Peer review process
Revised: This Reviewed Preprint has been revised by the authors in response to the previous round of peer review; the eLife assessment and the public reviews have been updated where necessary by the editors and peer reviewers.
Read more about eLife’s peer review process.Editors
- Reviewing EditorBrice BathellierCentre National de la Recherche Scientifique, Paris, France
- Senior EditorPanayiota PoiraziFORTH Institute of Molecular Biology and Biotechnology, Heraklion, Greece
Reviewer #1 (Public review):
The propagation of electrical signals within neuronal circuits is tightly regulated by the physical and molecular properties of neurons. Since neurons vary in size across species, the question arises whether propagation speed also varies to compensate for it. The present article compares numerous speed-related properties in human and rat neurons. They found that the larger size of human neurons seems to be compensated by a faster propagation within dendrites but not axons of these neurons. The faster dendritic signal propagation was found to arise from wider dendritic diameters and greater conductance load in human neurons. In addition, the article provides a careful characterization of human dendrites and axons, as the field has only recently begun to characterize post-operative human cells. There are only a few studies reporting dendritic properties and these are not all consistent, hence there is added value of reporting these findings, particularly given that the characterization is condensed in a compartmental model.
Strengths
The study was performed with great care using standard techniques in slice electrophysiology (pharmacological manipulation with somatic patch-clamp) as well as some challenging ones (axonal and dendritic patch-clamp). Modeling was used to parse out the role of different features in regulating dendritic propagation speed. The finding that propagation speed varies across species is novel as previous studies did not find a large change in membrane time constant nor axonal diameters (a significant parameter affecting speed). A number of possible, yet less likely factors were carefully tested (Ih, membrane capacitance). The main features outlined here are well known to regulate speed in neuronal processes. The modeling was also carefully done to verify that the magnitude of the effects is consistent with the difference in biophysical properties. Hence, the findings appear very solid to me.
Weaknesses
The role of diameter in regulating propagation speed is well known in the axon literature.
Comment on the revised version: the authors have now made clearer that the role of diameter was well known in the manuscript.
Reviewer #2 (Public review):
Summary:
In this paper, Oláh and colleagues introduce new research data on the cellular and biophysical elements involved in transmission within the pyramidal circuits of the human neocortex. They gathered a comprehensive set of patch-clamp recordings from human and rat pyramidal neurons to compare how the temporal aspect of neuronal processing is maintained in the larger human neocortex. A range of experimental techniques have been used, including two-photon guided dual whole-cell recordings, electron microscopy, complemented by theoretical and computational methods.
The authors find that synaptically connected pyramidal neurons within the human neocortex have longer intercellular path lengths. They go on to show that the short soma to soma latencies is not due to propagation velocity along the axon but instead reflects a higher propagation speed of synaptic potentials from dendrite to soma. Next, in a series of extensive computational modeling studies focusing on the synaptic potentials, the authors show that the shorter latency may be explained by larger diameters, affecting the cable properties and resulting is relatively faster propagation of EPSPs in the human neuron. The manuscript is well-written, and the physiological experiments and in-depth theoretical steps for the simulations are clear. Whether passive cable properties of the dendrites alone are responsible for higher velocities remains to be further investigated. Based on the present data the contribution of active membrane properties cannot be excluded.
Strengths:
The authors used complex 2P-guided dual whole-cell recordings in human neurons. In combination with detailed reconstructions, these approaches represent the next steps in unravelling the information processing in human circuits.
The computational modelling and cable theory application to the experimentally constrained simulations provide an integrated view of the passive membrane properties of human neurons.
Weaknesses:
There are concerns with the statistical analyses of the experimental data. The two-way analyses are not supporting that the backpropagation speed in human neurons is more affected by TTX-induced or after ZD remains higher. Significance of interaction is required, and the authors make errors in the interpretation and application of separate additional t-tests. Whether the cable properties alone are the main explanation for speeding the electrical signaling in human pyramidal neurons deserves further studies.
Comments on the latest version:
In my previous review I suggested the author read upon the need to perform two-way ANOVA for their experiments. Although I am glad this has now been done, I'm surprised to read the interpretation remains flawed and we are not provided with all the analyses. We need to know all the covariates and results of the post-hoc comparisons. What is written in the results is incomplete.
