Peer review process
Revised: This Reviewed Preprint has been revised by the authors in response to the previous round of peer review; the eLife assessment and the public reviews have been updated where necessary by the editors and peer reviewers.
Read more about eLife’s peer review process.Editors
- Reviewing EditorAndres Jara-OsegueraThe University of Texas at Austin, Austin, United States of America
- Senior EditorMerritt MadukeStanford University, Stanford, United States of America
Reviewer #1 (Public review):
Summary:
This manuscript reports the substrate-bound structure of SiaQM from F. nucleatum, which is the membrane component of a Neu5Ac-specific Tripartite ATP-dependent Periplasmic (TRAP) transporter. Until recently, there was no experimentally derived structural information regarding the membrane components of TRAP transporter, limiting our understanding of the transport mechanism. Since 2022, there have been 3 different studies reporting the structures of the membrane components of Neu5Ac-specific TRAP transporters. While it was possible to narrow down the binding site location by comparing the structures to proteins of the same fold, a structure with substrate bound has been missing. In this work, the authors report the Na+-bound state and the Na+ plus Neu5Ac state of FnSiaQM, revealing information regarding substrate coordination. In previous studies, 2 Na+ ion sites were identified. Here, the authors also tentatively assign a 3rd Na+ site. The authors reconstitute the transporter to assess the effects of mutating the binding site residues they identified in their structures. Of the 2 positions tested, only one of them appears to be critical to substrate binding.
Strengths:
The main strength of this work is the capture of the substrate bound state of SiaQM, which provides insight into an important part of the transport cycle.
Weaknesses:
The main weakness is the lack of experimental validation of the structural findings. The authors identified the Neu5Ac binding site, but only test 2 residues for their involvement in substrate interactions, which is quite limited. However, comparison with previous mutagenesis studies on homologues supports the location of the Neu5Ac binding site. The authors tentatively identified a 3rd Na+ binding site, which if true would be an impactful finding, but this site was not sufficiently experimentally tested for its contribution to Na+ dependent transport. This lack of experimental validation prevents the authors from unequivocally assigning this site as a Na+ binding site. However, the reporting of these new data is important as it will facilitate follow up studies by the authors or other researchers.
Comments on revisions:
Overall, the authors have done a good job of addressing the reviewers' comments. It's good to know that the authors are working on the characterisation of the potential metal binding site mutants - characterising just a few of these will provide much needed experimental support for this potential Na+ site.
The new MD simulations provide some additional support for the new Na+ site and could be included. However, as the authors know, direct experimental characterisation of mutants is the ideal evidence of the Na+ site.
Aside from the characterisation of mutants, which seems to be held up by technical issues, the only remaining issue is the comparison of the Na+- and Na+/Neu5Ac-bound states with ASCT2.
It still does not make sense to me why the authors are not directly comparing their Na+ only and Na+/Neu5Ac states with the structures of VcINDY in the Na+-only and Na+/succinate bound states. These VcINDY structures also revealed no conformational changes in the HP loops upon binding succinate, as the authors see for SiaQM. Therefore, this comparison is very supportive. It is understood that the similarity to the DASS structure is mentioned on p.17, but it is also interesting and useful to note that TRAP and DASS transporters also share a lack of substrate-induced local conformational changes, to the extent these things have been measured.
Reviewer #3 (Public review):
The manuscript by Goyal et al report substrate-bound and substrate-free structures of a tripartite ATP independent periplasmic (TRAP) transporter from a previously uncharacterized homolog, F. nucleatum. This is one of most mechanistically fascinating transporter families, by means of its QM domain (the domain reported in his manuscript) operating as a monomeric 'elevator', and its P domain functioning as a substrate-binding 'operator' that is required to deliver the substrate to the QM domain; together, this is termed an 'elevator with an operator' mechanism. Remarkably, previous structures had not demonstrated the substrate Neu5Ac bound. In addition, they confirm the previously reported Na+ binding sites, and report a new metal binding site in the transporter, which seems to be mechanistically relevant. Finally, they mutate the substrate binding site and use proteoliposomal uptake assays to show the mechanistic relevance of the proposed substrate binding residues.
Strengths:
The structures are of good quality, the presentation of the structural data has improved, the functional data is robust, the text is well-written, and the authors are appropriately careful with their interpretations. Determination of a substrate bound structure is an important achievement and fills an important gap in the 'elevator with an operator' mechanism.
Weaknesses:
Although the possibility of the third metal site is compelling, I do not feel it is appropriate to model in a publicly deposited PDB structure without directly confirming experimentally. The authors do not extensively test the binding sites due to technical limitations of producing relevant mutants; however, their model is consistent with genetic assays of previously characterized orthologs, which will be of benefit to the field. Finally, some clarifications of EM processing would be useful to readers, and it would be nice to have a figure visualizing the unmodeled lipid densities - this would be important to contextualize to their proposed mechanism.
