Peer review process
Not revised: This Reviewed Preprint includes the authors’ original preprint (without revision), an eLife assessment, public reviews, and a provisional response from the authors.
Read more about eLife’s peer review process.Editors
- Reviewing EditorJuan Alvaro GallegoChampalimaud Foundation, Lisbon, Portugal
- Senior EditorTamar MakinUniversity of Cambridge, Cambridge, United Kingdom
Reviewer #1 (Public Review):
Summary:
This manuscript provides potentially important new information about ipsilateral cortical impact on locomotion. A number of issues need to be addressed.
Strengths:
The primary appeal and contribution of this manuscript are that it provides a range of different measures of ipsilateral cortical impact on locomotion in the setting of impaired contralateral control. While the pathways and mechanisms underlying these various measures are not fully defined and their functional impacts remain uncertain, they comprise a rich body of results that can inform and guide future efforts to understand cortical control of locomotion and to develop more effective rehabilitation protocols.
Weaknesses:
1. The authors state that they used a cortical stimulation location that produced the largest ankle flexion response (lines 102-104). Did other stimulation locations always produce similar, but smaller responses (aside from the two rats that showed ipsilateral neuromodulation)? Was there any site-specific difference in response to stimulation location?
2. Figure 2: There does not appear to be a strong relationship between the percentage of spared tissue and the ladder score. For example, the animal with the mild injury (based on its ladder score) in the lower left corner of Figure 2A has less than 50% spared tissue, which is less spared tissue than in any animal other than the two severe injuries with the most tissue loss. Is it possible that the ladder test does not capture the deficits produced by this spinal cord injury? Have the authors looked for a region of the spinal cord that correlates better with the deficits that the ladder test produces? The extent of damage to the region at the base of the dorsal column containing the corticospinal tract would be an appropriate target area to quantify and compare with functional measures.
3. Lines 219-221: The authors state that "phase-coherent stimulation reinstated the function of this muscle, leading to increased burst duration (90{plus minus}18% of the deficit, p=0.004, t-test, Fig. 4B) and total activation (56{plus minus}13% of the deficit, p=0.014, t-test, Fig. 3B). This way of expressing the data is unclear. For example, the previous sentence states that after SCI, burst duration decreased by 72%. Does this mean that the burst duration after stimulation was 90% higher than the -72% level seen with SCI alone, i.e., 90% + -72% = +18%? Or does it mean that the stimulation recovered 90% of the portion of the burst duration that had been lost after SCI, i.e., -72% * (100%-90%)= -7%? The data in Figure 4 suggests the latter. It would be clearer to express both these SCI alone and SCI plus stimulation results in the text as a percent of the pre-SCI results, as done in Figure 4.
4. Lines 227-229: The authors claim that the phase-dependent stimulation effects in SCI rats are immediate, but they don't say how long it takes for these effects to be expressed. Are these effects evident in the response to the first stimulus train, or does it take seconds or minutes for the effects to be expressed? After the initial expression of these effects, are there any gradual changes in the responses over time, e.g., habituation or potentiation?
5. Awake motor maps (lines 250-277): The analysis of the motor maps appears to be based on measurements of the percentage of channels in which a response can be detected. This analytic approach seems incomplete in that it only assesses the spatial aspect of the cortical drive to the musculature. One channel could have a just-above-threshold response, while another could have a large response; in either case, the two channels would be treated as the same positive result. An additional analysis that takes response intensity into account would add further insight into the data, and might even correlate with the measures of functional recovery. Also, a single stimulation intensity was used; the results may have been different at different stimulus intensities.
6. Lines 858-860: The authors state that "All tests were one-sided because all hypotheses were strictly defined in the direction of motor improvement." By using the one-sided test, the authors are using a lower standard for assessing statistical significance that the overwhelming majority of studies in this field use. More importantly, ipsilateral stimulation of particular kinds or particular sites might conceivably impair function, and that is ignored if the analysis is confined to detecting improvement. Thus, a two-sided analysis or comparable method should be used. This appropriate change would not greatly modify the authors' current conclusions about improvements.
