Peer review process
Revised: This Reviewed Preprint has been revised by the authors in response to the previous round of peer review; the eLife assessment and the public reviews have been updated where necessary by the editors and peer reviewers.
Read more about eLife’s peer review process.Editors
- Reviewing EditorZiyue GaoUniversity of Pennsylvania, Philadelphia, United States of America
- Senior EditorMolly PrzeworskiColumbia University, New York, United States of America
Reviewer #1 (Public Review):
The revised manuscript new presented 1) a permutation-based test for the significance of the overlap between DEGs and genes with positive selection signals in Tibetans, and 2) polygenic adaptation test for the eQTLs. I make my suggestions in detail as below:
Major Comments
1. My previous concern regarding the DEG analysis remains unresolved. Although the authors agreed in their response that the difference between the male- and female-specific DEGs are insufficient to the difference between sex-combined and sex-specific DEGs (Figure S6). However, the results section still states the opposite pattern between males and females as a decisive reason for the difference (p. 9, lines 236-239). Again, I would like to recommend the authors to test alternative ways of analysis to boost statistical power for DEG detection other than simply splitting data into males and females and performing analysis in each subset. For example, the authors may consider utilizing gene by environment interaction analysis schemes here biological sex as an environmental factor.
2. Multiple testing schemes are still sub-optimal in some cases. Most of all, the p-values in the WGCNA analysis (p. 11), the authors corrected for the number of traits (n=12) after adjusting for the correlation between them. However, they did not mention whether they counted for the number of modules they tested at all (n=136 and 161 for males and females, respectively). Whether they account for the number of modules will make a substantial difference in the significance threshold, please incorporate and describe a proper multiple testing scheme for this analysis.
3. Evidence for natural selection on the observed DEG pattern is still weak and not properly described.
1) For the overlap between DEGs and TSNGs, the authors introduced a permutation-based test, but used a total set of genes in the human genome as a comparison set (p. 25, lines 699-700). I believe that the authors should sample random sets of genes from those already expressed in each tissue to make a fair comparison.
2) The entire polygraph analysis for polygenic adaptation is poorly described. The current version of the Methods does not clarify i) for which genes the eQTLs are discovered, 2) how the authors performed the eQTL analysis, iii) how the authors polarized the effect, and iv) how they set up a comparison between the eQTLs and the others.
Reviewer #2 (Public Review):
More than 80 million people live at high altitude. This impacts health outcomes, including those related to pregnancy. Longer-lived populations at high altitudes, such as the Tibetan and Andean populations show partial protection against the negative health effects of high altitude. The paper by Yue sought to determine the mechanisms by which the placenta of Tibetans may have adapted to minimise the negative effect of high altitude on fetal growth outcomes. It compared placentas from pregnancies from Tibetans to those from the Han Chinese. It employed RNAseq profiling of different regions of the placenta and fetal membranes, with some follow-up of histological changes in umbilical cord structure and placental structure. The study also explored the contribution of fetal sex in these phenotypic outcomes.
A key strength of the study is the large sample sizes for the RNAseq analysis, the analysis of different parts of the placenta and fetal membranes, and the assessment of fetal sex differences.
A main weakness is that this study, and its conclusions, largely rely on transcriptomic changes informed by RNAseq. Changes in genes and pathways identified through bioinformatic analysis were not verified by alternate methods, such as by western blotting, which would add weight to the strength of the data and its interpretations. There is also a lack of description of patient characteristics, so the reader is unable to make their own judgments on how placental changes may link to pregnancy outcomes. Another weakness is that the histological analyses were performed on n=5 per group and were rudimentary in nature.
Author Response
The following is the authors’ response to the original reviews.
Reviewer #1:
In this manuscript, the authors investigate differences between Tibetans and Han Chinese at altitude in terms of placental transcriptomes during full-term pregnancy. Most importantly, they found that the inter-population differentiation is mostly male-specific and the observed direction of transcriptional differentiation seems to be adaptive at high altitude. In general, it is of great importance and provides new insights into the functional basis of Tibetan high-altitude adaptations, which so far have been mostly studied via population genetic measures only. More specifically, I firmly believe that we need more phenotype data (including molecular phenotypes such as gene expression data) to fully understand Tibetan adaptations to high altitude, and this manuscript is a rare example of such a study. I have a few suggestions and/or questions with which I hope to improve the manuscript further, especially in terms of 1) testing if the observed DEG patterns are truly adaptive, and 2) how and whether the findings in this study can be linked to EPAS1 and EGLN1, the signature adaptation genes in Tibetans.
We appreciate the reviewer’s constructive comments. We have addressed these points and the details are discussed below.
