Peer review process
Revised: This Reviewed Preprint has been revised by the authors in response to the previous round of peer review; the eLife assessment and the public reviews have been updated where necessary by the editors and peer reviewers.
Read more about eLife’s peer review process.Editors
- Reviewing EditorEmilio KropffFundación Instituto Leloir, Buenos Aires, Argentina
- Senior EditorLaura ColginUniversity of Texas at Austin, Austin, United States of America
Reviewer #1 (Public Review):
Summary:
The authors provide very compelling evidence that the lateral septum (LS) engages in theta cycle skipping.
Strengths:
The data and analysis is highly compelling regarding the existence of cycle skipping.
Comments on the revised version:
All previous recommendations were addressed in this revision.
Reviewer #2 (Public Review):
Summary
Recent evidence indicates that cells of the navigation system representing different directions and whole spatial routes fire in a rhythmic alternation during 5-10 Hz (theta) network oscillation (Brandon et al., 2013, Kay et al., 2020). This phenomenon of theta cycle skipping was also reported in broader circuitry connecting the navigation system with the cognitive control regions (Jankowski et al., 2014, Tang et al., 2021). Yet nothing was known about the translation of these temporally separate representations to midbrain regions involved in reward processing as well as the hypothalamic regions, which integrate metabolic, visceral, and sensory signals with the descending signals from the forebrain to ensure adaptive control of innate behaviors (Carus-Cadavieco et al., 2017). The present work aimed to investigate theta cycle skipping and alternating representations of trajectories in the lateral septum, neurons of which receive inputs from large number of CA1 and nearly all CA3 pyramidal cells (Risold and Swanson, 1995). While spatial firing has been reported in the lateral septum before (Leutgeb and Mizumori, 2002, Wirtshafter and Wilson, 2019), its dynamic aspects have remained elusive. The present study replicates the previous findings of theta-rhythmic neuronal activity in the lateral septum and reports a temporal alternation of spatial representations in this region, thus filling an important knowledge gap and significantly extending the understanding of the processing of spatial information in the brain. The lateral septum thus propagates the representations of alternative spatial behaviors to its efferent regions. The results can instruct further research of neural mechanisms supporting learning during goal-oriented navigation and decision-making in the behaviourally crucial circuits entailing the lateral septum.
Strengths
To this end, cutting-edge approaches for high-density monitoring of neuronal activity in freely behaving rodents and neural decoding were applied. Strengths of this work include comparisons of different anatomically and probably functionally distinct compartments of the lateral septum, innervated by different hippocampal domains and projecting to different parts of the hypothalamus; large neuronal datasets including many sessions with simultaneously recorded neurons; consequently, the rhythmic aspects of the spatial code could be directly revealed from the analysis of multiple spike trains, which were also used for decoding of spatial trajectories; and comparisons of the spatial coding between the two differently reinforced tasks.
Weaknesses
Without using perturbation techniques, the present approach could not identify the aspects of the spatial code actually influencing the generation of behaviors by downstream regions.
Author response:
The following is the authors’ response to the original reviews.
We thank the reviewers for their constructive comments and suggestions. We have prepared a revised manuscript with updated quantification of theta cycle skipping, new statistical comparisons of the difference between the two behavioral tasks, and general improvements to the text and figures.
Reviewer #1 (Public Review):
Summary
The authors provide very compelling evidence that the lateral septum (LS) engages in theta cycle skipping.
Strengths
The data and analysis are highly compelling regarding the existence of cycle skipping.
Weaknesses
The manuscript falls short on in describing the behavioral or physiological importance of the witnessed theta cycle skipping, and there is a lack of attention to detail with some of the findings and figures:
More/any description is needed in the article text to explain the switching task and the behavioral paradigm generally. This should be moved from only being in methods as it is essential for understanding the study.
Following this suggestion, we have expanded the description of the behavioral tasks in the Results section.
An explanation is needed as to how a cell can be theta skipping if it is not theta rhythmic.
A cell that is purely theta skipping (i.e., always fires on alternating theta cycles and never on adjacent theta cycles) will only have enhanced power at half theta frequency and not at theta frequency. Such a cell will therefore not be considered theta rhythmic in our analysis. Note, however, that there is a large overlap between theta rhythmic and theta skipping cell populations in our data (Figure 3 - figure supplement 2), indicating that most cells are not purely theta skipping.
The most interesting result, in my opinion, is the last paragraph of the entire results section, where there is more switching in the alternation task, but the reader is kind of left hanging as to how this relates to other findings. How does this relate to differences in decoding of relative arms (the correct or incorrect arm) during those theta cycles or to the animal's actual choice? Similarly, how does it relate to the animal's actual choice? Is this phenomenon actually behaviorally or physiologically meaningful at all? Does it contribute at all to any sort of planning or decision-making?