Reviewer #2 (Public Review):
Summary:
The authors' long-term goals are to understand the utility of precisely phased cortex stimulation regimes on recovery of function after spinal cord injury (SCI). In prior work, the authors explored the effects of contralesion cortex stimulation. Here, they explore ipsilesion cortex stimulation in which the corticospinal fibers that cross at the pyramidal decussation are spared. The authors explore the effects of such stimulation in intact rats and rats with a hemisection lesion at the thoracic level ipsilateral to the stimulated cortex. The appropriately phased microstimulation enhances contralateral flexion and ipsilateral extension, presumably through lumbar spinal cord crossed-extension interneuron systems. This microstimulation improves weight bearing in the ipsilesion hindlimb soon after injury, before any normal recovery of function would be seen. The contralateral homologous cortex can be lesioned in intact rats without impacting the microstimulation effect on flexion and extension during gait. In two rats ipsilateral flexion responses are noted, but these are not clearly demonstrated to be independent of the contralateral homologous cortex remaining intact.
Strengths:
This paper adds to prior data on cortical microstimulation by the laboratory in interesting ways. First, the strong effects of the spared crossed fibers from the ipsi-lesional cortex in parts of the ipsi-lesion leg's step cycle and weight support function are solidly demonstrated. This raises the interesting possibility that stimulating the contra-lesion cortex as reported previously may execute some of its effects through callosal coordination with the ipsi-lesion cortex tested here. This is not fully discussed by the authors but may represent a significant aspect of these data. The authors demonstrate solidly that ablation of the contra-lesional cortex does not impede the effects reported here. I believe this has not been shown for the contra-lesional cortex microstimulation effects reported earlier, but I may be wrong.
Effects and neuroprosthetic control of these effects are explored well in the ipsi-lesion cortex tests here.
Weaknesses:
Some data is based on very few rats. For example (N=2) for ipsilateral flexion effects of microstimulation. N=3 for homologous cortex ablation, and only ipsi extension is tested it seems. There is no explicit demonstration that the ipsilateral flexion effects in only 2 rats reported can survive the contra-lateral cortex ablation.
Some improvements in clarity and precision of descriptions are needed, as well as fuller definitions of terms and algorithms.
Likely Impacts:
This data adds in significant ways to prior work by the authors, and an understanding of how phased stimulation in cortical neuroprosthetics may aid in recovery of function after SCI, especially if a few ambiguities in writing and interpretation are fully resolved.
Reviewer #3 (Public Review):
Summary:
This article aims to investigate the impact of neuroprosthesis (intracortical microstimulation) implanted unilaterally on the lesion side in the context of locomotor recovery following unilateral thoracic spinal cord injury.
Strength:
The study reveals that stimulating the left motor cortex, on the same side as the lesion, not only activates the expected right (contralateral) muscle activity but also influences unexpected muscle activity on the left (ipsilateral) side. These muscle activities resulted in a substantial enhancement in lift during the swing phase of the contralateral limb and improved trunk-limb support for the ipsilateral limb. They used different experimental and stimulation conditions to show the ipsilateral limb control evoked by the stimulation. This outcome holds significance, shedding light on the engagement of the "contralateral projecting" corticospinal tract in activating not only the contralateral but also the ipsilateral spinal network.
The experimental design and findings align with the investigation of the stimulation effect of contralateral projecting corticospinal tracts. They carefully examined the recovery of ipsilateral limb control with motor maps. They also tested the effective sites of cortical stimulation. The study successfully demonstrates the impact of electrical stimulation on the contralateral projecting neurons on ipsilateral limb control during locomotion, as well as identifying important stimulation spots for such an effect. These results contribute to our understanding of how these neurons influence bilateral spinal circuitry. The study's findings contribute valuable insights to the broader neuroscience and rehabilitation communities.
