Peer review process
Not revised: This Reviewed Preprint includes the authors’ original preprint (without revision), an eLife assessment, public reviews, and a provisional response from the authors.
Read more about eLife’s peer review process.Editors
- Reviewing EditorLeopoldo PetreanuChampalimaud Center for the Unknown, Lisbon, Portugal
- Senior EditorJohn HuguenardStanford University School of Medicine, Stanford, United States of America
Reviewer #1 (Public Review):
Summary:
The manuscript by Tuberosa et al outlines the generation of a new set of transgenic mice that express different recombinases specifically in Smim32 positive cells. They show that Smim32 is a useful marker of the mouse claustrum. Therefore, these mice could be useful for functional studies focused on measuring claustrum activity or manipulating the claustrum using optogenetic and pharmacogenetic tools.
Strengths:
The manuscript provides a new genetic approach to target claustrum neurons, using Smim32. The work may help future studies where claustrum excitatory neurons are measured or manipulated.
Weaknesses:
A toolbox is only useful if others can use it. Therefore, these mice should be made available to the community through commercial vendors. Without this added step, this toolbox and method does not provide any utility to the research community.
The data presented and quantified in each figure subpanel are from N = 1 mouse. This is not acceptable or conventional. Replication is an important aspect of any paper, and currently, there are no replicates contained in the manuscript. Additional examples of female mice should also be included and separately quantified. Mice from different litters should be used for replicates.
Given the preliminary nature of these data from the minimum possible number of mice, a better characterization of all data should be undertaken.
The tone of the paper implies that this is the superior way to locate the claustrum. A more balanced discussion of the strengths and weaknesses of these mice should be included. Several sentences highlighting the shortfalls of other approaches are overstated and should be toned down.
Reviewer #2 (Public Review):
Rodent studies of claustrum are complicated by the tube-like shape of this nucleus. As such, judicious viral strategies alone or in combination with existing Cre driver lines (Egr2, Gnb4, Slc17a6, and Tbx21) represent the current gold standard for claustrum structure and function investigation. Any improvement in tools that would allow better genetic access to the claustrum are always desired, as with any nucleus in brain. This paper describes the expression pattern of the gene Smim32 and characterization of new mouse transgenic lines expressing Cre/Flp recombinase driven by the Smim32 promoter. The authors should be applauded for the work to develop these new tools presented in the study. Overall, the strengths of the paper lie in the development of new mouse lines that are well-characterized in comparison to other molecular markers of the claustrum. Weaknesses lie in poor anatomical definitions of the claustrum (and endopiriform nucleus). Smim32 expression is used to define claustrum anatomical boundaries, rather than first using several structural, molecular, and connectivity lines of evidence to define the claustrum anatomically and then to assess whether Smim32 expression fits within this anatomical definition. Another major weakness is the fact that Cre/Flp expression driven by the Smim32 promoter is present in non-claustrum regions, including the neighboring cortex, striatum, and endopiriform nucleus as well as the more distant thalamic reticular nucleus. Despite this, the conclusion of the study, as communicated by the authors, is that selective interrogation of the claustrum is now possible with these Smim32-based tools. Therefore, the data do not support the claims and conclusions.
Very concerning is problematic language in the abstract and introduction sections that diminish the impact of several published studies (not cited) that have led to important findings regarding claustrum function. The authors Create an argument that all the research performed thus far on the claustrum is unreliable because targeting the structure has been sub-optimal. This is definitely not the case for several studies from multiple labs. If investigators new to the claustrum were to read this paper, they would conclude that all previous data hold little-to-no value and that using these tools set forth the possibility, at long last, to solve claustrum structural and functional queries. Here is an example from the abstract of the problematic language: "However, research on the CLA has been challenging due to difficulties in specifically and comprehensively targeting its neuronal populations. In various cases, this limitation has led to inconsistent findings and a lack of reliable data." (no references cited). Since Smim32 driven recombinase (in 61 or 62lrod) is not exclusively expressed in the claustrum, it is not clear how Smim32 is an advantage over possible Nr4a2 or, the more selective, GNB4 Cre driver lines. Taken together, the goal of the study as articulated in the Introduction: "Our goal was here to generate genetic tools capable of targeting the majority of mouse CLA projection neurons without affecting other brain cell populations, or tissues outside the brain" has not been met and, therefore, the conclusion of the study based on the data "With these genetic tools in hand, the comprehensive targeting and functional probing of the densely connected CLA is now possible" is unfortunately also unmet.
