Peer review process
Revised: This Reviewed Preprint has been revised by the authors in response to the previous round of peer review; the eLife assessment and the public reviews have been updated where necessary by the editors and peer reviewers.
Read more about eLife’s peer review process.Editors
- Reviewing EditorDaniel HenrionUniversité d'Angers, Angers, France
- Senior EditorMatthias BartonUniversity of Zurich, Zurich, Switzerland
Reviewer #1 (Public review):
Summary:
Assessment of cardiac LEC transcriptomes post-MI may yield new targets to improve lymphatic function. scRNAseq is a valid approach as cardiac LECs are rare compared to blood vessel endothelial cells.
Strengths:
Extensive bioinformatics approaches employed by the group
Weaknesses:
Too few cells included in scRNAseq data set and the spatial transcriptomics data that was exploited has little relevance, or rather specificity, for cardiac lymphatics. This study seems more a collection of preliminary transcriptomic data than a true scientific report to help advance the field.
Comments on revisions:
Thank you for the revision that helps clarify some outstanding questions.
(1) I still have questions relating to the relevance of the spatial maps generated and shown in fig 3C. They are supposedly generated using a 'molecular finger print' specific to each sub-cluster of LECs. However, given that at early stages postMI most populations are exceedingly rare in your analyses, could you please explain or comment on the relevance of the spatial maps?
(2) Fig 3 s1 would indicate that the population CaII is the majoritarian one in healthy hearts, while quantifications in Fig 3A show that rather the LEC Co subpopulation is majoritarian. Further, in mouse hearts histological analyses have demonstrated that cardiac lymphatics are restricted to the outer layers of the heart. This is not seen in your spatial maps. This seems to be the case only for the LEc Co population in healthy hearts, but not for other subpopulation signatures. Please explain.
(3) Further, the population of CaI, with 1 cell analysed in d3, but appears very prevalent in the spatial maps at d3. Please explain.
(4) In your list of 12 genes used as matrix anchors to identify LEC subpopulations in your screens, it is not apparent how LEC CaI, II and III differ so much as to allow selective detection of subpopulations. This similitude of profiles is supported by Fig 2F, and further explanations are needed to explain how the spatial maps of LEC ca subpopulations appear as distinct as shown in fig 3 S1 and Fig 3C.
Reviewer #2 (Public review):
Summary:
This study integrated single-cell sequencing and spatial transcriptome data from mouse heart tissue at different time points post-MI. They identified four transcriptionally distinct subtypes of lymphatic endothelial cells and localized them in space. They observed that LECs subgroups are localized in different zones of infarcted heart with functions. Specifically, they demonstrated that LEC ca III may be involved in directly regulating myocardial injuries in the infarcted zone concerning metabolic stress, while LEC ca II may be related to the rapid immune inflammatory responses of the border zone in the early stage of MI. LEC ca I and LEC collection mainly participate in regulating myocardial tissue edema resolution in the middle and late stages post-MI. Finally, cell trajectory and Cell-Chat analyses further identified that LECs may regulate myocardial edema through Aqp1, and likely affect macrophage infiltration through the galectin9-CD44 pathway. The authors concluded that their study revealed the dynamic transcriptional heterogeneity distribution of LECs in different regions of the infarcted heart and that LECs formed different functional subgroups that may exert different bioeffects in myocardial tissue post-MI.
Strengths:
The study addresses a significant clinical challenge, and the results are of great translational value. All experiments were carefully performed, and their data support the conclusion.
Editors' comments (Public review):
Weaknesses:
(1) Figure 7C, 7E, 7I, and "Figure7-figure supplement 1 ": All data in these data panels are based on only n=3, which is insufficient. Sample sizes of n=3 are too low to correctly assess normality of distribution and, as a consequence, do not allow to select the appropriate parametric/non-parametric tests. Accordingly, no statistical comparison can be performed and all p values and symbols currently indicating statistically significant differences between groups must be removed.
(2) Figure 3A, 3B, or 3C: No information about n numbers per group. Should n numbers per group be n=4 or less, no statistical comparison can be performed and all p values and symbols indicating statistically significant differences between groups must be removed.
(3) Figure 4 E and 4F: No information about n numbers per group. Should n numbers per group be n=4 or less, no statistical comparison can be performed and all p values and symbols indicating statistically significant differences between groups must be removed.
(4) Figure 5: No information about n numbers per group is provided. Should n numbers per group be n=4 or less, no statistical comparison can be performed and all p values and symbols indicating statistically significant differences between groups must be removed.