Author response:
The following is the authors’ response to the original reviews.
Reviewer #1 (Public Review):
This manuscript reports the substrate-bound structure of SiaQM from F. nucleatum, which is the membrane component of a Neu5Ac-specific Tripartite ATP-dependent Periplasmic (TRAP) transporter. Until recently, there was no experimentally derived structural information regarding the membrane components of the TRAP transporter, limiting our understanding of the transport mechanism. Since 2022, there have been 3 different studies reporting the structures of the membrane components of Neu5Ac-specific TRAP transporters. While it was possible to narrow down the binding site location by comparing the structures to proteins of the same fold, a structure with substrate bound has been missing. In this work, the authors report the Na+-bound state and the Na+ plus Neu5Ac state of FnSiaQM, revealing information regarding substrate coordination. In previous studies, 2 Na+ ion sites were identified. Here, the authors also tentatively assign a 3rd Na+ site. The authors reconstitute the transporter to assess the effects of mutating the binding site residues they identified in their structures. Of the 2 positions tested, only one of them appears to be critical to substrate binding.
Strengths:
The main strength of this work is the capture of the substrate-bound state of SiaQM, which provides insight into an important part of the transport cycle.
Weaknesses:
The main weakness is the lack of experimental validation of the structural findings. The authors identified the Neu5Ac binding site, but only tested 2 residues for their involvement in substrate interactions, which was very limited. The authors tentatively identified a 3rd Na+ binding site, which if true would be an impactful finding, but this site was not tested for its contribution to Na+ dependent transport, and the authors themselves report that the structural evidence is not wholly convincing. This lack of experimental validation undermines the confidence of the findings. However, the reporting of these new data is important as it will facilitate follow-up studies by the authors or other researchers.
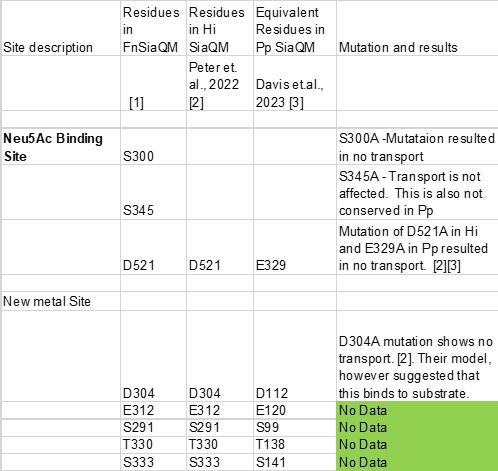
The main concern, also mentioned by other reviewers, is the lack of mutational data and functional studies on the identified binding sites. Two other structures of TRAP transporters have been determined, one from Haemophilus influenzae (Hi) and the other from Photobacterium profundum (Pp). We will refer to the references in this paper as [1], Peter et al. as [2], and Davies et al. as [3]. The table below lists all the mutations made in the Neu5Ac binding site, including direct polar interactions between Neu5Ac and the side chains, as well as the newly identified metal sites.
The structure of Fusobacterium nucleatum (Fn) that we have reported shows a significant sequence identity with the previously reported Hi structure. When we superimpose the Pp and Fn structures, we observe that nearly all the residues that bind to the Neu5Ac and the third metal site are conserved. This suggests that mutagenesis and functional studies from other research can be related to the structure presented in our work.
The table below shows that all three residues that directly interact with Neu5Ac have been tested by site-directed mutagenesis for their role in Neu5Ac transport. Both D521 and S300 are critical for transport, while S345 is not. We do not believe that a mutation of D521A in Fn, followed by transport studies, will provide any new information.
However, Peter et al. have mutated only one of the 5 residues near the newly identified metal binding site, which resulted in no transport. The rest of the residues have not been functionally tested. We propose to mutate these residues into Ala, express and purify the proteins, and then carry out transport assays on those that show expression. We will include this information in the revised manuscript.
Author response table 1.