Major Comments:
1) The DEG analysis is the most central result in this manuscript, but the discrepancy between sex-combined and sex-specific DEGs is quite mind-boggling. For those that were differentially expressed in the sex-specific sets but not in the sex-combined one, the authors suggest an opposite direction of DE as an explanation (page 11, Figure S5). But Figure S5A does not show such a trend, showing that down-regulated genes in males are mostly not at all differentially expressed in females. Figure S5B does show such a trend, but it doesn't seem to be a dominant explanation. I would like to recommend the authors test alternative ways of analysis to boost statistical power for DEG detection other than simply splitting data into males and females and performing analysis in each subset. For example, the authors may consider utilizing gene-by-environment interaction analysis schemes here biological sex as an environmental factor.
We agree with reviewer that the opposite direction of DEGs is likely only one of the possible explanations for the discrepancy between the sex-combined and the sex-specific DEGs. We have toned down the description of this point in the revised manuscripts.
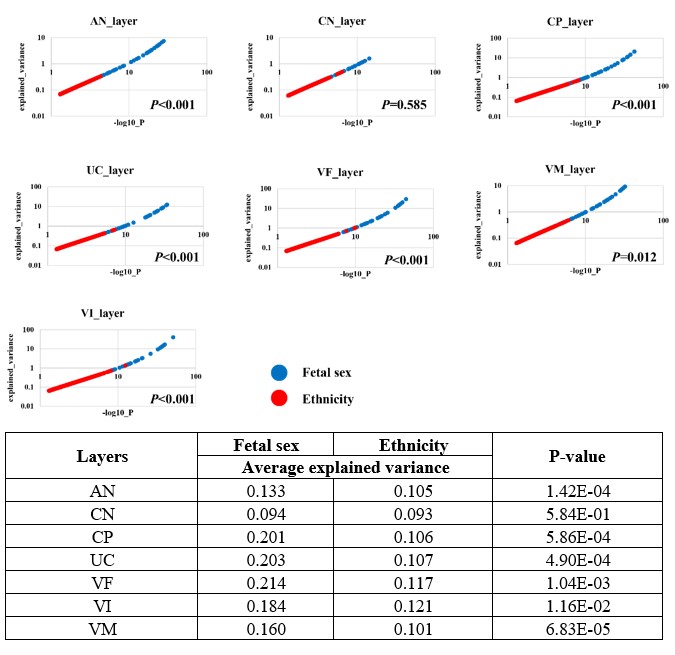
Following the suggestion of reviewer, we performed a ANCOVA analysis to evaluate the variance explained by sex from the expression data. For each gene, univariate comparisons of the average of gene expression between Tibetans and Han Chinese were made by using the ANCOVA test in R aov function with sex as covariates: aov (Expression ~ Ethnicity + Fetal sex). We observed a significantly higher variance explained by sex than by ethnicity in six layers of the placenta (except for the CN layer) (Author response image 1). For example, in the UC layer, fetal sex can explain ~0.203 variance, while the ethnicity explains ~0.107 variance (P-value = 4.9e-4). These results suggest a significant contribution of fetal sex for the observed variance of gene expression, consist with the observed sex-biased DEG patterns.
Author response image 1.
The ANCOVA results of the seven layers of placenta. The scatter plot shows the comparison of the explained variance (y-axis) and significance (x-axis, denoted by –log10(P-value)) between ethnicity (dots in red) and fetal sex (dots in blue). Each dot represents an investigated gene, and only genes with P<0.05 in significance are shown in the plots. The table is the summary statistics of the ANCOVA analysis.
2) Please clarify how the authors handled multiple testing correction of p-values.
There were three analyses involving multiple testing in this study: 1) for the differential expression analysis, we obtained the multiple corrected p-values by Benjamini-Hochberg FDR (false discovery rate) procedure; 2) for the GO enrichment analysis, we calculated the FDR-adjusted q-values from the overall p-values to correct for multiple testing.
