Peer review process
Not revised: This Reviewed Preprint includes the authors’ original preprint (without revision), an eLife assessment, public reviews, and a provisional response from the authors.
Read more about eLife’s peer review process.Editors
- Reviewing EditorAndrew WestDuke University, Durham, United States of America
- Senior EditorAmy AndreottiIowa State University, Ames, United States of America
Reviewer #1 (Public Review):
Summary:
The Roco proteins are a family of GTPases characterized by the conserved presence of an ROC-COR tandem domain. How GTP binding alters the structure and activity of Roco proteins remains unclear. In this study, Galicia C et al. took advantage of conformation-specific nanobodies to trap CtRoco, a bacterial Roco, in an active monomeric state and determined its high-resolution structure by cryo-EM. This study, in combination with the previous inactive dimeric CtRoco, revealed the molecular basis of CtRoco activation through GTP-binding and dimer-to-monomer transition.
Strengths:
The reviewer is impressed by the authors' deep understanding of the CtRoco protein. Capturing Roco proteins in a GTP-bound state is a major breakthrough in the mechanistic understanding of the activation mechanism of Roco proteins and shows similarity with the activation mechanism of LRRK2, a key molecule in Parkinson's disease. Furthermore, the methodology the authors used in this manuscript - using conformation-specific nanobodies to trap the active conformation, which is otherwise flexible and resistant to single-particle average - is highly valuable and inspiring.
Weakness:
Though written with good clarity, the paper will benefit from some clarifications.
1. The angular distribution of particles for the 3D reconstructions should be provided (Figure 1 - Sup. 1 & Sup. 2).
2. The B-factors for protein and ligand of the model, Map sharpening factor, and molprobity score should be provided (Table 1).
3. A supplemental Figure to Figure 2B, illustrating how a0-helix interacts with COR-A&LRR before and after GTP binding in atomic details, will be helpful for the readers to understand the critical role of a0-helix during CtRoco activation.
4. For the following statement, "On the other hand, only relatively small changes are observed in the orientation of the Roc a3 helix. This helix, which was previously suggested to be an important element in the activation of LRRK2 (Kalogeropulou et al., 2022), is located at the interface of the Roc and CORB domains and harbors the residues H554 and Y558, orthologous to the LRRK2 PD mutation sites N1337 and R1441, respectively."
It is not surprising the a3-helix of the ROC domain only has small changes when the ROC domain is aligned (Figure 2E). However, in the study by Zhu et al (DOI: 10.1126/science.adi9926), it was shown that a3-helix has a "see-saw" motion when the COR-B domain is aligned. Is this motion conserved in CtRoco from inactive to active state?
5. A supplemental figure showing the positions of and distances between NbRoco1 K91 and Roc K443, K583, and K611 would help the following statement. "Also multiple crosslinks between the Nbs and CtRoco, as well as between both nanobodies were found. ... NbRoco1-K69 also forms crosslinks with two lysines within the Roc domain (K583 and K611), and NbRoco1-K91 is crosslinked to K583".
6. It would be informative to show the position of CtRoco-L487 in the NF and GTP-bound state and comment on why this mutation favors GTP hydrolysis.
Reviewer #2 (Public Review):
Summary
The manuscript by Galicia et al describes the structure of the bacterial GTPyS-bound CtRoco protein in the presence of nanobodies. The major relevance of this study is in the fact that the CtRoco protein is a homolog of the human LRRK2 protein with mutations that are associated with Parkinson's disease. The structure and activation mechanisms of these proteins are very complex and not well understood. Especially lacking is a structure of the protein in the GTP-bound state. Previously the authors have shown that two conformational nanobodies can be used to bring/stabilize the protein in a monomer-GTPyS-bound state. In this manuscript, the authors use these nanobodies to obtain the GTPyS-bound structure and importantly discuss their results in the context of the mammalian LRRK2 activation mechanism and mutations leading to Parkinson's disease. The work is well performed and clearly described. In general, the conclusions on the structure are reasonable and well-discussed in the context of the LRRK2 activation mechanism.
Strengths:
The strong points are the innovative use of nanobodies to stabilize the otherwise flexible protein and the new GTPyS-bound structure that helps enormously in understanding the activation cycle of these proteins.
