epigenetic regulators which makes them attractive candidates for therapeutics is plasticity. Upon removal of the inhibitors, we expect that some of the changes in gene expression are reversible. To test for reversibility, we performed a recovery experiment following diacetyl exposure using the Drosophila model. We maintained 5-day diacetyl-exposed flies in clean air for 5 additional days (Figure 4A). In parallel, we performed age-matched mock experiments with paraffin oil solvent exposure alone. A large number of genes were down-regulated following the recovery in comparison to the 10-day-old flies in the mock condition (Figure 4B). Interestingly, there was a significant overlap of these down-regulated genes with the set that was up-regulated in the diacetyl treatment (Figure 4C). These results suggest that the effects of HDAC inhibitory odorant exposure are not permanent but dynamic, and removal of the odorant leads to subsequent changes in gene expression of the up-regulated set.
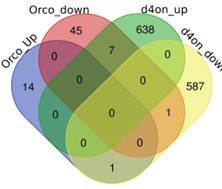
Drugs that inhibit HDACs are in various stages of approval as treatments for several diseases. The approved drugs include sodium butyrate and valproic acid. To compare the gene-regulatory effects of these drugs to diacetyl, we performed RNA-seq after raising the flies on food containing sodium butyrate (SB) or valproic acid (VA) (27) and compared gene expression in the antennae of flies raised on untreated food for 5 days. We next compared the up-regulated gene profiles following each treatment to the one induced by exposure to diacetyl. As expected, feeding SB and VA induced significant changes in expression levels of several genes (Figure 4D,E). Interestingly we found that 113 of diacetyl up-regulated genes were also up-regulated in either SB, VA or both treatment conditions (Figure 4F). Pairwise statistical analysis of each gene set revealed a significant overlap of diacetyl-induced genes with SB-induced genes (p=6x10-11) and with VA-induced genes (p=2x10-65) (Figure 4F). There was, as expected, also a significant overlap between SB- and VA-induced genes (p=1x10-52) (Figure 4F). This highly significant overlap among up-regulated genes lends further support to our model that diacetyl vapors act as an HDAC inhibitor in vivo. As expected, each of the 3 treatments also modulated a substantial number of unique genes (Figure 4G, H), suggesting that differences in oral vs. vapor delivery, molecular structure and inhibition profile across the repertoire of HDACs may contribute to differences in gene regulation. These results suggest that some volatile odorants, or the microbes that emit them, may impart therapeutic effects like other HDAC inhibitors from a distance, through the air.
Volatile Diacetyl alters gene expression in the mouse brain
One of the categories of challenging diseases that HDAC inhibitors are being tested for drugs are neurodegenerative diseases (28) where volatile odorants may provide a new class of inhaled therapeutics. A major design challenge for nervous system drugs is the ability to cross the blood-brain barrier. Due to the volatility and small size of volatile odorants, there is a possibility that they could diffuse through the intranasal route to the brain directly (29). We cannot tell a priori whether odorants like diacetyl can travel across the nasal membrane to the brain. In order to test whether cells in the brain of mice respond when presented with diacetyl vapors from a distance, we used gene expression as the final readout to measure it. We performed RNA-seq experiments on mice exposed only to aroma of diacetyl in the air for 5 days as before. Littermate controls were exposed in a similar manner to the solvent (PO) in the air. Interestingly, several genes were differentially expressed upon exposure to headspace from 0.1% (v/v) diacetyl passaged through the air in the holding cage where it is further diluted (49 up-regulated, 32 down regulated, |log2 fold-change| >1, FDR<0.05) or to headspace from 1% (v/v) diacetyl diluted in the holding cage (748 up-regulated, 1031 down regulated, |log2 fold-change| >1, FDR <0.05) (Figure 5B,C). GO analysis of the regulated genes in the exposed mouse brain transcriptome revealed several interesting sets of genes were significantly altered in each set (Figure 5E). These results indicate that a volatile odorant like diacetyl can reach the brain in mammals, and presumably alter gene expression in neuronal cells. Although the overall DEG sets were different across the lungs and brain, there was a statistically significant overlap, particularly in the 1% exposure, as would be expected from using the same mechanism of action. There was a highly significant overlap (p=2x10-101) between genes upregulated in the brain compared with the lungs (248 of 748), as well as for downregulated genes (p=3x10-163; 477 of 1031 genes) (Figure 5D).

Exposure to diacetyl vapor alters gene expression in the mouse brain
(A) Schematic of diacetyl exposure protocol for transcriptome analysis of mouse brain tissues. (B and C) Plot showing up- and down-regulated genes in the diacetyl-exposed groups. Red and blue dots represent up-regulated genes (FDR < 0.05, LFC > 1) and down-regulated genes (FDR < 0.05, LFC < 1), respectively in the brain. (D) Left: Table showing pairwise tests of significance of overlap between gene sets. P-values from Fisher’s exact test. Right: Venn diagrams showing the overlaps of DEGs between lung and brain groups. (E) Bar graphs denoting the protein classification of the brain genes up- and down-regulated after 1% diacetyl exposure. (F) Mean fold change of select neuroblastoma related genes in the brain of mice exposed to diacetyl vapors. (G) Cell counts of indicated cancer cell lines in tissue culture treated with solvent control or indicated concentration of diacetyl. N=3-6, p<0.001.