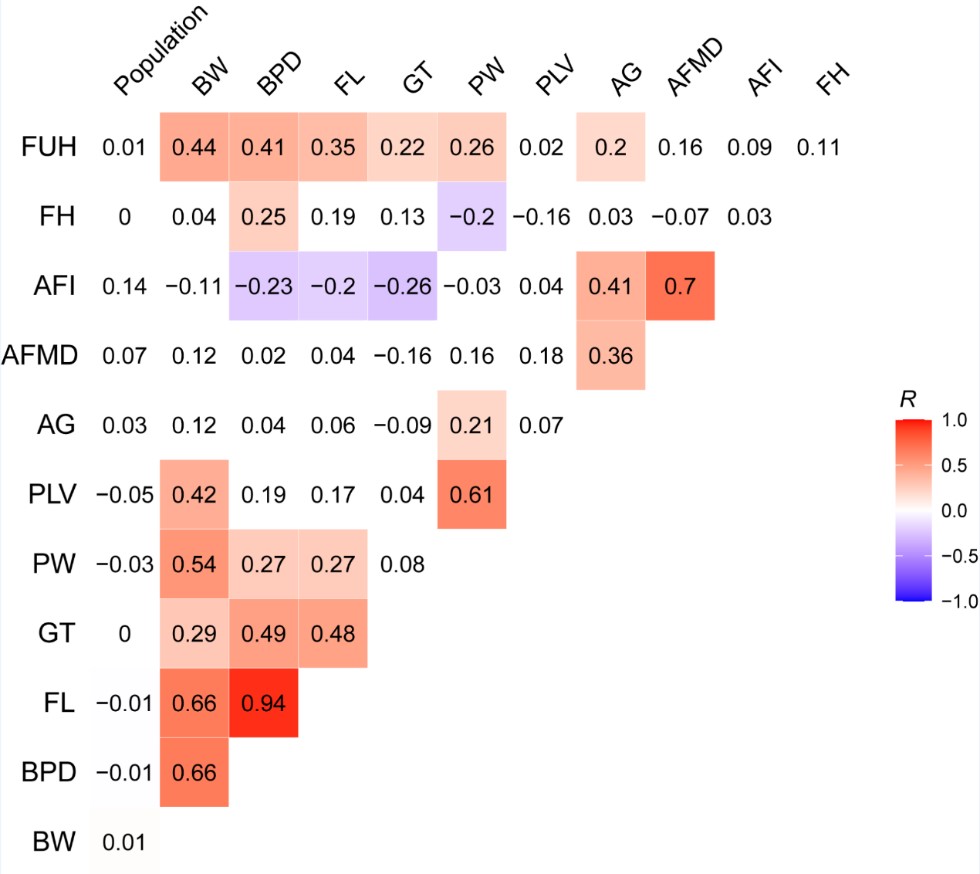
3) for the WGCNA analysis, considering the 12 traits were involved, including population, birth weight (BW), biparietal diameter (BPD), femur length (FL), gestation time (GT), placental weight (PW), placental volume (PLV), abdominal girth (AG), amniotic fluid maximcon depth (AFMD), amniotic fluid (AFI), fetal heart rate (FH) and fundal height (FUH). We calculated a Bonferroni threshold (p-value = 0.05/the number of independent traits) using the correlation matrix of the traits to evaluate the significant modules. We estimated the number of independent traits among the 12 investigated traits was 4 (Author response image 2). Therefore, we used a more stringent significant threshold p-value = 0.0125 (0.05/4) as the final threshold to correct the multiple testing brought by multiple traits in our WGCNA analyses. We have updated this section based on the new threshold.
Author response image 2.
The correlation matrix of 12 traits involved in the WGCNA analysis. The correlation coefficients larger than 0.2 (or smaller than -0.2) are regarded as significant correlation and marked in gradient colors.
3) The "natural selection acts on the placental DEGs ..." section is potentially misleading readers to assume that the manuscript reports evidence for positive selection on the observed DEG pattern between Tibetans and Han, which is not.
a) Currently the section simply describes an overlap between DEGs and a set of 192 genes likely under positive selection in Tibetans (TSNGs). The overlap is quite small, leading to only 13 genes in total (Figure 6). The authors are currently not providing any statistical measure of whether this overlap is significantly enriched or at the level expected for random sampling.
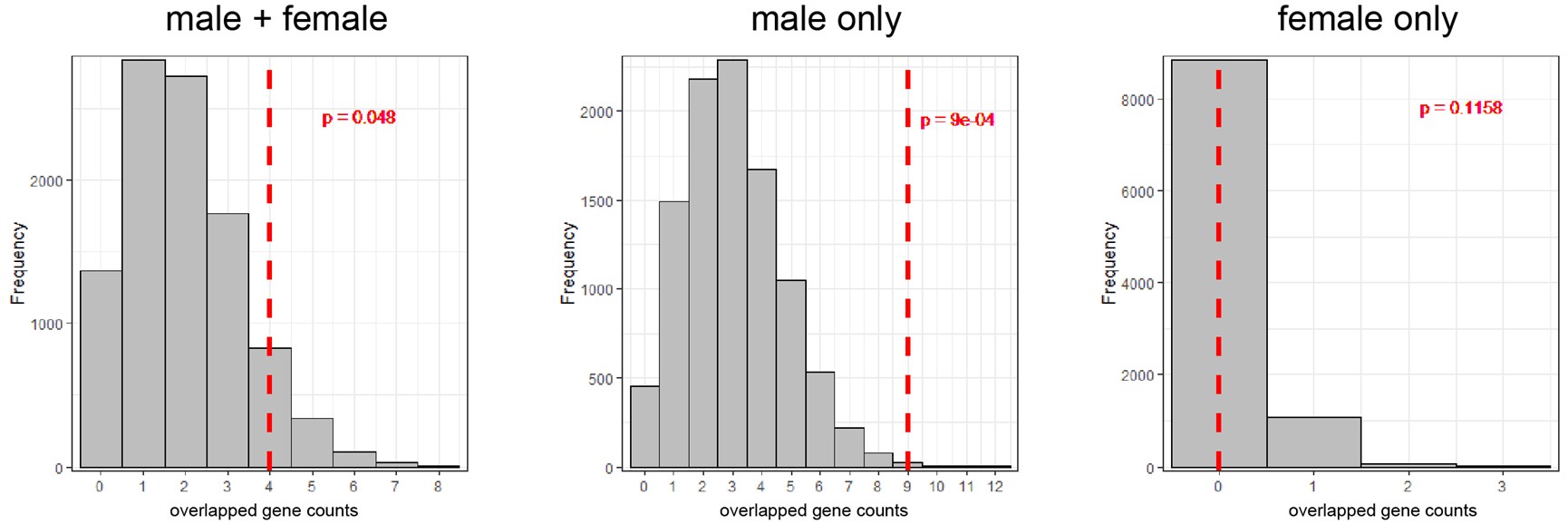
We understand the reviewer’s point that the observed gene counts overlapped between DEGs from the three sets (4 for female + male; 9 for male only and 0 for female only) with TSNGs should be tested using a statistical method. Therefore, we adopted permutation approach to evaluate the enrichment of the overlapped DEGs with TSNGs.