The manuscript does convincingly show that Smim32 targets excitatory neurons in the claustrum as evidenced by exclusive overlap of Smim32 expression with Vglut2 and not GAD (fig 1 and suppl fig 1). Additionally, the manuscript provides sufficient evidence that neurons in the claustrum area expressing Smim32 further co-express a number of other molecular markers of claustrum, including Nr4a2 (fig1), Lxn, Gnb4, and Oprk1 (fig 2), and Slc17a6 (suppl fig 1). The authors further show that Smim32 is not co-expressed with molecular markers of layer VI cortex like Ctgf and Rprm (fig 2). However, by limiting the line of evidence to molecular expression, the study fails to escape the limitations of molecular markers, which cannot by themselves be used to define the anatomical boundary of the claustrum. The expression of several of these markers in the neighboring endopiriform nucleus, including Smim32, is evidence that using molecular markers as a sole indicator of the anatomy of the claustrum is not warranted.
While the anatomical boundaries of the claustrum remain somewhat debated, several standards have emerged to delineate claustrum boundaries. These include immunoreactivity against Gng2 (or PV, especially in rat) to indicate claustrum or against Crym to counter-indicate claustrum. In addition, injection of retrograde tracers into the anterior cingulate cortex or retrosplenial cortex, for example, results in selective targeting of (large) subpopulations of claustrum neurons that help define claustrum location. Further targeting of neurons projecting to the anterior insula or thalamus has been used to delineate the boundaries of what some consider the claustrum shell and others consider the deep layers of the insula. The use of any of these approaches to delineate the claustrum anatomy should be used to describe the spatial distribution of Smim32 and Cre or FlpO in the transgenic lines.
The manuscript provides a description of Smim32 promoter-driven tdTomato in the three transgenic Cre lines during development. This shows strong expression in claustrum and not in surrounding regions. However, as the claustrum borders are not distinct without markers, the anatomical boundary of claustrum for this analysis is deemed arbitrary - an issue that is exacerbated when looking at the developing brain where atlases are less precise and boundaries of the claustrum are ill-defined.
Reviewer #3 (Public Review):
Summary:
In the manuscript by Tuberosa et al., the authors set out to identify a genetic marker for the claustrum to create transgenic mice as tools to study this challenging brain region. To achieve this, the authors first re-analyzed published scRNAseq datasets from mouse frontal cortex and identified a unique cluster expressing Smim32, which correlated with Nr4a2, a previously reported claustrum marker (though also expressed in layer 6 and elsewhere). Importantly, Smim32 was also found to strongly express in the layer 6 and the thalamic reticular nucleus (with weaker expression in other parts of cortex, striatum, thalamus, olfactory bulb and more). The authors then extensively characterize Smim32 expression relative to a few other genes associated with claustrum and layer 6, as well as creating several novel transgenic mice focused on the Smim32 gene.
Strengths:
The main strength of the paper is the well done scRNAseq analysis, the beautiful ISH images/reconstructions, and the assessment of gene expression throughout development. The main value of this paper is adding the Smim32 gene to the list of markers expressed in the claustrum, though it is not specific to the claustrum, showing extensive expression in TRN and layer 6 of cortex.
Weaknesses:
The main weaknesses are that the results do not support the conclusion, namely that the Smim32 gene is not specific to the claustrum and that no other orthogonal approaches were used to define the claustrum, such as retrograde neuroanatomical tracing from cortex. Also, these results are of limited applicability as the gene expression was only performed in mice, so it is unclear how Smim32 relates to claustrum in other mammalian species (e.g. primates), which have a very clearly defined claustrum. The article is also missing some key literature on the anatomical definition of claustrum, specifically as it relates to the endopiriform nucleus (which is putatively considered part of the claustrum in rodents).
Author response:
Data replicability
There are no replicates contained in the manuscript. (Reviewer #1)
We respectfully disagree with this statement. In this manuscript, we included both cell and animal replicates. For cell replicates, we analyzed over 50.000 cells using RNAscope and over 10.000 cells using RNAseq, employing two independent methods on different animals. We believe this extensive analysis is sufficient by any standards. Regarding animal replicates, we generated four different transgenic lines (two knockin lines and two BAC transgenic lines), which is an uncommon and rigorous effort. We analyzed dozens of animals, consistently observing the expression pattern of Smim32 and its derived transgenes across multiple experiments, including crosses between transgenics and various reporter lines, which is again an uncommon and rigorous effort. These experiments were conducted on animals from different litters to ensure robustness. Additionally, our longitudinal study, which includes 13 animals harvested at two-day intervals from E16 to P20, provides further consistency of our data.