We agree that the difference between the two behavioral tasks is very interesting. It may provide clues about the mechanisms that control the cycle-by-cycle expression of possible future paths and the potential impact of goal-directed planning and (recent) experience. In the revised manuscript, we have expanded the analysis of the differences in theta-cycle dynamics between the two behavioral tasks. First, we confirm the difference through a new quantification and statistical comparison. Second, we performed additional analyses to explore the idea that the alternation of non-local representations reflects the number of relevant paths available to the animal (Figure 11 – figure supplements 2 and 3), but this did not appear to be the case. However, these results provide a starting point for future studies to clarify the task dependence of the theta- cycle dynamics of spatial representations and to address the important question of behavioral/physiological relevance.
The authors state that there is more cycle skipping in the alternation task than in the switching task, and that this switching occurs in the lead-up to the choice point. Then they say there is a higher peak at ~125 in the alternation task, which is consistent. However, in the final sentence, the authors note that "This result indicates that the representations of the goal arms alternate more strongly ahead of the choice point when animals performed a task in which either goal arm potentially leads to reward." Doesn't either arm potentially lead to a reward (but different amounts) in the switching task, not the alternation task? Yet switching is stronger in the alternation task, which is not constant and contradicts this last sentence.
The reviewer is correct that both choices lead to (different amounts of) reward in the switching task. As written, the sentence that the reviewer refers to is indeed not accurate and we have rephrased it to: “This result indicates that the representations of the goal arms alternate more strongly ahead of the choice point when animals performed a task in which either goal arm potentially leads to a desirable high-value reward.”.
Additionally, regarding the same sentence - "representations of the goal arms alternate more strongly ahead of the choice point when the animals performed a task in which either goal arm potentially leads to reward." - is this actually what is going on? Is there any reason at all to think this has anything to do with reward versus just a navigational choice?
We appreciate the reviewer’s feedback and acknowledge that our statement needs clarification. At the choice point in the Y-maze there are two physical future paths available to the animal (disregarding the path that the animal took to reach the choice point) – we assume this is what the reviewer refers to as “a navigational choice”. One hypothesis could be that alternation of goal arm representations is present whenever there are multiple future paths available, irrespective of the animal’s (learned) preference to visit one or the other goal arm. However, the reduced alternation of goal arm representations in the switching task that we report, suggests that the animal’s recent history of goal arm visits and reward expectations likely do influence the theta-cycle representations ahead of the choice point. We have expanded our analysis to test if theta cycle dynamics differ for trials before and after a switch in reward contingency in the switching task, but there was no statistical difference in our data. We have rewritten and expanded this part of the results to make our point more clearly.
Similarly, the authors mention several times that the LS links the HPC to 'reward' regions in the brain, and it has been found that the LS represents rewarded locations comparatively more than the hippocampus. How does this relate to their finding?
Indeed, Wirtshafter and Wilson (2020) reported that lateral septum cells are more likely to have a place field close to a reward site than elsewhere in their double-sided T-maze. It is possible that this indicates a shift towards reward or value representations in the lateral septum. In our study we did not look at reward-biased cells and whether they are more or less likely to engage in theta cycle skipping. This could be a topic for future analyses. It should be noted that the study by Wirtshafter and Wilson (2020) reports that a reward bias was predominantly present for place fields in the direction of travel away from the reward site. These reward-proximate LS cells may thus contribute to theta-cycle skipping in the inbound direction, but it is not clear if these cells would be active during theta sweeps when approaching the choice point in the outbound direction.
Reviewer #2 (Public Review)
Summary
Recent evidence indicates that cells of the navigation system representing different directions and whole spatial routes fire in a rhythmic alternation during 5-10 Hz (theta) network oscillation (Brandon et al., 2013, Kay et al., 2020). This phenomenon of theta cycle skipping was also reported in broader circuitry connecting the navigation system with the cognitive control regions (Jankowski et al., 2014, Tang et al., 2021). Yet nothing was known about the translation of these temporally separate representations to midbrain regions involved in reward processing as well as the hypothalamic regions, which integrate metabolic, visceral, and sensory signals with the descending signals from the forebrain to ensure adaptive control of innate behaviors (Carus-Cadavieco et al., 2017). The present work aimed to investigate theta cycle skipping and alternating representations of trajectories in the lateral septum, neurons of which receive inputs from a large number of CA1 and nearly all CA3 pyramidal cells (Risold and Swanson, 1995). While spatial firing has been reported in the lateral septum before (Leutgeb and Mizumori, 2002, Wirtshafter and Wilson, 2019), its dynamic aspects have remained elusive. The present study replicates the previous findings of theta-rhythmic neuronal activity in the lateral septum and reports a temporal alternation of spatial representations in this region, thus filling an important knowledge gap and significantly extending the understanding of the processing of spatial information in the brain. The lateral septum thus propagates the representations of alternative spatial behaviors to its efferent regions. The results can instruct further research of neural mechanisms supporting learning during goal-oriented navigation and decision-making in the behaviourally crucial circuits entailing the lateral septum.
