Peer review process
Revised: This Reviewed Preprint has been revised by the authors in response to the previous round of peer review; the eLife assessment and the public reviews have been updated where necessary by the editors and peer reviewers.
Read more about eLife’s peer review process.Editors
- Reviewing EditorPanayiota PoiraziFORTH Institute of Molecular Biology and Biotechnology, Heraklion, Greece
- Senior EditorPanayiota PoiraziFORTH Institute of Molecular Biology and Biotechnology, Heraklion, Greece
Reviewer #1 (Public review):
Summary:
This study investigated spatial representations in deep feedforward neural network models (DDNs) that were often used in solving vision tasks. The authors create a three-dimensional virtual environment, and let a simulated agent randomly forage in a smaller two-dimensional square area. The agent "sees" images of the room within its field of view from different locations and heading directions. These images were processed by DDNs. Analyzing model neurons in DDNs, they found response properties similar to those of place cells, border cells and head direction cells in various layers of deep nets. A linear readout of network activity can decode key spatial variables. In addition, after removing neurons with strong place/border/head direction selectivity, one can still decode these spatial variables from remaining neurons in the DNNs. Based on these results, the authors argue that that the notion of functional cell types in spatial cognition is misleading.
Comments on the revision:
In the revision, the authors proposed that their model should be interpreted as a null model, rather than the actual model of the spatial navigation system in the brain. In the revision, the authors also argued that the criterion used in the place cell literature was arbitrary. However, the strength of the present work still depends on how well the null model can explain the experimental findings. It seems that currently the null model failed to explain important aspects of the response properties of different functional cell types in the hippocampus.
Strengths:
This paper contains interesting and original ideas, and I enjoy reading it. Most previous studies (e.g., Banino, Nature, 2018; Cueva & Wei, ICLR, 2018; Whittington et al, Cell, 2020) using deep network models to investigate spatial cognition mainly relied on velocity/head rotation inputs, rather than vision (but see Franzius, Sprekeler, Wiskott, PLoS Computational Biology, 2007). Here, the authors find that, under certain settings, visual inputs alone may contain enough information about the agent's location, head direction and distance to the boundary, and such information can be extracted by DNNs. This is an interesting observation from these models.
Weaknesses:
While the findings reported here are interesting, it is unclear whether they are the consequence of the specific model setting and how well they would generalize. Furthermore, I feel the results are over-interpreted. There are major gaps between the results actually shown and the claim about the "superfluousness of cell types in spatial cognition". Evidence directly supporting the overall conclusion seems to be weak at the moment.
Comments on the revision:
The authors showed that the results generalized to different types of networks. The results were generally robust to different types of deep network architectures. This partially addressed my concern. It remains unclear whether the findings would generalize across different types of environment. Regarding this point, the authors argued that the way how they constructed the environment was consistent with the typical experimental setting in studying spatial navigation system in rodents. After the revision, it remains unclear what the implications of the work is for the spatial navigation system in the brain, given that the null model neurons failed to reproduce certain key properties of place cells (although I agreed with the authors that examining such null models are useful and would encourage one to rethink about the approach used to study these neural systems).
Major concerns:
(1) The authors reported that, in their model setting, most neurons throughout the different layers of CNNs show strong spatial selectivity. This is interesting and perhaps also surprising. It would be useful to test/assess this prediction directly based on existing experimental results. It is possible that the particular 2-d virtual environment used is special. The results will be strengthened if similar results hold for other testing environments.
In particular, examining the pictures shown in Fig. 1A, it seems that local walls of the 'box' contain strong oriented features that are distinct across different views. Perhaps the response of oriented visual filters can leverage these features to uniquely determine the spatial variable. This is concerning because this is is a very specific setting that is unlikely to generalize.
[Updated after revision]: This concern is partially addressed in the revision. The authors argued that the way how they constructed the environment is consistent with the typical experimental setting in studying spatial navigation system in rodents.
(2) Previous experimental results suggest that various function cell types discovered in rodent navigation circuits persist in dark environments. If we take the modeling framework presented in this paper literally, the prediction would be that place cells/head direction cells should go away in darkness. This implies that key aspects of functional cell types in the spatial cognition are missing in the current modeling framework. This limitation needs to be addressed or explicitly discussed.
[Updated after revision]: The authors proposed that their model should be treated as a null model, instead of a candidate model for the brain's spatial navigation system. This clarification helps to better position this work. I would like to thank the authors for making this point explicit. However, this doesn't fully address the issues raised. The significance of the reported results still depend on how well the null model can explain the experimental findings. If the null model failed to explain important aspects of the firing properties of functional cell types, that would speak in favor of the usefulness of the concept of functional cell types.
(3) Place cells/border cell/ head direction cells are mostly studied in the rodent's brain. For rodents, it is not clear whether standard DNNs would be good models of their visual systems. It is likely that rodent visual system would not be as powerful in processing visual inputs as the DNNs used in this study.
[Updated after revision]: The authors didn't specifically address this. But clarifying their work as a null model partially addresses this concern.
