Peer review process
Revised: This Reviewed Preprint has been revised by the authors in response to the previous round of peer review; the eLife assessment and the public reviews have been updated where necessary by the editors and peer reviewers.
Read more about eLife’s peer review process.Editors
- Reviewing EditorJalees RehmanUniversity of Illinois Chicago, Chicago, United States of America
- Senior EditorSatyajit RathNational Institute of Immunology, New Delhi, India
Reviewer #1 (Public Review):
In this revised preprint the authors investigate whether a presumably allosteric P2RX7 activating compound that they previously discovered reduces fibrosis in a bleomycin mouse model. They chose this particular model as publicly available mRNA data indicate that the P2RX7 pathway is downregulated in idiopathic pulmonary fibrosis patients compared to control individuals. In their revised manuscript, the authors use three proxies of lung damage, Ashcroft score, collagen fibers, and CD140a+ cells, to assess lung damage following the administration of bleomycin. These metrics are significantly reduced on HEI3090 treatment. Additional data implicate specific immune cell infiltrates and cytokines, namely inflammatory macrophages and damped release of IL-17A, as potential mechanistic links between their compound and reduced fibrosis. Finally, the researchers transplant splenocytes from WT, NLRP3-KO, and IL-18-KO mice into animals lacking the P2RX7 receptor to specifically ascertain how the transplanted splenocytes, which are WT for P2RX7 receptor, respond to HEI3090 (a P2RX7 agonist). Based on these results, the authors conclude that HEI3090 enhanced IL-18 production through the P2RX7-NLRP3 inflammasome axis to dampen fibrosis.
These findings could be interesting to the field, as there are conflicting results as to whether NLRP3 activation contributes to fibrosis and if so, at what stage(s) (e.g., acute damage phase versus progression). The revised manuscript is more convincing in that three orthogonal metrics for lung damage were quantified. However, major weaknesses of the study still include inconsistent and small effect sizes of HEI3090 treatment versus either batch effects from transplanted splenocytes or the effects of different genetic backgrounds. Moreover, the fundamental assumption that HEI3090 acts specifically and functionally through the P2RX7 pathway in this model cannot be directly tested, as the authors now provide results indicating that P2RX7 knockout mice do not establish lung fibrosis on bleomycin treatment.
In order to provide clear evidence that HEI3090 functions through P2RX7, a different lung fibrosis model that does not require P2RX7 would be necessary. For example, in such a system the authors could demonstrate a lack of HEI3090-mediated therapeutic effect on P2RX7 knockout. Molecularly, additional evidence on specificity, such as thermal proteome profiling and direct biophysical binding experiments, would also enhance the authors' argument that the compound indeed binds P2RX7 directly and specifically. Since all small molecules have some degree of promiscuity, the absence of an additional P2RX7 modulator, or direct recombinant IL-18 administration (as suggested by another reviewer), is needed to orthogonally validate the functional importance of this pathway. Another way the authors could probe pathway specificity would involve co-administering α-IL-18 with HEI3090 in several key experiments (similar to Figure 4L).
Reviewer #2 (Public Review):
In the study by Hreich et al, the potency of P2RX7-specific positive modulator HEI3090, developed by the authors, for the treatment of Idiopathic pulmonary fibrosis (IPF) was investigated. Recently, the authors have shown that HEI3090 can protect against lung cancer by stimulating dendritic cell P2RX7, resulting in IL-18 production that stimulates IFN-γ production by T and NK cells (DOI: 10.1038/s41467-021-20912-2). Interestingly, HEI3090 increases IL-18 levels only in the presence of high eATP. Since the treatment options for IPF are limited, new therapeutic strategies and targets are needed. The authors first show that P2RX7/IL-18/IFNG axis is downregulated in patients with IPF. Next, they used a bleomycin-induced lung fibrosis mouse model to show that the use of a positive modulator of P2RX7 leads to the activation of the P2RX7/IL-18 axis in immune cells that limits lung fibrosis onset or progression. Mechanistically, treatment with HEI3090 enhanced IL-18-dependent IFN-γ production by lung T cells leading to a decreased production of IL-17 and TGFβ, major drivers of IPF. The major novelty is the use of the small molecule HEI3090 to stimulate the immune system to limit lung fibrosis progression by targeting the P2RX7, which could be potentially combined with current therapies available. Overall, the study was well performed and the manuscript is clear. However, there is need for more details on the description and interpretation of the adoptive transfer experiments, as well as the statistical analyses and number of replicate independent experiments.