Strengths
To this end, cutting-edge approaches for high-density monitoring of neuronal activity in freely behaving rodents and neural decoding were applied. Strengths of this work include comparisons of different anatomically and probably functionally distinct compartments of the lateral septum, innervated by different hippocampal domains and projecting to different parts of the hypothalamus; large neuronal datasets including many sessions with simultaneously recorded neurons; consequently, the rhythmic aspects of the spatial code could be directly revealed from the analysis of multiple spike trains, which were also used for decoding of spatial trajectories; and comparisons of the spatial coding between the two differently reinforced tasks.
Weaknesses
Possible in principle, with the present data across sessions, longitudinal analysis of the spatial coding during learning the task was not performed. Without using perturbation techniques, the present approach could not identify the aspects of the spatial code actually influencing the generation of behaviors by downstream regions.
Reviewer #3 (Public Review)
Summary
Bzymek and Kloosterman carried out a complex experiment to determine the temporal spike dynamics of cells in the dorsal and intermediate lateral septum during the performance of a Y-maze spatial task. In this descriptive study, the authors aim to determine if inputting spatial and temporal dynamics of hippocampal cells carry over to the lateral septum, thereby presenting the possibility that this information could then be conveyed to other interconnected subcortical circuits. The authors are successful in these aims, demonstrating that the phenomenon of theta cycle skipping is present in cells of the lateral septum. This finding is a significant contribution to the field as it indicates the phenomenon is present in neocortex, hippocampus, and the subcortical hub of the lateral septal circuit. In effect, this discovery closes the circuit loop on theta cycle skipping between the interconnected regions of the entorhinal cortex, hippocampus, and lateral septum. Moreover, the authors make 2 additional findings: 1) There are differences in the degree of theta modulation and theta cycle skipping as a function of depth, between the dorsal and intermediate lateral septum; and 2) The significant proportion of lateral septum cells that exhibit theta cycle skipping, predominantly do so during 'non-local' spatial processing.
Strengths
The major strength of the study lies in its design, with 2 behavioral tasks within the Y-maze and a battery of established analyses drawn from prior studies that have established spatial and temporal firing patterns of entorhinal and hippocampal cells during these tasks. Primary among these analyses, is the ability to decode the animal's position relative to locations of increased spatial cognitive demand, such as the choice point before the goal arms. The presence of theta cycle skipping cells in the lateral septum is robust and has significant implications for the ability to dissect the generation and transfer of spatial routes to goals within and between the neocortex and subcortical neural circuits.
Weaknesses
There are no major discernable weaknesses in the study, yet the scope and mechanism of the theta cycle phenomenon remain to be placed in the context of other phenomena indicative of spatial processing independent of the animal's current position. An example of this would be the ensemble-level 'scan ahead' activity of hippocampal place cells (Gupta et al., 2012; Johnson & Redish, 2007). Given the extensive analytical demands of the study, it is understandable that the authors chose to limit the analyses to the spatial and burst firing dynamics of the septal cells rather than the phasic firing of septal action potentials relative to local theta oscillations or CA1 theta oscillations. Yet, one would ideally be able to link, rather than parse the phenomena of temporal dynamics. For example, Tingley et al recently showed that there was significant phase coding of action potentials in lateral septum cells relative to spatial location (Tingley & Buzsaki, 2018). This begs the question as to whether the non-uniform distribution of septal cell activity within the Y-maze may have a phasic firing component, as well as a theta cycle skipping component. If so, these phenomena could represent another means of information transfer within the spatial circuit during cognitive demands. Alternatively, these phenomena could be part of the same process, ultimately representing the coherent input of information from one region to another. Future experiments will therefore have to sort out whether theta cycle skipping, is a feature of either rate or phase coding, or perhaps both, depending on circuit and cognitive demands.
The authors have achieved their aims of describing the temporal dynamics of the lateral septum, at both the dorsal extreme and the intermediate region. All conclusions are warranted.
Reviewer #1 (Recommendations For The Authors)
The text states: "We found that 39.7% of cells in the LSD and 32.4% of cells in LSI had significantly higher CSI values than expected by chance on at least one of the trajectories." The text in the supplemental figure indicates a p-value of 0.05 was used to determine significance. However, four trajectory categories are being examined so a Bonferroni correction should be used (significance at p<0.0125).
Indeed, a p-value correction for multiple tests should be performed when determining theta cycle skipping behavior for each of the four trajectories. We thank the reviewer for pointing out this oversight. We have implemented a Holm-Sidak p-value correction for the number of tested trajectories per cell (excluding trajectories with insufficient spikes). As a consequence, the number of cells with significant cycle-skipping activity decreased, but overall the results have not changed.
Figure 4 is very confusing as raster plots are displayed for multiple animals but it is unclear which animal the LFP refers to? The bottom of the plot is also referenced twice in the figure caption.