Weakness:
The term "ipsilateral" lacks a clear definition in the title, abstract, introduction, and discussion, potentially causing confusion for the reader.
The unexpected ipsilateral (left) muscle activity is most likely due to the left corticospinal neurons recruiting not only the right spinal network but also the left spinal network. This is probably due to the joint efforts of the neuroprosthesis and activation of spinal motor networks which work bilaterally at the spinal level.
However, in my opinion, readers can easily link the ipsilateral cortical network to the ipsilateral-projecting corticospinal tract, which is less likely to play a role in ipsilateral limb control in this study since this tract is disrupted by the thoracic spinal injury.
Author Response
We would like to thank the Editors and Reviewers for their comprehensive review of the manuscript. We appreciate your feedback, and we will carefully consider all your comments in the revision of the manuscript. Below are our provisional responses to your comments.
eLife assessment
This manuscript reveals important insights into the role of ipsilateral descending pathways in locomotion, especially following unilateral spinal cord injury. The study provides solid evidence that this method improves the injured side's ability to support weight, and as such the findings may lead to new treatments for stroke, spinal cord injuries, or unilateral cerebral injuries. However, the methods and results need to be better detailed, and some of the statistical analysis enhanced.
Thank you for your assessment. We will incorporate various textual enhancements in the final version of the manuscript to address the weaknesses you have pointed out. The specific improvements are outlined below.
Public Reviews:
Reviewer #1 (Public Review):
Summary:
This manuscript provides potentially important new information about ipsilateral cortical impact on locomotion. A number of issues need to be addressed.
Strengths:
The primary appeal and contribution of this manuscript are that it provides a range of different measures of ipsilateral cortical impact on locomotion in the setting of impaired contralateral control. While the pathways and mechanisms underlying these various measures are not fully defined and their functional impacts remain uncertain, they comprise a rich body of results that can inform and guide future efforts to understand cortical control of locomotion and to develop more effective rehabilitation protocols.
Weaknesses:
- The authors state that they used a cortical stimulation location that produced the largest ankle flexion response (lines 102-104). Did other stimulation locations always produce similar, but smaller responses (aside from the two rats that showed ipsilateral neuromodulation)? Was there any site-specific difference in response to stimulation location?
We derived motor maps in each rat, akin to the representation depicted in Fig 6. In each rat, alternative cortical sites did, indeed, produce distal or proximal contralateral leg flexion responses. Distal responses were more likely to be evoked in the rostral portion of the array, similarly to proximal responses early after injury. This distribution in responses across different cortical sites is reported in this study (Fig. 6) and is consistent with our prior work. The Results section will be revised to provide additional clarification and context for the data presented in Figure 6.
- Figure 2: There does not appear to be a strong relationship between the percentage of spared tissue and the ladder score. For example, the animal with the mild injury (based on its ladder score) in the lower left corner of Figure 2A has less than 50% spared tissue, which is less spared tissue than in any animal other than the two severe injuries with the most tissue loss. Is it possible that the ladder test does not capture the deficits produced by this spinal cord injury? Have the authors looked for a region of the spinal cord that correlates better with the deficits that the ladder test produces? The extent of damage to the region at the base of the dorsal column containing the corticospinal tract would be an appropriate target area to quantify and compare with functional measures.
In Fig. S6 of our 2021 publication "Bonizzato and Martinez, Science Translational Medicine", we investigated the predictive value of tissue sparing in specific sub-regions of the spinal cord for ladder performance. Specifically, we examined the correlation between the accuracy of left leg ladder performance in the acute state and the preservation of the corticospinal tract (CST). Our results indicated that dorsal CST sparing serves as a mild predictor for ladder deficits, confirming the results obtain in this study.
