Peer review process
Revised: This Reviewed Preprint has been revised by the authors in response to the previous round of peer review; the eLife assessment and the public reviews have been updated where necessary by the editors and peer reviewers.
Read more about eLife’s peer review process.Editors
- Reviewing EditorGustavo GoldmanUniversidade de Sao Paulo, Sao Paulo, Brazil
- Senior EditorJonathan CooperFred Hutch Cancer Center, Seattle, United States of America
Reviewer #1 (Public Review):
While I acknowledge the authors' effort in conducting Southern blot analysis to address my prior concern regarding the presence of dual copies of torA and tapA, I find their current resolution inadequate. Specifically, the simple deletion of the respective result sections for torA and tapA significantly impacts the overall significance of this study. The repeated unsuccessful attempts to generate correct mutants only offer circumstantial evidence, as technical issues may have been a contributing factor. Therefore, instead of merely removing these sections, it is essential for the authors to present more compelling experimental data demonstrating that torA and tapA are indeed vital for the viability of A. flavus. Such data would enhance the overall significance of this study.
Reviewer #2 (Public Review):
In this study, authors identified TOR, HOG and CWI signaling network genes as modulators of the development, aflatoxin biosynthesis and pathogenicity of A. flavus by gene deletions combined with phenotypic observation. They also analyzed the specific regulatory process and proposed that the TOR signaling pathway interacts with other signaling pathways (MAPK, CWI, calcineurin-CrzA pathway) to regulate the responses to various environmental stresses. Notably, they found that FKBP3 is involved in sclerotia and aflatoxin biosynthesis and rapamycin resistance in A. flavus, especially that the conserved site K19 of FKBP3 plays a key role in regulating aflatoxin biosynthesis. In general, the study involved a heavy workload and the findings are potentially interesting and important for understanding or controlling the aflatoxin biosynthesis. However, the findings have not been deeply explored and the conclusions mostly are based on parallel phenotypic observations.
Author Response
The following is the authors’ response to the original reviews.
Response to Reviewer 1 Comments (PublicReview)
Point 1: First, the authors should provide more convincing data showing that tor and tapA genes are indeed duplicated genes in A. flavus. The authors appeared to use the A. flavus PTS strain as a parental strain for constructing the tor and tapA mutants. If so, the A. flavus CA14 strain (Hua et al., 2007) should be the parental wild-type strain for the A. flavus PTS strain. I did a BLAST search in NCBI for the torA (AFLA_044350) and tapA (AFLA_092770) genes using the most recent CA14 genome assembly sequence (GCA_014784225.2) and only found one allele for each gene: torA on chromosome 7 and tapA on chromosome 3. I could not find any other parts with similar sequences. Even in another popular A. flavus wild-type strain, NRRL3357, both torA and tapA exist as a single allele. Based on the published genome assembly data for A. flavus, there is no evidence to support the idea that tor and tapA exist as copies of each other. Therefore, the authors could perform a Southern blot analysis to further verify their claim. If torA and tapA indeed exist as duplicate copies in different chromosomal locations, Southern blot data could provide supporting results.
Response 1: We thank the reviewer for their insightful observation. Based on the southern blot analysis results presented in Figure 1, we have determined that torA and tapA are single-copy genes. Additionally, we conducted protoplast transformation experiments repeated several times. which revealed that both torA and tapA transformants exhibited ectopic mutations. It is plausible that the deletion of torA and tapA genes may lead to the demise of A. flavus, this phenomenon is consistent with previous studies conducted on the fungus Fusarium graminearum[1].To ensure the rigor of the study, we have retracted the previously incorrect conclusion. We once again express our heartfelt appreciation to the experts for their valuable suggestions.
Author response image 1.
Fig.1 Southern blot hybridization analyses of WT, torA, and tapA transformants. (A) The structure diagram of the torA gene. (B) The structure diagram of the tapA gene. (C) Southern blot hybridization analyses of torA gene. (D) Southern blot hybridization analyses of tapA gene.