Diacetyl prevents proliferation of neuroblastoma cells
Among the significantly downregulated genes was MYCN, which is known to show increased expression and plays a central role in many types of cancers, suggesting that epigenetic regulation may affect tumor cells. Broad-spectrum HDAC inhibitors have been approved by the FDA for treatment of a variety of cancers. To test the exciting possibility that a volatile HDAC inhibitor, which affects gene expression in the brain, would have an effect against cancers as well, we performed a cell-based assay. We picked neuroblastoma as a candidate since it has a strong dependence on MYCN and develop from fetal adrenal neuroblasts (30). Several genetic and “omics” studies have identified genes that are upregulated in neuroblastomas, some of which are considered to be key causative factors (31). We evaluated the DEGs from the diacetyl-exposed mouse brain transcriptome for several of these key genes. Three of the six key genes identified across multiple studies were significantly altered, two of which were significantly downregulated, MYCN (P=2.06E-13, FDR=1.76E-12) and Alk (P=3.3E-5, FDR=9.48E-5) (Figure 5F). An unbiased computational study evaluated omics datasets and identified 4 genes that were the best predictors of clinical outcome (32), all of which are significantly different in the brains of the diacetyl exposed mice, strikingly 3 of which were significantly downregulated, NCAN (p=1.33E-16, FDR=1.59E-15), STK33 (p=4.6E-7, FDR=1.79E-6) and ERCC6L2 (p=4.57E-5, FDR=0.00012) (Figure 5F). These findings are extremely provocative, given that the downregulated genes, particularly MYCN play a role in neuroblastomas and in several other cancers.
We tested diacetyl for the ability to inhibit proliferation of a SH-SY5Y neuroblastoma cell line from a metastatic bone tumor. A significant reduction in cell count was seen in diacetyl treatments in the SH-SY5Y cancer cell lines (Figure 5G). To validate the initial result, we retested diacetyl with at two concentrations across 3 different cancer cell lines. Diacetyl showed a significant reduction in cell numbers for the neuroblastoma cell line specifically, and not any other cell lines, relative to the control (Figure 5G). These results demonstrate that the effect is specific to the SH-SY5Y cancer cell line, and unlikely to be caused by a non-specific general toxic effect. It also suggests that the potential epigenetic regulation of cancer promoting genes may be caused by exposure to volatiles like diacetyl in the environment.
Testing therapeutic effect in neuroblastoma and Huntington’s neurodegeneration
Since diacetyl can affect changes in a mammalian brain by crossing the blood brain barrier, we wanted to test if it can impart therapeutic effects in disorders of the brain like neurodegeneration for which HDAC inhibitors have been proposed as therapeutics. In order to test diacetyl in the context of neurodegenerative disease, we used the well-established Drosophila model of Huntington’s disease, which has previously been used to demonstrate the efficacy of sodium butyrate in slowing neurodegeneration (27, 33). The human Huntingtin protein with expanded poly-Q repeats is expressed in the neurons of the compound eye in Drosophila, causing progressive degeneration of the seven visible photoreceptor rhabdomere cells in each ommatidium (34). We selected this model in particular since polyglutamine disorders are well suited for targeting by inhibitors of HDAC1 and HDAC3 (35) such as diacetyl (Figure 1B). Moreover, previous studies have shown that orally administered HDAC inhibitors such as sodium butyrate and SAHA can significantly reduce photoreceptor degeneration in this model (27). When the transgenic flies expressing two copies of the human Huntingtin with poly-Q repeats (HTTQ120) under control of the eye-specific GMR promoter were raised at 18°C, the number of rhabdomeres in each ommatidium was similar to that normally observed in wild-type flies immediately post-eclosion (day 1, Figures 6A-C). When these HTTQ120 flies were moved to 25° C following eclosion (Figure 6A), they showed dramatic degeneration of rhabdomeres over a period of 10 days (Figures 6B, D-G). The mean number of rhabdomeres was reduced from 7 to ∼1 by day 10. The dose of the volatile in the chamber was measured in parallel experiments by tracking volatile loss and airflow (0.5 L/min) and determined to range from 260-95 ppm, an average of 125 ppm at the end of hour 1, tapering off to 0 ppm by hour 24. The odorant was replaced each day, leading to a dosage profile that would seem like a once-a-day treatment regimen (Supplemental Figure S3A).

Odor exposure slows Huntington’s neurodegeneration model in fly eye
(A) Schematic diagram showing temperature of experimental condition and timing of the eye examination in pGMR-HTTQ120 flies. (B) Bar graph showing mean number of rhabdomeres in each ommatidium in solvent paraffin oil (PO, blue) and diacetyl-exposed (red) pGMR-HTTQ120 flies at 1, 5, and 10 days after eclosion (AE). n = 600 ommatidia from 15 flies, **** p < 0.0001. (C) A representative image of ommatidia of pGMR-HTTQ120 flies at 1 day AE. (D and E) Representative images of ommatidia of pGMR-HTTQ120 flies exposed to paraffin oil (PO) or diacetyl at 5 (D) and 10 (E) days AE. (F and G) Histogram showing the percent of the ommatidium with a given number of rhabdomeres indicated on the x axis at 5 (F) and 10 (G) days AE.
Remarkably, when the Huntingtin (HTTQ120)-expressing flies were exposed immediately after eclosion to volatile headspace of 1% diacetyl (Figure 6A), they showed a substantial (∼50%) reduction of rhabdomere loss (Figures 6B, D-G). The majority of ommatidia retained 6-7 rhabdomeres at day 5 (Figures 6D and F). Even after 10 days, the majority of the ommatidia still had 2-3 rhabdomeres left in the odor-exposed flies, while the solvent controls had only ∼1-2 rhabdomeres (Figures 6E and G). These results demonstrate that exposure to volatile diacetyl in air slows down the photoreceptor degeneration caused in these Huntington’s disease model flies. Thus, odorants like diacetyl may have potential as a prophylactic against this neurodegenerative disorder. Therefore, the volatile odorant can both significantly alter gene expression from a distance as well as cause a phenotypic change in the fly with a human disease gene.