However, to underscore the consistency of endogenous Smim32 expression, when submitting a revised manuscript, we will present Smim32 expression levels across individuals in single-cell RNA-seq data. Furthermore, we will pool data from different transgenic animals to demonstrate interindividual variability in the claustrum of adult animals.
Additional examples of female mice should also be included and separately quantified. (Reviewer #1)
We initially analyzed both males and females for one line (the Smim32-Cre knock-in line). Since we observed no differences between males and females (which we will note in the revised manuscript), we subsequently limited our analyses to males to minimize the use of animals.
Claustrum definition
Weaknesses lie in poor anatomical definitions of the claustrum (and endopiriform nucleus). (Reviewer #2)
No other orthogonal approaches were used to define the claustrum, such as retrograde neuroanatomical tracing from cortex. (Reviewer #3)
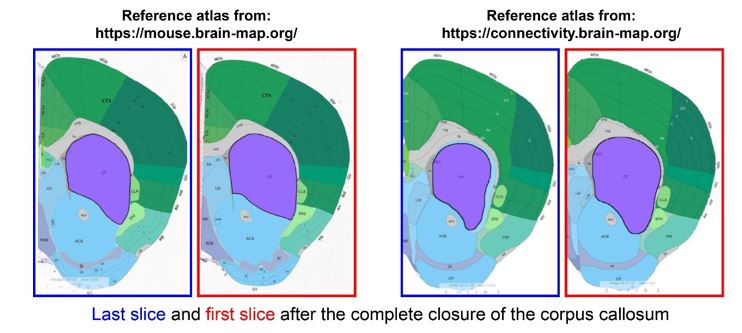
We share the reviewers’ opinion that the claustrum (CLA) and endopiriform nucleus (EN) are poorly defined anatomically in rodent brains due to the limited development of white matter tracts. This ambiguity has led to many conflicting descriptions of CLA/EN boundaries in various papers and atlases, including those by Paxinos and the Allen Brain Institute. Notably, the Allen Institute frequently updates the shape and anatomical location of the CLA/EN in their reference atlas, resulting in different websites displaying various versions (as illustrated in rebuttal figure 1 at comparable levels of the anteroposterior brain axis). It remains uncertain which version would most effectively satisfy the entire scientific community, if any. Indeed, after many years of working on these structures and surveying the literature, we regret to note that there is currently no consensus on the anatomical definition of the CLA and EN, even among expert laboratories using tracing or staining methods. At one end of the spectrum, some authors define the CLA as a small nucleus that could be, for example, characterized by the PVrich plexus. At the other end, other authors consider it part of a larger complex that includes the EN and extends dorsally to the S2 cortex. Additionally, differing definitions of the core and shell regions, as well as the precise anteroposterior extent of the nucleus, further complicate the issue.
Author response image 1.
Comparison of CLA and EN shapes in two recent versions of the Allen brain atlas
Given this lack of consensus, we deliberately opted for a molecular definition of the claustrum and its projection neurons. We used a set of well-documented canonical markers for the claustrum and neighboring neurons to determine the expression pattern of Smim32. The claustrum-specific markers we selected (Nr4a2, Lxn, Gnb4, Car3, etc.) have been extensively studied and allow us to distinguish claustrum projection neurons from neighboring and intermingled populations. Although none of these individual markers are exclusively specific to CLA and EN neurons, the combined expression of these markers provides greater confidence in identifying the different neuronal populations in space.
Smim32 expression is used to define claustrum anatomical boundaries, rather than first using several structural, molecular, and connectivity lines of evidence to define the claustrum anatomically and then to assess whether Smim32 expression fits within this anatomical definition. (Reviewer #2)
Contrary to the reviewer's suggestion, we do not define the claustrum based on Smim32 expression. Instead, Figures 1 and 2 demonstrate that Smim32 expression is highly correlated with the expression of known claustrum markers (Nr4a2, Lxn, Gnb4, Car3, etc.), both regionally and at the cellular level. As suggested by Peng et al. (2021, Fig. 4 and Extended Data Fig. 11), this population of cells, which includes the claustrum, a specific subset of cells in cortical layer 6, and the dorsal endopiriform nucleus, forms a discrete group of neurons sharing the same transcriptomic identity. Given what is known about the connectivity of claustrum and endopiriform nucleus projection neurons, this population obviously includes neurons projecting to various areas, likely fulfilling distinct functions. Whether these cells should be subdivided based on projection area, developmental origin, or structural features is beyond the scope of this article.