Reviewer #2 (Public Review):
In this exciting new paper from the Ramaswamy group at Purdue, the authors provide a new structure of the membrane domains of a tripartite ATP-independent periplasmic (TRAP) transporter for the important sugar acid, N-acetylneuraminic acid or sialic acid (Neu5Ac). While there have been a number of other structures in the last couple of years (the first for any TRAP-T) this is the first to trap the structure with Neu5Ac bound to the membrane domains. This is an important breakthrough as in this system the ligand is delivered by a substrate-binding protein (SBP), in this case, called SiaP, where Neu5Ac binding is well studied but the 'hand over' to the membrane component is not clear. The structure of the membrane domains, SiaQM, revealed strong similarities to other SBP-independent Na+-dependent carriers that use an elevator mechanism and have defined Na+ and ligand binding sites. Here they solve the cryo-EM structure of the protein from the bacterial oral pathogen Fusobacterium nucleatum and identify a potential third (and theoretically predicted) Na+ binding site but also locate for the first time the Neu5Ac binding site. While this sits in a region of the protein that one might expect it to sit, based on comparison to other transporters like VcINDY, it provides the first molecular details of the binding site architecture and identifies a key role for Ser300 in the transport process, which their structure suggests coordinates the carboxylate group of Neu5Ac. The work also uses biochemical methods to confirm the transporter from F. nucleatum is active and similar to those used by selected other human and animal pathogens and now provides a framework for the design of inhibitors of these systems.
The strengths of the paper lie in the locating of Neu5Ac bound to SiaQM, providing important new information on how TRAP transporters function. The complementary biochemical analysis also confirms that this is not an atypical system and that the results are likely true for all sialic acid-specific TRAP systems.
The main weakness is the lack of follow-up on the identified binding site in terms of structure-function analysis. While Ser300 is shown to be important, only one other residue is mutated and a much more extensive analysis of the newly identified binding site would have been useful.
Please see the comments above.
Reviewer #3 (Public Review):
The manuscript by Goyal et al reports substrate-bound and substrate-free structures of a tripartite ATP-independent periplasmic (TRAP) transporter from a previously uncharacterized homolog, F. nucleatum. This is one of the most mechanistically fascinating transporter families, by means of its QM domain (the domain reported in his manuscript) operating as a monomeric 'elevator', and its P domain functioning as a substrate-binding 'operator' that is required to deliver the substrate to the QM domain; together, this is termed an 'elevator with an operator' mechanism. Remarkably, previous structures had not demonstrated the substrate Neu5Ac bound. In addition, they confirm the previously reported Na+ binding sites and report a new metal binding site in the transporter, which seems to be mechanistically relevant. Finally, they mutate the substrate binding site and use proteoliposomal uptake assays to show the mechanistic relevance of the proposed substrate binding residues.
The structures are of good quality, the functional data is robust, the text is well-written, and the authors are appropriately careful with their interpretations. Determination of a substrate-bound structure is an important achievement and fills an important gap in the 'elevator with an operator' mechanism. Nevertheless, I have concerns with the data presentation, which in its current state does not intuitively demonstrate the discussed findings. Furthermore, the structural analysis appears limited, and even slight improvements in data processing and resulting resolution would greatly improve the authors' claims. I have several suggestions to hopefully improve the clarity and quality of the manuscript.
We appreciate your feedback and will make the necessary modifications to the manuscript incorporating most of the suggestions. We will submit the revised version once the experiments are completed. We are also working on improving the quality of the figures and have made several attempts to enhance the resolution using CryoSPARC or RELION, but without success. We will continue to explore newer methods in an effort to achieve higher resolution and to model more lipids, particularly in the binding pocket.
Reviewing Editor (Recommendations for the Authors):
After discussing the reviews, the reviewers and reviewing editor have agreed on a list of the most important suggested revisions for the authors, which, if satisfactorily addressed, would improve the assessment of the work. These suggested revisions are listed below. We also include the full Recommendations For The Authors from each of the individual reviewers.
(1) The authors tentatively identified a 3rd Na+ binding site, which if true would be an impactful finding, but this site was not tested for its contribution to Na+ dependent transport, and the authors themselves report that the structural evidence is not wholly convincing. Additional mutagenesis and activity experiments to test the contribution of this site to transport would strengthen the manuscript. Measuring Na+ concentration-response relations and calculating Hill slopes in WT vs. an M site mutant would be a good experiment. Given the lack of functional data and poor density, it does not seem appropriate to build the M site sodium in the PDB model.
The density is well defined to suggest a metal bound (waters would not be clearly defined at this resolution). While our modeling of the site as a Na+ is arbitrary, this was done to satisfy the refinement programs where we have a known scatterer modeled. We could model this density with other metals, but unlike crystallographic refinement, real-space refinement of cryoEM maps does not produce a difference map that might allow us to identify the metal but not conclusively. The density of the maps is good (we have added better figures to demonstrate this). We tried making multiple mutations to test for activity – unfortunately, we are still struggling to express proteins with mutations in this site in sufficient quantities to carry out transport assays.