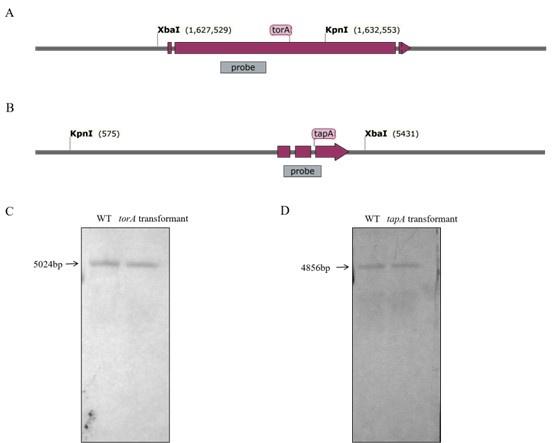
Point 2: Second, the authors should consider the possibility of aneuploidy for their constructed mutants. When an essential gene is targeted for deletion, aneuploidy often occurs even in a fungal strain without the "ku" mutation, which results in seemingly dual copies of the gene. As the authors appear to use the A. flavus PTS strain having the "ku" mutation, the parental strain has increased genome instability, which may result in enhanced chromosomal rearrangements. So, it will be necessary to Illumina-sequence their tor and tapA mutants to make sure that they are not aneuploidy.
Response 2: Thank you for your comment. Based on the sequencing results of the torA and tapA mutants, it was determined that the torA and tapA genes were still present in both mutants. In this case, it suggests that the torA and tapA genes may have undergone a genetic rearrangement or insertion at a different site in the mutant strains.
Point 3: Furthermore, the genetic nomenclature +/- and -/- should be reserved for heterozygous and homozygous mutants in a diploid strain. As A. flavus is not a diploid strain, this type of description could cause confusion for the readers.
Response 3: Thank you for your suggestion. We acknowledge your concerns about potential confusion caused by using this type of description, and we agree that it is best to avoid any misunderstandings for readers. Therefore, we have decided to remove this part of the content from the manuscript.
Response to Reviewer 2 Comments (PublicReview)
Point 1: However, findings have not been deeply explored and conclusions are mostly are based on parallel phenotypic observations. In addition, there are some concerns for the conclusions.
Response 1: We are grateful for the suggestion. We conduct additional experiments and analyses to provide a more comprehensive understanding and address concerns raised.
Response to Reviewer 3 Comments (PublicReview)
Point 1: The paper by Li et al. describes the role of the TOR pathway in Aspergillus flavus. The authors tested the effect of rapamycin in WT and different deletion strains. This paper is based on a lot of experiments and work but remains rather descriptive and confirms the results obtained in other fungi. It shows that the TOR pathway is involved in conidiation, aflatoxin production, pathogenicity, and hyphal growth. This is inferred from rapamycin treatment and TOR1/2 deletions. Rapamycin treatment also causes lipid accumulation in hyphae. The phenotypes are not surprising as they have been shown already for several fungi. In addition, one caveat is in my opinion that the strains grow very slowly and this could cause many downstream effects. Several kinases and phosphatases are involved in the TOR pathway. They were known from S. cerevisiae or filamentous fungi. The authors characterized them as well with knock-out approaches.
Response 1: Thank you for your comment. The role of the target of rapamycin (TOR) signaling pathway is of fundamental importance in the physiological processes of diverse eukaryotic organisms. Nevertheless, its precise involvement in regulating the developmental and virulent characteristics of opportunistic pathogenic fungi, such as A. flavus, has yet to be fully elucidated. Furthermore, the mechanistic underpinnings of TOR pathway activity specifically in A. flavus remain largely unresolved. Consequently, our study represents a significant contribution as the first comprehensive exploration of the conserved TOR signaling pathway encompassing a majority of its constituent genes in A. flavus.
Response to Reviewer 1 Comments (Recommendations For The Authors)
Point 1: In Table S3, the authors indicated that the Δku70 ΔniaD ΔpyrG::pyrG strain is A. flavus wild-type strain. However, this strain is not a wild-type strain because it seems like a control strain after introducing the pyrG gene into the A. flavus PTS strain (Δku70 ΔniaD ΔpyrG). So please indicate the real wild-type A. flavus strain name to help readers find out its original genome sequence data. Also, the reference for this Δku70 ΔniaD ΔpyrG::pyrG strain is "saved in our lab". This is not an eligible reference. If you use this control strain for the first time in this study, it should be described as "In this study". Otherwise, please indicate the proper reference for which the strain was first used.