- Lines 219-221: The authors state that "phase-coherent stimulation reinstated the function of this muscle, leading to increased burst duration (90{plus minus}18% of the deficit, p=0.004, t-test, Fig. 4B) and total activation (56{plus minus}13% of the deficit, p=0.014, t-test, Fig. 3B). This way of expressing the data is unclear. For example, the previous sentence states that after SCI, burst duration decreased by 72%. Does this mean that the burst duration after stimulation was 90% higher than the -72% level seen with SCI alone, i.e., 90% + -72% = +18%? Or does it mean that the stimulation recovered 90% of the portion of the burst duration that had been lost after SCI, i.e., -72% * (100%-90%)= -7%? The data in Figure 4 suggests the latter. It would be clearer to express both these SCI alone and SCI plus stimulation results in the text as a percent of the pre-SCI results, as done in Figure 4.
Your assessment is correct; we intended to report that the stimulation recovered 90% of the portion of the burst duration that had been lost after SCI. This point will be addressed in the revision of the manuscript.
- Lines 227-229: The authors claim that the phase-dependent stimulation effects in SCI rats are immediate, but they don't say how long it takes for these effects to be expressed. Are these effects evident in the response to the first stimulus train, or does it take seconds or minutes for the effects to be expressed? After the initial expression of these effects, are there any gradual changes in the responses over time, e.g., habituation or potentiation?
The effects are immediately expressed at the very first occurrence of stimulation. We never tested a rat completely naïve to stimuli, as each treadmill session involves prior cortical mapping to identify a suitable active site for involvement in locomotor experiments. Yet, as demonstrated in Supplementary Video 1 accompanying our 2021 publication on contralateral effects of cortical stimulation, "Bonizzato and Martinez, Science Translational Medicine," the impact of phase-dependent cortical stimulation on movement modulation is instantaneous and ceases promptly upon discontinuation of the stimulation. We did not quantify potential gradual changes in responsiveness over time, but we cannot exclude that for long stimulation sessions (e.g., 30 min or more), stimulus amplitude may need to be slightly increased over time to compensate habituation.
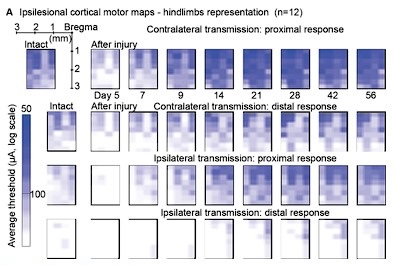
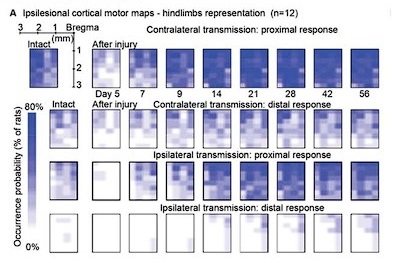
- Awake motor maps (lines 250-277): The analysis of the motor maps appears to be based on measurements of the percentage of channels in which a response can be detected. This analytic approach seems incomplete in that it only assesses the spatial aspect of the cortical drive to the musculature. One channel could have a just-above-threshold response, while another could have a large response; in either case, the two channels would be treated as the same positive result. An additional analysis that takes response intensity into account would add further insight into the data, and might even correlate with the measures of functional recovery. Also, a single stimulation intensity was used; the results may have been different at different stimulus intensities.
We confirm that maps of cortical stimulation responsiveness may vary at different stimulus amplitudes. To establish an objective metric of excitability, we identified 100µA as a reliable stimulation amplitude across rats and used this value to build the ipsilateral motor representation results in Figure 6. This choice allows direct comparison with Figure 6 of our 2021 article, related to contralateral motor representation. The comparison reveals a lack of correlation with functional recovery metrics in the ipsilateral case, in contrast to the successful correlation achieved in the contralateral case.