In the absence of being able to do the experiments, we did MD simulations (carried out by Senwei Quan and Jane Allison at University of Auckland). Our results are shown below – we are not certain without further studies that these should be included in the current paper (we will add them as authors if the editor feels that this evidence is critical).
Author response table 2.
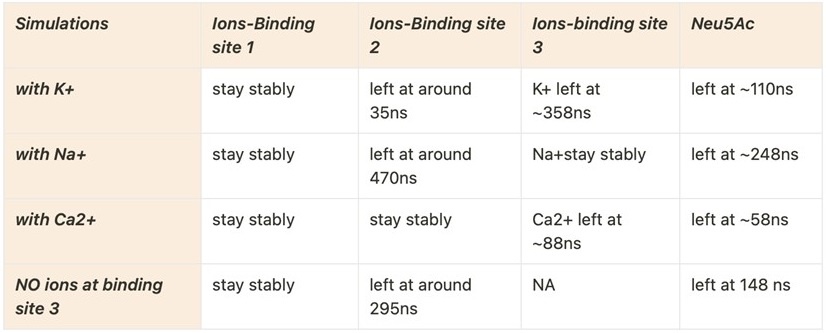
We are showing this for review to suggest that K+, Ca2+, and Na+ were tried, and only Na+ stays stably in the binding pocket. The rest of the results will also have to be explained, which would change the focus of the paper.
We also provided the sequence to Alphafold3 and asked it to identify the possible metal binding sites—when the input was Na+, it found all three binding sites.
Summary: Both our experimental data and computational studies suggest the observed metal binding site is real but at the moment, it is not possible to refine the structure and put an unidentified metal. Computational studies suggest that this is a high-probability Na+ site.
Demonstration of cooperativity between the Na+ site and transport require carrying out these experiments with mutations in these sites in a concentration-dependent manner. Unfortunately, our inability to produce well-expressed and purified proteins with mutations in a short time frame failed.
(2) The authors identified the Neu5Ac binding site but only tested 2 residues for their involvement in substrate interactions, which was very limited. Given that the major highlight of this paper is the identification of the Neu5Ac binding site, it would strengthen the manuscript if the authors provided a more extensive series of mutagenesis experiments - testing at least the effect of D521A would be important. One inconsistency is Ser345 mutagenesis not affecting transport, and the authors should further discuss in the text why they think that is.
D521A has been tested in H. influenzae, and this mutation results in loss of transport. This residue is highly conserved and occupies the same position. We expect the result to remain the same.
We have added a few extra lines to discuss Serine 345: “Ser 345 OG is 3.5Å away from the C1-carboxylate oxygen – a distance that would result in a weak interaction between the two groups. It is, therefore, not surprising that the mutation into Ala did not affect transport. The space created by the mutation can be occupied by a water molecule.”
(3) The purification and assessment of the stability of the protein are described in text alone with no accompanying data. It would be beneficial to include these data (e.g. in the Supplementary info) as it allows the reader to evaluate the protein quality.
This is now added as Supplementary Figure 2.
(4) The structural figures throughout the paper could benefit from more clarity to better support the conclusions. Specific critiques are listed below:
- Figure 1: since the unbound map has a similar reported resolution, displaying the unbound structure's substrate binding site with the same contour would clearly demonstrate that the appearance of this density is substrate-dependent.
- Figure 1: the atomic fit of the ligand to the density, and the suggested coordination by side chain and backbone residues, would be useful in this figure.
- Figure 1: I think it would be more intuitive to compare apo and bound structures with the same local resolution scale.
We have remade Figure 1 “Architecture of FnSiaQM with nanobody. (A and B) Cryo-EM maps of FnSiaQM unliganded and sialic acid bound at 3.2 and 3.17 Å, respectively. The TM domain of FnSiaQM is colored using the rainbow model (N-terminus in blue and C-terminus in red). The nanobody density is colored in purely in red. The density for modeled lipids is colored in tan and the unmodelled density in gray. The figures were made with Chimera at thresholds of 1.2 and 1.3 for the unliganded and sialic acid-bound maps. (C and D) The cytoplasmic view of apo and sialic acid bound FnSiaQM, respectively. Color coding is the same as in panels A and B. The density corresponding to sialic acid and sodium ions are in purple. The substrate binding sites of apo and sialic acid bound FnSiaQM are shown with key residues labeled. The density (blue mesh) around these atoms was made in Pymol with 2 and 1.5 s for the apo and the sialic acid, respectively, with a carve radius of 2 Å.”
The local resolution maps have been moved to Supplementary Figure 3.
- Figure 3, Figure 5a: The mesh structures throughout the manuscript are blocky and very difficult to look at and interpret, especially for the ion binding sites, which are currently suggestive of but not definitively ion densities. Either using transparent surfaces, higher triangle counts, or smoothing the surface might help this.