Specificity issues
Cre/Flp expression driven by the Smim32 promoter is present in non-claustrum regions, including the neighboring cortex, striatum, and endopiriform nucleus as well as the more distant thalamic reticular nucleus. (Reviewer #2)
The Smim32 gene is not specific to the claustrum. (Reviewer #3)
We do not claim that endogenous Smim32 expression is exclusive to the claustrum or that the knock-in lines, by themselves, are sufficient to isolate claustrum neurons without combined approaches based on the transgenic lines presented here. However, there are significant differences in the expression pattern between endogenous Smim32 and the expression of Cre in the various derived transgenic lines, which might not have been clear in the current manuscript. Notably, there is no expression of Cre in the striatum and the thalamic reticular nucleus, and only sparse expression in the endopiriform nucleus in Tg61(Smim32-cre). Each transgenic line provides different levels of overlap with the endogenous Smim32 expression, with the Tg61(Smim32-cre) line allowing for the most specific genetic access to claustrum neurons. Again, for greater specificity, any of these lines could be used in combined approaches, such as viral targeting (as shown in Figure 6A and B) or using transgenic intersectional (dual recombinase) approaches based on Cre- and Flp-expressing mice with an overlap in the claustrum, leading to circuit-specific and/or claustrum-only labeling.
This means that our claims are supported by the observed data. However, we acknowledge that we may not have clearly explained the specificity of the random transgenes, which could have led some reviewers to believe that « the data do not support the claims ».
We will clarify these points in the revised manuscript and include additional examples and quantifications to highlight the differences between endogenous Smim32 expression and Cre expression in the transgenic Tg61(Smim32-cre) line.
Regarding Cre-expressing cells in the neighboring cortex (layer 6 projection neurons), these cells are genetically distinct from other layer 6 cortical neurons and express the same canonical markers as claustrum projection neurons, likely sharing also the same transcriptomic identity. We will provide a more detailed characterization of these cells in the revised manuscript.
Since Smim32 driven recombinase (in 61 or 62lrod) is not exclusively expressed in the claustrum, it is not clear how Smim32 is an advantage over possible Nr4a2 or, the more selective, GNB4 Cre driver lines. (Reviewer #2)
Over the years, we have found a limited number of Cre lines used in the literature for targeting claustrum neurons. These include Gnb4-cre, Slc17a6-cre (also known as Vglut2-cre), Egr2-cre, Tg(Tbx21-cre), Ntng2-cre, Cux2-cre and Esr2-cre lines. We have not found any study describing and/or using an Nr4a2-cre line. Although a Nr4a2-Dre line exists (that we have studied in our laboratories), caution is warranted in its use, as it lacks the complete coding sequence of the Nr4a2 gene.
One problem with Nr4a2 is its documented expression in the adjacent Layer 6b cortical neurons, which discards it as a suitable candidate to selectively target the claustrum. Furthermore, Nr4a2 is also expressed in a majority of the endopiriform nucleus neurons, whereas endogenous Smim32 is expressed in a smaller proportion of these cells, and is restricted mainly to the dorsal endopiriform nucleus. These reasons led us to select Smim32 over Nr4a2.
Author response image 2.
(A) In situ hybridization for various CLA/EN marker genes. (B) Developmental recombination observed outside the CLA/EN in various cre lines (all data from the Allen brain databases)
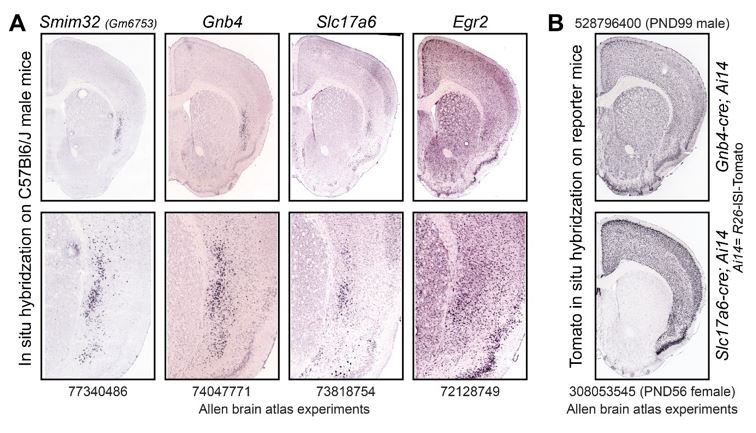
What are the advantages of using the different Smim32-cre lines over the existing Cre lines mentioned above?