One cannot perform two-way ANOVA and subsequently performing t-tests on computed differences. Figures 3C and F are irrelevant. Instead, we need to know the Bonferroni post-hoc results for all comparisons.
If there is an interaction significance then the authors will have to conclude the Na+ channels have a larger contribution to the backpropagation.
Line187 "It therefore be argued that HCN channels may contribute to the higher conduction velocities in human dendrites, but do not by themselves explain the differences between the two species."
One wonders whether supplementary figures are required.
Reviewer #3 (Public review):
Summary:
This study indicates that connections across human cortical pyramidal cells have identical latencies despite a larger mean dendritic and axonal length between somas in human cortex. A precise demonstration combining detailed electrophysiology and modeling, indicates that this property is due to faster propagation of signals in proximal human dendrites. This faster propagation is itself due to a slightly thicker dendrite, to a larger capacitive load, and to stronger hyperpolarizing currents. Hence, the biophysical properties of human pyramidal cells are adapted such that they do not compromise information transfer speed.
Strengths:
The manuscript is clear and very detailed. The authors have experimentally verified a large number of aspects that could affect propagation speed and have pinpointed the most important one. This paper provides an excellent comparision of biophysical properties between rat and human pyramidal cells. Thanks to this approach a comprehensive description of the mechanisms underlying the acceleration of propagation in human dendrite is provided.
Weaknesses:
The weaknesses I had identified have been addressed by the authors.
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public Review):
The propagation of electrical signals within neuronal circuits is tightly regulated by the physical and molecular properties of neurons. Since neurons vary in size across species, the question arises whether propagation speed also varies to compensate for it. The present article compares numerous speed-related properties in human and rat neurons. They found that the larger size of human neurons seems to be compensated by a faster propagation within dendrites but not the axons of these neurons. The faster dendritic signal propagation was found to arise from wider dendritic diameters and greater conductance load in human neurons. In addition, the article provides a careful characterization of human dendrites and axons, as the field has only recently begun to characterize post-operative human cells. There are only a few studies reporting dendritic properties and these are not all consistent, hence there is the added value of reporting these findings, particularly given that the characterization is condensed in a compartmental model.
Strengths:
The study was performed with great care using standard techniques in slice electrophysiology (pharmacological manipulation with somatic patch-clamp) as well as some challenging ones (axonal and dendritic patch-clamp). Modeling was used to parse out the role of different features in regulating dendritic propagation speed. The finding that propagation speed varies across species is novel as previous studies did not find a large change in membrane time constant or axonal diameters (a significant parameter affecting speed). A number of possible, yet less likely factors were carefully tested (Ih, membrane capacitance). The main features outlined here are well-known to regulate speed in neuronal processes. The modeling was also carefully done to verify that the magnitude of the effects is consistent with the difference in biophysical properties. Hence, the findings appear very solid to me.
Weaknesses:
The role of diameter in regulating propagation speed is well-known in the axon literature.
We thank the reviewer for this comment. This is indeed true. The paper does not claim that this is new – we just refereed to Waxman’s book to acknowledge this established effect. Our main emphasize is on the impact of dendritic (rather than axonal) diameter – highlighting the faster EPSP speed near the input synapse and converging to steady-state value further away from the soma and using this to explore the impact of differences in dendritic diameter of rat vs. human on EPSP latency and velocity. We now made this point clearer in the revised text.
Reviewer #2 (Public Review):
Summary:
In this paper, Oláh and colleagues introduce new research data on the cellular and biophysical elements involved in transmission within the pyramidal circuits of the human neocortex. They gathered a comprehensive set of patch-clamp recordings from human and rat pyramidal neurons to compare how the temporal aspect of neuronal processing is maintained in the larger human neocortex. A broad range of experimental, theoretical, and computational methods are used, including two-photon guided dual whole-cell recordings, electron microscopy, and computational simulations of reconstructed neurons.
Recordings from synaptically connected pyramidal neurons revealed longer intercellular path lengths within the human neocortex. Further, by using dual whole-cell recordings from somadendrite and soma-axon locations, they found that short latencies from soma to soma can be partly attributed to an increased propagation speed for synaptic potentials, but not for the propagation of action potentials along the axon.