Author response:
The following is the authors’ response to the previous reviews.
Reviewer #1 (Recommendations for The Authors):
Q1: In response to reviewers you noted totally 292 sequenced LECs, however in reviewer figure 3 B the numbers seem to add up to 221. Please include mention of the total number of LEC sequences. Please mention line 119, page 4 the total number of explored LEC transcriptomes
Thank you for your carefully review. We have updated Fig 2A, 2C and 2E. It was 242 (not 292) LECs included in our initial analysis, which contains the sample of d5 post MI in raw data (E-MTAB-7895). We dropped d5 in our subsequent analysis because the change in d5 did not significant differ from d3. Therefore, we included 221 LECs in our final analysis as we updated in Fig2A, 2C and 2E.
Q2-1: Figure 3A supposedly shows % of LEC subpopulations relative to their numbers found in day 0 samples. However, there seem to be some errors, because for example the subpop LEC Cap I include 13 cells day 1 and 6 cells day 1, which corresponds to 46% of initial numbers. However, from your graph 3B the blue population seems to occupy 10%. Please revise or explain how these relative % were calculated.
Thank you for your question. In the Figure 3A, each column was calculated by dn/d0*100%, that is d0=57/57*100%=100%, and d1= 21/57*100%=36.84%, d3=9/57*100%=15.79%, d7, d14, d28...Therefor, Cap I in d0 (13 cells) is 13/57*100%=22.81%, and Cap I in d1(6 cells) is 6/57*100%= 10.53%.
Q2-2: Further, based on the relative % of LEC subpopulations, using the numbers mentioned in Fig 3B, it would appear that the relative frequency LEC cap II population is actually stable at around 20-30% of all LECs per time point throughout the study (except day 1 drop). This contrasts with line 136 p. 4 statement. I would also urge caution for interpreting too much into the variation of relative levels of LEC co, as these represent exceeding rare cells in your samples, and could reflect technical issues rather than true biological variation (total LEC co numbers analyzed ranging from 1-24 cells/ time point). The same could be said of LEC cap II and cap III.
We strongly agree with your comment on the proportion of LEC cell subtypes post MI. As you pointed out, we have revised the result description on Page 4, line 137-143 as followed.
“In the early stages of myocardial infarction (D1 and D3), the quantity of LECs decreased sharply. The number of LECs gradually increasing from day 7 and returning to normal levels by day 14 after MI. Moreover, from day 14 onwards, the number and proportion of Ca I type LECs significantly increased.”
Q3: Please list in supplement the gene features used to identify in spatial transcriptomics the different LEC subpopulations, as their profiles (notably for capillary LECs) don't appear to be very different based on data in Fig 2F.
We have supplied gene features in supplementary materials.
Q4: In section 2.7 you refer to Gal9 secretion. Please replace with expression as no measure of protein levels from LECs has been described in your study.
Thank you for your suggestion, we have replaced secretion with expression.
Q5: The updated method to exclude non-lymphatic cells from lymphatic vessel analyses by incorporating pdpn as an additional marker ('present costained areas wherever possible' line 350 p 10)
Thank you for your correction. We have updated the description as follows and lighted them in the manuscript: rabbit anti-Lyve1 (1:300, ab14917, Abcam, UK), [Syrian hamster anti-Podoplanin (1:100, 53-5381-82, Thermo, USA), rabbit anti-Prox1(1:300, ab199359, Abcam, UK), both anti-podoplain and anti-prox1 are additional markers co-stained with Lyve1 to exclude non-lymphatic cells from lymphatic vessel].
Q6: Fig 1B, it is highly surprising to see the lymphatic density in the BZ go from 25 um² at day 3 to more than 1000 um² only four days later (day 7). Is it possible that your day 3 measurements were in the infarct area, and not BZ area? The H&E image shown in Fig1a for d3 sample would seem to indicate the analysis was done in a dead area, rather than BZ. Please revise (perhaps select similar zone as shown for d1 in fig 6D, adjusted for subepicardial region and not mid-myocardial as seems to be the case currently), and also provide lymphatic area measures in healthy myocardium for day 0 samples. The unit used (um²) also would depend on the size of the area examined. Is this unit per image? If so please report total imaged area as a reference.
A6: Thank you for your reminding and advises. We have labeled each zone on H&E and IF images in Fig1-supplementary Fig2B, and updated a clearer histological photo taken at 3 days post MI in Fig1A. Furthermore, we recalculated the lymphatic vessel area ratio as you suggested by calculating the ratio of LEC co-stained area to total imaged area under 100-fold magnification.