Response 1: Thank you for your valuable feedback on our manuscript. We appreciate your attention to detail and the opportunity to clarify the information regarding the strain in Table S3. The A. flavus CA14 strain which produces aflatoxins and large sclerotia was isolated from a pistachio bud in the Wolfskill Grant Experimental Farm (University of Davis, Winters, California, USA)[2]. The A. flavus CA14 strain is the parental wild-type strain for the A. flavus CA14 PTs (Δku70, ΔniaD, ΔpyrG) strain. The recipient strain CA14 PTs has been used satisfactorily in gene knockout and subsequent genetic complementation experiments[3]. In this study, the A. flavus CA14 PTs strain was used as the transformation recipient strain, and the control strain (Δku70, ΔniaD, ΔpyrG::pyrG) created by introducing the pyrG gene into the A. flavus CA14 PTs strain. Refer to previously published literature[4],this control strain (Δku70, ΔniaD, ΔpyrG::pyrG) was named wild-type strain. Therefore, this control strain was also named wild-type strain in this study. As this control strain is indeed used in this study, we will revise the reference to "In this study" Once again, we appreciate your keen attention to detail and thank you for bringing these issues to our attention.
Response to Reviewer 2 Comments (Recommendations For The Authors)
Point 1: As in response: However, the tor gene in A. flavus exhibited varying copy numbers, as was confirmed by absolute quantification PCR at the genome level (Table S1). However, it is hard to understand Table S1: Estimation of copy number of tor gene in A. flavus toro and sumoo stand for the initial copy number, and the data are figured as the mean {plus minus} 95% confidence limit. CN is copy number. As indicated in the section of Method, using sumo gene as reference, the tor and tapA gene copy number was calculated by standard curve. In Table S1 of WT, for tor gene, CN value is 1412537 compared to 1698243 in tor+/-, for the reference gene sumo,794328 compared to1584893, how these data could support copy gene numbers of tor?
Response 1: Thank you for your suggestion. We understand the confusion with the data presented in Table S1 regarding the copy number estimation of the tor gene in A. flavus. We apologize for not providing a clear explanation for the data in the table. Quantitative real-time PCR (qPCR) is widely used to determine the copy number of a specific gene. It involves amplifying the gene of interest and a reference gene simultaneously using specific primers and probes. By comparing the amplification curves of the gene of interest and the reference gene, you can estimate the relative copy number of the gene.
To address your concern and provide more accurate information, we have re-performed the copy number analysis using southern blot. Southern blot analysis allows for the direct estimation of gene copy number by hybridizing genomic DNA with a specific probe for the gene. This method provides more reliable and accurate results in determining gene copy numbers. The southern blot analysis results are presented in Figure 1.
We appreciate your input and apologize for any confusion caused by the earlier presentation of the data.
Point 2: In response: For the knockout of the FRB domain, we used the homologous recombination method, but because tor genes are double-copy genes, there are also double copies in the FRB domain. Despite our efforts, we encountered challenges in precisely determining the location of the other copy of the tor gene. I could not understand these consistent data, why not for using sequencing?
Response 2: Thank you for your comment. We observed that the torA gene is a single copy. We removed this part of the results to avoid any ambiguity or potential misinterpretation.
Point 3: Response in Due to the large number of genes involved, we did not perform a complementation experiment. If there were no complementation data, how to demonstrate data are solid?
Response 3: Thank you for your important suggestion. We understand that complementation experiments are commonly used to validate gene deletions. Therefore, to ensure the reliability of our data, we have conducted supplementary experiments on specific gene deletions, such as ΔsitA-C and Δppg1-C. Thank you again for your positive comments and valuable suggestions to improve the quality of our manuscript.
References:
(1) Yu F, Gu Q, Yun Y, et al. The TOR signaling pathway regulates vegetative development and virulence in Fusarium graminearum. New Phytol. 2014; 203(1): 219-32.
(2) Hua SS, Tarun AS, Pandey SN, Chang L, Chang PK. Characterization of AFLAV, a Tf1/Sushi retrotransposon from Aspergillus flavus. Mycopathologia. 2007 Feb;163(2):97-104.
(3) Chang PK, Scharfenstein LL, Mack B, Hua SST. Genome sequence of an Aspergillus flavus CA14 strain that is widely used in gene function studies. Microbiol Resour Announc. 2019 Aug 15;8(33):e00837-19.
(4) Zhu Z, Yang M, Yang G, Zhang B, Cao X, Yuan J, Ge F, Wang S. PP2C phosphatases Ptc1 and Ptc2 dephosphorylate PGK1 to regulate autophagy and aflatoxin synthesis in the pathogenic fungus Aspergillus flavus. mBio. 2023 Oct 31;14(5):e0097723.