Next, in a series of extensive computational modeling studies focusing on the synaptic potentials, the authors observe that the short-latency within large human pyramidal neural circuits may have a passive origin. For a wide array of local synaptic input sites, the authors show that the conductance load of the dendrites, electrically coupled to a large diameter apical dendrite, affects the cable properties. The result is a relatively faster propagation of EPSPs in the human neuron.
The manuscript is well-written and the physiological experiments and biophysical arguments are very well explained. I appreciated the in-depth theoretical steps for the simulations. That passive cable properties of the dendrites are causing a higher velocity in human dendrites is interesting but there is a disconnect between the experimental findings and the model simulations. Based on the present data the contribution of active membrane properties cannot be dismissed and deserves further experiments.
See our response below
Strengths:
The authors present state-of-the-art 2P-guided dual whole-cell recordings in human neurons. In combination with detailed reconstructions, these approaches represent the next steps in unravelling the information processing in human circuits.
The computational modeling based on cable theory and experimentally constrained simulations provides an excellent integrated view of the passive membrane properties.
Weaknesses:
There are smaller and larger issues with the statistical analyses of the experimental data which muddles the interim conclusions.
That the cable properties alone are the main explanation for speeding the electrical signaling in human pyramidal neurons appears inconsistent with the experimental data.
This is an excellent point – we indeed performed analysis on only passive cases – highlighting (and now also ranking) the impact of the various morpho-electrical properties of the neurons on the differences in signal latency in human vs. rats. We did explored (not shown) the effect of active channels in the dendrites (including the h-current); as expected the results strongly depend on channel density and their spatial distribution over the dendritic tree. As we do not know these parameters for the modelled cells, we decided to remain focus on the impact of passive/morphological parameters. We also note that the experimental results (page 4-5 in manuscript) show minor contribution of h-current emphasizing that the passive properties have the main role in differentiating human and rats. differences between human and rat.
Some of the electrophysiological experiments require further control experiments to make robust conclusions.
Reviewer #3 (Public Review):
Summary:
This study indicates that connections across human cortical pyramidal cells have identical latencies despite a larger mean dendritic and axonal length between somas in the human cortex. A precise demonstration combining detailed electrophysiology and modeling indicates that this property is due to faster propagation of signals in proximal human dendrites. This faster propagation is itself due to a slightly thicker dendrite, a larger capacitive load, and stronger hyperpolarizing currents. Hence, the biophysical properties of human pyramidal cells are adapted such that they do not compromise information transfer speed.
Strengths:
The manuscript is clear and very detailed. The authors have experimentally verified a large number of aspects that could affect propagation speed and have pinpointed the most important one. This paper provides an excellent comparison of biophysical properties between rat and human pyramidal cells. Thanks to this approach a comprehensive description of the mechanisms underlying the acceleration of propagation in human dendrite is provided.
Weaknesses:
Several aspects having an impact on propagation speed are highlighted (dendritic diameter, ionic channels, capacitive load) and there is no clear ranking of their impact on signal propagation speed. It seems that the capacitive load plays a major role, much more than dendritic diameter for which only a 10% increase is observed across species. Both aspects actually indicate that there is an increase in passive signal propagation speed with bigger cells at least close to the soma. This suggests that bigger cells are mechanically more rapid. An intuitive reason why capacitive load increases speed would also help the reader follow the demonstration.
We thank the referee for both these excellent points. In response to them:
(i) We now performed a new comprehensive statistical analysis and show the ranking of the effect of the different morphological/cable factors on EPSP propagation. This analysis appears in both Supp. Table 5& 6, Fig. S16 and also in the main text as follows:
To rank the impact of the various factors affecting EPSP propagation latency in human and rat neurons, we conducted a comprehensive statistical analysis using two complementary approaches: the generalized linear model (GLM) (Kiebel & Holmes, 2007) as well as SHAP (SHapley Additive exPlanations) (Lundberg & Lee, 2017) based on fitting Gradient Tree Boosting (Friedman, 2002)model. We began by fitting a GLM without interaction terms among the factors affecting EPSP latency (Suppl. Table 5). This enables us to quantify the primary individual factors affecting EPSP propagation. Our analysis revealed the following ranking order: 1) physical distance of synapses from soma had the strongest effect; 2) species differences; 3) conductance load, as demonstrated by our “hybrid cells” manipulation; 4) radii of the apical dendrite, affecting the cables’ space constant, λ; and 5) the specific cable parameters, as revealed when using per-cell fitted parameters versus uniform cable parameters, was minimal. We next performed GLM analysis with interaction terms showing that, as expected, there are significant interactions between the factors affecting EPSP latency (Suppl. Table 6). To further validate the above ranking while incorporating the interactions between the various factors affecting EPSP latency, we performed a SHAP analysis. Notably, even with interactions included, the ranking of the factors affecting signal propagation are aligned with the results from the analysis based on the GLM without interaction terms (see Fig S.16).