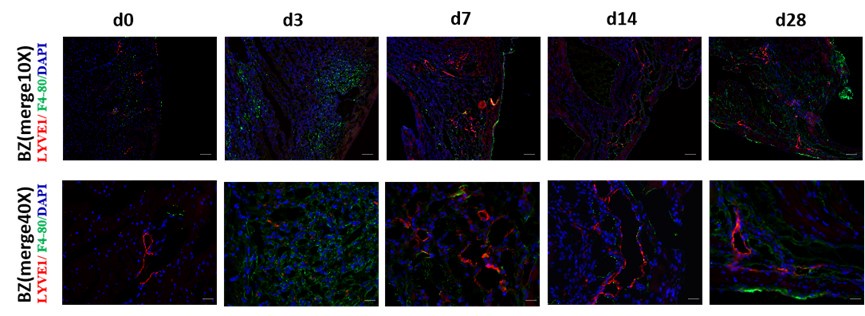
Q7: The mention that CD68 antibody isn't compatible with lyve1 antibody could easily have been bridged by using other macrophage markers, such as F4/80, which is readily available and often used marker for macs in mice and comes notably as a rat anti-mouse F4-80. It would have added much more relevant information to exclude Lyve1-/F4/80+ cells as compared to the current analysis, which may indeed include in area measures Lyve1+ /Pdpn- single cells erroneously spotted as 'lymphatic vessels'
Thank you for your excellent suggestion. We co-stained the sample with F4/80 and LYVE1 and supplied in the Fig1-supplementary Figure 1E, as shown in Author response image 1.
Author response image 1.
Immunofluorescence (IF) co-staining of tissue section with F4/80 and LYVE1 in sham and MI mice model at d3, d7, d14, and d28 post-MI. LYVE1: lymphatic vessel endothelial hyaluronan receptor 1; DAPI: 4’6-diamidino-2-phenylindole; scale bar in 10×-100 μm, 40×-25μm.
Reviewer 2 (Recommendations for The Authors):
Q1: Language expression must be improved. Many incomplete sentences exist throughout the manuscript. A few examples: Line 70-71: In order to further elucidate the effects and regulatory mechanisms of the lymphatic vessels in the repair process of myocardial injury following MI. Line 71-73. This study, integrated single-cell sequencing and spatial transcriptome data from mouse heart tissue at different timepoints after MI from publicly available data (E-MTAB-7895, GSE214611) in the ArrayExpress and gene expression omnibus (GEO) databases. Line 88-89: Since the membrane protein LYVE1 can present lymphatic vessel morphology more clearly than PROX1.
Thank you for your correction. We have carefully inspected and corrected the whole manuscript.
Q2: The type of animal models (i.e., permanent MI or MI plus reperfusion) included in Array Express and gene expression omnibus (GEO) databases must be clearly defined as these two models may have completely different effects on lymphatic vessel development during post-MI remodeling.
Thank you for your excellent suggestion. The animal models used in both E-MTAB-7895 and GSE214611 are permanent MI. We have modified the model information in the methodology section (page 12, line 400-401).
Q3: Line 119-120: Caution must be taken regarding Cav1 as a lymphocyte marker because Cav1 is expressed in all endothelial cells, not limited to LEC.
Thanks for your reminding. Cav 1 used in our clustering is one of the marker gene for its different expression in sub-types of LECs, referred in article PMID: 31402260
Q4: Figure 1 legend needs to be improved. RZ, BZ, and IZ need to be labeled in all IF images. Day 0 images suggest that RZ is the tissue section from the right ventricle.
Thank you for your suggestion. We have labeled and updated the regions of RZ, BZ, and IZ in H&E and IF image in Figure1-Figure supplement 2B.
Q5: The discussion section needs to be improved and better focused on the findings from the current study.
Thank you for your good comment. Based on your suggestion, we have revised the first paragraph of the discussion from lines 250-256 (Page 7) as followed:
Cardiac lymphatics play an important role in myocardial edema and inflammation. This study, for the first time, integrated single-cell sequencing data and spatial transcriptome data from mouse heart tissue at different time points of post-MI, and identified four transcriptionally distinct subtypes of LECs and their dynamic transcriptional heterogeneity distribution in different regions of myocardial tissue post-MI. These subgroups of LECs were shown to form different function involved in the inflammation, apoptosis, ferroptosis, and water absorption related regulation of vasopressin during the process of myocardial repair after MI.