We apologize for the confusion. We have removed this figure in the revised manuscript, as it was not necessary to make the point about the spatial distribution of theta cycle skipping. Instead, we show examples of spatially-resolved cycle skipping in Figure 4 (formerly Figure 5 - supplementary figures 1 and 2) and we have added a plot with the spatially-resolved cycle skipping index for all analyzed cells in Figure 5A.
Figure 6 has, I think, an incorrect caption or figure. Only A and B are marked in the figure but A-G are mentioned in the caption but do not appear to correspond to anything in the figure.
Indeed, the caption was outdated. This has now been corrected.
Figure 8 is also confusing for several reasons: how is the probability scale on the right related to multiple semi-separate (top and middle) figures? In the top and bottom figures, it is not clear what the right and left sides refer to. It is also unclear why a probability of 0.25 is used for position (seems potentially low). The caption also mentions Figure A but there are no lettered "sub" figures in Figure 8.
The color bar on the right applies to both the top plot (directional decoding) and the middle plot (positional decoding). However, the maximum probability that is represented by black differs between the top and middle plots. We acknowledge that a shared color bar may lead to confusion and we have given each of the plots a separate color bar.
As for the maximum probability of 0.25 for position: this was a typo in the legend. The correct maximum value is 0.5. In general, the posterior probability will be distributed over multiple (often neighboring) spatial bins, and the distribution of maximum probabilities will depend on the number of spatial bins, the level of spatial smoothing in the decoding algorithm, and the amount of decodable information in the data. It would be more appropriate to consider the integrated probability over a small section of the maze, rather than the peak probability that is assigned to a single 5 cm bin. Also, note that a posterior probability of 0.5 is many times higher than the probability associated with a uniform distribution, which is 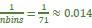 in our case.
in our case.
The left and right sides of the plots represent two different journeys that the animal ran. On the left an outbound journey is shown, and on the right an inbound journey. We have improved the figure and the description in the legend to make this clearer.
The reviewer is correct that there are no panels in Figure 8 and we have corrected the legend.
Some minor concerns
The introduction states that "a few studies have reported place cell-like activity in the lateral septum (Tingley and Buzsaki, 2018; Wirtshafter and Wilson, 2020, 2019)." However, notably and controversially, the Tingley study is one of the few studies to find NO place cell activity in the lateral septum. This is sort of mentioned later but the citation in this location should be removed.
The reviewer is correct, Tingley and Buzsaki reported a spatial phase code but no spatial rate code. We have removed the citation.
Stronger position/direction coding in the dLS consistent with prior studies and they should be cited in text (not a novel finding).
Thank you for pointing out this omission. Indeed, a stronger spatial coding in the dorsal lateral septum has been reported before, for example by Van der Veldt et al. (2021). We now cite this paper when discussing these findings.
Why is the alternation task administered for 30m but the switching task for 45m?
The reason is that rats received a larger reward in the switching task (in the high-reward goal arm) and took longer to complete trials on average. To obtain a more-or-less similar number of trials per session in both tasks, we extended the duration of switching task sessions to 45 minutes. We have added this explanation to the text.
Regarding the percentage of spatially modulated cells in the discussion, it is also worth pointing out that bits/sec information is consistent with previous studies.
Thank you for the suggestion. We now point out that the spatial information in our data is consistent with previous studies.
Reviewer #2 (Recommendations For The Authors)
While the results of the study are robust and timely, further details of behavioural training, additional quantitative comparisons, and improvements in the data presentation would make the study more comprehensible and complete.
Major comments
(1) I could not fully comprehend the behavioural protocols. They require a clearer explanation of both the specific rationale of the two tasks as well as a more detailed presentation of the protocols. Specifically:
(1.1) In the alternation task, were the arms baited in a random succession? How many trials were applied per session? Fig 1D: how could animals reach high choice accuracy if the baiting was random?
We used a continuous version of the alternation task, in which the animals were rewarded for left→home→right and right→home→left visit sequences. In addition, animals were always rewarded on inbound journeys. There was no random baiting of goal arms. Perhaps the confusion stems from our use of the word “trial” to refer to a completed lap (i.e., a pair of outbound/inbound journeys). On average, animals performed 54 of such trials per 30-minute session in the alternation task. We have expanded the description of the behavioral tasks in the Results and further clarified these points in the Methods section.
(1.2) Were they rewarded for correct inbound trials? If there was no reward, why were they considered correct?
Yes, rats received a reward at the home platform for correct inbound trials. We have now explicitly stated this in the text.
(1.3) In the switch alternation protocol, for how many trials was one arm kept more rewarding than the other, and how many trials followed after the rewarding value switch?