(ii) As for the intuitive explanation required by the referee. We added the following paragraph In the Discussion:
The intuitive reason for this enhancement is that the large conductance load (the “leaky end” boundary conditions) more effectively “steals” the synaptic (axial) current (like water pouring faster into a large pool). The more mathematical intuition is that the large soma (sink) adds fast time constants to the system (see also related explanation in Fig. 4 in Eyal et al., 2014).
We thank the editors for considering and revising our manuscript for publication in eLife. We appreciate the positive appreciation of the work and the critical points raised by the reviewers. We have responded in detail to all the excellent comments from all reviewers. We believe that these revisions have significantly improved the quality of our study.
Recommendations for the authors:
Reviewer #1 (Recommendations For The Authors):
There are two points that could improve the reading experience of this nice manuscript. These should be easily addressed with minor re-phrasing.
Credit to conduction velocity literature. Less widely known in the dendrite literature, in the axon literature, the relationship between propagation speed and process diameter is well established. I thought the two articles cited (Jack Noble Tsien and Agmon-Snir & Segev) were not as direct in the treatment of this relationship. The work of Stephen Waxman, for instance, made clear how axon diameter tightly controls propagation speed (see for instance the Scholarpedia entry by Swadlow and Waxman). In my opinion, this is a widely known piece of work, that is part of some introductory books to neuroscience. While the article does not claim they found this relationship, parts of the presentation are better understood if we ignore this well-known fact. I am referring to the abstract, intro, and the beginning of results where 'larger' is presented as synonymous with 'slower'. For instance 'to compensate for the increase neurons' size' (abstract) or 'the increase in size of dendrites and axons might come with a cost of longer signal propagation times' only makes sense if 'size' refers to spatial extent, not diameter.
We thank for this valid point; leaving out axon diameter references was not intentional. We have now added the suggested reference to our manuscript. In the size comparisons, we have only pointed out the obvious size differences between the body and the dendritic processes. We have reworded sentences with size comparisons.
In Abstract (lines 1-6):
Human-specific cognitive abilities depend on information processing in the cerebral cortex, where neurons are significantly larger, their processes are longer and sparser compared to rodents. We found that, in synaptically-connected layer 2/3 pyramidal cells (L2/3 PCs), soma-tosoma signal propagation delay is similar in humans and rodents. Thus, to compensate for the increase in neurons’s longer processes, membrane potential changes must propagate faster in human axons and/or dendrites.
In section “Effect of dendritic thickness” in Results we have modified it as follows:
The relationship between conduction velocity and axon diameter is well known for small myelinated and unmyelinated axons (Waxman and Bennett, 1972). Anatomical features of neuronal processes dendrites also have a major influence on signal propagation properties 5,19, thus …
Waxman, S. G. and Bennett, M. V. L. Relative conduction velocity of small myelinated and nonmyelinated fibres in the central nervous system. Nature New Biol., 238217-219, 1972.
Two or four dendritic factors? The study identifies two major dendritic factors influencing the propagation speed (diameter and load), however the end of the results highlights four factors. I did not understand how factor 2 was different than factor 1. Neither did I understand how factor 4 was different from the other factors. There seemed to be a little redundancy here that could be streamlined.
We thank the reviewer for pointing this out. We now have changes the respective text, added the ranking statistics (see above) to assess the effect of the different parameters on signal propagation in dendrites.
Microcircuits? The study found that the changes in speed arise from the dendrites rather than the axons, as such it seems it would be more precise to replace 'microcircuits' with 'dendrites'.
We are thankful for this suggestion. We change the title to Accelerated signal propagation speed in human neocortical dendrites.
Typos
P3 line 24 'find significant difference the propagation'.
P6 line 35 'how morphological differences' it would be useful to specify which morphological difference here.
