Peer review process
Revised: This Reviewed Preprint has been revised by the authors in response to the previous round of peer review; the eLife assessment and the public reviews have been updated where necessary by the editors and peer reviewers.
Read more about eLife’s peer review process.Editors
- Reviewing EditorBruno LemaitreÉcole Polytechnique Fédérale de Lausanne, Lausanne, Switzerland
- Senior EditorWendy GarrettHarvard T.H. Chan School of Public Health, Boston, United States of America
Reviewer #2 (Public review):
The resubmitted version of the paper by Yan et al. titled "Frequent intertrophic transmission of Wolbachia by parasitism but not predation" contains all the major flaws I found in the original submission. As far as I could see, the authors did not address my original concerns.
In short:
(1) A control of Portiera MUST be included in the FISH experiments, if the claim that Wolbachia is not only transferred from a parasitoid to the whitefly, but finds its way to the bacteriocytes. This is especially true for the Q, a biotype for which the pattern of Wolbachia distribution has been documented as scattered in naturally infected populations. The very strong signal in the whitefly bacteriocytes implies Portiera.
(2) In my original review I wrote: "The authors fail to discuss, or even acknowledge, a number of published studies that specifically show no horizontal transmission such as the one claimed to be detected in the study presented." In return the authors wrote in their rebuttal letter: "We have made corresponding modifications to the discussion section (Lines 256-271in the revised manuscript) and have discussed the published studies that report no evidence of horizontal transmission (Lines 260-263 in the revised manuscript)." However, the stated lines are concerned with a different subject. In addition, in their letter the authors write "Additionally, some experiments have found no evidence of horizontal transmission of Wolbachia (39- 42) (Lines 260-263 in the revised manuscript)." Beside the fact that the line numbers are wrong, the papers cited are entirely irrelevant as they do not discuss parasitoids.
(3) My original comment on the origin of sequences used for the phylogenetical analysis still stands. It is hard to claim a data-based search, when most of the data originate in the authors lab. The explanation of the confusion with the Qi et al. (2019) paper should at least be mentioned in the M&M. Apologies if it has been included and I missed it.
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public Review):
Summary and Strengths:
The ability of Wolbachia to be transmitted horizontally during parasitoid wasp infections is supported by phylogenetic data here and elsewhere. Experimental analyses have shown evidence of wasp-to-wasp transmission during coinfection (eg Huigins et al), host to wasp transmission (eg Heath et al), and mechanical ('dirty needle') transmission from host to host (Ahmed et al). To my knowledge this manuscript provides the first experimental evidence of wasp to host transmission. Given the strong phylogenetic pattern of host-parasitoid Wolbachia sharing, this may be of general importance in explaining the distribution of Wolbachia across arthropods. This is of interest as Wolbachia is extremely common in the natural world and influences many aspects of host biology.
Weaknesses:
The first observation of the manuscript is that the Wolbachia strains in hosts are more closely related to those in their parasitoids. This has been reported on multiple occasions before, dating back to the late 1990s. The introduction cites five such papers (the observation is made in other studies too that could be cited) but then dismisses them by stating "However, without quantitative tests, this observation could simply reflect a bias in research focus." As these studies include carefully collected datasets that were analysed appropriately, I felt this claim of novelty was rather strong. It is unclear why downloading every sequence in GenBank avoids any perceived biases, when presumably the authors are reanalysing the data in these papers.
Thank you for bringing this to our attention. In this study, we downloaded all wsp sequences from GenBank and conducted a systematic analysis. We acknowledge that there could still be a bias in research focus, but a systematic analysis, compared to a limited dataset, may reduce this bias. We agree with the reviewer's point, and we have revised this statement to make it more accurate. Now the new sentence reads: "However, there is still a lack of systematic statistical analyses to support this hypothesis." (Lines 69–70 in the revised manuscript)
I do not doubt the observation that host-parasitoid pairs tend to share related Wolbachia, as it is corroborated by other studies, the effect size is large, and the case study of whitefly is clearcut. It is also novel to do this analysis on such a large dataset. However, the statistical analysis used is incorrect as the observations are pseudo-replicated due to phylogenetic non-independence. When analysing comparative data like this it is essential to correct for the confounding effects of related species tending to be similar due to common ancestry. In this case, it is well-known that this is an issue as it is a repeated observation that related hosts are infected by related Wolbachia. However, the authors treat every pairwise combination of species (nearly a million pairs) as an independent observation. Addressing this issue is made more complex because there are both the host and symbiont trees to consider. The additional analysis in lines 123-124 (including shuffling species pairs) does not explicitly address this issue.