Discussion
In this study, we discovered that eukaryotic cells could alter gene expression in response to an odorant, in a manner independent of traditional neuronal activity-induced pathways. Members of a conserved family of HDACs detected the concentration of diacetyl differentially. Diacetyl exposure increased levels of H3K9 acetylation in the nucleus in a dose-dependent manner and led to changes in gene expression. The odorant diacetyl occurs naturally and is generated from the metabolism of a variety of food components, including triglycerides, sugars and amino acids (5). Moreover, the chemical is produced by the activity of microorganisms such as yeasts and lactic bacteria during fermentation in many foods and beverages (5). Although beyond the scope of this study, the discovery that cells can alter gene expression upon prolonged exposure to a common naturally occurring odorant raises important questions about the potential physiological consequences of the expression changes. Given our repeated exposure to particular flavors and fragrances, the findings outlined here highlight a new consideration for evaluating the safety of certain volatile chemicals that can cross the cell membrane.
In mammals, upregulation of circulating β-hydroxybutyrate during fasting or calorie restriction induces changes in the expression of a set of genes (21). The acetylation mark specificity and IC50 of β-hydroxybutyrate is similar to that of diacetyl. Any beneficial physiological effects of odor-detection at lower odor concentrations would also be countered by studies of potential risks at higher concentrations. In fact, the deleterious effect of exposure to high levels to diacetyl causing bronchiolitis obliterans, or “popcorn lung”, and its toxicity in cultured cells is already known (36). Even so it is present in several foods we eat and is on the GRAS (Generally Regarded as Safe) list of Flavor and Extract Manufacturers Association (FEMA) for use as a flavoring ingredient at low concentrations. While the specific mechanism of “popcorn lung” is unknown, it has previously been proposed that carbonyl groups of diacetyl can react to modify amino acid side chains on proteins or generate reactive dicarbonyl and reactive oxygen species, leading to excessive cytokine production and inflammation (37). Our results raise the possibility that the molecular mechanisms underlying this disorder could also be partially attributed to the unusually high HDAC-mediated genetic response to diacetyl by cells in the lungs. Several studies using rodent animal models have shown toxicity of high levels of diacetyl in the respiratory tracts including the nose and lungs (38, 39). The toxic effect of higher dosages has been observed for many types of HDAC inhibitors. For example, in one Drosophila study, feeding of a high dosage of 4-phenylbutyrate reduces survival rate, while a lower dosage extends longevity (40). Another concern is that one of the highest known levels of diacetyl exposure to humans is through smoking cigarettes, where exposure from even a single cigarette (250-361ppm) is high (41, 42): Salem (128 μg/cigarette), Camel (128.1 μg/cigarette), Carlton (45.6 μg/cigarette) (42). Different types of diets and lifestyle choices have been known to have effects on health and well-being due to differences in balance of different health promoting ingredients, yet little is known about role of such flavors, fragrances and metabolites acting on epigenetic marks and altering gene expression. Our findings raise the possibility that diacetyl and structurally related odors in the environment may affect gene expression directly through absorption/transport into these cells. In mammals, the epithelium-lined lungs and nasal/oral pathway is directly exposed to odorants presenting an opportunity to affect gene expression and physiology.
Eukaryotes primarily detect volatile odorants in the environment using olfactory neurons that express a variety of transmembrane receptors, a family of genes that have independently evolved multiple times in different phyla. Examples of these unrelated genes include: the ionotropic 7-transmembrane (TM) odorant receptor (Or) family, which is insect-specific; 3-TM ionotropic receptors (IRs), which are present across most arthropods; the nematode-divergent 7-TM GPCRs belonging to the str, sra, srg, srw, srz, srbc, srsx and srr families; the mammalian 7-TM GPCR olfactory receptor (OR) family; and the trace amine-associated receptor (TAAR) family (43–47). The activation of these receptors induces neuronal action potentials, and this information is conveyed to higher brain centers where olfactory perception is generated (48). This second response pathway which we have discovered is much slower and has an enzymatic mechanism which draws parallels with the multiple pathways in place to detect light. In the specialized cells of the eye, detection of light occurs via rhodopsins (7-transmembrane GPCRs) and leads to neuronal activity and behavioral responses. However, other cells, including in plants, are also able to respond to changes in light intensity of certain wavelengths using the ancient, conserved cryptochrome proteins that are related to photolyases (49). This suggests that vital sensory modalities can be detected via multiple pathways. Our observations raise the question whether HDACs represent a conserved detection mechanism for odorants like diacetyl that link odor-detection to a specific gene expression response. HDAC proteins, however, are an ancient family of genes that predate even histones themselves (50, 51). It is conceivable that odor detection mechanisms that emerged in ancient forms may not have involved specialized transmembrane receptors, or neurons, or resulted in rapid behavioral movements.
Our data suggests that an HDAC inhibitory odorant also affects gene expression in the lungs and brain of mice, including significantly decreasing the expression level of the ACE2 receptor, whose ortholog in humans is the entry receptor for the SARS-CoV-2 virus that causes COVID-19. The applied impact for the future could be far-reaching, as HDACs are known to affect a wide variety of pathways in eukaryotes, and inhibitors are candidates for novel inhaled therapeutics as treatment of many conditions such as COVID-19, cancers, neurodegeneration and a variety of infectious diseases (52–60).
Our studies also show that one of the volatiles is protective against the neurodegenerative effects of Huntington’s disease model and neuroblastoma. Exposure to diacetyl volatiles in the fly model of Huntington’s disease reduces cell degeneration, as has been previously observed with orally administered HDAC inhibitors like sodium butyrate and SAHA in this genetic model (27). Previous studies indicate that the inhibition of HDACs counter the acetyltransferase inhibitory activity of the polyglutamine-domain of the Htt protein which binds to p300, P/CAF and CBP (27).