Author Response
The following is the authors’ response to the original reviews.
eLife assessment
This study presents a potentially valuable discovery which indicates that activation of the P2RX7 pathway can reduce the lung fibrosis after its establishment by inflammatory damage. If confirmed, the study could clarify the role of specific immune networks in the establishment and progression of lung fibrosis. However, the presented data and analyses are incomplete as they primarily rely on limited pharmacological treatments with modest effect sizes. I hope you will be convinced by the validity of our approaches with the following explanation/information and I remain at your disposal to discuss
Public Reviews:
Reviewer #1 (Public Review):
In this revised preprint the authors investigate whether a presumably allosteric P2RX7 activating compound that they previously discovered reduces fibrosis in a bleomycin mouse model. They chose this particular model as publicly available mRNA data indicate that the P2RX7 pathway is downregulated in idiopathic pulmonary fibrosis patients compared to control individuals. In their revised manuscript, the authors use three proxies of lung damage, Ashcroft score, collagen fibers, and CD140a+ cells, to assess lung damage following the administration of bleomycin. These metrics are significantly reduced on HEI3090 treatment. Additional data implicate specific immune cell infiltrates and cytokines, namely inflammatory macrophages and damped release of IL-17A, as potential mechanistic links between their compound and reduced fibrosis. Finally, the researchers transplant splenocytes from WT, NLRP3-KO, and IL-18-KO mice into animals lacking the P2RX7 receptor to specifically ascertain how the transplanted splenocytes, which are WT for P2RX7 receptor, respond to HEI3090 (a P2RX7 agonist). Based on these results, the authors conclude that HEI3090 enhanced IL-18 production through the P2RX7-NLRP3 inflammasome axis to dampen fibrosis.
These findings could be interesting to the field, as there are conflicting results as to whether NLRP3 activation contributes to fibrosis and if so, at what stage(s) (e.g., acute damage phase versus progression). The revised manuscript is more convincing in that three orthogonal metrics for lung damage were quantified. However, major weaknesses of the study still include inconsistent and small effect sizes of HEI3090 treatment versus either batch effects from transplanted splenocytes or the effects of different genetic backgrounds. Moreover, the fundamental assumption that HEI3090 acts specifically and functionally through the P2RX7 pathway in this model cannot be directly tested, as the authors now provide results indicating that P2RX7 knockout mice do not establish lung fibrosis on bleomycin treatment.
I’m particularly concerned by the assumption made by reviewer 1 concerning the fact that P2RX7 knockout mice do not establish lung fibrosis on bleomycin treatment.
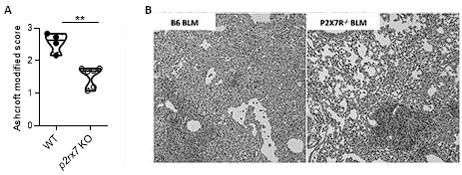
Indeed, what we showed in the point-to-point response is that BLM induces fibrosis in both WT and P2RX7 KO mice, but the intensity of the fibrosis is reduced in P2RX7KO mice, panel A. Therefore, as discussed in our first response, our results confirmed the previous publication of Riteau et al, that P2RX7 participates in BLM-induced lung fibrosis (see panel B).
Author response image 1.
Bleomycin induced lung fibrosis in WT versus p2rx7 KO mice. A: lung from BLM-treated mice were stained with HE and fibrosis was quantified using the Ashcroft protocol. Result showed that fibrosis induced by BLM in KO mice is reduced as compared to WT mice. B: Representative images of lung sections at day 14 after BLM treatment stained with H&E as published in Riteau et al. and illustrating that fibrosis induced by BLM in KO mice is reduced as compared to WT mice. WT mice vehicle (n=4) or p2rx7 KO (n=6) mice. Two-tailed Mann-Whitney test, p values: **p < 0.01.