(4) The overall claim that the functional cell types defined in spatial cognition are superfluous seems to be too strong based on the results reported here. The paper only studied a particular class of models, and arguably, the properties of these models have a major gap to those of real brains. Even though that, in the DNN models simulated in this particular virtual environment, (i) most model neurons have strong spatial selectivity; (ii) removing model neurons with the strongest spatial selectivity still retain substantial spatial information, why is this relevant to the brain? The neural circuits may operate in a very different regime. Perhaps a more reasonable interpretation of the results would be: these results raise the possibility that those strongly selective neurons observed in the brain may not be essential for encoding certain features, as something like this is observed in certain models. It is difficult to draw definitive conclusions about the brain based on the results reported.
[Updated after revision]: The authors clarified that their model should be interpreted as a null model. This partially addresses the concern raised here. However, some concerns remain- it remains unclear what new insights the current work offers in terms of understanding the spatial navigation systems. It seems that this work concerns more about the approach to studying the neural systems. Perhaps this point could be made even more clear.
Reviewer #3 (Public review):
Summary:
In this paper, the authors demonstrate the inevitability of the emergence of spatial information in sufficiently complex systems, even those that are only trained on object recognition (i.e. not a "spatial" system). As such, they present an important null hypothesis that should be taken into consideration for experimental design and data analysis of spatial tuning and its relevance for behavior.
Strengths:
The paper's strengths include the use of a large multi-layer network trained in a detailed visual environment. This illustrates an important message for the field: that spatial tuning can be a result of sensory processing. While this is a historically recognized and often-studied fact in experimental neuroscience, it is made more concrete with the use of a complex sensory network. Indeed, the manuscript is a cautionary tale for experimentalists and computational researchers alike against blindly applying and interpreting metrics without adequate controls. The addition of the deep network, i.e. the argument that sufficient processing increases the likelihood of such a confound, is a novel and important contribution.
Weaknesses:
However, the work has a number of significant weaknesses. Most notably: the spatial tuning that emerges is precisely that we would expect from visually-tuned neurons, and they do not engage with literature that controls for these confounds or compare the quality or degree of spatial tuning with neural data; the ability to linearly decode position from a large number of units is not a strong test of spatial cognition; and the authors make strong but unjustified claims as to the implications of their results in opposition to, as opposed to contributing to, work being done in the field.
The first weakness is that the degree and quality of spatial tuning that emerges in the network is not analyzed to the standards of evidence that have been used in well-controlled studies of spatial tuning in the brain. Specifically, the authors identify place cells, head direction cells, and border cells in their network, and their conjunctive combinations. However, these forms of tuning are the most easily confounded by visual responses, and it's unclear if their results will extend to observed forms of spatial tuning that are not.
For example, consider the head direction cells in Figure 3C. In addition to increased activity in some directions, these cells also have a high degree of spatial nonuniformity, suggesting they are responding to specific visual features of the environment. In contrast, the majority of HD cells in the brain are only very weakly spatially selective, if at all, once an animal's spatial occupancy is accounted for (Taube et al 1990, JNeurosci). While the preferred orientation of these cells are anchored to prominent visual cues, when they rotate with changing visual cues the entire head direction system rotates together (cells' relative orientation relationships are maintained, including those that encode directions facing AWAY from the moved cue), and thus these responses cannot be simply independent sensory-tuned cells responding to the sensory change) (Taube et al 1990 JNeurosci, Zugaro et al 2003 JNeurosci, Ajbi et al 2023).
As another example, the joint selectivity of detected border cells with head direction in Figure 3D suggests that they are "view of a wall from a specific angle" cells. In contrast, experimental work on border cells in the brain has demonstrated that these are robust to changes in the sensory input from the wall (e.g. van Wijngaarden et al 2020), or that many of them are are not directionally selective (Solstad et al 2008).
The most convincing evidence of "spurious" spatial tuning would be the emergence of HD-independent place cells in the network, however, these cells are a very small minority (in contrast to hippocampal data, Thompson and Best 1984 JNeurosci, Rich et al 2014 Science), the examples provided in Figure 3 are significantly more weakly tuned than those observed in the brain.
Indeed, the vast majority of tuned cells in the network are conjunctively selective for HD (Figure 3A). While this conjunctive tuning has been reported, many units in the hippocampus/entorhinal system are not strongly hd selective (Muller et al 1994 JNeurosci, Sangoli et al 2006 Science, Carpenter et al 2023 bioRxiv). Further, many studies have been done to test and understand the nature of sensory influence (e.g. Acharya et al 2016 Cell), and they tend to have a complex relationship with a variety of sensory cues, which cannot readily be explained by straightforward sensory processing (rev: Poucet et al 2000 Rev Neurosci, Plitt and Giocomo 2021 Nat Neuro). E.g. while some place cells are sometimes reported to be directionally selective, this directional selectivity is dependent on behavioral context (Markus et al 1995, JNeurosci), and emerges over time with familiarity to the environment (Navratiloua et al 2012 Front. Neural Circuits). Thus, the question is not whether spatially tuned cells are influenced by sensory information, but whether feed-forward sensory processing alone is sufficient to account for their observed turning properties and responses to sensory manipulations.