Corrected.
Reviewer #2 (Recommendations For The Authors):
(1) The statistical analyses should be changed. T-testing populations and comparing visual differences of differences ("human minus rats") is a common but egregious error in the field of neurosciences (see doi:10.1038/nn.2886). The conclusion that HCN channels "... do not by themselves explained the differences between the two species" (lines 174-176) is not compelling. The design of the experiments presented in Figure 3 is paired recordings and the addition of a blocker (ZD7288 or TTX cocktail). These are classic 2 x 2 factorial designs (species x drug). The authors will need to perform a repeated-measured analysis of variance (RM-ANOVA) and provide information on the interaction significance. Please revise the figures and improve statistical reporting. Post-hoc comparisons of the velocity populations are required to support the idea of whether h-channels are explaining the observed differences.
Thank you for drawing our attention to this error. The statistical analysis of the pharmacological experiments was re-performed as suggested. After the 2-way ANOVA with repeated measures and Bonferroni post-hoc correction, we can indeed find significant differences only in the control group, namely that the propagation speed of bAPs in human dendrites was significantly higher. The implementation of the proposed statistical analysis demonstrates that the administration of ZD has no statistically significant effect on the propagation speed of human or rat dendrites. The treatment with TTX cocktail resulted in a significant difference in signal propagation in humans but not in rodents. However the trend is discernible and the P = 0.0588 value is close to the widely accepted 0.05 threshold. After the TTX cocktail treatment, the speed of signal propagation did not differ significantly between the two species. However, on average, the human dendrites remained faster. These alterations in P-values do not affect our primary conclusions. The MS text has been modified accordingly.
(2) Although ZD7288, in my opinion, influences the bAP (see point #1) the authors subsequently leave the h-current unblocked in the experiments in Figures 3D, E. Here, they use sodium, potassium, and calcium currents as well as synaptic conductances. I am puzzled why (in line 188) they claim the dendrites are "passive" although the data show h-currents are contributing to the shape of the bAP in human neurons. In line 196 they conclude voltage-gated conductances have a "minor" contribution and passive properties a main role. Please revise conclusions or provide better experimental support.
Thank you for this point. We meant to refer to the state in which no action potential can be generated, although the word 'passive' might be misleading in this context; we rephrase these sentences in the MS accordingly.
(3) A major concern is the injection of an AP in voltage-clamp mode. Although this is the right choice and I'm in support of the experiment, it is technically challenging to space clamp the soma and fully recapitulate the speed and amplitude of a 100 mV depolarization. The voltage drop in peak amplitude as well as the increased delay between the baseline AP (current clamp) and AP in blocker conditions (voltage clamp) could be fully explained by switching between current- and voltage-clamp modes. In additional control experiments, the authors should add a second voltage follower electrode (CC) at the soma showing whether the authors can preserve the original AP (from CC) in VC/blocker condition. It may well be they need to adjust the injection protocol.
Our experiments were designed to replicate the work of Stuart et al. (1994), in which they compared the attenuation of active and passive backpropagating signals. When they blocked Na+ channels with TTX they injected simulated action potentials in voltage-clamp mode. They concluded that TTX-sensitive Na+ channels cause somatic action potential entry into the dendritic compartment. They found a comparable attenuation of the backward propagating action potential in the dendrites in control conditions (~70 %).
We performed control recordings based on the reviewer’s suggestion (Author response image 1).
Author response image 1.
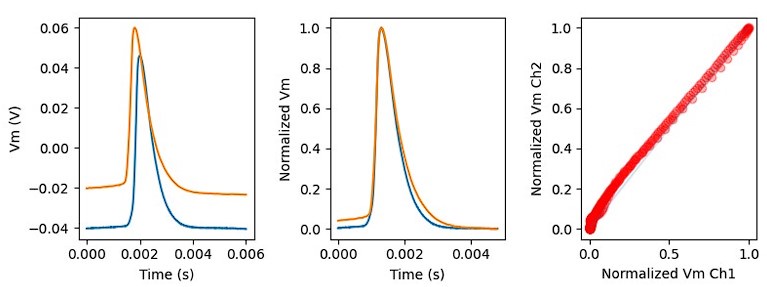
Injection of the previously recorded AP (blue) in VC mode produced a completely similar somatic AP in CC mode (orange). The slight temporal delay between the two signal caused by the different position of the pipettes on the cell body. The right panel shows the plot of the two peak-aligned APs as a function of each other, close to the blue ‘equality’ line. We concluded that the original AP is well preserved in VC/blocker condition.