For each permutation, we randomly extracted 192 genes from the human genome, then overlapped with DEGs of the three sets (female + male; female only and male only) and counted the gene numbers. After 10,000 permutations, we constructed a null distribution for each set, and found that the overlaps between DEGs and TSNGs were significantly enriched in the “female + male” set (p-value = 0.048) and the “male only” set (p-value = 9e-4), but not in the “female only” set (p-value = 0.1158) (Author response image 3). This result suggests that the observed DEGs are significantly enriched in TSNGs when compared to random sampling, especially for the male DEGs. We added this analysis in the revised manuscript.
Author response image 3.
The distribution of 10,000 permutation tests of counts of the overlapped genes between DEGs and the 192 randomly selected genes in the genome. The red-dashed lines indicate the observed values based on the 192 TSNGs.
b) The authors are describing sets of DEGs that seem to affect important phenotypic changes in a consistent and adaptive direction. A relevant form of natural selection for this situation may be polygenic adaptation while the authors only consider strong positive selection at a single variant/gene level.
We agree with reviewer that polygenic adaptation might be a potential mechanism for DEGs to take effect on the adaptive phenotypes. Therefore, following the suggestion in the comment below, we conducted a polygenic adaptation analysis using eQTL information.
c) The manuscript is currently providing no eQTL information that can explain the differential expression of key genes. The authors can actually do this based on the genotype and expression data of the individuals in this study. Combining eQTL info, they can set up a test for polygenic adaptation (e.g., Berg and Coop; https://journals.plos.org/plosgenetics/article?id=10.1371/journal.pgen.1004412). This will provide a powerful and direct test for the adaptiveness of the observed DEG pattern.
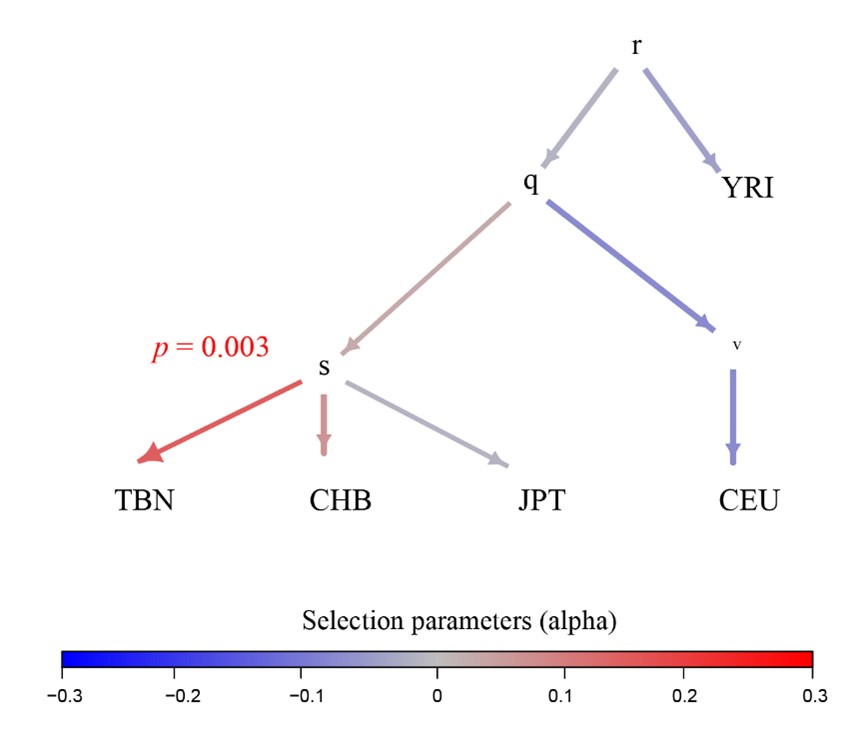
Following the reviewer’s suggestion, we employed the PolyGraph (Racimo et al., 2018) tool to identify the signatures of polygenic selection in Tibetans using eQTL information. We conducted eQTL analysis for the seven layers, and collected a set of 5,251 eQTLs, covering the SNPs associated with gene expression with a significanct p-value < 5e-8. To obtain a list of independent eQTLs, we removed those SNPs in linkage disequilibrium (r2 > 0.2 in 1000 Genome Project). Finally, we obtained 176 independent eQTLs. At the same time, we generated a set of 1,308,436 independent SNPs of Tibetans as the control panel. The PolyGraph result showed that Tibetans have a clear signature of polygenic selection on gene expression (Bonferroni-correction p-value = 0.003) (Author response image 4).