This newly discovered activity in this class of volatile chemicals will also allow us to understand how their presence in our environment, diet and microbiome shapes the epigenetic landscape in the lungs and affects inflammation and immune responses. Our approach to investigate consequences in model systems allows for a foundation for future research and allow identification of key molecular biomarkers that can be associated with observation of epigenetic effects, toxic or not, in individuals. The public health impact of these findings can be far-reaching, both from the perspective of toxicity as well as therapeutics. Investigating the health safety concerns of exposure to such odorants at high doses or prolonged periods is essential to understand potential for damage. Conversely, at non-toxic concentrations HDAC inhibitors can be used for treatment of many debilitating conditions in the lungs such as COPD, inflammation, and COVID-19 infection (61).
Taken together, our discovery that cells can alter gene expression in response to an odor that acts as an HDAC inhibitor and reprogram gene expression, promises the pursuit of new types of odor-based therapeutics already approved for human consumption as prophylactics for a myriad of diseases. Simultaneously, they raise the concern of a new type of environmental agent which can reprogram gene expression and have widespread effects that are yet unknown.
Methods
Drosophila Stocks and Manipulations
Fly stocks were maintained on conventional fly food under a 12-hr. light:12 hr. dark cycle at 18°C or 25°C. The fly strain of w1118 backcrossed 5 times to Canton-S (wCS) was used in all the Drosophila transcriptome experiments. P{GMR-HTT.Q120}2.4 (Bloomington # 8533) were used for neurodegeneration experiments. For imaging Gr63a-Gal4; UAS-mcd8:GFP, Gr21a-Gal4; UAS-mcd8:GFP stocks were obtained as a kind gift from the Carlson lab, Yale.
Odor exposure experiments for imaging
Newly eclosed male wCS flies were sorted in groups of 30 into vials with standard cornmeal medium. Vials were closed with two overlapping 3’’x3’’polypropylene mesh squares and tied with cotton twine. Vials were used within 6 hours. Four vials were placed in a 1000ml Nalgene straight sided jar with a 10ml glass beaker with 10ml of diacetyl 10-2 in paraffin oil. Jars were closed for exposure periods ranging from 2-6 days. In recovery experiments, vials were removed from the jars and were placed in a 25°C incubator for the remainder of the recovery period. Flies were anesthetized on ice, sacrificed in ethanol, and then immediately put into 1X phosphate buffered saline with 0.3% triton-x (PTX). Brains or antenna were put in 4% paraformaldehyde in PTX and incubated for 30min while rotating at 25°C. Samples were washed 5 times in PTX, and blocked in 5% natural goat serum in PTX for 1-hr while rotating at 25°C. Samples were then incubated in primary antibody with 5% goat serum in PTX, mouse-nc82 (1:20) (Development Studies Hybridoma Bank, University of Iowa) and rabbit-antiGFP (1:150) (Invitrogen) in PTX for 48 hours in 4°C while rotating. After washing 5 times in PTX samples were incubated in secondary antibody with 5% goat serum in PTX, rabbit-anti-Alexa488 (1:400) (Invitrogen) and mouse-antiAlexa568 (1:400) (Invitrogen) for 48 hours in 4°C. Samples are washed 5 times in PTX and stored in 70% glycerol in PTX. Images were taken using a Ziess 510 laser scanning confocal microscope, and image analysis was done using Image J software.
QRT-PCR
Drosophila heads (with antenna) were collected on liquid nitrogen and stored at -80°C until processing. 300 heads were collected for each sample, and ground on liquid nitrogen. mRNA was extracted using a PolyATract kit (Promega) and was reverse transcribed using Superscript III (Invitrogen) according to the manufacturer’s protocol. The quality of the cDNA was assessed on a 1.5% agarose gel. cDNA was added to individual reactions of SYBR green master mix (BioRad) and run on a BioRad-MyQ thermocycler. The program began with a single cycle for 3-min at 95°C, followed by 40 cycles of 15-sec at 95°C, 30-sec at 58°C and 30-sec at 72°C. Afterward, the PCR products were heated to 95°C for 1-min and cooled to 55°C for 1-min, to measure the dissociation curves. The efficiency of each primer set was first validated by constructing a standard curve and three 10x serial dilutions of first strand cDNA. For each serial dilution, the CT value was plotted against the log (template dilution) and the slope and r2 value of each regression line calculated. Expression of each Or or Gr gene was assessed in triplicate. Dissociation curves were used to assess the purity of the PCR reactions. Or and Gr transcript levels were normalized by using the transcript levels of ribosomal protein 49. Expression levels were calculated using the Pfaffl equation.
In general, primers were designed by using coding sequences close to the 3’ end of the gene, and where possible, primers spanned an intron.