We agree with your point about the non-independence of data due to phylogenetic relationships. In the analysis of species traits, a conventional phylogenetic correction assumes that traits follow a Brownian motion model (Felsenstein, 1985). The variance of the trait values for a species i is given by:
Var[Yi]=σ2Ti,
Where Ti represents the time from the root to the tip for species i. Consequently, the covariance between traits of species i and j is:
Cov[Yij,Yj]=σ2Tii,
where Tij is the time from the root to the most recent common ancestor (MRCA) of species i and j. Linear model analysis incorporates the covariance matrix to correct for the effects of non-independence. Mathematically, this method is equivalent to the independent contrasts approach (Felsenstein, 1985).
In our analysis, we treat the minimum interspecific wsp distance between two species as a trait for the species pair (i, j). Similarly, for any two pairs of species (i, j) and (k, l), we postulate that the covariance between their traits is given by:
Cov[Yij,Ykl]=σ2⋅(Tik+Tjl),
where Tik denotes the time from the root to the MRCA of species i and k, and Tjl represents the time from the root to the MRCA of species j and l. This covariance matrix is then incorporated into our linear model analysis to account for the effects of phylogenetic non-independence.
However, when extending trait analysis to pairs of species, the computational demands increase substantially. For instance, with a dataset of 1,377 species, forming all possible pairs yields 947,376 unique species combinations. Consequently, constructing a covariance matrix for these pairs would necessitate storing 897,521,285,376 entries, a requirement that far exceeds the memory capabilities of standard computing systems.
To address this, we randomly sampled 1,000 pairs from the total of 947,376 species pairs within the 'Others' category, thereby reducing the computational load without compromising the representativeness of our analysis. Ultimately, even after accounting for phylogenetic correction using covariance, the effect of parasitism remains highly significant (p < 0.0001).
We have added a “Phylogenetic correction” section to Materials and Methods (Lines 392–405 in the revised manuscript). The corresponding results are described on lines 120–121 and in supplementary Note 1. The data and scripts for this analysis are available at https://doi.org/10.6084/m9.figshare.24718119.
REFERENCE
Felsenstein J, 1985. Phylogenies and the comparative method. The American Naturalist, 125(1), 1-15.
The sharing of Wolbachia between whitefly and their parasitoids is very striking, although this has been reported before (eg the authors recently published a paper entitled "Diversity and Phylogenetic Analyses Reveal Horizontal Transmission of Endosymbionts Between Whiteflies and Their Parasitoids"). In Lines 154-164 it is suggested that from the tree the direction of transfer between host and parasitoid can be inferred from the data. This is not obvious to me given the poor resolution of the tree due to low sequence divergence. There are established statistical approaches to test the direction of trait changes on a tree that could have been used (a common approach is to use the software BEAST).
We thank the reviewer for this constructive feedback on our interpretation of Wolbachia transfer between whiteflies and their parasitoids. Inspired by the reviewer's comments, we have now incorporated a trait-based approach, using the taxonomic order of the source species of the wsp gene as a discrete trait for ancestral state reconstruction on the wsp tree. The estimated ancestral trait state for one clade, which clusters wsp sequences from whiteflies and parasitoids, is Hymenoptera, suggesting that within this clade, the direction of Wolbachia transfer may have been from parasitoids to hosts. Conversely, in another clade characterized by the ancestral trait state of Hemiptera, the inferred direction of transfer appears to be from hosts to parasitoids. We have added a “Ancestral state reconstruction” section to Materials and Methods (Lines 406–412 in the revised manuscript). The corresponding results are described on lines 159–163 and 167–168. The data and script for this analysis is available at https://doi.org/10.6084/m9.figshare.24718119.
Reviewer #2 (Public Review):
The paper by Yan et al. aims to provide evidence for horizontal transmission of the intracellular bacterial symbiont Wolbachia from parasitoid wasps to their whitefly hosts. In my opinion, the paper in its current form consists of major flaws.
Weaknesses:
The dogma in the field is that although horizontal transmission events of Wolbachia occur, in most systems they are so rare that the chances of observing them in the lab are very slim.
For the idea of bacteria moving from a parasitoid to its host, the authors have rightfully cited the paper by Hughes, et al. (2001), which presents the main arguments against the possibility of documenting such transmissions. Thus, if the authors want to provide data that contradict the large volume of evidence showing the opposite, they should present a very strong case.
In my opinion, the paper fails to provide such concrete evidence. Moreover, it seems the work presented does not meet the basic scientific standards.