(5) From the paragraph entitled "Modeling EPSP propagation in dendrites" and onwards the authors make countless conclusions based on theory and modelling results but without any statistical support. Multiple neurons are used thus it is rather straightforward to provide numerical support for the assertions. For example, but this is not an exhaustive list, how should we interpret that latency ranges are different (line 240, line 253) etc.? Or were the estimated Cm values of human and rat neurons (0.6 versus 1.1) significantly different? And if so, how does this align with the Cm estimates in the nucleated patch experiments?
We thank the referee for this comment and now added a set of statistical analyses. The results appear now throughout the whole theoretical paper in revised article. In particular with respect to Figs. 6&7 where we now show that, indeed, our various manipulations (e.g., hybrid vs. original cells) as well as the cable parameters (Cm, Rm) are indeed significantly different between human and rats whereas the membrane time constant is not significantly different between human and rat. As for Cm in human. Our limited sample size shows significant difference between human and rat. Yet, the range of values for Cm that we found in our modeling study does fall within the experimental range reported in the present study.
Minor
Line 44. The "simulated EPSP" example in Figure 2C is not a command waveform for an EPSC. Line 526 in the methods states that also ramp currents were used. Please revise to clarify the main text.
Thank you for bringing this discrepancy to our attention. In the experiments, we used ramp injections. We have made this clear in the main text as follows: ”... we tested orthodromic or forward propagating signal propagation velocity by injecting short-duration current ramps to simulate EPSP (sEPSP) signals in the dendrites and recorded the resultant subthreshold voltage response in the soma”
Line 522. The authors state the recordings were all carried out "in current clamp mode" but detailed VC method information is lacking. Did they use series resistance compensation?
We did not use series resistance compensation.
Line 479 From which region(s) where human "neocortical slices" sampled? Please add this information.
We have added regions of origin to the Methods section: frontal (n = 21), temporal (n = 20), parietal (n = 20), and occipital (n = 1).
Please show higher temporal resolution example traces, for example in Figure 3. Differences are at the micrometer scale, but APs are shown at the millisecond scale. Hard to judge the quality of the data. Showing the command potentials (inset Figure 3D, E) is misleading (see major point #3).
In response to the reviewer's request, we have redrawn the example traces in Figure 3.
Please check the labeling of figures. There is information missing. For example, in Figure 5 A to C I am missing information and the units of the axes.
In the black plots on the right side of panels B and C, the y-axis shows the thickness measurements for the given dendrite stacked on top of each other and the x-axis shows the measurement values, the units for the x-axis are µm as mentioned in the figure legend.
Line 981 "scalebars" should read scale bars."
Line 986 "bootstraped" should read "bootstrapped".
Done.
Are the dendritic diameters increased for all basal and apical higher-order branches? It is unclear how the model simulations were built on diameters of primary and higher-order branches.
In our modelling study we took the actual diameter of the reconstructed PCs in both proximal and higher order branches. We did compare per-distance differences in diameter – but it is automatically incorporated into the computation of the basal load (“equivalent cables” in Figs 6&8).
The velocity calculation for axonal propagation (yielding a ~0.9 m/s conduction velocity, Figure 2B) is incorrect. Using the peak of the action potentials between soma and axon misses the fact that action potentials start earlier and spatially distally from the soma in the axon. Please revise the calculation to include the temporal delay and actual distance travelled by the forward propagating action potential.
Thank you for this question. We are aware that the AP is generated at the AIS and that it is located between the two recording electrodes and we have to take into account that the signal propagates from the AIS to the soma and this may shorten the delay in the system. To the best of our knowledge, there is no experimental evidence of the location of the AP generation site on the AIS in layer 2-3 pyramidal cells in the human neocortex, so we assumed that it is located 35 microns from the soma, and that the propagation speed from the AIS to the two directions is the same. Consequently, we have corrected our propagation velocity values as follows:
“For the axon bleb recordings we assumed that the axon initial segment (AIS) of the cells are 35 µm from the axon hillock, and the APs propagate to forward (to the bleb) and backward (to the soma) at the same speed. For the correction of the AIS we used the following formula: (2)
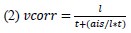
where vcorr is the corrected propagation speed for AIS position, l is the axonal distance between the soma and the axon bleb, t is the latency between the two measuring point, ais is the assumed position of the AIS alongside the axon (35 µm).”