Let’s first consider the Gnb4-cre line, which is considered one of the best available. Although the endogenous Gnb4 gene appears to have a similar expression pattern to Nr4a2, Slc17a6, and Smim32 in the striato-claustro-insular region of adult mice (Rebuttal Figure 2A), the results observed with the Gnb4-cre line either shows otherwise, or indicate that the Cre line does not fully recapitulate Gnb4 endogenous expression (Rebuttal Figure 3). Indeed some neurons in the insular cortex, piriform cortex, and putamen express the Cre recombinase (possibly due to low Gnb4 expression not detected in the in situ hybridization data of the Allen brain institute or due to nonspecific transgene expression) and will recombine viral vectors injected in adult mice (Rebuttal Figure 3). Therefore, this Cre expression outside the CLA/EN neurons in the Gnb4-cre line presents complications for data interpretation, depending on the viral injection coordinates and the quantity of injected vectors.
Author response image 3.
Specificity of the Gnb4-Cre line tested with viral transduction in adult mice (all data from the Allen Brain Institute database). The top and middle rows display the same data but with different scaling of the lookup tables to highlight either the patterns of axonal projections (top) or the infected neurons themselves (middle). The bottom row shows a higher magnification of the infection site. Note that individual neurons cannot be resolved in experiment 485903475 due to signal saturation.
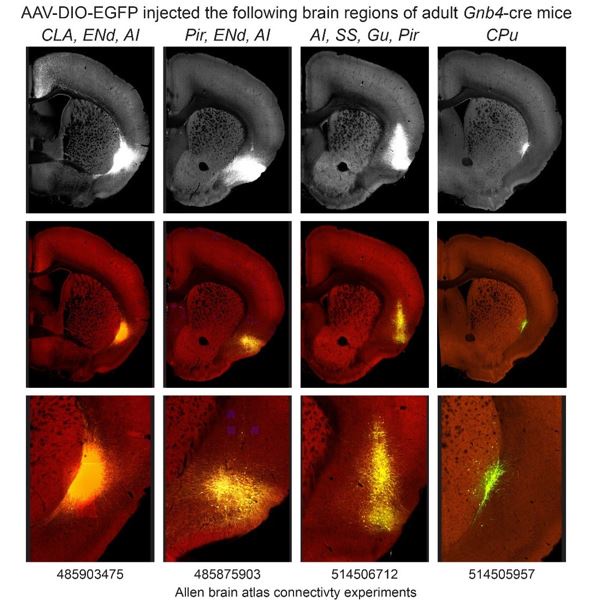
Cre expression in the CLA appears more specific in the various Smim32-cre transgenic lines than in many of the lines mentioned above. Although we have no doubt that the different existing transgenic lines can target CLA neurons, the selectivity of the targeting (for example, the fraction and types of CLA neurons versus potential non-CLA neurons) remains to be fully described for most of the lines. It is particularly true in the case of Tbx21 and Esr2 (used as drivers for the Tg(Tbx21-cre) and Esr2-cre transgenic lines). Tbx21 is not endogenously expressed in adult CLA neurons (evaluated by in situ and RNAseq data) and Egr2, if expressed in the claustrum, is not restricted to CLA neurons as it is an immediate early gene expressed in recently active neurons (Rebuttal Figure 2A).
Cre expression in the EN is observed in all Cre-expressing transgenic lines used to target the claustrum (with the exception of Slc17a6-cre). This can naturally be problematic for some approaches. Luckily, the random integrant Tg61(Smim32-cre) we describe in our manuscript shows a strong expression in the claustrum, and very limited expression outside the CLA (a very weak activity in the EN), representing a novel tool with improved claustrum selectivity. An advantage of the Tg61(Smim32-cre) over the Slc17a6-cre is that more CLA neurons can be targeted with the Tg61(Smim32-cre) line.