We are grateful for your critical perspective on our work. Nonetheless, we are confident in the credibility of our findings regarding the horizontal transmission of Wolbachia from En. formosa to B. tabaci. Our study has documented this phenomenon through phylogenetic tree analyses, and we have further substantiated our observations with rigorous experiments in both cages and petri dishes. The horizontal transfer of Wolbachia was confirmed via PCR, with the wsp sequences in B. tabaci showing complete concordance with those in En. formosa. Additionally, we utilized FISH, vertical transmission experiments, and phenotypic assays to demonstrate that the transferred Wolbachia could be vertically transmitted and induce significant fitness cost in B. tabaci. All experiments were conducted with strict negative controls and a sufficient number of replicates to ensure reliability, thereby meeting basic scientific standards. The collective evidence we present points to a definitive case of Wolbachia transmission from the parasitoid En. formosa to the whitefly B. tabaci.
My main reservations are:
- I think the distribution pattern of bacteria stained by the probes in the FISH pictures presented in Figure 4 looks very much like Portiera, the primary symbiont found in the bacterium of all whitefly species. In order to make a strong case, the authors need to include Portiera probes along with the Wolbachia ones.
We thank you for your critical evaluation regarding the specificity of FISH in our study. We assure the reliability of our FISH results based on several reasons.
(1) We implemented rigorous negative controls which exhibited no detectable signal, thereby affirming the specificity of our hybridization. (2) The central region of the whitefly nymphs is a typical oviposition site for En. formosa. Post-parasitism, we observed FISH signals around the introduced parasitoid eggs, distinct from bacteriocyte cells which are rich in endosymbionts including Portiera (Fig 3e-f). This observation supports the high specificity of our FISH method. (3) In the G3 whiteflies, we detected the presence of Wolbachia in bacteriocytes in nymphs and at the posterior end of eggs in adult females (Fig. 4). This distribution pattern aligns with previously reported localizations of Wolbachia in B. tabaci (Shi et al., 2016; Skaljac et al., 2013). Furthermore, the distribution of Wolbachia in the whiteflies does indeed exhibit some overlap with that of Portiera (Skaljac et al., 2013; Bing et al., 2014). 4) The primers used in our FISH assays have been widely cited (Heddi et al., 1999) and validated in studies on B. tabaci and other systems (Guo et al., 2018; Hegde et al., 2024; Krafsur et al., 2020; Rasgon et al., 2006; Uribe-Alvarez et al., 2019; Zhao et al., 2013).
Taking all these points into consideration, we stand by the reliability of our FISH results.
REFERENCES
Bing XL, Xia WQ, Gui JD, et al., 2014. Diversity and evolution of the Wolbachia endosymbionts of Bemisia (Hemiptera: Aleyrodidae) whiteflies. Ecol Evol, 4(13):2714-37.
Guo Y, Hoffmann AA, Xu XQ, et al., 2018. Wolbachia-induced apoptosis associated with increased fecundity in Laodelphax striatellus (Hemiptera: Delphacidae). Insect Mol Biol, 27:796-807.
Heddi A, Grenier AM, Khatchadourian C, Charles H, Nardon P, 1999. Four intracellular genomes direct weevil biology: nuclear, mitochondrial, principal endosymbiont, and Wolbachia. Proc Natl Acad Sci USA, 96:6814-6819.
Hegde S, Marriott AE, Pionnier N, et al., 2024. Combinations of the azaquinazoline anti-Wolbachia agent, AWZ1066S, with benzimidazole anthelmintics synergise to mediate sub-seven-day sterilising and curative efficacies in experimental models of filariasis. Front Microbiol, 15:1346068.
Krafsur AM, Ghosh A, Brelsfoard CL, 2020. Phenotypic response of Wolbachia pipientis in a cell-free medium. Microorganisms, 8.
Rasgon JL, Gamston CE, Ren X, 2006. Survival of Wolbachia pipientis in cell-free medium. Appl Environ Microbiol, 72:6934-6937.
Shi P, He Z, Li S, et al., 2016. Wolbachia has two different localization patterns in whitefly Bemisia tabaci AsiaII7 species. PLoS One, 11: e0162558.
Skaljac M, Zanić K, Hrnčić S, et al., 2013. Diversity and localization of bacterial symbionts in three whitefly species (Hemiptera: Aleyrodidae) from the east coast of the Adriatic Sea. Bull Entomol Res, 103(1):48-59.
Uribe-Alvarez C, Chiquete-Félix N, Morales-García L, et al., 2019. Wolbachia pipientis grows in Saccharomyces cerevisiae evoking early death of the host and deregulation of mitochondrial metabolism. MicrobiologyOpen, 8: e00675.
Zhao DX, Zhang XF, Chen DS, Zhang YK, Hong XY, 2013. Wolbachia-host interactions: Host mating patterns affect Wolbachia density dynamics. PLoS One, 8: e66373.