We have made Figure 3 again with higher triangle counts. We tried all three suggestions and this provided the best figure. We have replaced Figure 5A with density for Neu5Ac and residues around it.
- Figure 5A: It would be important to show the densities of the entire binding pocket, especially coordinating side chains, to show the reader what is and isn't demonstrated by this structure.
- It's not clear how Figure 5D is supposed to show that the cavity can accommodate Neu5Gc, as suggested by the text - please make the discussed cavity clearer in the Figure.
We have now marked with an arrow the Methyl Carbon where the hydroxyl group is added. We have mentioned that in the legend. It is open to the periplasmic side of the cavity.
- Supplementary Figure 4: Please label coordinating residue sites.
Labels have been added to Supplementary Figure 6 which was earlier Supplementary Figure 4.
(5) Intro section: the authors should introduce the work on HiSiaP around the role of the R147 residue in high-affinity Neu5Ac binding, which coordinates the carboxylate of Neu5Ac, and which is a generally conserved mechanism for organic acid binding in other TRAP transporters. This context will help magnify their discovery later that in the membrane domains, it is a key serine and not an arginine that coordinates the carboxylate group (probably as the local concentration of Neu5Ac is high and tight binding site is not desirable for rapid transport, which is mentioned in the discussion).
Thank you for pointing this out. We have added a new sentence to the introduction.
“All the SiaP structures show the presence of a conserved Arginine that binds to the C1-carboxylate of Neu5Ac, and this Arg residue is critical as the high electrostatic affinity may be important to have a strong binding affinity that sequesters the small amounts that reach the bacterial periplasmic space (Glaenzer et al., 2017).”
(6) TRAP transporters exist for many organic compounds and not just sialic acid, which might be nice to make the reader aware of.
We initially did not do this as this is an advance paper and this was discussed in the earlier paper (Currie et. al., 2024). However, we have now added a sentence to the introduction. “Additionally, amino acids, C4-dicarboxylates, aromatic substrates and alpha-keto acids are also transported by TRAP transporters (Vetting et al., 2015). “
(7) On p. 12, the authors describe the Neu5Ac binding site as a large solvent-exposed vestibule, having previously described the substrate-bound state as occluded. These descriptions should be adjusted to make clear which structure is being referenced. The clarity of this would be substantially improved if the authors included a figure that showed this occlusion - currently none of the structure figures clearly demonstrate what the authors are referring to. There are several conspicuous unmodeled densities proximal to the substrate, reminiscent of lipids (in between transport and scaffold domain) and possibly waters/ions. Given this, it is really surprising that the substrate binding site is described as "solvent-exposed" since the larger molecules seem to occlude the pocket. The authors should further process their dataset and discuss the implications of these surrounding densities.
We have processed the data sets carefully both with cryosparc and relion and the resolution described here is same with both software with the cryosparc maps slightly better in terms of interpretability of peripheral helices and described in the manuscript. The current sample (FnTRAP) with the nanobody is a relatively stable sample (in our experience with other similar proteins) as evident from the number of images and particles to achieve a decent resolution and thus the workflow is straightforward and simple. There are number of non-protein densities, which in principle can be modelled but we have chosen a conservative approach not to model these extra densities (except for the two lipids, few ions) due to limit of the resolution. It is possible that increasing the number of particles will result in an increase in resolution but from the estimated B-factor (125 or 135 Å2 for unliganded and liganded), this will certainly require lot of more images with no guarantee of increased resolution.
The question of outward open Vs outward occluded is a valid point. We have now modified this in the manuscript. “The Neu5Ac binding site has a large solvent-exposed vestibule towards the cytoplasmic side, while its periplasmic side is sealed off. Cryo-EM map shows the presence of multiple densities that could be modeled as lipids, possibly preventing the substrate from leaving the transporter. However, the densities are not well defined to model them as specific lipids, hence they have not been modeled. We describe this as the “inward-facing open state” with the substrate-bound.”
(8) On p.15, the activity of FnSiaPQM in liposomes is reported, although the impetus for this study is not clear. Presumably, the reason for its inclusion is to ensure that the structurally characterized protein is active. It would be useful to say this at the start of the section if this is the case. This study nicely shows that the energetics and requirements of transport are identical to all the previous studies on Neu5Ac TRAP transporters - it would be good to acknowledge this somewhere in this section as well.
These changes have been incorporated. We have added a line to say why we did this and added as the last line that this is similar to other SiaPQM’s characterized.