These issues indicate a more significant underlying issue of scientific methodology relating to the interpretation of their result and its impact on neuroscientific research. Specifically, in order to make strong claims about experimental data, it is not enough to show that a control (i.e. a null hypothesis) exists, one needs to demonstrate that experimental observations are quantitatively no better than that control.
Where the authors state that "In summary, complex networks that are not spatial systems, coupled with environmental input, appear sufficient to decode spatial information." what they have really shown is that it is possible to decode some degree of spatial information. This is a null hypothesis (that observations of spatial tuning do not reflect a "spatial system"), and the comparison must be made to experimental data to test if the so-called "spatial" networks in the brain have more cells with more reliable spatial info than a complex-visual control.
Further, the authors state that "Consistent with our view, we found no clear relationship between cell type distribution and spatial information in each layer. This raises the possibility that "spatial cells" do not play a pivotal role in spatial tasks as is broadly assumed." Indeed, this would raise such a possibility, if 1) the observations of their network were indeed quantitatively similar to the brain, and 2) the presence of these cells in the brain were the only evidence for their role in spatial tasks. However, 1) the authors have not shown this result in neural data, they've only noticed it in a network and mentioned the POSSIBILITY of a similar thing in the brain, and 2) the "assumption" of the role of spatially tuned cells in spatial tasks is not just from the observation of a few spatially tuned cells. But from many other experiments including causal manipulations (e.g. Robinson et al 2020 Cell, DeLauilleon et al 2015 Nat Neuro), which the authors conveniently ignore. Thus, I do not find their argument, as strongly stated as it is, to be well-supported.
An additional weakness is that linear decoding of position is not a measure of spatial cognition. The ability to decode position from a large number of weakly tuned cells is not surprising. However, based on this ability to decode, the authors claim that "'spatial' cells do not play a privileged role in spatial cognition". To justify this claim, the authors would need to use the network to perform e.g. spatial navigation tasks, then investigate the networks' ability to perform these tasks when tuned cells were lesioned.
Finally, I find a major weakness of the paper to be the framing of the results in opposition to, as opposed to contributing to, the study of spatially tuned cells. For example, the authors state that "If a perception system devoid of a spatial component demonstrates classically spatially-tuned unit representations, such as place, head-direction, and border cells, can "spatial cells" truly be regarded as 'spatial'?" Setting aside the issue of whether the perception system in question does indeed demonstrate spatially-tuned unit representations comparable to those in the brain, I ask "Why not?" This seems to be a semantic game of reading more into a name than is necessarily there. The names (place cells, grid cells, border cells, etc) describe an observation (that cells are observed to fire in certain areas of an animal's environment). They need not be a mechanistic claim (that space "causes" these cells to fire) or even, necessarily, a normative one (these cells are "for" spatial computation). This is evidenced by the fact that even within e.g. the place cell community, there is debate as to these cells' mechanisms and function (eg memory, navigation, etc), or if they can even be said to only serve a single one function. However, they are still referred to as place cells, not as a statement of their function but as a history-dependent label that refers to their observed correlates with experimental variables. Thus, the observation that spatially tuned cells are "inevitable derivatives of any complex system" is itself an interesting finding which contributes to, rather than contradicts, the study of these cells. It seems that the authors have a specific definition in mind when they say that a cell is "truly" "spatial" or that a biological or artificial neural network is a "spatial system", but this definition is not stated, and it is not clear that the terminology used in the field presupposes their definition.
In sum, the authors have demonstrated the existence of a control/null hypothesis for observations of spatially-tuned cells. However, 1) It is not enough to show that a control (null hypothesis) exists, one needs to test if experimental observations are no better than control, in order to make strong claims about experimental data, 2) the authors do not acknowledge the work that has been done in many cases specifically to control for this null hypothesis in experimental work or to test the sensory influences on these cells, and 3) the authors do not rigorously test the degree or source of spatial tuning of their units.
Comments on revisions:
While I'm happy to admit that standards of spatial tuning are not unified or consistent across the field, I do not believe the authors have addressed my primary concern: they have pointed out a null model, and then have constructed a strong opinion around that null model without actually testing if it's sufficient to account for neural data. I've slightly modified my review to that effect.
I do think it would be good for the authors to state in the manuscript what they mean when they say that a cell is "truly" "spatial" or that a biological or artificial neural network is a "spatial system". This is implied throughout, but I was unable to find what would distinguish a "truly" spatial system from a "superfluous" one.
Author response:
The following is the authors’ response to the original reviews.
Reviewer #1 (Public Review):
but see Franzius, Sprekeler, Wiskott, PLoS Computational Biology, 2007
We have discussed the differences with this work in the response to Editor recommendations above.
While the findings reported here are interesting, it is unclear whether they are the consequence of the specific model setting, and how well they would generalize.
We have considered deep vision models across different architectures in our paper, which include traditional feedforward convolutional neural networks (VGG-16), convolutional neural networks with skip connections (ResNet-50) and the Vision Transformer (VIT) which employs self-attention instead of convolution as its core information processing unit.