We have added this result in the revised manuscript (Figure S4), and added a detailed description of polygenic adaption in the Methods section.
Author response image 4.
Polygraphs for the eQTLs that show evidence for polygenic adaptation in the five-leaf tree built using the allele frequency data of 1001 Tibetans (Zheng et al. 2023) and 1000 Genome Project. The colors indicate the marginal posterior mean estimate of the selection parameter for variants associated with the gene expression. r, q, s and v in the tree nodes refer to the nodes in terminal branches and internal branches. TBN, Tibetans; CHB, Han Chinese in Beijing; JPT, Japanese in Tokyo, Japan; CEU, Northern Europeans from Utah; YRI, Yoruba in Ibadan, Nigeria.
4) The manuscript is currently only minimally discussing how findings are linked to EPAS1 and EGLN1 genes, which show the hallmark signature of positive selection in Tibetans. In fact, the authors' group previously reported male-specific association between EPAS1 SNPs and blood hemoglobin level. Many readers will be intrigued to see a discussion about this point.
According to the reviewer’s suggestion, in the revised manuscript, we added a paragraph to discuss the relationship between our transcriptomic data and the two genes with strong selective signals, i.e. EPAS1 and EGLN1.
“As the gene with the strongest signal of natural selection in Tibetans, EPAS1 has been reported in numerus studies on its contribution to high altitude adaptation. In this study, we detected a significant expression reduction of EPAS1 in the Tibetan UC compared to the high-altitude Han. It was reported that the selected-for EPAS1 variants/haplotype were associated with lower hemoglobin levels in the Tibetan highlanders with a major effect (Beall et al., 2010; Peng et al., 2017), and the low hemoglobin concentration of Tibetans is causally associated with a better reproductive success (Cho et al., 2017). Therefore, we speculate that the selective pressure on EPAS1 is likely through its effect on hemoglobin, rather than directly on the reproductive traits. The down-regulation of EPAS1 in placentas likely reflects a blunted hypoxic response that may improve vasodilation of UC for better blood flow, and eventually leading to the higher BW in Tibetans (He et al., 2023). For EGLN1, another well-known gene in Tibetans, we detected between-population expression difference in the male UC layer, but not in other placental layers. Considering the known adaptation mechanism of EGLN1 is attributed to the two Tibetan-enriched missense mutations, the contribution of EGLN1 to the gene expression changes in the Tibetan UC is unexpected and worth to be explored in the future.”
Reviewer #2 (Public Review):
In this manuscript, the authors use newly-generated, large-scale transcriptomic data along with histological data to attempt to dissect the mechanisms by which individuals with Tibetan ancestry are able to mitigate the negative effects of high elevation on birth weight. They present detailed analyses of the transcriptomic data and find significant sex differences in the placenta transcriptome.
I have significant concerns about the conclusions that are presented. The analyses also lack the information necessary to evaluate their reliability.
The experimental design does not include a low elevation comparison and thus cannot be used to answer questions about how ancestry influences hypoxia responses and thus birthweight at high elevations. Importantly, because the placenta tissues (and trophoblasts specifically) are quickly evolving, there are a priori good reasons to expect to find population differences irrespective of adaptive evolution that might contribute to fetal growth protection. There are also significant details missing in the analyses that are necessary to substantiate and replicate the analyses presented.
Although the datasets are ultimately valuable as reference sets, the absence of low elevation comparisons for Tibetans and Han Chinese individuals undermines the ability of the authors to assess whether differences observed between populations are linked to hypoxia responses or variation in the outcomes of interest (i.e., hypoxia-dependent fetal growth restriction).