- If I understand the methods correctly, the phylogeny presented in Figure 2a is supposed to be based on a wide search for Wolbachia wsp gene done on the NCBI dataset (p. 348). However, when I checked the origin of some of the sequences used in the tree to show the similarity of Wolbachia between Bemisia tabaci and its parasitoids, I found that most of them were deposited by the authors themselves in the course of the current study (I could not find this mentioned in the text), or originated in a couple of papers that in my opinion should not have been published to begin with.
We appreciate your meticulous examination of the sources for our sequence data. All the sequences included in our phylogenetic analysis were indeed downloaded from the NCBI database as of July 2023. The sequences used to illustrate the similarity of Wolbachia between B. tabaci and its parasitoids include those from our previously published study (Qi et al., 2019), which were sequenced from field samples. Additionally, some sequences were also obtained from other laboratories (Ahmed et al., 2009; Baldo et al., 2006; Van Meer et al., 1999). We acknowledge that in our prior research (Qi et al., 2019), the sequences were directly submitted to NCBI and, regrettably, we did not update the corresponding publication information after the article were published. It is not uncommon for sequences on NCBI, with some never being followed by a published paper (e.g., FJ710487- FJ710511 and JF426137-JF426149), or not having their associated publication details updated post-publication (for instance, sequences MH918776-MH918794 from Qi et al., 2019, and KF017873-KF017878 from Fattah-Hosseini et al., 2018). We recognize that this practice can lead to confusion and apologize for the oversight in our work.
REFERENCES
Ahmed MZ, Shatters RG, Ren SX, Jin GH, Mandour NS, Qiu BL, 2009. Genetic distinctions among the Mediterranean and Chinese populations of Bemisia tabaci Q biotype and their endosymbiont Wolbachia populations. J Appl Entomol, 133:733-741.
Baldo L, Dunning Hotopp JC, Jolley KA, et al., 2006. Multilocus sequence typing system for the endosymbiont Wolbachia pipientis. Appl Environ Microbiol. 72(11):7098-110.
Fattah-Hosseini S, Karimi J, Allahyari H, 2014. Molecular characterization of Iranian Encarsia formosa Gahan populations with natural incidence of Wolbachia infection. J Entomol Res Soc, 20(1):85–100.
Qi LD, Sun JT, Hong XY, Li YX, 2019. Diversity and phylogenetic analyses reveal horizontal transmission of endosymbionts between whiteflies and their parasitoids. J Econ Entomol, 112(2):894-905.
Van Meer MM, Witteveldt J, Stouthamer R, 1999. Phylogeny of the arthropod endosymbiont Wolbachia based on the wsp gene. Insect Mol Biol, 8(3):399-408.
- The authors fail to discuss or even acknowledge a number of published studies that specifically show no horizontal transmission, such as the one claimed to be detected in the study presented.
Thank you for bringing this to our attention. We have made corresponding modifications to the discussion section (Lines 256–271 in the revised manuscript) and have discussed the published studies that report no evidence of horizontal transmission (Lines 260–263 in the revised manuscript). The added sentences read: “Experimental confirmations of Wolbachia horizontal transfer remain relatively rare, with only a limited number of documented cases (24, 27, 37, 38). Additionally, some experiments have found no evidence of horizontal transmission of Wolbachia (39-42).” (Lines 260–263 in the revised manuscript)
Reviewer #3 (Public Review):
This is a very ordinary research paper. The horizontal of endosymbionts, including Wolbachia, Rickettsia etc. has been reported in detail in the last 10 years, and parasitoid vectored as well as plant vectored horizontal transmission is the mainstream of research. For example, Ahmed et al. 2013 PLoS One, 2015 PLoS Pathogens, Chiel et al. 2014 Enviromental Entomology, Ahmed et al. 2016 BMC Evolution Biology, Qi et al. 2019 JEE, Liu et al. 2023 Frontiers in Cellular and Infection Microbiology, all of these reported the parasitoid vectored horizontal transmission of endosymbiont. While Caspi-Fluger et al. 2012 Proc Roy Soc B, Chrostek et al. 2017 Frontiers in Microbiology, Li et al. 2017 ISME Journal, Li et al. 2017 FEMS, Shi et al. 2024 mBio, all of these reported the plant vectored horizontal transmission of endosymbiont. For the effects of endosymbiont on the biology of the host, Ahmed et al. 2015 PLoS Pathogens explained the effects in detail.