Another advantage of our four transgenic lines is their versatility; they can be used to recombine reporter lines as well as FRT-floxed and loxP-floxed knockouts in limited neuronal populations. They will be employed in the future for intersectional genetics to exclusively target CLA neurons. Existing transgenic lines cannot offer these possibilities because their marker genes are broadly expressed in the brain during embryogenesis, leading to the impact on a large number of non-CLA/EN neurons. This is evident in the Gnb4-cre and Slc17a6-cre lines crossed with the Ai14 reporter line expressing the fluorescent protein tomato (Rebuttal Figure 2B, right panels). Similar observations have been made for the Ntng2-Cre and Cux2-cre lines (see the Allen Brain Institute database for these data). Alternatively, inducible recombinase systems, such as the Gnb4-IRES2CreERT2-D line, could be used. However, the Gnb4-IRES2-CreERT2-D line requires tamoxifen to induce Cre recombination, which can be problematic depending on the research context, as well as recombinations in the absence of tamoxifen treatment (see experiments 560948627 and 560948194 in the Allen Brain database).
It is unclear how Smim32 relates to claustrum in other mammalian species (e.g. primates) (Reviewer #3)
As mentioned in the last paragraph of the introduction of the initial manuscript, Smim32 is specifically expressed in the claustrum of a primate species, Homo sapiens (reference 37 of the initially submitted manuscript).
Availability of the transgenic mice
These mice should be made available to the community through commercial vendors. (Reviewer #1 and #2 in private comments)
We are pleased to see that two of the three reviewers would like to see these mice available. These mice will not be kept for ourselves, and we will distribute them at some point in time, but this will naturally occur after the publication of the revised manuscript.
Critical comments on discussion and other topics
A clear description of the search in the Allen Mouse Brain Atlas is missing. A search for Smim32 in the ISH mouse atlas did not provide any hits and so it would be useful to include in the methods or results section the exact query used for examination of Smim32 expression as well as other genes identified in this process. (Reviewer #2)
Smim32 has been referred to by different names in various versions of the mouse genome. For the readers not versed in navigating genomes and annotations, before being officially named Smim32, this gene was originally called Gm6753 (as noted in the Allen Brain Institute database, see Rebuttal Figure 2A for an example of their in situ data) and later Gm45623.
Several sentences highlighting the shortfalls of other approaches are overstated and should be toned down. (Reviewer #1)
Very concerning is problematic language in the abstract and introduction sections that diminish the impact of several published studies (not cited) that have led to important findings regarding claustrum function. The authors create an argument that all the research performed thus far on the claustrum is unreliable because targeting the structure has been sub-optimal. (Reviewer #2)
A more balanced discussion of the strengths and weaknesses of these mice should be included. (Reviewer #1)
We regret if our choice of language inadvertently appeared to undermine the contributions of our colleagues; that was certainly not our intention. The paragraph in question was meant to address certain studies that we believe have led to inconsistent findings and unreliable data due to a lack of rigorous methodology in targeting claustrum projection neurons. To avoid singling out specific works, we chose not to cite them directly. We understand that some colleagues whose research does not fall under the “various cases” mentioned may feel unfairly targeted by this statement. We will revise this section to better clarify our intent and ensure it is respectful of all contributions. We will rephrase passages in the abstract, introduction, and discussion to provide a balanced view of the strengths and weaknesses of these mice.
Our main goal is to provide tools to specifically target claustrum cells based on their transcriptomic identity, which we believe is the best means to assess the function of any neuronal population. Due to the intermingling of claustrum neurons with neighboring populations, employing stereotaxic injections in the claustrum without genetic segregation will always infect and label physically adjacent cells that do not belong to the claustrum, ontologically and functionally speaking.
Similarly, targeting claustrum neurons retrogradely by injecting into claustrum projection sites likely labels neurons from different populations. For instance, as reviewer 1 mentions Erwin et al. (2021), infecting retrosplenial projections without genetic specificity labels many claustrum Synpr+ neurons (considered the claustrum core), a small proportion of claustrum Nnat+ neurons (considered the claustrum shell by some, and non-claustrum neurons by others), and some neighboring cortical L6b neurons. These three populations have very different transcriptomic identities, connectivity patterns, and likely distinct functions.
Thus, we believe that genetic specificity provides an important added value for selectively targeting the claustrum or claustro-insular complex.
A better characterization of all data should be undertaken. (Reviewer #1)
Having generated hundreds of transgenic lines over the years, we have never performed a more thorough analysis of transgenic lines, nor have a recollection of reading a publication evaluating at such a precise level the expression pattern of transgenes in mice. We, therefore, do not see exactly what the reviewer means by this remark. It is possible, not being native English speakers, that we did not grasp a certain form of joke.