What explains the strongly attenuated axonal action potential at the bleb? Is this representative?
The strongly attenuated axonal action potential at the bleb can be explained by a few key factors:
(1) Membrane Integrity: Bleb formation often indicates some level of membrane damage or alteration. This can disrupt the normal ionic gradients across the membrane, leading to a failure in generating or propagating action potentials effectively.
(2) Current Leakage: Bleb formation may create additional pathways for ion leakage, which can dissipate the electrical current that would normally propagate the action potential. This leakage reduces the overall amplitude of the action potential.
Line 275 "To our delight", please rephrase.
Corrected.
Reviewer #3 (Recommendations For The Authors):
- In Figure 1, the number of cells used to assess intersomatic distance is quite low. A larger number of neuron pairs should be analyzed to be more representative. Or at least an explanation of why such a low sampling can be conclusive.
We appreciate the reviewer’s concerns on sample sizes of the first set of experiments, where the anatomical pathways were measured through the synapses of coupled cells with electrophysiological recordings. We acknowledge that this is a limitation of our study. However, in this series of experiments, we simply wanted to experimentally confirm already known results which consisted of two parts: first, that in humans the dendrites and axons of neurons are longer, and second, that they have the same time delay in terms of synaptic latency.
The reported similarity in synaptic latencies is consistent with the results of a recent study by Campagnola et al. (2022) showing that EPSP latencies of local connections between layer 2/3 pyramidal cells are in the same range in humans and mice (human median latency = 1.73 ms vs. mouse median latency = 1.49 ms). We came to the same conclusion in our previous work where we compared pyramidal basket cell synaptically coupled pairs in human and rat pairs (Molnár et al. 2016).
On the other hand, we report interspecific differences in cable pathways from soma to soma, again consistent with the literature suggesting that the length of pyramidal neural processes is longer in humans than in rodents (see Supplementary Figure 1 and e.g. Berg et al. 2021).
From a practical point of view the collection of experimental data in this hard won experiment is particularly difficult. The electrophysiological recording of a connected pair with an appropriate pre- and postsynaptic series resistance, where human tissue samples are limited, is the first step here. To obtain information about the path of the signals between pre- and postsynaptic cells, an anatomical reconstruction is required. This requires a) a high-quality recovery of postsynaptic dendrites and presynaptic axons, b) successful tracing of all potential contact points between presynaptic axons and postsynaptic dendrites back to the pre- and postsynaptic soma. The difficulty of the latter point in particular arises from the fact that parts of the presynaptic axonal arbor are myelinated and the success of biocytin-based tracing depends on the length of the myelinated axon branches. The success/failure of complete axonal tracing only becomes apparent at the end of these efforts.
- The author should provide an intuitive explanation of why capacitive load accelerates propagation in the dendrite.
See answer above
- The author should more clearly rank the contribution of each difference between rat and human neurons. The 10% increase in dendritic diameter which affects velocity only via a square root seems a very weak contribution. This should be clarified.
We now added a set of statistical methods to perform such a ranking in the theoretical part of this study, as described above (and in a new paragraph, attached above) in the revised article.
References
Eyal, G., Mansvelder, H. D., de Kock, C. P. J., & Segev, I. (2014). Dendrites impact the encoding capabilities of the axon. Journal of Neuroscience, 34(24), 8063–8071. https://doi.org/10.1523/JNEUROSCI.5431-13.2014
Friedman, J. H. (2002). Stochastic gradient boosting. In Computational Statistics & Data Analysis (Vol. 38). www.elsevier.com/locate/csda
Kiebel, S. J., & Holmes, A. P. (2007). The General Linear Model. In K. Friston, J. Ashburner, S. Kiebel, T. Nichols, & P. William (Eds.), Statistical Parametric Mapping (pp. 101–125). Academic Press.
Lundberg, S. M., & Lee, S.-I. (2017). A unified approach to interpreting model predictions. Proceedings of the 31st International Conference on Neural Information Processing Systems, 4768–4777.