Primers:
Gr21a F: CGATCGTCTTTCCGAATCTC
Gr21a R: GGCTCAGATCCACCCATAGA
Gr63a F: AAATGAACTCCGCCTCCTTT
Gr63a R: CGCAATTTCAGAGGCAAACT
RP49 F: CTGCCCACCGGATTCAAG
RP49 R: GTTTCATGCGGCGAGATCG
Or47a F: ATCACAGGCCACATTGAACA
Or47a R: TCCCCGCAGTAGCAGTAGAT
Or88a F: TTAAAGTGGCCTTCCTGGTG
Or88a R: ATGCGGCAATAAAGTTCCAC
Or83b F: TTCTTGGCATTCGCTTTTCT
Or83b R: TCCCTGGATTTGTTTGCTTC
Odor Exposure Protocol for Transcriptome Analysis
Flies were exposed to diacetyl (B85307, Sigma-Aldrich, St. Louis, MO) by placing them in vials in a cylindrical closed container (112 mm diameter x 151 mm height) along with an odor-containing glass vial. The odorant was dissolved in 5 mL paraffin oil at 1% dilution. For a given exposure protocol, two groups of flies were prepared: those exposed to 1% diacetyl headspace and those exposed to paraffin oil headspace alone (control flies). Adult male flies aged 1 d were transferred to fly vials containing fresh medium and put into the container with the odor vial. At the end of the fifth day of exposure, flies were collected, and their antennae were dissected for RNA extraction. All treatments and experiments were performed at room temperature. For the recovery experiment, flies were transferred to a container with a glass vial of paraffin oil after 5 days of diacetyl exposure. At the end of the fifth day of recovery, flies were collected, and their antennae were dissected for total RNA extraction. The second and third antennal segments from 40-60 male flies after treatment were carefully hand-dissected from the head and collected in 1.5 ml microfuge tubes kept cold in liquid nitrogen. Antennae were mechanically crushed with disposable RNAse-free plastic pestles, and total RNA was isolated using a Trizol-based protocol. cDNA libraries were prepared from total RNA using the Illumina TruSeq RNA Sample Preparation Kit (v2) and 50 bps single- and paired-end sequencing was done using the HighSeq2000. Two biological replicates were sequenced for each condition, with an average of 27 million reads / replicate, and with an average of 84% mapped.
Two-month-old C57BL/6 male mice (2-3 for each condition in a single cage) were continually exposed to air flowing over headspace of paraffin oil (solvent control), 0.1%, or 1% diacetyl over a period of 5 days, then euthanized for collecting the lung and brain tissues and processing for mRNA isolation. All protocols for animal use and euthanasia were approved by the Institutional Animal Care and Use Committee (#20150028) and were in accordance with the provisions established by the Animal Welfare Act and the Public Health Services (PHS) Policy. In the transcriptome analysis, two replicates were performed for each condition, with an average of 123,687,411 reads / replicate, with an average of 88% mapped. Multiplexed libraries were made from total RNA input using the Illumina TruSeq RNA sample preparation kit (v2) and 50 bps single-end sequencing was done using the NextSeq500.
Calculation ppm in air based on weight loss of odorant over time
The odor solution in PO in each setup (Drosophila and mouse) was weighed using a sensitive balance. PO is non-volatile and therefore weight loss is attributable to diacetyl volatilization. Volatile Headspace exposure (ppm) is calculated through odor cartridge weight change from pure odor compound volatizing out (Inamdar et al., 2012).

Where ppm is the concentration of tested VOC in parts per million (v/v); w is the weight of the tested chemical in grams; MW is the molecular weight of the tested VOC (grams/mole); v is the total volume of the respective exposure chambers; and Vm is the molecular volume under the tested condition.

Where 24.45 = gram molecular volume, under standard Lab conditions of 760 mm Hg, 25°C; P = ambient pressure, in mm Hg; and t = ambient temperature, in °C. Odor cartridges, consisting of varying odor volume/volume dilutions, were weighed periodically under standard ambient temperature and pressure. Under these conditions, the pure volatile odor compounds volatize out of the odor cartridge and the evaporation rates were used for our exposure calculations. Based on Ideal Gas Law and experimental conditions, ppm is calculated based on volatized odor compound and experimental chamber volume based on airflow.
HDAC inhibitor Treatment Protocol for Transcriptome Analysis
Sodium butyrate (B5887, Sigma-Aldrich) or valproic acid (P4543, Sigma-Aldrich) were dissolved in normal fly food medium at the final concentration of 10 mM. Three groups of flies were prepared: those treated with one of the HDAC inhibitors and those without HDAC inhibitor treatment (control flies). Adult flies aged 1 d were transferred to fly vials containing medium with or without a HDAC inhibitor. At the end of the fifth day of treatment, flies were collected, and their antennae were dissected for RNA extraction. All treatments and experiments were performed at room temperature. Two biological replicates with 60 flies/replicate were performed for each condition, with an average of 23 million reads / replicate, and with an average of 92% mapped. Multiplexed libraries were made from total RNA input using the Illumina TruSeq RNA sample preparation kit (v2) and 50 bps paired-end sequencing was done using the HighSeq2000.
Bioinformatic analysis of RNA-seq experiments
Reads aligned to the latest release of each of the genomes used (dm6 for the Drosophila genome, GRCm38 for the Mus Musculus genome) and quantified with kallisto (version 0.43.1) (62) Only libraries for which we obtained >75 % alignment were used for downstream analysis. Transcript counts were summarized to gene-level using tximport package (version 1.4.0)(63). For any instances of detected batch effects, we removed unwanted variation using RuvR in the RuvSeq package (version 1.10.0)(64). Differentially expressed gene (DEG) analysis was performed with the edgeR package (version 3.18.1)(65) using low count filtering (cpm >0.5) and TMM normalization. Protein classification analysis was performed with PANTHER (version 13.1)(66). All significance analyses of gene overlap were done using the GeneOverlap package in R package (version 1.14.0). GO enrichment analysis was performed using clusterProfiler (version 3.6.0)(67).
HDAC Activity Assays
HDAC activity of class I HDACs (HDAC1, 2, 3 and 8) was measured with the fluorometric HDAC Activity Assay kit: HDAC1 (10011563, Cayman Chemical, Ann Arbor, MI), HDAC2, HDAC 3, and HDAC 8 (50062, 50073 and 50068, BPS Bioscience, San Diego, CA), according to the manufacturer’s instructions. HDAC activity of class II HDACs (HDAC4 and HDAC6) were measured with the fluorometric HDAC Activity Assay kit: HDAC4, HDAC 4 (50064 and 50076, BPS Bioscience, San Diego, CA), according to the manufacturer’s instructions. HDAC activity in these 96-well format kits are directly correlated with fluorescence produced by the enzyme and substrate interaction. Each test compound was tested in duplicates, minimum. Due to interference, concentrations of >30 mM were not used in the IC50 calculation. To account for any potential absorbance or fluorescence contributed by the test chemicals interacting with the assay substrate, all baseline background measurements of test compound + substrate control wells (everything, but no HDAC enzyme) were performed and subtracted against the respective test compound wells. It is fair to note that no significant changes in background fluorescence were seen with tested compounds in comparison to control background fluorescence. HDAC activity was calculated based on averaged maximum fluorescence from negative control wells and the averaged fluorescence in test compound wells.