In particular, examining the pictures shown in Fig. 1A, it seems that local walls of the ’box’ contain strong oriented features that are distinct across different views. Perhaps the response of oriented visual filters can leverage these features to uniquely determine the spatial variable. This is concerning because this is a very specific setting that is unlikely to generalize.
The experimental set up is based on experimental studies of spatial cognition in rodents. They are typically foraging in square or circular environments. Indeed, square environments will have more borders and corners that will provide information about the spatial environment, which is true in both empirical studies and our simulations. In any navigation task, and especially more realistic environments, visual information such as borders or landmarks likely play a major role in spatial information available to the agent. In fact, studies that do not consider sensory information to contribute to spatial information are likely missing a major part of how animals navigate.
The prediction would be that place cells/head direction cells should go away in darkness. This implies that key aspects of functional cell types in the spatial cognition are missing in the current modeling framework.
We addressed this comment in our response to the editor’s highlight. To briefly recap, we do not intend to propose a comprehensive model of the brain that captures all spatial phenomena, as we would not expect this from an object recognition network. Instead, we show that such a simple and nonspatial model can reproduce key signatures of spatial cells, raising important questions about how we interpret spatial cell types that dominate current research.
Reviewer #2 (Public Review):
The network used in the paper is still guided by a spatial error signal [...] one could say that the authors are in some way hacking this architecture and turning it into a spatial navigation one through learning.
To be clear, the base networks we use do not undergo spatial error training. They have either been pre-trained on image classification tasks or are untrained. We used a standard neuroscience approach: training linear decoders on representations to assess the spatial information present in the network layers. The higher decoding errors in early layer representations (Fig. 2A) indicate that spatial information differs across layers—an effect that cannot be attributed to the linear decoder alone.
My question is whether the paper is fighting an already won battle.
Intuitive cell type discovery are still being celebrated. Concentrating on this kind of cell type discovery has broader implications that could be deleterious to the future of science. One point to note is that this issue depends on the area or subfield of neuroscience. In some subfields, papers that claim to find cell types with a strong claim of specific functions are relatively rare, and population coding is common (e.g., cognitive control in primate prefrontal cortex, neural dynamics of motor control). Although rodent neuroscience as a field is increasingly adopting population approaches, influential researchers and labs are still publishing “cell types” and in top journals (here are a few from 2017-2024: Goal cells (Sarel et al., 2017), Object-vector cells (Høydal et al., 2019), 3D place cells (Grieves et al., 2020), Lap cells (Sun et al., 2020), Goal-vector cells (Ormond and O’Keefe, 2022), Predictive grid cells (Ouchi and Fujisawa, 2024).
In some cases, identification of cell types is only considered a part of the story, and there are analyses on behavior, neural populations, and inactivationbased studies. However, our view (and suggest this is shared amongst most researchers) is that a major reason these papers are reviewed and accepted to top journals is because they have a simple, intuitive “cell type” discovery headline, even if it is not the key finding or analysis that supports the insightful aspects of the work. This is unnecessary and misleading to students of neuroscience, related fields, and the public, it affects private and public funding priorities and in turn the future of science. Worse, it could lead the field down the wrong path, or at the least distribute attention and resources to methods and papers that could be providing deeper insights. Consistent with the central message of our work, we believe the field should prioritize theoretical and functional insights over the discovery of new “cell types”.
Reviewer #3 (Public Review):
The ability to linearly decode position from a large number of units is not a strong test of spatial information, nor is it a measure of spatial cognition
Using a linear decoder to test what information is contained in a population of neurons available for downstream areas is a common technique in neuroscience (Tong and Pratte, 2012; DiCarlo et al., 2012) including spatial cells (e.g., Diehl et al. 2017; Horrocks et al. 2024). A linear decoder is used because it is a direct mapping from neurons to potential output behavior. In other words, it only needs to learn some mapping to link one set of neurons to another set which can “read out” the information. As such, it is a measure of the information contained in the population, and it is a lower bound of the information contained - as both biological and artificial neurons can do more complex nonlinear operations (as the activation function is nonlinear).
We understand the reviewer may understand this concept but we explain it here to justify our position and for completeness of this public review.
For example, consider the head direction cells in Figure 3C. In addition to increased activity in some directions, these cells also have a high degree of spatial nonuniformity, suggesting they are responding to specific visual features of the environment. In contrast, the majority of HD cells in the brain are only very weakly spatially selective, if at all, once an animal’s spatial occupancy is accounted for (Taube et al 1990, JNeurosci). While the preferred orientation of these cells are anchored to prominent visual cues, when they rotate with changing visual cues the entire head direction system rotates together (cells’ relative orientation relationships are maintained, including those that encode directions facing AWAY from the moved cue), and thus these responses cannot be simply independent sensory-tuned cells responding to the sensory change) (Taube et al 1990 JNeurosci, Zugaro et al 2003 JNeurosci, Ajbi et al 2023).