(9) Figure 5C. The authors show the transport activity with and without valinomycin. The authors do not explain the rationale for testing and reporting both conditions for these mutants; an explanation is required, or the data should be simplified. The expected membrane potential induced by valinomycin should be mentioned in the legend.
We have simplified Figure 5C and added the expected membrane potential value.
(10) The authors state that the S300A mutant is inactive. However, unless the authors also measured the background binding/transport of radiolabelled substrate in the absence of protein, then the accuracy of this statement is not clear because Figure 5C does indicate some activity for S300A, albeit much lower than WT. This is an important point in light of the authors' suggestion that the membrane protein does not need a binding site of high affinity or stringent selectivity.
We thank the reviewer for pointing this out we have now added a line in the experimental protocols “The experimental values were corrected by subtracting the control, i.e. the radioactivity taken up in liposomes reconstituted in the absence of protein. The radioactivity associated with the control samples, i.e. empty liposomes was less than 10% with respect to proteoliposomes.”.
(11) There are several issues and important omissions in the work cited:
- It is not normal practice to cite a reference in the abstract and the citation is only to the second structure of HiSiaQM, which does not fairly reflect previous work in the field by only referring to their own work. Also throughout the article, it is normal practice with in-text citations to order them chronologically, i.e. earliest first. Please update this.
This article was submitted as an “Research advance article”. The instructions specifically say that “Research advance article should cite the article in eLife this paper advances. Hence the citation of the “second structure of HiSiaQM”. In fact, in the manuscript we explicitly say “The first structure of _Hi_SiaQM (4.7 Å resolution) demonstrated that it is composed of 15 transmembrane helices and two helical hairpins.” We are following the policy laid out.
Zotero organizes multiple references in alphabetical order, we did not choose to do it that way – the suggestion of bias is not true. The final version of the accepted paper will have numbers, and this argument will automatically be corrected.
- Intro: please cite the primary papers discovering other families of sialic acid transporters.
- Intro: When introducing information on the binding site, dissociation constant of Neu5Ac, and thermodynamics of ligand binding to SiaP, the authors should also include references to the work done by others in addition to their own work.
The Setty et al. paper was the first to demonstrate that the two-component systems are distinct, and that the binding protein of the TRAP system binds enthalpically while the binding protein of the ABC system binds entropically (SiaP vs SatA). As the reviewer points out, this is significant because it highlights how the Arg binding to the carboxylate, which is the enthalpic driver in this case and contributes to the difference between sugar binding to SiaP and SatA. Many studies have published binding affinities of molecules to SiaP, but this paper offers valuable insight into the differences between these systems. We have cited a number of the SiaP papers from other groups, including acknowledging the first structure of SiaP from H. influenzae by Muller et al., in 2006.
- p.5 "TRAP transporters are postulated to employ an elevator-type mechanism...". This postulation has been experimentally tested and published, so should be discussed and referenced (Peter et al. 2024. https://doi.org/10.1038/s41467-023-44327-3).
We have now corrected this error. We removed “are postulated to” and added the reference.
- p.5 "Notably, the transport of Neu5Ac by TRAP transporters requires at least two sodium ions (Davies et al., 2023)." The requirement for at least 2 Na+ ions for Neu5Ac transport was first demonstrated in Mulligan et al. PNAS 2009, so should also be cited (for completion, so should Mulligan et al. JBC. 2012 and Currie et al. elife 2023, which have also shown this requirement is a commonality amongst all Neu5Ac TRAP transporters).
Added.
- P.12, Mulligan et al, JBC, 2012 should be added to the citations in the first sentence.
Added.
- p.19 "Interestingly, even the dicarboxylate transporter from V. cholerae (VcINDY) binds to its ligand via electrostatic interactions with both carboxylate groups". Other references are more appropriate than the one used to support this statement.
Also added references for Mancusso et. al, 2012, Nie et.al, 2017 and Sauer et.al., 2022 here.
- p.19. "The structure of the protein in the outward-facing conformation is unknown". The authors do not discuss the mechanistic findings from Peter et al 2024 Nat Comm here. The work described in that paper revealed an experimentally verified model of the OFS of HiSiaQM, so really needs to be included.
This is not an experimentally determined 3D structure. They have shown the possible existence of this by microscopy, but the structure is not determined. The work mentioned is a wonderful piece of work, but it does not report the three-dimensional structure of the protein in the outward-facing conformation to allow us to understand the nature of the molecular interactions.
- The reference to Kinz-Thompson et al 2022 on p. 6 is not appropriate - neither the HiSiaQM papers nor the PpSiaQM paper makes reference to this work when identifying the binding site. More suitable references are used, for example, Mancusso et al 2012, Nie et al 2017 and Sauer et al 2022; this should be reported accurately.