We understand the reviewer’s concern about the lack of low-altitude comparison. For the placenta transcriptomic data, actually, we previously studied the comparison of placenta from high-altitude Tibetans and low-altitude Han Chinese, including 63 placentas of Tibetans living at Lhasa (elevation: 3650m) and 14 placentas of Han in Kunming (elevation: 1800m) (Peng et al. 2017). The main finding was that in general, the expression profiles are similar between the high-altitude Tibetans and the low-altitude Han. In particular, most high-altitude Tibetans have a similar level of EPAS1 expression in the placenta as the lowlander Han Chinese, a reflection of Tibetans’ adaptation at altitude. In other words, (Peng et al. 2017). In this study, we observed a significant down-regulation of EPAS1 in the Tibetan UC when compared to Han Chinese living at the same high altitude. Therefore, the observed differences between Tibetans and Han Chinese placenta at high altitude are due to the adaptation of Tibetans.
For phenotypic data, we made a systematical comparison of reproductive outcomes in our previous studies (He et al., 2023; He et al., 2022). We proved that polygenic adaptation of reproduction in Tibetans tends to reduce the chance of preterm birth and eliminate the restriction on fetal development at high altitude. Compared to the high-altitude Han Chinese migrants, the high-altitude Tibetans exhibit a less birth weight reduction and infant mortality induced by hypoxia, similar with the lowland Han Chinese as reference.
In summary, although we cannot make combination analysis with our high-altitude data and the published low-altitude data because of batch effect and difference of sampling strategy, we obtained more supportive evidence for the adaptation of placenta expression regulation in Tibetans. To be objective, we have discussed the limitation of the lack of lowlander placenta data in the Discussion section.
The authors attempt to tackle this phenotypic association by looking for correlations between gene networks (WGCNA) and individual genes with birthweight and other measurements collected at birth. I have some reservations about this approach with only two groups (i.e., missing the lowland comparison), but it is further problematic that the authors do not present data demonstrating that there are differences in birthweight or any other traits between the populations in the samples they collected.
Throughout, I thus find conclusions about the adaptive value and hypoxia-responses made by the authors to be unsubstantiated and/or the data to be inadequate. There are also a gratuitous number of speculative statements about mechanisms by which differential gene expression leads to the protection of birthweight that are not evaluated and thus cannot be substantiated by the data presented.
As currently presented and discussed, these results thus can only be used to evaluate population differences and tissue-specific variation therein.
We understand the reviewer’s point that the observed differences of gene expression between Tibetan natives and Han immigrants living at high altitude might be explained by ancestral divergence, rather than hypoxia-associated response and genetic adaptation of native Tibetans.
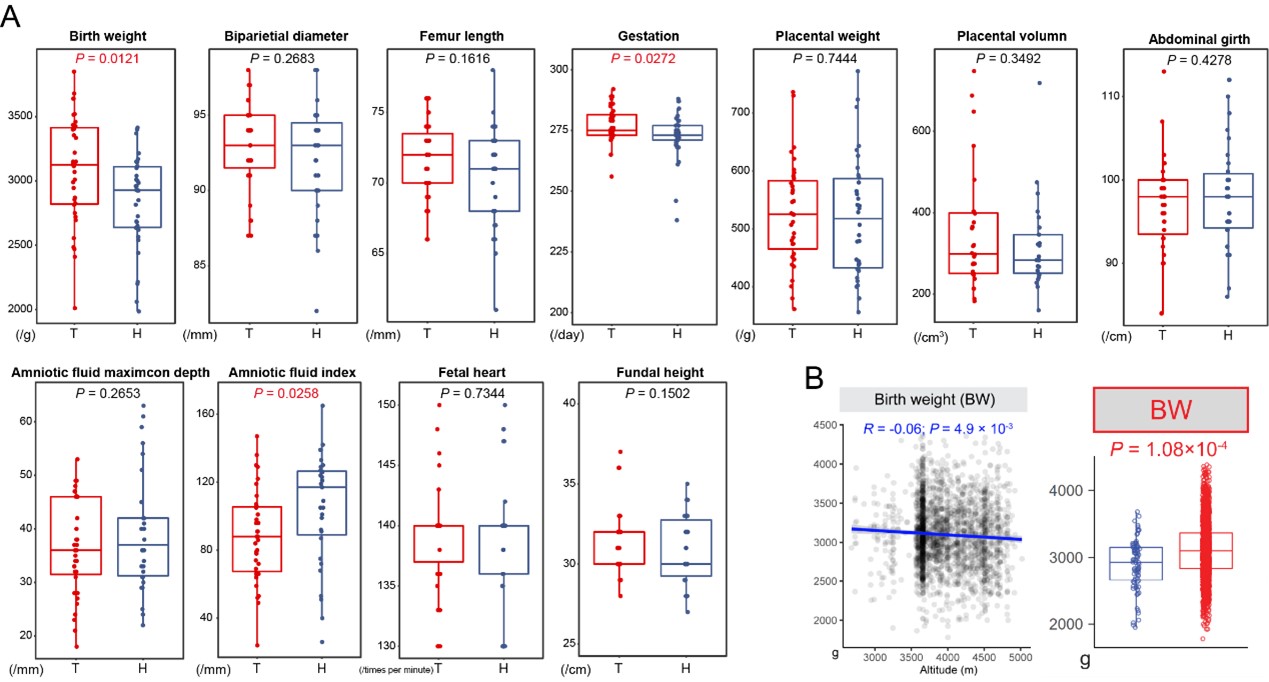
Firstly, we conclude that Tibetans have a better reproductive outcome, not only based on the two highlander groups living at the same altitude, but also relied on the change direction compared to the lowland level. For example, we observed a significant higher BW in Tibetans than Han migrants in our dataset (35 Tibetans vs. 34 Han: p-value = 0.012) (Author response image 5), and in a larger dataset (He et al. 2023) (1,317 Tibetans vs. 87 Han: p-value = 1.1e-6), suggesting an adaptation of Tibetans because BW decreases with the increase of altitude. The logic was the same to the other traits. Following the suggestion of reviewer, we added these phenotype comparisons in the revised manuscripts. The detailed information of the investigated samples and the statistic results were also added as supplementary tables in the revised version.