As we have noted in our response to the editor, one of the main issues is how the criteria to assess what they are interested in is created in a subjective, and biased way, in a circular fashion (seeing spatial-like responses, developing criteria to determine a spatial response, select a threshold).
All the examples the reviewer provides concentrate on strict criteria developed after finding such cells. What is the purpose of these cells for function, for behavior? Just finding a cell that looks like it is tuned to something does not explain its function. Neuroscience began with tuning curves in part due to methodological constraints, which was a promising start, but we propose that this is not the way forward.
The metrics used by the authors to quantify place cell tuning are not clearly defined in the methods, but do not seem to be as stringent as those commonly used in real data. (e.g. spatial information, Skaggs et al 1992 NeurIPS).
We identified place cells following the definition from Tanni et al. (2022), by one of the leading labs in the field. Since neurons in DNNs lack spikes, we adapted their criteria by focusing on the number of spatial bins in the ratemap rather than spike-based measures. However, our central argument is that the very act of defining spatial cells is problematic. Researchers set out to find place cells to study spatial representations, find spatially selective cells with subjective, qualitative criteria (sometimes combined with prior quantitative criteria, also subjectively defined), then try to fine-tune the criteria to more “stringent” criteria, depending on the experimental data at hand. It is not uncommon to see methodological sections that use qualitative judgments, such as: “To avoid bias ... we applied a loose criteria for place cells” Tanaka et al. (2018) , which reflects the lack of clarity for and subjectivity of place cell selection criteria.
A simple literature survey reveals inconsistent criteria across studies. For place field selection, Dombeck et al. (2010) required mean firing rates exceeding 25% of peak rate, while Tanaka et al. (2018) used a 20% threshold. Speed thresholds also vary dramatically: Dombeck et al. (2010) calculated firing rates only when mice moved faster than 8.3 cm/s, whereas Tanaka et al. (2018) used 2 cm/s. Additional criteria differ further: Tanaka et al. (2018) required firing rates between 1-10 Hz and excluded cells with place fields larger than 1/3 of the area, while Dombeck et al. (2010) selected fields above 1.5 Hz, and Tanni et al. (2022) used a 10 spatial bins to 1/2 area threshold. As Dombeck et al. (2010) noted, differences in recording methods and place field definitions lead to varying numbers of identified place cells. Moreover, Grijseels et al. (2021) demonstrated that different detection methods produce vastly different place cell counts with minimal overlap between identified populations.
This reflects a deeper issue. Unlike structurally and genetically defined cell types (e.g., pyramidal neurons, interneurons, dopamingeric neurons, cFos expressing neurons), spatial cells lack such clarity in terms of structural or functional specialization and it is unclear whether such “cell types” should be considered cell types in the same way. While scientific progress requires standardized definitions, the question remains whether defining spatial cells through myriad different criteria advances our understanding of spatial cognition. Are researchers finding the same cells? Could they be targeting different populations? Are they missing cells crucial for spatial cognition that they exclude due to the criteria used? We think this is likely. The inconsistency matters because different criteria may capture genuinely different neural populations or computational processes.
Variability in definitions and criteria is an issue in any field. However, as we have stated, the deeper issue is whether we should be defining and selecting these cells at all before commencing analysis. By defining and restricting to spatial “cell types”, we risk comparing fundamentally different phenomena across studies, and worse, missing the fundamental unit of spatial cognition (e.g., the population).
We have added a paragraph in Discussion (lines 357-366) noting the inconsistency in place cell selection criteria in the literature and the consequences of using varying criteria.
We have also added a sentence (lines 354-356) raising the comparison of functionally defined spatial cell types with structurally and genetically defined cell types in the Discussion.
Thus, the question is not whether spatially tuned cells are influenced by sensory information, but whether feed-forward sensory processing alone is sufficient to account for their observed turning properties and responses to sensory manipulations.
These issues indicate a more significant underlying issue of scientific methodology relating to the interpretation of their result and its impact on neuroscientific research. Specifically, in order to make strong claims about experimental data, it is not enough to show that a control (i.e. a null hypothesis) exists, one needs to demonstrate that experimental observations are quantitatively no better than that control.
Where the authors state that ”In summary, complex networks that are not spatial systems, coupled with environmental input, appear sufficient to decode spatial information.” what they have really shown is that it is possible to decode *some degree* of spatial information. This is a null hypothesis (that observations of spatial tuning do not reflect a ”spatial system”), and the comparison must be made to experimental data to test if the so-called ”spatial” networks in the brain have more cells with more reliable spatial info than a complex-visual control.
We agree that good null hypotheses with quantitative comparisons are important. However, it is not clear that researchers in the field have not been using a null hypothesis, rather they make the assumption that these cell types exist and are functional in the way they assume. We provide one null hypothesis. The field can and should develop more and stronger null hypotheses.
In our work, we are mainly focusing on criteria of finding spatial cells, and making the argument that simply doing this is misleading. Researcher develop criteria and find such cells, but often do not go further to assess whether they are real cell “types”, especially if they exclude other cells which can be misleading if other cells also play a role in the function of interest.