Weakness:
The strong point of the use of nanobodies is also a potential weak point; these nanobodies may have induced some conformational changes in a part of the protein that will not be present in a GTPyS-bound protein in the absence of nanobodies.
Two major points need further attention.
1. Several parts of the protein are very flexible during the monomer-dimer activity cycle. This flexibility is crucial for protein function, but obviously hampers structure resolution. Forced experiments to reduce flexibility may allow better structure resolution, but at the same time may impede the activation cycle. Therefore, careful experiments and interpretation are very critical for this type of work. This especially relates to the influence of the nanobodies on the structure that may not occur during the "normal" monomer-dimer activation cycle in the absence of the nanobodies (see also point 2). So what is the evidence that the nanobody-bound GTPyS-bound state is biochemically a reliable representative of the "normal" GTP-bound state in the absence of nanobodies, and therefore the obtained structure can be confidentially used to interpret the activation mechanism as done in the manuscript.
2. The obtained structure with two nanobodies reveals that the nanobodies NbRoco1 and NbRoco2 bind to parts of the protein by which a dimer is impossible, respectively to a0-helix of the linker between Roc-COR and LRR, and to the cavity of the LRR that in the dimer binds to the dimerizing domain CORB. It is likely the open monomer GTP-bound structure is recognized by the nanobodies in the camelid, suggesting that overall the open monomer structure is a true GTP-bound state. However, it is also likely that the binding energy of the nanobody is used to stabilize the monomer structure. It is not automatically obvious that in the details the obtained nonobody-Roco-GTPyS structure will be identical to the "normal" Roco-GTPyS structure. What is the influence of nanobody-binding on the conformation of the domains where they bind; the binding energy may be used to stabilize a conformation that is not present in the absence of the nanobody. For instance, NbRoco1 binds to the a0 helix of the linker; what is here the "normal" active state of the Roco protein, and is e.g. the angle between RocCOR and LRR also rotated by 135 degrees? Furthermore, nanobody NbRoco2 in the LRR domain is expected to stabilize the LRR domain; it may allow a position of the LRR domain relative to the rest of the protein that is not present without nanobody in the LRR domain. I am convinced that the observed open structure is a correct representation of the active state, but many important details have to be supported by e,g, their CX-MS experiments, and in the end probably need confirmation by more structures of other active Roco proteins or confirmation by a more dynamic sampling of the active states by e.g. molecular dynamics or NMR.
Author Response
Reviewer #1 (Public Review):
Summary:
The Roco proteins are a family of GTPases characterized by the conserved presence of an ROC-COR tandem domain. How GTP binding alters the structure and activity of Roco proteins remains unclear. In this study, Galicia C et al. took advantage of conformation-specific nanobodies to trap CtRoco, a bacterial Roco, in an active monomeric state and determined its high-resolution structure by cryo-EM. This study, in combination with the previous inactive dimeric CtRoco, revealed the molecular basis of CtRoco activation through GTP-binding and dimer-to-monomer transition.
Strengths:
The reviewer is impressed by the authors' deep understanding of the CtRoco protein. Capturing Roco proteins in a GTP-bound state is a major breakthrough in the mechanistic understanding of the activation mechanism of Roco proteins and shows similarity with the activation mechanism of LRRK2, a key molecule in Parkinson's disease. Furthermore, the methodology the authors used in this manuscript - using conformation-specific nanobodies to trap the active conformation, which is otherwise flexible and resistant to single-particle average - is highly valuable and inspiring.
Weakness:
Though written with good clarity, the paper will benefit from some clarifications.
1) The angular distribution of particles for the 3D reconstructions should be provided (Figure 1 - Sup. 1 & Sup. 2).
The supplementary figures will be adapted to include particle distribution plots.
2) The B-factors for protein and ligand of the model, Map sharpening factor, and molprobity score should be provided (Table 1).
The map used to interpret the model was post-processed by density modification, therefore no sharpening factor was obtained. This information will be included in Table 1, together with B-factors and molprobity scores.
3) A supplemental Figure to Figure 2B, illustrating how a0-helix interacts with COR-A&LRR before and after GTP binding in atomic details, will be helpful for the readers to understand the critical role of a0-helix during CtRoco activation.