Regarding the incorporation of stimulation amplitudes into the analysis, as detailed in the Method section (lines 770-771), we systematically tested various stimulation amplitudes to determine the minimal threshold required for eliciting a muscle twitch, identified as the threshold value. This process was conducted for each electrode site. Upon reviewing these data, we considered the possibility of presenting an additional assessment of ipsilateral cortical motor representation based on stimulation thresholds. However, the representation depicted in the figure did not differ significantly from the data presented in Figure 6A. Furthermore, this representation introduced an additional weakness, as it was unclear how to represent the absence of a response in the threshold scale. We chose to arbitrarily designate it as zero on the inverse logarithmic scale, where, for reference, 100 µA is positioned at 0.2 and 50 µA at 0.5.
In conclusion, we believe that the conclusions drawn from this analysis align substantially with those in the text. The addition of the threshold analysis, in our assessment, would not contribute significantly to improving the manuscript.
Author response image 1.
Threshold analysis
Author response image 2.
Original occurrence probability analysis, for comparison.
- Lines 858-860: The authors state that "All tests were one-sided because all hypotheses were strictly defined in the direction of motor improvement." By using the one-sided test, the authors are using a lower standard for assessing statistical significance that the overwhelming majority of studies in this field use. More importantly, ipsilateral stimulation of particular kinds or particular sites might conceivably impair function, and that is ignored if the analysis is confined to detecting improvement. Thus, a two-sided analysis or comparable method should be used. This appropriate change would not greatly modify the authors' current conclusions about improvements.
Our original hypothesis, drawn from previous studies involving cortical stimulation in rats and cats, as well as other neurostimulation research for movement restoration, posited a favorable impact of neurostimulation on movement. Consistent with this hypothesis, we designed our experiments with a focus on enhancing movement, emphasizing a strict direction of improvement.
It's important to note that a one-sided test is the appropriate match for a one-sided hypothesis, and it is not a lower standard in statistics. Each experiment we conducted was constructed around a strictly one-sided hypothesis: the inclusion of an extensor-inducing stimulus would enhance extension, and the inclusion of a flexion-inducing stimulus would enhance flexion. This rationale guided our choice of the appropriate statistical test.
We acknowledge your concern regarding the potential for ipsilateral stimulation to have negative effects on locomotion, which might not be captured when designing experiments based on one-sided hypotheses. This concern is valid, and we will explicitly mention it in the statistics section. Nonetheless, even if such observations were made, they could serve as the basis for triggering an ad-hoc follow-up study.
Reviewer #2 (Public Review):
Summary:
The authors' long-term goals are to understand the utility of precisely phased cortex stimulation regimes on recovery of function after spinal cord injury (SCI). In prior work, the authors explored the effects of contralesion cortex stimulation. Here, they explore ipsilesion cortex stimulation in which the corticospinal fibers that cross at the pyramidal decussation are spared. The authors explore the effects of such stimulation in intact rats and rats with a hemisection lesion at the thoracic level ipsilateral to the stimulated cortex. The appropriately phased microstimulation enhances contralateral flexion and ipsilateral extension, presumably through lumbar spinal cord crossed-extension interneuron systems. This microstimulation improves weight bearing in the ipsilesion hindlimb soon after injury, before any normal recovery of function would be seen. The contralateral homologous cortex can be lesioned in intact rats without impacting the microstimulation effect on flexion and extension during gait. In two rats ipsilateral flexion responses are noted, but these are not clearly demonstrated to be independent of the contralateral homologous cortex remaining intact.
Strengths:
This paper adds to prior data on cortical microstimulation by the laboratory in interesting ways. First, the strong effects of the spared crossed fibers from the ipsi-lesional cortex in parts of the ipsi-lesion leg's step cycle and weight support function are solidly demonstrated. This raises the interesting possibility that stimulating the contra-lesion cortex as reported previously may execute some of its effects through callosal coordination with the ipsi-lesion cortex tested here. This is not fully discussed by the authors but may represent a significant aspect of these data. The authors demonstrate solidly that ablation of the contra-lesional cortex does not impede the effects reported here. I believe this has not been shown for the contra-lesional cortex microstimulation effects reported earlier, but I may be wrong. Effects and neuroprosthetic control of these effects are explored well in the ipsi-lesion cortex tests here.