HDAC % inhibition was directly calculated from obtained HDAC activity %.
Cell Culture and Treatment
Human embryonic kidney 293 (HEK293) cells were grown in 100 mm cell culture dishes with Dulbecco’s modified Eagle’s medium (DMEM) (10-013, Corning, Manassas, VA), supplemented with 10% fetal bovine serum (FBS) (26140-079, Gibco, Carlsbad, CA) at 37°C with 5% CO2. Cells that were ∼80% confluent were treated with freshly prepared medium supplemented with diacetyl at concentrations indicated. The cells for mock controls were handled in the same manner without adding diacetyl to the medium. To prevent diffusion of diacetyl odor from the treatment dishes to the ones of mock control, the cell culture dishes in different conditions were cultured in separate CO2 incubators.
Cell proliferation assays were performed to assess effects of compounds against cancer cell lines. Cells were maintained in a 37°C incubator with 5% CO2 throughout the whole experiment. Compound stocks are dissolved in 10% DMSO, and final concentrations made fresh in complete media as needed. 10% DMSO solvent is used as control in complete media. Cells were seeded onto 12-well plates and allowed 12-24 hours until treatment to allow cells to adhere to wells.
This is followed by minimum 5-day treatment of compounds, changing the media with treatment every 48 hours. Cell lines A549, PC3, and SK-MEL-5 will be seeded at 8000 cells/well, which was determined to provide 80-100% confluency after 6-day growth. SH-SY5Y was seeded at 8000 cells/well and 16,000 cells/well due to their known slower division rate and given at least 10 days for sufficient growth. After allotted growth times, cell counts were performed using the CountessTM automated cell counter. All assays had a minimum of three replicates for all conditions and analyzed through GraphPad Prism One-way ANOVA for significance.
Odor Exposure Protocol for Huntington’s Disease Model Flies
Flies were exposed to diacetyl in a cylindrical container (112 mm diameter x 151 mm height). Each container was tightly closed but had 2 holes, one of which connected to an air suction port, and the other to a vial containing either of 5 mL paraffin oil or 5 mL 1% diacetyl in paraffin oil. A gentle suction was applied to pull the headspace from the odor or paraffin oil vials into the cylindrical structure. pGMR-HTTQ120 flies were maintained at 18°C. Adult flies aged 1 d were transferred to fly vials containing fresh medium and put into the odor-filled container at room temperature. Paraffin oil and 1% diacetyl solution were prepared and replaced every day. At the end of the fifth day of exposure, half of the flies were collected and subjected to pseudopupil analysis as before (27). The remaining flies were transferred to fresh medium and exposed to the odors for an additional 5 days. All treatments and experiments were performed at room temperature. The number of rhabdomeres in each ommatidium was counted using the pseudopupil analysis, and statistical significance was determined by unpaired t test (two-tailed) against PO at each time point.
Preparation of Nuclear Extracts from HEK293 Cells
Nuclear extracts of HEK293 cells were prepared according to a protocol described previously (68), with minor modifications. In brief, HEK293 cells were washed twice with cold phosphate-buffered saline (PBS) and lysed with hypotonic buffer (10 mM HEPES-KOH [pH 7.9], 1.5 mM MgCl2, 10 mM KCl, protease inhibitor cocktail (04693159001, Roche, Indianapolis, IN), 1 mM DTT, 1 mM TSA). Following a brief centrifugation, the pellet was resuspended in hypertonic buffer (20 mM HEPES-KOH [pH 7.9], 25% glycerol, 420 mM NaCl 1.5 mM MgCl2, 0.2 mM EDTA, protease inhibitor cocktail, 1 mM DTT, 1 mM TSA). The supernatant was recovered as nuclear extract.
Western Blot Analysis
Proteins in the nuclear extracts (60 μg protein) were separated by SDS–PAGE gels (456-1043, Bio-Rad, Hercules, CA), transferred onto PVDF membranes (162-0174, Bio-Rad), and incubated with anti-histone antibodies: acetylated H3K9 (1/2000: ab4441, abcam, Cambridge, MA), acetylated H3K14 (1/5000: 06-911, EMD Millipore, Billerica, MA), acetylated H4K5 (1/2000: 07-327, EMD Millipore). Bound antibody was detected by horseradish peroxidase-conjugated anti-rabbit secondary antibody (1/20000: 1705046, Bio-Rad) and developed using Clarity™ Western ECL Substrate (1705060, Bio-Rad). Signals were detected and captured by ImageQuant™ LAS 4000 mini (GE healthcare, Pittsburgh, PA), and band intensities were quantified with ImageJ software. H3K9 acetylation intensity in individual lanes was reported relative to the normalized Mock treatment and calculated using this formula: Relative H3K9ace intensity for each timepoint = (H3K9ace / PCNA) / averaged (Mock H3K9ace / Mock PCNA). Statistical significance was determined by unpaired t test (two-tailed) against mock at each time point.
Data availability
All transcriptome data sets in this publication have been deposited in the GEO repository with accession number GSE116502. All other data is presented in the figures, tables, and supplementary materials.
Supporting information
Competing interests
A.R. is founder of startups: Sensorygen working on computational neurobiology of olfaction and taste, and Remote Epigenetics working on small molecule enhancement of agricultural production. A.R., S.T.C., S.H.Y., R.N-F., are listed as inventors on patents filed by University of California.