A supplemental figure will be prepared to illustrate this in the revised document.
4) For the following statement, "On the other hand, only relatively small changes are observed in the orientation of the Roc a3 helix. This helix, which was previously suggested to be an important element in the activation of LRRK2 (Kalogeropulou et al., 2022), is located at the interface of the Roc and CORB domains and harbors the residues H554 and Y558, orthologous to the LRRK2 PD mutation sites N1337 and R1441, respectively."
It is not surprising the a3-helix of the ROC domain only has small changes when the ROC domain is aligned (Figure 2E). However, in the study by Zhu et al (DOI: 10.1126/science.adi9926), it was shown that a3-helix has a "see-saw" motion when the COR-B domain is aligned. Is this motion conserved in CtRoco from inactive to active state?
We indeed describe the conformational changes from the perspective of the Roc domain. When using the COR-B domain for structural alignment, a rotational movement of Roc (including a “seesaw”-like movement of the α3-helix helix around His554) with respect to COR-B is correspondingly observed. We will include this in the revised document.
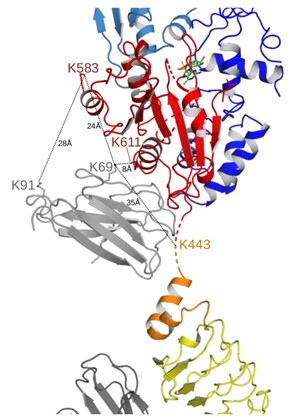
5) A supplemental figure showing the positions of and distances between NbRoco1 K91 and Roc K443, K583, and K611 would help the following statement. "Also multiple crosslinks between the Nbs and CtRoco, as well as between both nanobodies were found. ... NbRoco1-K69 also forms crosslinks with two lysines within the Roc domain (K583 and K611), and NbRoco1-K91 is crosslinked to K583".
A provisional figure displaying these crosslinks is already provided below, and we will also consider including this in the revised manuscript. However, in interpreting these crosslinks it should be taken into consideration that the additive length of the DSSO spacer and the lysine side chains leads to a theoretical upper limit of ∼26 Å for the distance between the α carbon atoms of cross-linked lysines (and even a cut-off distance of 35 Å when taking into account protein dynamics).
Author response image 1.
6) It would be informative to show the position of CtRoco-L487 in the NF and GTP-bound state and comment on why this mutation favors GTP hydrolysis.
We will create an additional figure showing the position of L487, and discuss possible mechanisms for the observed effect of a mutation on GTPase activity.
Reviewer #2 (Public Review):
Summary
The manuscript by Galicia et al describes the structure of the bacterial GTPyS-bound CtRoco protein in the presence of nanobodies. The major relevance of this study is in the fact that the CtRoco protein is a homolog of the human LRRK2 protein with mutations that are associated with Parkinson's disease. The structure and activation mechanisms of these proteins are very complex and not well understood. Especially lacking is a structure of the protein in the GTP-bound state. Previously the authors have shown that two conformational nanobodies can be used to bring/stabilize the protein in a monomer-GTPyS-bound state. In this manuscript, the authors use these nanobodies to obtain the GTPyS-bound structure and importantly discuss their results in the context of the mammalian LRRK2 activation mechanism and mutations leading to Parkinson's disease. The work is well performed and clearly described. In general, the conclusions on the structure are reasonable and well-discussed in the context of the LRRK2 activation mechanism.
Strengths:
The strong points are the innovative use of nanobodies to stabilize the otherwise flexible protein and the new GTPyS-bound structure that helps enormously in understanding the activation cycle of these proteins.
Weakness:
The strong point of the use of nanobodies is also a potential weak point; these nanobodies may have induced some conformational changes in a part of the protein that will not be present in a GTPyS-bound protein in the absence of nanobodies.
Two major points need further attention.