But from many other experiments including causal manipulations (e.g. Robinson et al 2020 Cell, DeLauilleon et al 2015 Nat Neuro), which the authors conveniently ignore. Thus, I do not find their argument, as strongly stated as it is, to be well-supported.
We acknowledge that there are several studies that have performed inactivation studies that suggest a strong role for place cells in spatial behavior. Most studies do not conduct comprehensive analyses to confirm that their place cells are in fact crucial for the behavior at hand.
One question is how the criteria were determined. Did the researchers make their criteria based on what “worked”, so they did not exclude cells relevant to the behavior? What if their criteria were different, then the argument could have been that non-place cells also contribute to behavior.
Another question is whether these cells are the same kinds of cells across studies and animals, given the varied criteria across studies? As most studies do not follow the same procedures, it is unclear whether we can generalize these results across cells and indeed, across task and spatial environments.
Finally, does the fact that the place cells – the strongly selective cells with a place field – have a strong role in navigation provide any insight into the mechanism? Identifying cells by itself does not contribute to our understanding of how they work. Consistent with our main message, we argue that performing analyses and building computational models that uncover how the function of interest works is more valuable than simply naming cells.
Finally, I find a major weakness of the paper to be the framing of the results in opposition to, as opposed to contributing to, the study of spatially tuned cells. For example, the authors state that ”If a perception system devoid of a spatial component demonstrates classically spatially-tuned unit representations, such as place, head-direction, and border cells, can ”spatial cells” truly be regarded as ’spatial’?” Setting aside the issue of whether the perception system in question does indeed demonstrate spatiallytuned unit representations comparable to those in the brain, I ask ”Why not?” This seems to be a semantic game of reading more into a name then is necessarily there. The names (place cells, grid cells, border cells, etc) describe an observation (that cells are observed to fire in certain areas of an animal’s environment). They need not be a mechanistic claim... This is evidenced by the fact that even within e.g. the place cell community, there is debate about these cells’ mechanisms and function (eg memory, navigation, etc), or if they can even be said to serve only a single function. However, they are still referred to as place cells, not as a statement of their function but as a history-dependent label that refers to their observed correlates with experimental variables. Thus, the observation that spatially tuned cells are ”inevitable derivatives of any complex system” is itself an interesting finding which *contributes to*, rather than contradicts, the study of these cells. It seems that the authors have a specific definition in mind when they say that a cell is ”truly” ”spatial” or that a biological or artificial neural network is a ”spatial system”, but this definition is not stated, and it is not clear that the terminology used in the field presupposes their definition.
We have to agree to disagree with the reviewer on this point. Although researchers may reflect on their work and discuss what the mechanistic role of these cells are, it is widely perceived that cell type discovery is perceived as important to journals and funders due to its intuitive appeal and easy-tounderstand impact – even if there is no finding of interest to be reported. As noted in the comment above, papers claiming cell type discovery continue to be published in top journals and is continued to be funded.
Our argument is that maybe “cell type” discovery research should not celebrated in the way it is, and in fact they shouldn’t be discovered when they are not genuine cell types like structural or genetic cell types. By using this term it make it appear like they are something they are not, which is misleading. They may be important cells, but providing a name like a “place” cell also suggests other cells are not encoding space - which is very unlikely to be true.
In sum, our view is that finding and naming cells through a flawed theoretical lens that may not actually function as their names suggests can lead us down the wrong path and be detrimental to science.
Reviewer #1 (Recommendations For The Authors):
The novelty of the current study relative to the work by Franzius, Sprekeler, Wiskott (PLoS Computational Biology, 2007) needs to be carefully addressed. That study also modeled the spatial correlates based on visual inputs.
Our work differs from Franzius et al. (2007) on both theoretical and experimental fronts. While both studies challenge the mechanisms underlying spatial cell formation, our theoretical contributions diverge. Franzius et al. (2007) assume spatial cells are inherently important for spatial cognition and propose a sensory-driven computational mechanism as an alternative to mainstream path integration frameworks for how spatial cells arise and support spatial cognition. In contrast, we challenge the notion that spatial cells are special at all. Using a model with no spatial grounding, we demonstrate that 1) spatial cells as naturally emerge from complex non-linear processing and 2) are not particularly useful for spatial decoding tasks, suggesting they are not crucial for spatial cognition.
Our approach employs null models with fixed weights—either pretrained on classification tasks or entirely random—that process visual information non-sequentially. These models serve as general-purpose information processors without spatial grounding. In contrast, Franzius et al. (2007)’s model learns directly from environmental visual information, and the emergence of spatial cells (place or head-direction cells) in their framework depends on input statistics, such as rotation and translation speeds. Notably, their model does not simultaneously generate both place and head-direction cells; the outcome varies with the relative speed of rotation versus translation. Their sensory-driven model indirectly incorporates motion information through learning, exhibiting a time-dependence influenced by slow-feature analysis.
Conversely, our model simultaneously produces units with place and headdirection cell profiles by processing visual inputs sampled randomly across locations and angles, independent of temporal or motion-related factors. This positions our model as a more general and fundamental null hypothesis, ideal for challenging prevailing theories on spatial cells due to its complete lack of spatial or motion grounding.