Added the suggested references. We think the paper (Kinz-Thomposin et al 2022) is relevant and have also kept that reference.
- Garaeva et al report the opposite of what the authors mention - "In the human neutral amino acid transporter (ASCT2), which also uses the elevator mechanism, the HP1 and HP2 loops have been proposed to undergo conformational changes to enable substrate binding and release (Garaeva et al., 2019)." In fact, this paper suggested a one-gate model of transport (HP2), where HP1 seems uninvolved in gating.
The Reviewer is correct. We were wrong and not clear. The entire paragraph has been rewritten.
“While, both the HP1 and HP2 loops have been hypothesized to be involved in gating, in the human neutral amino acid transporter (ASCT2), (which also uses the elevator mechanism), only the HP2 loops have been shown to undergo conformational changes to enable substrate binding and release (Garaeva et al., 2019). Hence, it is suggested that there is a single gate that controls substrate binding. Superposition of the _Pp_SiaQM and _Hi_SiaQM structures do not reveal any change in these loop structures upon substrate binding. For TRAP transporters, the substrate is delivered to the QM protein by the P protein; hence, these loop changes may not play a role in ligand binding or release. This may support the idea that there is minimal substrate specificity within SiaQM and that it will transport the cargo delivered by SiaP, which is more selective.”
- p.19 "suggesting that SSS transporters have probably evolved to transport nine-carbon sugars such as Neu5Ac (Wahlgren et al, 2018)." Surely this goes without saying since Wahlgren et al 2018 demonstrated that SiaT, an SSS, could transport sialic acid? It's unclear why this was included here - perhaps it needs to be rewritten to make the point more clearly, but as it stands, this statement appears self-evident. Furthermore, these proteins can transport all kinds of molecules (see TCDB 2.A.21). This statement needs to be clarified.
This was a comparison to other Neu5Ac binding sites in other Neu5Ac transporters. We have modified the sentence. “The polar groups bind to both the C1-caboxylate side of the molecule and the C8-C9 carbonyls, suggesting that Proteus mirabilis Neu5Ac transporter (SSS type) evolved specifically to transport nine-carbon sugars such as Neu5Ac (Wahlgren et al., 2018)”. These were arguments we were making to suggest that the lack of tight binding could also mean reduced specificity.
- The authors reconstitute the FnSiaQM and measure transport with SiaP, which resembles closely what is known for both HiSiaPQM, VcSiaPQM, which is not cited (https://doi.org/10.1074/jbc.M111.281030).
- Regarding lipids between transport and scaffold domains: there is precedent for such lipids in the elevator transporter GltPh, Wang, and Boudker (eLife 2020) proposed similar displacements during transport and would be appropriate to cite here.
We have now cited the reference to the Mulligan et al., 2012 paper. We also added a sentence on the findings of GltPh paper by Wang and Boudker. Thank you for pointing this out.
(12) p.9 "TRAP transporters, as their name suggests, comprise three units: a substrate-binding protein (SiaP) and two membrane-embedded transporter units (SiaQ and SiaM) (Severi et al., 2007)." This is somewhat odd phrasing because the existence of fused membrane components has been well-documented for a long time. The addition of "Many" at the start of the sentence fixes this.
Added Many.
(13) On p.12 the authors compare the ligand-induced conformational changes of FnSiaQM with ASCT2, citing Garaeva et al, 2019. This comparison does not make sense considering TRAP transporters and ASCT2 do not share a common fold. A far superior comparison is with DASS transporters, which actually do have the same fold as TRAP transporters. And, importantly, the Na+ and substrate-induced conformational changes have been investigated for DASS transporters revealing a unique mechanism likely shared by TRAP transporters (Sauer et al, Nat Comm, 2022). The text on p.12 should be adjusted to replace the ASCT comparison with a VcINDY comparison.
The purpose of citing the ASCT2 paper was only concerning the HP1 and HP2 gates. The authors show that HP2 changes conformation only. Comparing the two FnSiaQM structures – with and without ligand, we see no change in either the HP1 or the HP2 loops. On Page 17, when we describe the structure, we do specifically mention that the overall architecture is similar to VcINDY and the DASS transporters.
(14) p.12 "For TRAP transporters, the substrate is delivered to the QM protein by the SiaP" protein;" "SiaP protein" should be "P protein"
Corrected.
(15) p.18. "periplasmic membrane" should be "cytoplasmic membrane".
Corrected.
(16) p.19. "This prevents Neu5Ac from binding..." There is no evidence for this so this needs to be softened, e.g. "This likely prevents Neu5Ac from...".
Agree – Modified.