1) Several parts of the protein are very flexible during the monomer-dimer activity cycle. This flexibility is crucial for protein function, but obviously hampers structure resolution. Forced experiments to reduce flexibility may allow better structure resolution, but at the same time may impede the activation cycle. Therefore, careful experiments and interpretation are very critical for this type of work. This especially relates to the influence of the nanobodies on the structure that may not occur during the "normal" monomer-dimer activation cycle in the absence of the nanobodies (see also point 2). So what is the evidence that the nanobody-bound GTPyS-bound state is biochemically a reliable representative of the "normal" GTP-bound state in the absence of nanobodies, and therefore the obtained structure can be confidentially used to interpret the activation mechanism as done in the manuscript.
See below for an answer to remark 1 and 2.
2) The obtained structure with two nanobodies reveals that the nanobodies NbRoco1 and NbRoco2 bind to parts of the protein by which a dimer is impossible, respectively to a0-helix of the linker between Roc-COR and LRR, and to the cavity of the LRR that in the dimer binds to the dimerizing domain CORB. It is likely the open monomer GTP-bound structure is recognized by the nanobodies in the camelid, suggesting that overall the open monomer structure is a true GTP-bound state. However, it is also likely that the binding energy of the nanobody is used to stabilize the monomer structure. It is not automatically obvious that in the details the obtained nonobody-Roco-GTPyS structure will be identical to the "normal" Roco-GTPyS structure. What is the influence of nanobody-binding on the conformation of the domains where they bind; the binding energy may be used to stabilize a conformation that is not present in the absence of the nanobody. For instance, NbRoco1 binds to the a0 helix of the linker; what is here the "normal" active state of the Roco protein, and is e.g. the angle between RocCOR and LRR also rotated by 135 degrees? Furthermore, nanobody NbRoco2 in the LRR domain is expected to stabilize the LRR domain; it may allow a position of the LRR domain relative to the rest of the protein that is not present without nanobody in the LRR domain. I am convinced that the observed open structure is a correct representation of the active state, but many important details have to be supported by e,g, their CX-MS experiments, and in the end probably need confirmation by more structures of other active Roco proteins or confirmation by a more dynamic sampling of the active states by e.g. molecular dynamics or NMR.
Recently, nanobodies have increasingly been used successfully to obtain structural insights in protein conformational states (reviewed in Uchański et al, Curr. Opin. Struc. Biol. 2020). As reviewer # 2 points out, the concern is sometimes raised that antibodies could distort a protein into non-native conformations. Here, it is important to note that the nanobodies were raised by immunizing a llama with the fully native CtRoco protein bound to a non-hydrolysable GTP analogue, after which the nanobodies were selected by phage display using the same fully native and functional form of the protein. As clearly explained in Manglik et al. Annu Rev Pharmacol Toxicol. 2017, the probability of an in vivo matured nanobody inducing a non-native conformation of the antigen is low, although it is possible that it selects a high-energy, low-population conformation of a dynamic protein. Immature B cells require engagement of displayed antibodies with antigen to proliferate and differentiate during clonal selection. Antibodies that induce non-native conformations of the antigen pay a substantial energetic penalty in this process, and B cell clones displaying such antibodies will have a significantly lower probability of proliferation and differentiation into mature antibody-secreting B lymphocytes. Hence, many recent experiments and observation give credence to the notion that nanobodies bind antigens primarily by conformational selection and not induced fit (e.g. Smirnova et al. PNAS 2015).
Extrapolated to the case of CtRoco, which is clearly very flexible in its GTP-bound form, this means that the nanobodies are able to trap and stabilize one conformational state that is representative of the “active state” ensemble of the protein. In this respect, it is clear from our experiments (XL-MS, affinity and effect on GTPase activity) that the effects of NbRoco1 and NbRoco2 are additive (or even cooperative), meaning that both nanobodies recognize different features of the same CtRoco “active state”. Correspondingly, the monomeric, elongated “open” conformation is also observed in the structure of CtRoco bound to NbRoco1 only (Figure1 - supplement 2), albeit that this structure still displays more flexibility. The monomerization and conformational changes that we observe and describe in the current paper at high resolution are also in very good agreement with earlier observations for CtRoco in the GTP-bound form in absence of any nanobodies, including negative stain EM (Deyaert et al. Nature Commun, 2017), hydrogen-deuterium exchange experiments (Deyaert et al. Biochem. J. 2019) and native MS (Leemans et al. Biochem J. 2020).
In the revised document we will include some additional text to address and clarify these aspects.