Thank you for the insightful comments and for highlighting the relevant literature in the field of horizontal transmission of endosymbionts, including Wolbachia and Rickettsia. After careful consideration of the studies mentioned in the commences, we believe that our work presents significant novel contributions to the field. 1) Regarding the parasitoid-mediated horizontal transmission of Wolbachia, most of the cited articles, such as Ahmed et al. 2013 in PLoS One and Ahmed et al. 2016 in BMC Evolutionary Biology, propose hypotheses but do not provide definitive evidence. The transmission of Wolbachia within the whitefly cryptic species complex (Ahmed et al. 2013) or between moths and butterflies (Ahmed et al. 2016) could be mediated by parasitoids, plants, or other unknown pathways. 2) Chiel et al. 2014 in Environmental Entomology reported “no evidence for horizontal transmission of Wolbachia between and within trophic levels” in their study system. 3) The literature you mentioned about Rickettsia, rather than Wolbachia, indirectly reflects the relative scarcity of evidence for Wolbachia horizontal transmission. For example, the evidence for plant-mediated transmission of Wolbachia remains isolated, with Li et al. 2017 in the ISME Journal being one of the few reports supporting this mode of transmission. 4) While the effects of endosymbionts on their hosts are not the central focus of our study, the effects of transgenerational Wolbachia on whiteflies are primarily demonstrated to confirm the infection of Wolbachia into whiteflies. Furthermore, the effects we report of Wolbachia on whiteflies are notably different from those reported by Ahmed et al. 2015 in PLoS Pathogens, likely due to different whitefly species and Wolbachia strains. 6) More importantly, our study reveals a mechanism of parasitoid-mediated horizontal transmission of Wolbachia that is distinct from the mechanical transmission suggested by Ahmed et al. 2015 in PLoS Pathogens. Their study implies transmission primarily through dirty needle, without Wolbachia infection of the parasitoid, suggesting host-to-host transmission at the same trophic level, where parasitoids serve as phoretic vectors. In contrast, our findings demonstrate transmission from parasitoids to hosts through unsuccessful parasitism, which represents cross-trophic level transmission. To our knowledge, this is the first experimental evidence that Wolbachia can be transmitted from parasitoids to hosts. We believe these clarifications and the novel insights provided by our research contribute valuable knowledge to the field.
REFERENCES
Ahmed MZ, De Barro PJ, Ren SX, Greeff JM, Qiu BL, 2013. Evidence for horizontal transmission of secondary endosymbionts in the Bemisia tabaci cryptic species complex. PLoS One, 8(1):e53084.
Ahmed MZ, Li SJ, Xue X, Yin XJ, Ren SX, Jiggins FM, Greeff JM, Qiu BL, 2015. The intracellular bacterium Wolbachia uses parasitoid wasps as phoretic vectors for efficient horizontal transmission. PLoS Pathog, 10(2):e1004672.
Ahmed MZ, Breinholt JW, Kawahara AY, 2016. Evidence for common horizontal transmission of Wolbachia among butterflies and moths. BMC Evol Biol, 16(1):118.
Caspi-Fluger A, Inbar M, Mozes-Daube N, Katzir N, Portnoy V, Belausov E, Hunter MS, Zchori-Fein E, 2012. Horizontal transmission of the insect symbiont Rickettsia is plant-mediated. Proc Biol Sci, 279(1734):1791-6.
Chiel E, Kelly SE, Harris AM, Gebiola M, Li X, Zchori-Fein E, Hunter MS, 2014. Characteristics, phenotype, and transmission of Wolbachia in the sweet potato whitefly, Bemisia tabaci (Hemiptera: Aleyrodidae), and its parasitoid Eretmocerus sp. nr. emiratus (Hymenoptera: Aphelinidae). Environ Entomol, 43(2):353-62.
Chrostek E, Pelz-Stelinski K, Hurst GDD, Hughes GL, 2017. Horizontal transmission of intracellular insect symbionts via plants. Front Microbiol, 8:2237.
Li SJ, Ahmed MZ, Lv N, Shi PQ, Wang XM, Huang JL, Qiu BL, 2017. Plant-mediated horizontal transmission of Wolbachia between whiteflies. ISME J, 11(4):1019-1028.
Li YH, Ahmed MZ, Li SJ, Lv N, Shi PQ, Chen XS, Qiu BL, 2017. Plant-mediated horizontal transmission of Rickettsia endosymbiont between different whitefly species. FEMS Microbiol Ecol, 93(12).
Liu Y, He ZQ, Wen Q, Peng J, Zhou YT, Mandour N, McKenzie CL, Ahmed MZ, Qiu BL, 2023. Parasitoid-mediated horizontal transmission of Rickettsia between whiteflies. Front Cell Infect Microbiol, 12:1077494.