For the WGCNA, we agree with the reviewer that the detected modules both showing significant correlation with population and other reproductive traits cannot be fully explained by adaptation of Tibetans. Therefore, we tuned down the description of this section and added other possible explanations, such as population differences, in the discussion.
Author response image 5.
Comparison of 11 reproductive traits between Tibetans and Han immigrants. (A) comparison based on the dataset of this study (35 Tibetans vs. 34 Han); (B) correlation between BW and altitude (left panel) and comparison analysis based on the larger sample size (the data were retrieved from (He et al., 2023)). Univariate comparisons of the average of each trait cross population were made by using the ANCOVA test in R aov function with fetal sex and maternal age as covariates.
There is also some important methodological information missing that makes it difficult or impossible to assess the quality of the underlying data and/or reproduce the analyses, further limiting the potential impact of these data:
1) Transcriptome data processing and analyses: RNA quality information is not mentioned (i.e., RIN). What # of reads are mapped to annotated regions? How many genes were expressed in each tissue (important for contextualizing the # of DE genes reported - are these a significant proportion of expressed genes or just a small subset?).
According to the reviewer’s suggestion, we added more information about transcriptome data processing and analyses in the revised Methods and Results:
“After RNA extraction, we assessed the RNA integrity and purity using 1% agarose gel electrophoresis. The RIN value of extracted RNA was 7.56 ± 0.71.”
“In total, 10.6 billion reads were mapped to the annotated regions, and 17,283 genes express in all the investigated placenta.”
“We identified 579 differentially expressed genes (DEGs) between Tibetans and Han, accounting for 3.4% of the total number of expressed genes.”
2) The methods suggest that DE analyses were run using data that were normalized prior to reading them into DESeq2. DESeq2 has an internal normalization process and should not be used on data that was already normalized. Please clarify how and when normalization was performed.
Actually, we made raw read count matrix as input file when conducting differential analysis using DESeq2, rather than using the normalized data. We have updated our description in the method section of the revised manuscript.
3) For enrichment analyses, the background gene set (all expressed genes? all genes in the genome? or only genes expressed in the tissue of interest?) has deterministic effects on the outcomes. The background sets are not specified for any analyses.
Actually, we utilized the genes expressed in placenta as the background gene set for enrichment analyses. The genes with more than two transcripts per million transcripts (TPM) were regarded as an expressed gene, which is commonly used criteria for RNA-seq data.
4) In the WGCNA analysis, P-values for correlations of modules with phenotype data (birthweight etc.) should be corrected for multiple testing (i.e., running the module correlation for each outcome variables) and p.adjust used to evaluate associations to limit false positives given the large number of correlations being run.
As we explained in response to comment#2 of Reviwer-1, we used a more stringent significant threshold of p-value = 0.0125 (0.05/4) as the final threshold to correct the multiple testing brought by multiple traits in the WGCNA analysis.
5) The plots for umbilical histological data (Fig 5 C) contain more than 5 points, but the use of replicate sections is not specified. If replicate sections were used, the authors should control for non-independence of replicate sections in their analyses (i.e., random effects model).
We did not use replicate sections. Figure 5C shows the umbilical artery intima and media. Because each human umbilical cord includes two umbilical arteries, the 5 vs. 5 individual comparison generates 10 vs. 10 umbilical artery comparison. To be clearer, we added an explanation in the revised manuscript.
On more minor notes:
There is significant and relevant published data on sex differences and hypoxia in rodents (see Cuffe et al 2014, "Mid- to late-term hypoxia in the mouse alters placental morphology, glucocorticoid regulatory pathways, and nutrient transporters in a sex-specific manner" and review by Siragher and Sferuzzi-Perro 2021, "Placental hypoxia: What have we learnt from small animal models?"), and historical work reporting sex differences in placental traits associated with high elevation adaptation in Andeans (series of publications by Moira Jackson in the late 1980s, reviewed in Wilsterman and Cheviron 2021, "Fetal growth, high altitude, and evolutionary adaptation: A new perspective").