A switch was triggered when rats (of their own volition) visited the high-reward goal arm eight times in a row. Following a switch, the animals could complete as many trials as necessary until they visited the new high- reward goal arm in eight consecutive trials, which triggered another switch. As can be seen in Figure 1D, at the population level, animals needed ~13 trials to fully commit to the high-reward goal arm following a switch. We have further clarified the switching task protocol in the Results and Methods sections.
(1.4) What does the phrase "the opposite arm (as 8 consecutive visits)" exactly mean? Sounds like 8 consecutive visits signalled that the arm was rewarded (as if were not predefined in the protocol).
The task is self-paced and the animals initially visit both goal arms, before developing a bias for the high- reward goal arm. A switch of reward size was triggered as soon as the animal visited the high-reward goal arm for eight consecutive trials. We have rewritten the description of the switching task protocol, including this sentence, which hopefully clarifies the procedure.
(1.5) P. 15, 1st paragraph, Theta cycle skipping and alternation of spatial representations is more prominent in the alternation task. Why in the switching task, did rats visit the left and right arms approximately equally often if one was more rewarding than the other? How many switches were applied per recording session, and how many trials were there in total?
Both the left and right goal arms were sampled more or less equally by the animals because both goal arms at various times were associated with a large reward following switches in reward values during sessions. The number of switches per session varied from 1 to 3. Sampling of both goal arms was also evident at the beginning of each session and following each reward value switch, before animals switched their behavior to the (new) highly rewarded goal arm. In Table 1, we have now listed the number of trials and the number of reward-value switches for all sessions.
(1.6) Is the goal arm in figures the rewarded/highly rewarded arm only or are non-baited arms also considered here?
Both left and right arms are considered goal arms and were included in the analyses, irrespective of the reward that was received (or not received).
(2) The spatial navigation-centred behavioural study design and the interpretation of results highlight the importance of the dorsal hippocampal input to the LS. Yet, the recorded LSI cells are innervated by intermediate and ventral aspects of the hippocampus, and LS receives inputs from the amygdala and the prefrontal cortex, which together may together bring about - crucial for the adaptive behaviours regulated by the LS - reward, and reward-prediction-related aspects in the firing of LS cells during spatial navigation. Does success or failure to acquire reward in a trial modify spatial coding and cycle skipping of LSD vs. LSI cells in ensuing inbound and outbound trials?
This is an excellent question and given the length of the current manuscript, we think that exploration of this question is best left for a future extension of our study.
A related question: in Figure 10, it is interesting that cycle skipping is prominent in the goal arm for outbound switching trials and inbound trials of both tasks. Could it be analytically explained by task contingencies and behaviour (e.g. correct/incorrect trial, learning dynamics, running speed, or acceleration)?
Our observation of cycle skipping at the single-cell level in the goal arms is somewhat surprising and, we agree with the reviewer, potentially interesting. However, it was not accompanied by alternation of representations at the population level. Given the current focus and length of the manuscript, we think further investigation of cycle skipping in the goal arm is better left for future analyses.
(3) Regarding possible cellular and circuit mechanisms of cycle skipping and their relation to the alternating representations in the LS. Recent history of spiking influences the discharge probability; e.g. complex spike bursts in the hippocampus are associated with a post-burst delay of spiking. In LS, cycle skipping was characteristic for LS cells with high firing rates and was not uniformly present in all trajectories and arms. The authors propose that cycle skipping can be more pronounced in epochs of reduced firing, yet the opposite seems also possible - this phenomenon can be due to an intermittently increased drive onto some LS cells. Was there a systematic relationship between cycle skipping in a given cell and the concurrent firing rate or a recent discharge with short interspike intervals?
In our discussion, we tried to explain the presence of theta cycle skipping in the goal arms at the single-cell level without corresponding alternation dynamics at the population level. We mentioned the possibility of a decrease in excitatory drive. As the reviewer suggests, an increase in excitatory drive combined with post- burst suppression or delay of spiking is an alternative explanation. We analyzed the spatial tuning of cells with theta cycle skipping and found that, on average, these cells have a higher firing rate in the goal arm than the stem of the maze in both outbound and inbound run directions (Figure 5 – figure supplement 1). In contrast, cells that do not display theta cycle skipping do not show increased firing in the goal arm. These results are more consistent with the reviewer’s suggested mechanism and we have updated the discussion accordingly.
(4) Were the differences between the theta modulation (cycle skipping) of local vs. non-local representations (P.14, line 10-12, "In contrast...", Figure 9A) and between alternation vs. switching tasks (Figure 10 C,D) significantly different?
We have added quantification and statistical comparisons for the auto- and cross-correlations of the local/non-local representations. The results indeed show significantly stronger theta cycle skipping of the non-local representations as compared to the local representations (Figure 10 - figure supplement 1A), a stronger alternation of non-local representations in the outbound direction (Figure 10 - figure supplement 1B), and significant differences between the two tasks (Figure 11E,F).