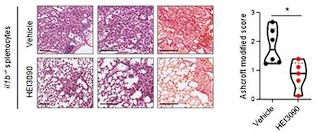
Importantly, this lower intensity of lung fibrosis in P2RX7 KO mice, does not interfere with the capacity of our molecule to attenuate lung fibrosis, as demonstrated in the adoptive transfer of IL1B KO splenocytes in P2RX7 KO mice, in which HEI3090 decreases the Ashcroft score, the % of fibrosis and the collagen fibers (see below).
Author response image 2.
HEI3090 activity requires P2RX7’s expressing immune cells: Experimental design. p2rx7-/- mice were given 3.106 il1β-/- splenocytes i.v. one day prior to BLM delivery (i.n. 2.5 U/kg). Mice were treated daily i.p. with 1.5 mg/kg HEI3090 or vehicle for 14 days. (C) Representative images of lung sections at day 14 after treatment stained with H&E and Sirius Red with il1β-/- splenocytes, bar= 100 µm (left) and fibrosis score assessed by the Ashcroft method, the % of fibrosis and the content of collagen fibers (right). Each point represents one mouse (n=2 in WT and NLRP3 experiment, n =1 in IL18 and IL1B experiment), data represented as violin plot or mean±SEM, two-tailed Mann-Whitney test, *p < 0.05. WT: Wildtype, KO: P2RX7 knock-out
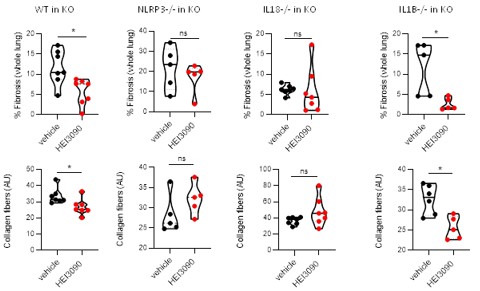
Importantly, in the same experimental setting, e.g adoptive transfer of splenocytes from different genetic backgrounds, HEI3090 decreases the fibrosis intensity only with WT and IL1B KO splenocytes and not with NLRP3 KO and IL18KO splenocytes.
Author response image 3.
HEI3090 activity requires P2RX7’s expressing immune cells: Experimental design. p2rx7-/- mice were given 3.106 WT, NLRP3-/-, IL18-/- or IL1β-/- splenocytes i.v. one day prior to BLM delivery (i.n. 2.5 U/kg). Mice were treated daily i.p. with 1.5 mg/kg HEI3090 or vehicle for 14 days. Fibrosis in whole lung was assessed by the % of fibrosis (upper panel) and the content of collagen fibers (lower panel). Each point represents one mouse (n=2 in WT and NLRP3 experiments, n =1 in IL18 and IL1B experiment). Data represented as violin plot or mean±SEM, two-tailed Mann-Whitney test, *p < 0.05. WT: Wildtype, KO: P2RX7 knock-out
In order to provide clear evidence that HEI3090 functions through P2RX7, a different lung fibrosis model that does not require P2RX7 would be necessary. For example, in such a system the authors could demonstrate a lack of HEI3090-mediated therapeutic effect on P2RX7 knockout.
Since BLM induces lung fibrosis in P2RX7 KO mice as we showed in this manuscript and as already published by Riteau in 2010, shown earlier in our response (first figure) and because HEI3090 is able to decrease the intensity of fibrosis in WT and IL1B-/- → P2RX7 KO mice but not in KO, NLRP3-/- → P2RX7 KO and IL18-/- → P2RX7 KO mice we believe that our data sustain the conclusion that
-
HEI3090 required the expression of P2RX7 in immune cells to mediate the antifibrotic activity,
-
IL1B is not a crucial effector mediating the antifibrotic effect of HEI3090.
Molecularly, additional evidence on specificity, such as thermal proteome profiling and direct biophysical binding experiments, would also enhance the authors' argument that the compound indeed binds P2RX7 directly and specifically. Since all small molecules have some degree of promiscuity, the absence of an additional P2RX7 modulator, or direct recombinant IL-18 administration (as suggested by another reviewer), is needed to orthogonally validate the functional importance of this pathway. Another way the authors could probe pathway specificity would involve co-administering α-IL-18 with HEI3090 in several key experiments (similar to Figure 4L).