Finally, unlike Franzius et al. (2007), who do not evaluate the functional utility of their spatial representations, we test whether the emergent spatial cells are useful for spatial decoding. We find that not only do spatial cells emerge in our non-spatial model, but they also fail to significantly aid in location or head-direction decoding. This is the central contribution of our work: spatial cells can arise without spatial or sensory grounding, and their functional relevance is limited. We have updated the manuscript to clarify the novelty of the current contribution to previous work (lines 324-335).
In Fig. 2, it may be useful to plot the error in absolute units, rather than the normalized error. The direction decoding can be quantified in terms of degree Also, it would be helpful to compare the accuracy of spatial localization to that of the actual place cells in rodents.
We argue it makes more sense and put comparison in perspective when we normalize the error by dividing the maximal error possible under each task. For transparency, we plot the errors in absolute physical units used by the Unity game engine in the updated Appendix (Fig. 1).
Reviewer #2 (Recommendations For The Authors):
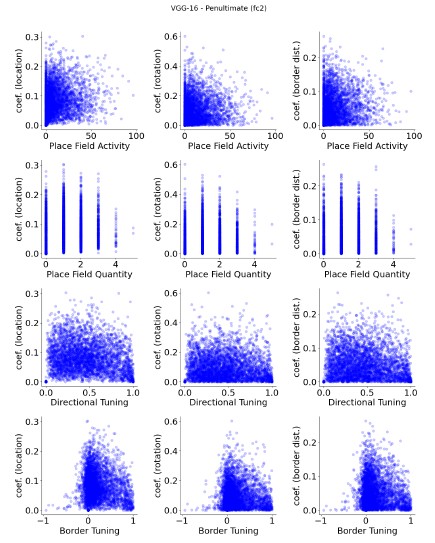
Regarding the involvement of ’classified cells’ in decoding, I think a useful way to present the results would be to show the relationship between ’placeness’, ’directioness’ and ’borderness’ and the strength of the decoder weights. Either as a correlation or as a full scatter plot.
We appreciate your suggestion to visualize the relationship between units’ spatial properties and their corresponding decoder weights. We believe it would be an important addition to our existing results. Based on the exclusion analyses, we anticipated the correlation to be low, and the additional results support this expectation.
As an example, we present unit plots below for VGG-16 (pre-trained and untrained, at its penultimate layer with sampling rate equals 0.3; Author response image 1 and 2). Additional plots for various layers and across models are included in the supplementary materials (Fig. S12-S28). Consistently across conditions, we observed no significant correlations between units’ spatial properties (e.g., placeness) and their decoding weight strengths. These results further corroborate the conclusions drawn from our exclusion analyses.
Reviewer #3 (Recommendations For The Authors):
My main suggestions are that the authors: -perform manipulations to the sensory environment similar to those done in experimental work, and report if their tuned cells respond in similar ways -quantitatively compare the degree of spatial tuning in their networks to that seen in publicly available data -re-frame the discussion of their results to critically engage with and contribute to the field and its past work on sensory influences to these cells
As we noted in our opening section, our model is not intended as a model of the brain. It is a non-spatial null model, and we present the surprising finding that even such a model contains spatial cell-like units if identified using criteria typically used in the field. This raises the question whether simply finding cells that show spatial properties is sufficient to grant the special status of “cell type” that is involved in the brain function of interest.
Author response image 1.
VGG-16 (pre-trained), penultimate layer units, show no apparent relationship between spatial properties and their decoder weight strengths.
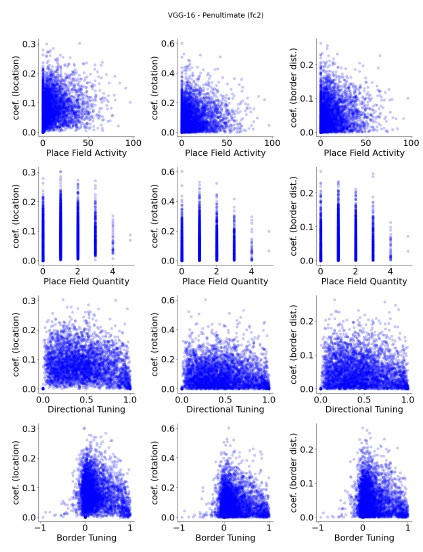
Author response image 2.
VGG-16 (untrained), penultimate layer units, show no apparent relationship between spatial properties and their decoder weight strengths.
Furthermore, our main simulations were designed to be compared to experimental work where rodents foraged around square environments in the lab. We did not do an extensive set of simulations as the purpose of our study is not to show that we capture exactly every single experimental finding, but rather raise the issues with the functional cell type definition and identification approach for progressing neuroscientific knowledge.
Finally, as we note in more detail below, different labs use different criteria for identifying spatial cells, which depend both on the lab and the experimental design. Our point is that we can identify such cells using criteria set by neuroscientists, and that such cell types may not reflect any special status in spatial processing. Additional simulations that show less alignment with certain datasets will not provide support for or against our general message.