(5) Regarding the possibility of prospective coding in LS, is the accurate coding of run direction not consistent with prospective coding? Can the direction be decoded from the neural activity in the start arm? Are the cycling representations of the upcoming arms near the choice point equally likely or preferential for the then- selected arm?
The coding of run direction (outbound or inbound) is distinct from the prospective/retrospective coding of the goal arm. As implemented, the directional decoding model does not differentiate between the two goal arms and accurate decoding of direction with this model can not inform us whether or not there is prospective (or retrospective) coding. To address the reviewer’s comments, we performed two additional analyses. First, we analyzed the directional (outbound/inbound) decoding performance as a function of location in the maze (Figure 6 - figure supplement 3E). The results show that directional decoding performance is high in both stem and goal arms. Second, we analyzed how well we can predict the trajectory type (i.e., to/from the left or right goal arm) as a function of location in the maze, and separately for outbound and inbound trajectories (Figure 6 - figure supplement 3C,D). The results show that on outbound journeys, decoding the future goal arm is close to chance when the animals are running along the stem. The decoding performance goes up around the choice point and reaches the highest level when animals are in the goal arm.
(6) Figure 10 seems to show the same or similar data as Figures 5 (A,B) and 9 (C,D).
Figure 10 (figure 11 in revised manuscript) re-analyzes the same data as presented in Figures 5 and 9, but separates the experimental sessions according to the behavioral task. We now explicitly state this.
Minor comments
(1) If cycle skipping in the periodicity of non-local representations was more prominent in alternation than in the switching task, one might expect them to be also prominent in early trials of the switching task, when the preference of a more rewarding arm is not yet established. Was this the case?
The reviewer makes an interesting suggestion. Indeed, if theta cycle skipping and the alternation of non-local representations reflect that there are multiple paths that the animal is considering, one may predict that the theta skipping dynamics are similar between the two tasks in early trials (as the reviewer suggests). Similarly, one may predict that in the switching task, the alternation of non-local representations is weaker immediately before a reward contingency switch (when the animal has developed a bias towards the goal arm with a large reward) as compared to after the switch.
We have now quantified the theta cycle dynamics of spatial representations in the early trials in each session of both tasks (Figure 11 - figure supplement 2) and in the trials before and after each switch in the switching task (Figure 11 - figure supplement 3).
The results of the early trial analysis indicate stronger alternation of non-local representations in the alternation task than in the switching task (consistent with the whole session analysis), which is contrary to the prediction.
The pre-/post-switch analysis did not reveal a significant difference between the trials before and after a reward contingency switch. If anything, there was a trend towards stronger theta cycle skipping/alternation in the trials before a switch, which would be opposite to the prediction.
These results do not appear to support the idea that the alternation of non-local representations reflects the number of relevant paths available to the animal. We have updated the text to incorporate these new data and discuss the implications.
(2) Summary: sounds like the encoding of spatial information and its readout in the efferent regions are equally well established.
Thank you for pointing this out.
(3) Summary: "motivation and reward processing centers such as the ventral tegmental area." How about also mentioning here the hypothalamus, which is a more prominent output of the lateral septum than the VTA?
We have now also mentioned the hypothalamus.
(4) "lateral septum may contribute to the hippocampal theta" - readers not familiar with details of the medial vs. lateral septum research may misinterpret the modest role of LS in theta compared to MS.
We have added “in addition to the strong theta drive originating from the medial septum” to make clear that the lateral septum has a modest role in hippocampal theta generation.
(5) "(Tingley and Buzsáki, 2018) found a lack of spatial rate coding in the lateral septum and instead reported a place coding by specific phases of the hippocampal theta rhythm (Rizzi-Wise and Wang, 2021) " needs rephrasing.
Thank you, we have rephrased the sentence.
(6) Figure 4 is a bit hard to generalize. The authors may additionally consider a sorted raster presentation of the dataset in this main figure.
We have removed this figure in the revised manuscript, as it was not necessary to make the point about the location of theta cycle skipping. Instead, we show examples of spatially-resolved cycle skipping in Figure 4 (formerly Figure 5 - supplementary figures 1 and 2), and, following the reviewer’s suggestion, we have added a plot with the spatially-resolved cycle skipping index for all analyzed cells (Figure 5A).
(7) It would help if legends of Figure 5 (and related supplementary figures) state in which of the two tasks the data was acquired, as it is done for Figure 10.
Thank you for the suggestion. The legends of Figure 4A,B (formerly Figure 5 – supplemental figures 1 and 2) and Figure 5 now include in which behavioral task the data was acquired.
(8) Page 10, "Spatial coding...", 1st Citing the initial report by Leugeb and Mizumori would be appropriate here too.
The reviewer is correct. We have added the citation.
(9) The legend in Figure 6 (panels A-G) does not match the figure (only panels A,B). What is shown in Fig. 6B, the legend does not seem to fully match.
Indeed, the legend was outdated. This has now been corrected.