Acknowledgements
We would like to thank Dr. Larry Marsh for advice about the fly eye assay and HDAC inhibitors, Joel Kowalewski for help with critical comments, Dr. Thomas Girke for guidance in bioinformatics analysis, Drs. Anupama Dahanukar, Naoki Yamanaka, Chih-cheng Tsai, Karine LeRoch and Adler R. Dillman for sharing equipment and reagents. This work was partially supported by funds provided by the National Institutes of Health, NIAID (R01AI153195 to M.G.N and R01AI087785 to A.R), and HATCH funds from the Agricultural Experimentation Station UC Riverside to A.R.
References
- 1.Animals in a bacterial world, a new imperative for the life sciencesProceedings of the National Academy of Sciences 110:3229–3236http://www.pnas.org/content/3110/3229/3229Google Scholar
- 2.Finding the missing links among metabolites, microbes, and the hostImmunity 40:824–832Google Scholar
- 3.Metabolomics analysis reveals large effects of gut microflora on mammalian blood metabolitesProceedings of the National Academy of Sciences 106:3698–3703http://www.pnas.org/content/3106/3610/3698Google Scholar
- 4.Gut microbiota, metabolites and host immunityNat Rev Immunol 16:341–352Google Scholar
- 5.Diacetyl: occurrence, analysis, and toxicityJ Agric Food Chem 62:4048–4053Google Scholar
- 6.Odor exposure causes central adaptation and morphological changes in selected olfactory glomeruli in DrosophilaJournal of Neuroscience 21:6274–6282Google Scholar
- 7.Activity-dependent plasticity in an olfactory circuitNeuron 56:838–850Google Scholar
- 8.Odor exposure causes central adaptation and morphological changes in selected olfactory glomeruli in DrosophilaJ Neurosci 21:6274–6282Google Scholar
- 9.Reassessment of the Influence of Malolactic Fermentation on the Concentration of Diacetyl in WinesAm J Enol Vitic 46:385–388Google Scholar
- 10.Distilled beverages. Volatile Compounds in Foods and Beverages, ed Maarse H (Dekker IncNew York :548–580Google Scholar
- 11.Flavour Determinants of Beer Quality. Beer: Quality, Safety and Nutritional AspectsCamdridge, UK: The Royal Society od Chemistry pp. 40–73Google Scholar
- 12.Modification of CO2 avoidance behaviour in Drosophila by inhibitory odorantsNature 461:277–281Google Scholar
- 13.Functions of site-specific histone acetylation and deacetylationAnnu Rev Biochem 76:75–100Google Scholar
- 14.Histone acetylation: molecular mnemonics on the chromatinNat Rev Neurosci 14:97–111Google Scholar
- 15.Histone deacetylase inhibitors and the promise of epigenetic (and more) treatments for cancerNat Rev Cancer 6:38–51Google Scholar
- 16.Therapeutic application of histone deacetylase inhibitors for central nervous system disordersNat Rev Drug Discov 7:854–868Google Scholar
- 17.Multiple roles of HDAC inhibition in neurodegenerative conditionsTrends Neurosci 32:591–601Google Scholar
- 18.Two chemosensory receptors together mediate carbon dioxide detection in DrosophilaNature 445:86–90Google Scholar
- 19.Wiring Stability of the Adult Drosophila Olfactory Circuit after LesionJ. Neurosci 26:3367–3376Google Scholar
- 20.Activity-Dependent Changes to the Brain and Behavior of the Honey Bee, Apis mellifera (L.)J. Neurosci 17:7148–7156Google Scholar
- 21.Suppression of oxidative stress by beta-hydroxybutyrate, an endogenous histone deacetylase inhibitorScience 339:211–214Google Scholar
- 22.Breath gas metabolites and bacterial metagenomes from cystic fibrosis airways indicate active pH neutral 2,3-butanedione fermentationISME J 8:1247–1258Google Scholar
- 23.Influence of valine and other amino acids on total diacetyl and 2,3-pentanedione levels during fermentation of brewer’s wortAppl Microbiol Biotechnol 97:6919–6930Google Scholar
- 24.Chemical signaling between gut microbiota and host chromatin: What is your gut really saying?J Biol Chem 292:8582–8593Google Scholar
- 25.Microbiological and biochemical origins of human axillary odourFEMS Microbiol Ecol 83:527–540Google Scholar
- 26.Massive production of butanediol during plant infection by phytopathogenic bacteria of the genera Dickeya and PectobacteriumMolecular microbiology 82:988–997Google Scholar
- 27.Histone deacetylase inhibitors arrest polyglutamine-dependent neurodegeneration in DrosophilaNature 413:739–743Google Scholar
- 28.[HDAC inhibitors as therapy for neural disorders. Discovery of a new therapy]Pharm Unserer Zeit 39:204–209Google Scholar
- 29.Brain Uptake of Neurotherapeutics after Intranasal versus Intraperitoneal Delivery in MiceJ Neurol Neurosurg 2:1Google Scholar
- 30.Single-cell transcriptomic analyses provide insights into the developmental origins of neuroblastomaNature Genetics 53:683–693Google Scholar
- 31.The genetic landscape of high-risk neuroblastomaNature Genetics 45:279–284Google Scholar
- 32.Identification of Potential Prognostic Genes for NeuroblastomaFront Genet 9Google Scholar
- 33.Drosophila in the study of neurodegenerative diseaseNeuron 52:169–178Google Scholar
- 34.Polyglutamine-expanded human huntingtin transgenes induce degeneration of Drosophila photoreceptor neuronsNeuron 21:633–642Google Scholar
- 35.Involvement of HDAC1 and HDAC3 in the Pathology of Polyglutamine Disorders: Therapeutic Implications for Selective HDAC1/HDAC3 InhibitorsPharmaceuticals (Basel 7:634–661Google Scholar