We thank the reviewer for the constructive comments on literature review. We have cited and discussed them in the revised manuscript.
Reviewer #3 (Public Review):
More than 80 million people live at high altitude. This impacts health outcomes, including those related to pregnancy. Longer-lived populations at high altitudes, such as the Tibetan and Andean populations show partial protection against the negative health effects of high altitude. The paper by Yue sought to determine the mechanisms by which the placenta of Tibetans may have adapted to minimise the negative effect of high altitude on fetal growth outcomes. It compared placentas from pregnancies from Tibetans to those from the Han Chinese. It employed RNAseq profiling of different regions of the placenta and fetal membranes, with some follow-up of histological changes in umbilical cord structure and placental structure. The study also explored the contribution of fetal sex in these phenotypic outcomes.
A key strength of the study is the large sample sizes for the RNAseq analysis, the analysis of different parts of the placenta and fetal membranes, and the assessment of fetal sex differences.
A main weakness is that this study, and its conclusions, largely rely on transcriptomic changes informed by RNAseq. Changes in genes and pathways identified through bioinformatic analysis were not verified by alternate methods, such as by western blotting, which would add weight to the strength of the data and its interpretations. There is also a lack of description of patient characteristics, so the reader is unable to make their own judgments on how placental changes may link to pregnancy outcomes. Another weakness is that the histological analyses were performed on n=5 per group and were rudimentary in nature.
For the weakness raised by the reviewer, here are our responses:
(1) Considering that our conclusions largely rely on the transcriptomic data, we agree with reviewer that more experiments are needed to validate the results from our transcriptomic data. However, this study was mainly aimed to provide a transcriptomic landscape of high-altitude placenta, and to characterize the gene-expression difference between native Tibetans and Han migrants. The molecular mechanism exploration is not the main task of this study, and more validation experiments are warranted in the future.
(2) For the lack of description of patient characteristics, actually, we provided three level results on the placental changes of Tibetans: macroscopic phenotypes (higher placental weight and volume), histological phenotypes (larger umbilical vein walls and umbilical artery intima and media; lower syncytial knots/villi ratios) and transcriptomic phenotypes (DEG and differential modules). Combined with the previous studies, these placenta changes suggest a better reproductive outcome. For example, the placenta volume shows a significantly positive correlation with birth weight (R = 0.31, p-value = 2.5e-16), therefore, the larger placenta volume of Tibetans is beneficial to fetal development at high altitude. In addition, the larger umbilical vein wall and umbilical artery intima and media of Tibetans can explain their adaptation in preventing preeclampsia.
(3) For the sample size of histological analyses, we understand the reviewer’s concern that 5 vs. 5 samples are not large in histological analyses. This is because it was difficult to collect high-altitude Han placenta samples, and we only got 13 Han samples, from which we selected 5 infant sex matched samples.
References
Beall, C.M., Cavalleri, G.L., Deng, L.B., Elston, R.C., Gao, Y., Knight, J., Li, C.H., Li, J.C., Liang, Y., McCormack, M., et al. (2010). Natural selection on EPAS1 (HIF2 alpha) associated with low hemoglobin concentration in Tibetan highlanders. P Natl Acad Sci USA 107, 11459-11464.
Cho, J.I., Basnyat, B., Jeong, C., Di Rienzo, A., Childs, G., Craig, S.R., Sun, J., and Beall, C.M. (2017). Ethnically Tibetan women in Nepal with low hemoglobin concentration have better reproductive outcomes. Evol Med Public Health 2017, 82-96. He, Y., Guo, Y., Zheng, W., Yue, T., Zhang, H., Wang, B., Feng, Z., Ouzhuluobu, Cui, C., Liu, K., et al. (2023). Polygenic adaptation leads to a higher reproductive fitness of native Tibetans at high altitude. Curr Biol.
He, Y., Li, J., Yue, T., Zheng, W., Guo, Y., Zhang, H., Chen, L., Li, C., Li, H., Cui, C., et al. (2022). Seasonality and Sex-Biased Fluctuation of Birth Weight in Tibetan Populations. Phenomics 2, 64-71.
Peng, Y., Cui, C., He, Y., Ouzhuluobu, Zhang, H., Yang, D., Zhang, Q., Bianbazhuoma, Yang, L., He, Y., et al. (2017). Down-Regulation of EPAS1 Transcription and Genetic Adaptation of Tibetans to High-Altitude Hypoxia. Mol Biol Evol 34, 818-830.
Racimo, F., Berg, J.J., and Pickrell, J.K. (2018). Detecting Polygenic Adaptation in Admixture Graphs. Genetics 208, 1565-1584.