(10) 7 suppl., if extended to enable comparisons, could be a main figure. Presently, Figure 7C does not account for the confounding effect of population size and is therefore difficult to interpret without complex comparisons with the Supplementary Figure which is revealing per se.
We thank the reviewer for their suggestion. We have changed Figure 7 such that it only shows the analysis of decoding performed with all LSD and LSI cells. Figure 7 – supplemental figure 1 has been transformed into main Figure 8, with the addition of a panel to show a statistical comparison between decoding performance in LSD and LSI with a fixed number of cells.
(11) 14, line 10 there is no Figure 8A
This has been corrected.
(12) 15 paragraph 1, is the discussed here model the one from Kay et al?
From Kay et al. (2020) and also Wang et al. (2020). We have added the citations.
(13) Figure 5 - Figure Supplement 1 presents a nice analysis that, in my view, can merit a main figure. I could not find the description of the colour code in CSI panels, does grey/red refer to non/significant points?
Indeed, grey/red refers to non-significant points and significant points respectively. We have clarified the color code in the figure legend. Following the reviewer’s suggestion, we have made Figure 5 Supplement 1 and 2 a main figure (Figure 4).
(14) Figure 5 -Figure Supplement 2. Half of the cells (255 and 549) seems not to be representative of the typically high SCI in the goal arm in left and right inbound trials combined (Figure 5 A). Were the changes in CSI in the right and left inbound trials similar enough to be combined in Fig 5A? Otherwise, considering left and right inbound runs separately and trying to explain where the differences come from would seem to make sense.
Figure 5 – figure supplement 2 is now part of the new main Figure 4. Originally, the examples were from a single session and the same cells as shown in the old Figure 4. However, since the old Figure 4 has been removed, we have selected examples from different sessions and both left/right trajectories that are more representative of the overall distribution. We have further added a plot with the spatially-resolved cycle skipping for all analyzed cells in Figure 5A.
(15) In the second paragraph of the Discussion, dorso-ventral topography of hippocampal projections to the LS (Risold and Swanson, Science, 90s) could be more explicitly stated here.
Thank you for the suggestion. We have now explicitly mentioned the dorsal-ventral topography of hippocampal-lateral septum projections and cite Risold & Swanson (1997).
(16) Discussion point: why do the differences in spatial information of cells in the ventral/intermediate vs. dorsal hippocampus not translate into similarly prominent differences in LSI vs. LSD?
In our data, we do observe clear differences in spatial coding between LSD and LSI. Specifically, cell activity in the LSD is more directional, has higher goal arm selectivity, and higher spatial information (we have now added statistical comparisons to Figure 6 – figure supplement 1). As a result, spatial decoding performance is much better for LSD cell populations than LSI cell populations (see updated Figure 8, with statistical comparison of decoding performance). Spatial coding in the LS is not as strong as in the hippocampus, likely because of the convergence of hippocampal inputs, which may give the impression of a less prominent difference between the two subregions.
(17) Discussion, last paragraph: citation of the few original anatomical and neurophysiological studies would be fitting here, in addition to the recent review article.
Thank you for the suggestion. We have added selected citations of the original literature.
(18) Methods, what was the reference electrode?
We used an external reference electrode that was soldered to a skull screw, which was positioned above the cerebellum. We have added this to the Methods section.
(19) Methods, Theta cycle skipping: bandwidth = gaussian kerner parameter?
The bandwidth is indeed a parameter of the Gaussian smoothing kernel and is equal to the standard deviation.
Reviewer #3 (Recommendations For The Authors)
Below I offer a short list of minor comments and suggestions that may benefit the manuscript.
(A) I was not able to access the Open Science Framework Repository. Can this be rectified?
Thank you for checking the OSF repository. The data and analysis code are now publicly available.
(B) In the discussion the authors should attempt to flesh out whether they can place theta cycle skipping into context with left/right sweeps or scan ahead phenomena, as shown in the Redish lab.
Thank you for the excellent suggestion. We have now added a discussion of the possible link between theta cycle skipping and the previously reported scan-ahead theta sweeps.
(C) What is the mechanism of cycle skipping? This could be relevant to intrinsic vs network oscillator models. Reference should also be made to the Deshmukh model of interference between theta and delta (Deshmukh, Yoganarasimha, Voicu, & Knierim, 2010).
We had discussed a potential mechanism in the discussion (2nd to last paragraph in the revised manuscript), which now includes a citation of a recent computational study (Chu et al., 2023). We have now also added a reference to the interference model in Deshmukh et al, 2010.
(D) Little background was given for the motivation and expectation for potential differences between the comparison of the dorsal and intermediate lateral septum. I don't believe that this is the same as the dorsal/ventral axis of the hippocampus, but if there's a physiological justification, the authors need to make it.