In the revised version of the manuscript, we will incorporate various text improvements to address the points you have highlighted below. Additionally, we will integrate the suggested discussion topic on callosal coordination related to contralateral cortical stimulation.
Weaknesses:
Some data is based on very few rats. For example (N=2) for ipsilateral flexion effects of microstimulation. N=3 for homologous cortex ablation, and only ipsi extension is tested it seems. There is no explicit demonstration that the ipsilateral flexion effects in only 2 rats reported can survive the contra-lateral cortex ablation. We agree with this assessment. The ipsilateral flexion representation is here reported as a rare but consistent phenomenon, which we believe to have robustly described with Figure 7 experiments. We will underline in the text that the ablation experiment did not conclude on the unilateral-cortical nature of ipsilateral flexion effects.
Some improvements in clarity and precision of descriptions are needed, as well as fuller definitions of terms and algorithms.
Likely Impacts: This data adds in significant ways to prior work by the authors, and an understanding of how phased stimulation in cortical neuroprosthetics may aid in recovery of function after SCI, especially if a few ambiguities in writing and interpretation are fully resolved.
The manuscript text will be revised in its final version, and we seek to eliminate any ambiguity in writing, data interpretation and algorithms.
Reviewer #3 (Public Review):
Summary:
This article aims to investigate the impact of neuroprosthesis (intracortical microstimulation) implanted unilaterally on the lesion side in the context of locomotor recovery following unilateral thoracic spinal cord injury.
Strength:
The study reveals that stimulating the left motor cortex, on the same side as the lesion, not only activates the expected right (contralateral) muscle activity but also influences unexpected muscle activity on the left (ipsilateral) side. These muscle activities resulted in a substantial enhancement in lift during the swing phase of the contralateral limb and improved trunk-limb support for the ipsilateral limb. They used different experimental and stimulation conditions to show the ipsilateral limb control evoked by the stimulation. This outcome holds significance, shedding light on the engagement of the "contralateral projecting" corticospinal tract in activating not only the contralateral but also the ipsilateral spinal network.
The experimental design and findings align with the investigation of the stimulation effect of contralateral projecting corticospinal tracts. They carefully examined the recovery of ipsilateral limb control with motor maps. They also tested the effective sites of cortical stimulation. The study successfully demonstrates the impact of electrical stimulation on the contralateral projecting neurons on ipsilateral limb control during locomotion, as well as identifying important stimulation spots for such an effect. These results contribute to our understanding of how these neurons influence bilateral spinal circuitry. The study's findings contribute valuable insights to the broader neuroscience and rehabilitation communities.
Thank you for your assessment of this manuscript. The final version of the manuscript will incorporate your suggestions for improving term clarity and will also enhance the discussion on the mechanism of spinal network engagement, as outlined below.
Weakness:
The term "ipsilateral" lacks a clear definition in the title, abstract, introduction, and discussion, potentially causing confusion for the reader. In the next revision of the manuscript, we will provide a clear definition of the term "ipsilateral."
The unexpected ipsilateral (left) muscle activity is most likely due to the left corticospinal neurons recruiting not only the right spinal network but also the left spinal network. This is probably due to the joint efforts of the neuroprosthesis and activation of spinal motor networks which work bilaterally at the spinal level. However, in my opinion, readers can easily link the ipsilateral cortical network to the ipsilateral-projecting corticospinal tract, which is less likely to play a role in ipsilateral limb control in this study since this tract is disrupted by the thoracic spinal injury.
We agree with your assessment. The discussion section paragraph presenting putative mechanisms of cortico-spinal transmission in the effects presented in the results will be enhanced to reflect these suggestions.