References
Banino A, Barry C, Uria B, Blundell C, Lillicrap T, Mirowski P, Pritzel A, Chadwick MJ, Degris T, Modayil J, Wayne G, Soyer H, Viola F, Zhang B, Goroshin R, Rabinowitz N, Pascanu R, Beattie C, Petersen S, Sadik A, Gaffney S, King H, Kavukcuoglu K, Hassabis D, Hadsell R, Kumaran D (2018) Vector-based navigation using grid-like representations in artificial agents. Nature 557(7705):429–433, DOI 10.1038/s41586-018-0102-6, URL http://www.nature.com/articles/s41586-018-0102-6
DiCarlo JJ, Zoccolan D, Rust NC (2012) How Does the Brain Solve Visual Object Recognition? Neuron 73(3):415–434, DOI 10.1016/J.NEURON.2012.01.010, URL https://www.cell.com/neuron/fulltext/S0896-6273(12)00092-X
Diehl GW, Hon OJ, Leutgeb S, Leutgeb JK (2017) Grid and Nongrid Cells in Medial Entorhinal Cortex Represent Spatial Location and Environmental Features with Complementary Coding Schemes. Neuron 94(1):83– 92.e6, DOI 10.1016/j.neuron.2017.03.004, URL https://linkinghub.elsevier.com/retrieve/pii/S0896627317301873
Dombeck DA, Harvey CD, Tian L, Looger LL, Tank DW (2010) Functional imaging of hippocampal place cells at cellular resolution during virtual navigation. Nature Neuroscience 13(11):1433–1440, DOI 10.1038/nn.2648, URL https://www.nature.com/articles/nn.2648
Ebitz RB, Hayden BY (2021) The population doctrine in cognitive neuroscience. Neuron 109(19):3055–3068, DOI 10.1016/j.neuron. 2021.07.011, URL https://linkinghub.elsevier.com/retrieve/pii/S0896627321005213
Grieves RM, Jedidi-Ayoub S, Mishchanchuk K, Liu A, Renaudineau S, Jeffery KJ (2020) The place-cell representation of volumetric space in rats. Nature Communications 11(1):789, DOI 10.1038/s41467-020-14611-7, URL https://www.nature.com/articles/s41467-020-14611-7
Grijseels DM, Shaw K, Barry C, Hall CN (2021) Choice of method of place cell classification determines the population of cells identified. PLOS Computational Biology 17(7):e1008835, DOI 10.1371/journal.pcbi.1008835, URL https://dx.plos.org/10.1371/journal.pcbi.1008835
Horrocks EAB, Rodrigues FR, Saleem AB (2024) Flexible neural population dynamics govern the speed and stability of sensory encoding in mouse visual cortex. Nature Communications 15(1):6415, DOI 10.1038/s41467-024-50563-y, URL https://www.nature.com/articles/s41467-024-50563-y
Høydal , Skytøen ER, Andersson SO, Moser MB, Moser EI (2019) Objectvector coding in the medial entorhinal cortex. Nature 568(7752):400– 404, DOI 10.1038/s41586-019-1077-7, URL https://www.nature.com/articles/s41586-019-1077-7
Ormond J, O’Keefe J (2022) Hippocampal place cells have goal-oriented vector fields during navigation. Nature 607(7920):741–746, DOI 10.1038/s41586-022-04913-9, URL https://www.nature.com/articles/s41586-022-04913-9
Ouchi A, Fujisawa S (2024) Predictive grid coding in the medial entorhinal cortex. Science 385(6710):776–784, DOI 10.1126/science.ado4166, URL https://www.science.org/doi/10.1126/science.ado4166
Sarel A, Finkelstein A, Las L, Ulanovsky N (2017) Vectorial representation of spatial goals in the hippocampus of bats. Science 355(6321):176–180, DOI 10.1126/science.aak9589, URL https://www.science.org/doi/10.1126/science.aak9589
Sun C, Yang W, Martin J, Tonegawa S (2020) Hippocampal neurons represent events as transferable units of experience. Nature Neuroscience 23(5):651–663, DOI 10.1038/s41593-020-0614-x, URL https://www.nature.com/articles/s41593-020-0614-x
Tanaka KZ, He H, Tomar A, Niisato K, Huang AJY, McHugh TJ (2018) The hippocampal engram maps experience but not place. Science 361(6400):392–397, DOI 10.1126/science.aat5397, URL https://www.science.org/doi/10.1126/science.aat5397
Tanni S, De Cothi W, Barry C (2022) State transitions in the statistically stable place cell population correspond to rate of perceptual change. Current Biology 32(16):3505–3514.e7, DOI 10.1016/j.cub. 2022.06.046, URL https://linkinghub.elsevier.com/retrieve/pii/S0960982222010089
Tong F, Pratte MS (2012) Decoding Patterns of Human Brain Activity. Annual Review of Psychology 63(1):483–509, DOI 10.1146/annurev-psych-120710-100412, URL https://www.annualreviews.org/doi/10.1146/annurev-psych-120710-100412