Qi LD, Sun JT, Hong XY, Li YX, 2019. Diversity and phylogenetic analyses reveal horizontal transmission of endosymbionts between whiteflies and their parasitoids. J Econ Entomol, 112(2):894-905.
Shi PQ, Wang L, Chen XY, Wang K, Wu QJ, Turlings TCJ, Zhang PJ, Qiu BL, 2024. Rickettsia transmission from whitefly to plants benefits herbivore insects but is detrimental to fungal and viral pathogens. mBio, 15(3):e0244823.
Weaknesses:
In the current study, the authors downloaded the MLST or wsp genes from a public database and analyzed the data using other methods, and I think the authors may not be familiar with the research progress in the field of insect symbiont transmission, and the current stage of this manuscript lacking sufficient novelty.
We appreciate your critical perspective on our study. However, we respectfully disagree with the viewpoint that our manuscript lacks sufficient novelty.
Recommendations for the authors:
Reviewer #1 (Recommendations For The Authors):
The data and scripts from the experimental section of the paper are not made publicly available. This would be good practice. It may well be a requirement for this journal too, but I have not read the journal policy on this matter.
Thank you for the kind reminder, we have uploaded the data and scripts to the public database at https://doi.org/10.6084/m9.figshare.24718119.
• Line 16 should read 'intertrophic' not 'intertropical'.
Corrected.
• Line 50 should not say 'the most infectious' as this is an incorrect use of the word 'infectious'. Maybe 'common'? Should also add something like 'likely' here.
Corrected. The new sentence reads “Together, these characteristics make Wolbachia likely the most common microbe on Earth in terms of the number of species it infects (7, 8).” (Lines 47–49 in the revised manuscript).
• Line 54 These references are all about mosquito disease vectors, not pests. More generally, in this paragraph, the research interest in Wolbachia relates overwhelmingly to blocking arbovirus transmission and not controlling pest populations.
To enhance consistency with our statements, we have revised the supporting references as follows:
X. Zheng et al., "Combined incompatible and sterile insect techniques eliminate mosquitoes," Nature 572, 56-61 (2019).
A. A. Hoffmann et al., "Wolbachia establishment in Aedes populations to suppress dengue transmission," Nature 476, 454-457 (2011).
J. T. Gong, T. P. Li, M. K. Wang, X. Y. Hong, "Prospects of Wolbachia in agricultural Pest Control," Current Opinion in Insect Science 57, 101039 (2023).J. T. Gong et al., "Stable integration of plant-virus-inhibiting Wolbachia into planthoppers for rice protection," Current Biology 30, 4837-4845.e4835 (2020).
Regarding the content of the articles:
Zheng et al. (2019) detail the successful suppression of wild mosquito populations through the release of male mosquitoes artificially infected with Wolbachia.
Gong et al. (2020) present the potential of releasing Wolbachia-infected brown planthoppers to inhibit plant viruses and control pest populations.
Gong et al. (2023) provide a comprehensive review on the application and future of Wolbachia in managing agricultural pests.
• Line 60-61. This sentence seems poorly supported by theory or data. I suggest it is deleted. Why should CI cause extinction, and why would it have a major effect on genetic diversity beyond mtDNA?
We have deleted the statements about extinction or genetic diversity. Now the sentence reads “It may also spread to nontarget organisms, potentially disrupting their population dynamics.” (Lines 57–58 in the revised manuscript)
• Line 66. Reword to make clear these routes are not an exhaustive list.
We have reworded these sentences. The new sentences now read “Similar to other symbionts, Wolbachia host shifts may occur through three main routes: parasitism, predation, and shared plant or other food sources (17). However, it is important to note that these are not the only routes through which transmission may occur, and the specific contributions of each to the overall process of host shift are not yet fully understood.” (Lines 62–66 in the revised manuscript).
• Line 77-79. This could do with mentioning studies of parasitoid-to-host transmission like Ahmedd et al given that it is common knowledge that insects commonly survive parasitoid attacks.
We have added sentences acknowledging the common occurrence of insects surviving parasitoid attacks and referenced and described the Ahmed et al. 2015 study. The added sentences read:
“However, it is common in nature for hosts to survive parasitoid attacks (27-29). For example, whiteflies can survive after attacks of Eretmocerus parasitoids (27). These parasitoids can act as phoretic vectors, facilitating the spread of Wolbachia within whitefly populations through the contamination of their mouthparts and ovipositors with Wolbachia during the probing process (27).” (Lines 77–82 in the revised manuscript).