- 36.The butter flavorant, diacetyl, forms a covalent adduct with 2-deoxyguanosine, uncoils DNA, and leads to cell deathJ Agric Food Chem 60:3311–3317Google Scholar
- 37.Diacetyl exposure as a pneumotoxic factor: a reviewRocz Panstw Zakl Hig 65:87–92Google Scholar
- 38.Respiratory toxicity of diacetyl in C57BL/6 miceToxicol Sci 103:169–180Google Scholar
- 39.Severe airway epithelial injury, aberrant repair and bronchiolitis obliterans develops after diacetyl instillation in ratsPLoS One 6:e17644Google Scholar
- 40.Life extension in Drosophila by feeding a drugProc Natl Acad Sci U S A 99:838–843Google Scholar
- 41.Emission of diacetyl (2,3 butanedione) from natural butter, microwave popcorn butter flavor powder, paste, and liquid productsInt J Occup Environ Health 16:291–302Google Scholar
- 42.Diacetyl and 2,3-pentanedione exposures associated with cigarette smoking: implications for risk assessment of food and flavoring workersCrit Rev Toxicol 44:420–435Google Scholar
- 43.A Novel Multigene Family May Encode Odorant Receptors: A Molecular Basis for Odor RecognitionCell 65:175–187Google Scholar
- 44.A novel family of divergent seven-transmembrane proteins: candidate odorant receptors in DrosophilaNeuron 22:327–338Google Scholar
- 45.Divergent seven transmembrane receptors are candidate chemosensory receptors in C. elegansCell 83:207–218Google Scholar
- 46.Variant Ionotropic Glutamate Receptors as Chemosensory Receptors in DrosophilaCell 136:149–162Google Scholar
- 47.A spatial map of olfactory receptor expression in the Drosophila antennaCell 96:725–736Google Scholar
- 48.Mapping odor valence in the brain of flies and miceCurr Opin Neurobiol 24:34–38Google Scholar
- 49.CRYPTOCHROME-mediated phototransduction by modulation of the potassium ion channel beta-subunit redox sensorProc Natl Acad Sci U S A 112:2245–2250Google Scholar
- 50.Molecular evolution of the histone deacetylase family: functional implications of phylogenetic analysisJ Mol Biol 338:17–31Google Scholar
- 51.The evolutionary history of histone H3 suggests a deep eukaryotic root of chromatin modifying mechanismsBMC Evol Biol 10:259Google Scholar
- 52.HDAC inhibitors: magic bullets, dirty drugs or just another targeted therapyCancer Lett 280:123–124Google Scholar
- 53.Potential advantages of CUDC-101, a multitargeted HDAC, EGFR, and HER2 inhibitor, in treating drug resistance and preventing cancer cell migration and invasionMol Cancer Ther 12:925–936Google Scholar
- 54.Sirtinol, a class III HDAC inhibitor, induces apoptotic and autophagic cell death in MCF-7 human breast cancer cellsInt J Oncol 41:1101–1109Google Scholar
- 55.HDAC inhibitor M344 suppresses MCF-7 breast cancer cell proliferationBiomed Pharmacother 66:232–236Google Scholar
- 56.A new synthetic HDAC inhibitor, MHY218, induces apoptosis or autophagy-related cell death in tamoxifen-resistant MCF-7 breast cancer cellsInvest New Drugs 30:1887–1898Google Scholar
- 57.Discovery of 7-(4-(3-ethynylphenylamino)-7-methoxyquinazolin-6-yloxy)-N-hydroxyheptanamide (CUDc-101) as a potent multi-acting HDAC, EGFR, and HER2 inhibitor for the treatment of cancerJ Med Chem 53:2000–2009Google Scholar
- 58.Histone deacetylase inhibitors: mechanisms and clinical significance in cancer: HDAC inhibitor-induced apoptosisAdv Exp Med Biol 615:261–298Google Scholar
- 59.The HDAC inhibitor trichostatin A inhibits growth of small cell lung cancer cellsJ Surg Res 142:219–226Google Scholar
- 60.SAHA, a HDAC inhibitor, has profound anti-growth activity against non-small cell lung cancer cellsOncol Rep 15:187–191Google Scholar
- 61.Histone deacetylases as regulators of inflammation and immunityTrends Immunol 32:335–343Google Scholar
- 62.Near-optimal probabilistic RNA-seq quantificationNat Biotechnol 34:525–527Google Scholar
- 63.Differential analyses for RNA-seq: transcript-level estimates improve gene-level inferencesF1000Res 4:1521Google Scholar
- 64.Normalization of RNA-seq data using factor analysis of control genes or samplesNat Biotechnol 32:896–902Google Scholar
- 65.edgeR: a Bioconductor package for differential expression analysis of digital gene expression dataBioinformatics 26:139–140Google Scholar
- 66.PANTHER version 11: expanded annotation data from Gene Ontology and Reactome pathways, and data analysis tool enhancementsNucleic Acids Res 45:D183–D189Google Scholar
- 67.clusterProfiler: an R package for comparing biological themes among gene clustersOMICS 16:284–287Google Scholar
- 68.A rapid micropreparation technique for extraction of DNA-binding proteins from limiting numbers of mammalian cellsNucleic Acids Res 19:2499Google Scholar
Article and author information
Author information
Version history
- Preprint posted:
- Sent for peer review:
- Reviewed Preprint version 1:
- Reviewed Preprint version 2:
- Version of Record published:
Cite all versions
You can cite all versions using the DOI https://doi.org/10.7554/eLife.86823. This DOI represents all versions, and will always resolve to the latest one.
Copyright
© 2023, Haga-Yamanaka et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.