At the moment we have no funds to do these experiments and given the high competition, we have decided to publish our story without these new data.
Reviewer #2 (Public Review):
In the study by Hreich et al, the potency of P2RX7-specific positive modulator HEI3090, developed by the authors, for the treatment of Idiopathic pulmonary fibrosis (IPF) was investigated. Recently, the authors have shown that HEI3090 can protect against lung cancer by stimulating dendritic cell P2RX7, resulting in IL-18 production that stimulates IFN-γ production by T and NK cells (DOI: 10.1038/s41467-021-20912-2). Interestingly, HEI3090 increases IL-18 levels only in the presence of high eATP. Since the treatment options for IPF are limited, new therapeutic strategies and targets are needed. The authors first show that P2RX7/IL-18/IFNG axis is downregulated in patients with IPF. Next, they used a bleomycin-induced lung fibrosis mouse model to show that the use of a positive modulator of P2RX7 leads to the activation of the P2RX7/IL-18 axis in immune cells that limits lung fibrosis onset or progression. Mechanistically, treatment with HEI3090 enhanced IL-18-dependent IFN-γ production by lung T cells leading to a decreased production of IL-17 and TGFβ, major drivers of IPF. The major novelty is the use of the small molecule HEI3090 to stimulate the immune system to limit lung fibrosis progression by targeting the P2RX7, which could be potentially combined with current therapies available. Overall, the study was well performed, and the manuscript is clear.
We thank the reviewer for this very positive comments.
However, there is need for more details on the description and interpretation of the adoptive transfer experiments, as well as the statistical analyses and number of replicate independent experiments.
I’m concerned by the reviewer’s comments, and I would like to bring additional information/explanation, which I hope will convince you on the validity of our approaches.
Author response image 4.
Adoptive transfer experiment. Adoptive transfer experiments are classically used to document which immune cells participate in immune cell responses (with more than 150 publications in pubmed with the key words adoptive transfer and onco immunology) and intravenous administration is a common route to trigger lungs (PMID: 23336716). To characterize the molecular effector (P2RX7, NLRP3, IL18 and IL1B) accounting for the antifibrotic effect of HEI3090 we purified splenocytes from donor mice and administrated them intra venously in P2RX7 KO mice. As shown in Author response image 4, HEI3090 has no antifibrotic activity when splenocyte isolated from mice invalidated for p2rx7 are iv into P2RX7 KO mice (KO in KO). By contrast, HEI3090 has antifibrotic activity when WT splenocytes expressing P2RX7 (isolated from WT mice) are transferred into P2RX7 KO mice (WT in KO).
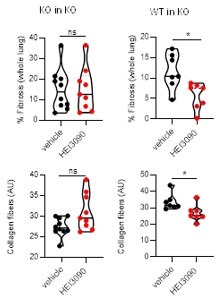
This experiment brings strong evidence to demonstrate the efficacy of adoptive transfer approach to identify molecular effector required to mediate the antifibrotic effect of HEI3090.
Statistical analyses and number of replicate independent experiments
We thank the reviewer for his comment, and we apologize to not have been sufficiently clear in our previous response with this miss phrased statement “the experiment was stopped when significantly statistical results were observed” when we should have written “the experiment was stopped when each experimental group contained at least 5 mice”.
To define the size of experimental groups we did a pilot experiment, with 4 WT mice (e.g. 4 biological replicates) in each group (as shown aside), and a statistical forecasting based on the result of the pilot experiment (40% difference, standard error: 0.9, α risk: 0.05, power: 0.8). Since we focused on the effect of HEI3090 we based our statistical analysis on a one-way ANOVA analysis comparing in each experiment the vehicle and the treated group.
The pilot experiment and statistical forecasting indicated 4 mice per group to characterize the effect of HEI3090 on BLM-induced lung fibrosis. Each experiment was started with 6 to 8 mice per group. Being aware that 30% of mice can unexpectedly dye due to BLM treatment, we duplicated the experiment, when necessary, to include at least 5 mice in each group of each experiment meaning 5 biological replicates, knowing that 4 mice are sufficient to statistically analyze the results. In each experiment we have checked for the presence of outlier, using the ROULT method, and removed the outliers when necessary.