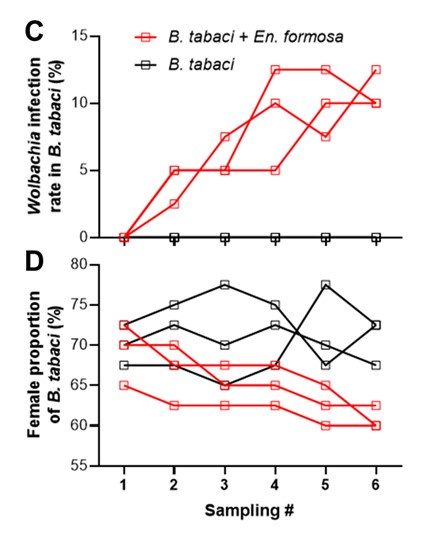
• Line 173. Mention that there are three replicates of each cage. In Figures 2C and D, it is better to show each replicate as a separate line to see how consistent they are.
In accordance with the reviewer's suggestion, we have included a statement highlighting the replication of our experiments: “Notably, each cage setup was replicated three times to ensure experimental rigor.” (Lines 179–180 in the revised manuscript).
Regarding Figures 2C and D, we have revised the figures to display each replicate as a separate line, as suggested. However, we have encountered a visual clutter that may detract from the clarity of the figures. Additionally, in Figure C, the three black lines, all representing zero values, do not allow for the distinction of individual trends. Therefore, we recommend retaining the original figure format. In accordance with eLife's data policy, we have also provided the source data for all figures, ensuring that readers can access to the detailed data, thus balancing the need for visual simplicity with the provision of comprehensive data.
Author response image 1.
• The GloBI database is central to the phylogenetic analysis and it would be helpful to have a few words in the results stating where this information comes from.
The revised sentence now reads: “To investigate potential horizontal transmission of Wolbachia, we retrieved 4685 wsp sequences from the NCBI database, and species interaction relationships were extracted from the GloBI database (for details, see Methods and Materials).” (Lines 94–96 in the revised manuscript).
Reviewer #3 (Recommendations For The Authors):
To improve the quality of this manuscript, I have some questions and suggestions.
Introduction:
Line 41-42, I don't agree with this statement, as mentioned above, the ways of insect symbiont transmission have been studied in the last 10 years.
According to the reviewer’s suggestion, we have deleted this statement.
Line 75-76, Again, the statement is not correct, many studies have clearly revealed and confirmed that Wolbachia CAN be transferred from parasitoid to their insect hosts including whitefly Bemisia tabaci.
Thank you for your insightful comments. After careful consideration of the studies you have mentioned above, none of these articles provided definitive evidence supporting the transfer of Wolbachia from parasitoids to their insect hosts. A closely related study is Ahmed et al. (2015) in PLoS Pathogens. This article demonstrates that parasitoid wasps can act as phoretic vectors mediating the transmission of Wolbachia between whiteflies. However, Wolbachia did not infect the parasitoid wasps themselves. Therefore, this study does not provide evidence for intertrophic transmission of Wolbachia from parasitoids to their hosts. To avoid confusion, we have cited the Ahmed et al. (2015) reference following this statement and described its findings accordingly. (Lines 88-92 in revised manuscript).
Results:
Line 133-134, Ahmed et al. 2016 BMC Evolution Biology, clearly revealed and confirmed the "common horizontal transmission of Wolbachia between butterflies and moths".
We thank you for guiding us to the relevant study. Ahmed et al. 2016 BMC Evolution Biology suggested common horizontal transmission of Wolbachia between butterflies and moths and proposed that this horizontal transmission might be caused by parasitoid wasps. Here, we present the potential Wolbachia transfer between Trichogramma and their lepidopteran hosts (Lines 135–136 in revised manuscript). Integrating the results from Ahmed et al. 2016, our result also suggests that Trichogramma wasps may be the vectors for horizontal transmission of Wolbachia among lepidopteran hosts. We have discussed this point in the discussion section and cited Ahmed et al. 2016 BMC Evolution Biology (Lines 239–246 in revised manuscript).
Line 176-177, as we know Wolbachia in Encarsia formosa is a strain of parthenogenesis, why did it reduce the female ratio of whitefly progeny after it was transmitted to whitefly B. tabaci, it needs a convincing explanation.
Wolbachia induces parthenogenesis in En. formosa. However, we observed that Wolbachia from En. formosa failed to induce parthenogenesis in B. tabaci, possibly due to the requirement for host gene compatibility. Additionally, we noted a reduced female ratio in B. tabaci infected with En. formosa Wolbachia. We speculate that this might result from the burden imposed by En. formosa Wolbachia on the new host, potentially reducing fertilization success rates and indirectly leading to a decrease in the female ratio. Similarly, we observed a decline in female fecundity, egg hatching rate, and immature survival rate in B. tabaci infected with En. formosa Wolbachia. The mechanisms underlying these fitness costs remain unclear and warrant further in-depth research.