(17) Figure 2B is rather small, cramped, and difficult to see. We suggest that the authors make that panel larger, or include it as a stand-alone supplementary figure.
We have moved this figure into a supplementary figure as suggested by the reviewer.
(18) The authors describe the Neu5Ac binding site in SiaQM. It would be helpful if the authors provided a figure in support of the statement that the Neu5Ac binding site architecture is similar to dicarboxylate in VcINDY (especially as Neu5Ac is a monocarboxylate).
The Neu5Ac binding site is NOT similar to the VcINDY binding site. But, we understand the origin of the comment. We have now changed the sentence: “The overall architecture of the Neu5Ac binding site is similar to that of citrate/malate/fumarate in the di/tricarboxylate transporter of V. cholerae (Vc_INDY), but the residues involved in providing specificity are different (Kinz-Thompson _et al., 2022; Mancusso et al., 2012; Nie et al., 2017; Sauer et al., 2022). Neu5Ac binds to the transport domain without direct interactions with the residues in the scaffold domain. The majority of the interactions are with residues in the HP1 and HP2 loops of the transport domain (Figure 5B). Asp521 (HP2), Ser300 (HP1), and Ser345 (helix 5) interact with the substrate through their side chains, except for one interaction between the main chain amino group of residue 301 and the C1-carboxylate oxygen of Neu5Ac. Mutation of the residue equivalent to Asp521 has been shown to result in loss of transport (Peter et al., 2022). To evaluate the role of residues Ser-300 and Ser-345, we mutated them to alanine and performed the transport assays.”
(19) When comparing the binding modes of Neu5Ac to different proteins in Figure 6, it would be helpful to include the structure in this paper as well.
The Neu5Ac binding site is present in figure 5. We would prefer not to show it again in Figure 6.
Additionally, there is a clear binding mode of Neu5Ac in Figure 1 as well.
(20) The manuscript would benefit from a more detailed comparison between Na+-bound (described as apo) and Na+/Neu5Ac structures, especially the prospective gates. If this transporter behaves anything like the archetypical ion-coupled glutamate transporters, some structural changes in the gates might be expected to facilitate transport domain movement when the substrate is loaded, but not when only Na+ is bound. It would be important to discuss and visualize these changes.
We have described in the manuscript that there is NO change in the HP1 and HP2 gates between the unliganded structure and the Neu5Ac bound structure. The major difference we observe is the ordering of the third metal binding site.
A figure comparing the substrate binding pockets between the different high-resolution structures would also be informative. Do the bonding distances between ligands and side chains significantly change between homologs?
This is the only Neu5Ac bound structure. Since the specificity to the substrate comes from the variability of the residues that interact it, we do not believe that this figure would not add much value.
(21) A supplementary figure (or an inset to Figure 2) showing pairwise percent identity between different characterized QM transporters would be useful.
We have now added a Supplementary Figure 4 showing the comparison of the three QM sequences whose structures have been determined.
(22) There is relatively minimal EM processing. More rigorous processing would require relatively little effort and could boost resolution, making this a vastly improved manuscript with a much more confident interpretation of structures.
We described the overall workflow. The processing was rigorous. After obtaining the first maps, we created templates with the structure and did template-based picking. We then did several rounds of 2D classification followed by homogenous refinement, Non-Uniform Refinement. We then made masks and carried out local refinement. We then got the best maps and did a 3D classification. Refined the 3D classes independently. Then, we regrouped them based on how similar they were. We then went back and picked particles again (we used different methods of particle picking, but template-based picking resulted in the final set of particles used) and went through the whole process again. At the end of the refinement, we carried out global and local CTF refinement followed by reference-based motion correction. The final refinement was then done with the Bayesian polished particles. The final refinement was local refinement with a mask over only the transporter and the nano-body. After the reviews came, we tried multi-body refinement in Relion5. It did not improve resolution. We have expanded the legend to supplementary Figure 2 (without listing all the different things we tried). The best resolution we obtained for the structure was 3.1 Å. However, it is important to note that the local resolution of the map around the ligand is good.
We realized this is not easy to depict in a local resolution map. So, we wrote a script to take every atom, then take a radius of 5 Å (again we tried different radii and used the optimal one; we are preparing a manuscript to describe this), take all the local resolution values within the 5 Å spere and average it and add it as B-factor that atom. We have moved the local resolution map figure to the supplement and replaced Figure 1 with a Cartoon, where the color represents the local resolution in which the atom is.
(23) Calling the structure without Neu5Ac bound an "apo" structure is confusing since it indeed has the ligand Na+ present and bound. "Na+" and "Na+/Neu5Ac" structures would be more appropriate.
Changed all “apo” to “unliganded”.