We have added a paragraph to the introduction to explain the anatomical and physiological differences across the lateral septum subregions that provide our rationale for comparing dorsal and intermediate lateral septum (we excluded the ventral lateral septum because the number of cells recorded in this region was too low).
(E) It would help to label "outbound" and "inbound" on several of the figures. All axes need to be labeled, with appropriate units indicated.
We have carefully checked the figures and added inbound/outbound labels and axes labels where appropriate.
(F) In Figure 6, the legend doesn't match the figure.
Indeed, the legend was outdated. This has now been corrected.
(G) The firing rate was non-uniform across the Y-maze. Does this mean that the cells tended to fire more in specific positions of the maze? If so, how would this affect the result? Would increased theta cycle skipping at the choice point translate to a lower firing rate at the choice point? Perhaps less overdispersion of the firing rate (Fenton et al., 2010)?
Individual cells indeed show a non-uniform firing rate across the maze. To address the reviewer’s comment and test if theta cycle skipping cells were active preferentially near the choice point or other locations, we computed the mean-corrected spatial tuning curves for cell-trajectory pairs with and without significant theta cycle skipping. This additional analysis indicates that, on average, the population of theta cycle skipping cells showed a higher firing rate in the goal arms than in the stem of the maze as compared to non-skipping cells for outbound and inbound directions (shown in Figure 5 - figure supplement 1).
(H) As mentioned above, it could be helpful to look at phase preference. Was there an increased phase preference at the choice point? Would half-cycle firing correlate with an increased or decreased phase preference? Based on prior work, one would expect increased phase preference, at least in CA1, at the choice point (Schomburg et al., 2014). In contrast, other work might predict phasic preference according to spatial location (Tingley & Buzsaki, 2018). Including phase analyses is a suggestion, of course. The manuscript is already sufficiently novel and informative. Yet, the authors should state why phase was not analyzed and that these questions remain for follow-up analyses. If the authors did analyze this and found negative results, it should be included in this manuscript.
We thank the reviewer for their suggestion. We have not yet analyzed the theta phase preference of lateral septum cells or other relations to the theta phase. We agree that this would be a valuable extension of our work, but prefer to leave it for future analyses.
(I) One of the most important aspects of the manuscript, is that there is now evidence of theta cycle skipping in the circuit loop between the EC, CA1, and LS. This now creates a foundation for circuit-based studies that could dissect the origin of route planning. Perhaps the authors should state this? In the same line of thinking, how would one determine whether theta cycle skipping is necessary for route planning as opposed to a byproduct of route planning? While this question is extremely complex, other studies have shown that spatial navigation and memory are still possible during the optogenetic manipulation of septal oscillations (Mouchati, Kloc, Holmes, White, & Barry, 2020; Quirk et al., 2021). However, pharmacological perturbation or lesioning of septal activity can have a more profound effect on spatial navigation (Bolding, Ferbinteanu, Fox, & Muller, 2019; Winson, 1978). As a descriptive study, I think it would be helpful to remind the readers of these basic concepts.
We thank the reviewer for their comment and for pointing out possible future directions for linking theta cycle skipping to route planning. Experimental manipulations to directly test this link would be very challenging, but worthwhile to pursue. We now mention how circuit-based studies may help to test if theta cycle skipping in the broader subcortical-cortical network is necessary for route planning. Given that the discussion is already quite long, we decided to omit a more detailed discussion of the possible role of the medial septum (which is the focus of the papers cited by the reviewer).
Very minor points
(A) In the introduction, "one study" begins the sentence but there is a second reference.
Thank you, we have rephrased the sentence.
(B) Also in the introduction, it could be helpful to have an operational definition of theta cycle skipping (i.e., 'enhanced rhythmicity at half theta frequency').
We followed the reviewer’s suggestion.
(C) The others should be more explicit in the introduction about their main question. Theta cycle skipping exists in CA1, and then import some of the explanations mentioned in the discussion to the introduction (i.e., attractors states of multiple routes). The main question is then whether this phenomenon, and others from CA1, translate to the output in LS.
We have edited the introduction to more clearly state the main question of our study, following the suggestion from the reviewer.
(D) There are a few instances of extra closing parentheses.
We checked the text but did not find instances of erroneous extra closing parentheses. There are instances of nested parentheses, which may have given the impression that closing parentheses were duplicated.
(E) The first paragraph of the Discussion lacks sufficient references.
We have now added references to the first paragraph of the discussion.
(F) At the end of the 2nd paragraph in the Discussion, the comparison is missing. More than what? It's not until the next reference that one can assume that the authors are referring to a dorsal/ventral axis. However, the physiological motivation for this comparison is lacking. Why would one expect a dorsal/intermediate continuum for theta modulation as there is along the dorsal/ventral axis of the hippocampus?
Thank you for spotting this omission. We have rewritten the paragraph to more clearly make the parallel between dorsal-ventral gradients in the lateral septum and hippocampus and how this relates to the topographical connections between the two structures.