Line 189-190, do the authors have convincing evidence that the 60Gy irradiation only has effects on the reproduction of En. formosa, but does not have any negative effects on the activity of Wolbachia? I think there may be.
We observed that after irradiation, the titer of Wolbachia within En. formosa significantly decreased (Fig S3). We agree that the irradiation may cause other negative effects on Wolbachia which is worth of close investigation. However, even with a significant reduction in Wolbachia titer, irradiation increased the infection rate of Wolbachia in surviving B. tabaci after wasp attacks (Fig 3C). We speculate that this may be due to irradiation of En. formosa increasing the rate of parasitic failure. While the full extent of the effects of irradiation on Wolbachia is not yet clear in our experiments, it does not alter our conclusion that Wolbachia can be transmitted from En. formosa to whitefly hosts through failed parasitism.
Discussion:
Line 289-290, I don't understand, why the authors think from parasitoid Eretmocerus to whitefly, and from Trichogramma to moth, are the same trophic level, they are indeed two different trophic levels.
Thank you for your feedback. We have conducted a thorough search but were unable to locate the specific statement you are referring to. If there has been any ambiguity in our manuscript that has led to confusion, we sincerely apologize for any misunderstanding it may have caused. We agree with your perspective and have always considered the parasitoid Eretmocerus and whitefly, as well as Trichogramma and moth, to be at different trophic levels. However, in the context of specific references, such as Ahmed et al. 2015 in PLoS Pathogens, we believe that Wolbachia is transmitted within the same trophic level without infecting the parasitoid Eretmocerus, merely serving as a phoretic vector to facilitate the spread of Wolbachia among whitefly hosts. Similarly, in the case of Huigens et al. 2000 in Nature, Wolbachia uses lepidopteran hosts as vectors to promote its transmission among Trichogramma without the need to infect the lepidopteran hosts themselves.
Materials and Methods
Line 348, what is tblastn?
We have corrected tblastn to TBLASTN. We are grateful to the reviewer for pointing this out. Here, we utilized TBLASTN instead of BLASTN, to avoid missing the rapidly evolving wsp sequences. Because alignment at the protein level is generally more sensitive than at the nucleotide level. TBLASTN is a bioinformatics tool within the BLAST (Basic Local Alignment Search Tool) suite used for comparing a protein query sequence against a nucleotide database. Specifically, TBLASTN aligns a given protein sequence with nucleotide sequences in a database by translating the nucleotide sequences into all possible protein sequences (considering different reading frames) and comparing them to the query protein sequence.
Line 383, how was the Wolbachia-free line of B. tabaci established, by antibiotics? If so, how do we ensure the antibiotic does not have any negative to other symbionts in whitefly B. tabaci?
The Wolbachia-free line of B. tabaci was collected from field, without the treatment of antibiotics. We have made revisions in the Materials and Methods section to clarify this, stating, "An iso-female line of B. tabaci, which is naturally Wolbachia-free and has not been treated with antibiotics, was established." (Lines 417–418 in the revised manuscript)
Line 419-421 as I mentioned before, the irradiation may have negative effects on Wolbachia too, so change the biology of both Encarsia and whitefly host.
We observed that after irradiation, the titer of Wolbachia within En. formosa significantly decreased (Fig S3). However, even with a significant reduction in Wolbachia titer, irradiation increased the infection rate of Wolbachia in surviving B. tabaci after wasp attacks (Fig 3C). We speculate that this may be due to irradiation of En. formosa increasing the rate of parasitic failure. While the full extent of the effects of irradiation on Wolbachia is not yet clear in our experiments, it does not alter our conclusion that Wolbachia can be transmitted from En. formosa to whitefly hosts through failed parasitism.
Line 452-453, From egg to eclosion, it needs about 21 days to understand suitable temperature and other conditions, during this period, the egg and nymphs can not move, so how to keep the cut-leaf fresh enough in a Petri dish for 21 days?
We apologize for not clearly describing the materials and methods. By using wet cotton to wrap the end of petiole of the leaf, we can keep the leaves fresh for up to a month. We have included this detail in the materials and methods to enhance the reproducibility of the experiment. “A single irradiated wasp was subsequently introduced into a Petri dish, which contained a tomato leaf infested with Wolbachia-free third or fourth instar whitefly nymphs, and wet cotton was used to wrap the end of the leaf petiole to keep the leaf fresh.” (Lines 455–458 in the revised manuscript)



