Peer review process
Revised: This Reviewed Preprint has been revised by the authors in response to the previous round of peer review; the eLife assessment and the public reviews have been updated where necessary by the editors and peer reviewers.
Read more about eLife’s peer review process.Editors
- Reviewing EditorBavesh KanaUniversity of the Witwatersrand, Johannesburg, South Africa
- Senior EditorBavesh KanaUniversity of the Witwatersrand, Johannesburg, South Africa
Reviewer #1 (Public review):
Summary:
Lodhiya et al. demonstrate that antibiotics with distinct mechanisms of action, norfloxacin and streptomycin, cause similar metabolic dysfunction in the model organism Mycobacterium smegmatis. This includes enhanced flux through the TCA cycle and respiration as well as a build-up of reactive oxygen species (ROS) and ATP. Genetic and/or pharmacologic depression of ROS or ATP levels protect M. smegmatis from norfloxacin and streptomycin killing. Because ATP depression is protective, but in some cases does not depress ROS, the authors surmise that excessive ATP is the primary mechanism by which norfloxacin and streptomycin kill M. smegmatis. In general, the experiments are carefully executed; alternative hypotheses are discussed and considered; the data are contextualized within the existing literature.
Strengths:
The authors tackle a problem that is both biologically interesting and medically impactful, namely, the mechanism of antibiotic-induced cell death.
Experiments are carefully executed, for example, numerous dose- and time-dependency studies; multiple, orthogonal readouts for ROS; and several methods for pharmacological and genetic depletion of ATP.
There has been a lot of excitement and controversy in the field, and the authors do a nice job of situating their work in this larger context.
Inherent limitations to some of their approaches are acknowledged and discussed e.g., normalizing ATP levels to viable counts of bacteria.
Weaknesses:
All of the experiments performed here were in the model organism M. smegmatis. As the authors point out, the extent to which these findings apply to other organisms (most notably, slow-growing pathogens like M. tuberculosis) is to be determined. To avoid the perception of overreach, I would recommend substituting "M. smegmatis" for Mycobacteria (especially in the title and abstract).
At first glance, a few of the results in the manuscript seem to conflict with what has been previously reported in the (referenced) literature. In their response to reviewers, the authors addressed my concerns. It would also be ideal to include a few lines in the manuscript briefly addressing these points. (Other readers may have similar concerns)
In the first round of review, I suggested that the authors consider removing Figs. 9 and 10A-B as I believe they distract from the main point of the paper and appear to be the beginning of a new story rather than the end of the current one. I still hold this opinion. However, one of the strengths of the eLife model is that we can agree to disagree.
Reviewer #2 (Public review):
Summary:
The authors are trying to test the hypothesis that ATP bursts are the predominant driver of antibiotic lethality of Mycobacteria
Strengths:
No significant strengths in the current state as it is written.
Weaknesses:
A major weakness is that M. smegmatis has a doubling time of three hours and the authors are trying to conclude that their data would reflect the physiology of M. tuberculossi that has a doubling time of 24 hours. Moreover, the authors try to compare OD measurements with CFU counts and thus observe great variabilities.
Comments on revisions:
I am surprised that the authors simply did not repeat the study in figure one with CFU counts and repeated in triplicate. Since this is M. smegmatis, it would take no longer than two weeks to repeat this experiment and replace the figure. I understand that obtaining CFU counts is much more laborious than OD measurements but it is necessary. Your graph still says that there is 0 bacteria at time 0, yet in your legend it says you started with 600,000 CFU/ml. I don't understand why this experiment was not repeated with CFU counts measured throughout. This is not a big ask since this is M. smegmatis but it appears that the authors do not want to repeat this experiment. Minimally, fix the graph to represent the CFU.
Author response:
The following is the authors’ response to the original reviews.
Reviewer #1 (Public review):
Summary:
Lodhiya et al. demonstrate that antibiotics with distinct mechanisms of action, norfloxacin, and streptomycin, cause similar metabolic dysfunction in the model organism Mycobacterium smegmatis. This includes enhanced flux through the TCA cycle and respiration as well as a build-up of reactive oxygen species (ROS) and ATP. Genetic and/or pharmacologic depression of ROS or ATP levels protect M. smegmatis from norfloxacin and streptomycin killing. Because ATP depression is protective, but in some cases does not depress ROS, the authors surmise that excessive ATP is the primary mechanism by which norfloxacin and streptomycin kill M. smegmatis. In general, the experiments are carefully executed; alternative hypotheses are discussed and considered; the data are contextualized within the existing literature. Clarification of the effect of 1) ROS depression on ATP levels and 2) ADP vs. ATP on divalent metal chelation would strengthen the paper, as would discussion of points of difference with the existing literature. The authors might also consider removing Figures 9 and 10A-B as they distract from the main point of the paper and appear to be the beginning of a new story rather than the end of the current one. Finally, statistics need some attention.
Strengths:
The authors tackle a problem that is both biologically interesting and medically impactful, namely, the mechanism of antibiotic-induced cell death.
Experiments are carefully executed, for example, numerous dose- and time-dependency studies; multiple, orthogonal readouts for ROS; and several methods for pharmacological and genetic depletion of ATP.
There has been a lot of excitement and controversy in the field, and the authors do a nice job of situating their work in this larger context.
Inherent limitations to some of their approaches are acknowledged and discussed e.g., normalizing ATP levels to viable counts of bacteria.
We sincerely appreciate the reviewer’s encouraging feedback.
Weaknesses:
The authors have shown that treatments that depress ATP do not necessarily repress ROS, and therefore conclude that ATP is the primary cause of norfloxacin and streptomycin lethality for M. smegmatis. Indeed, this is the most impactful claim of the paper. However, GSH and dipyridyl beautifully rescue viability. Do these and other ROS-repressing treatments impact ATP levels? If not, the authors should consider a more nuanced model and revise the title, abstract, and text accordingly.
We thank the reviewer for asking this question. In the revised version of the manuscript, we have included data on the impact of the antioxidant GSH on antibiotic-induced ATP levels as the supplementary figure (S9C)
Does ADP chelate divalent metal ions to the same extent as ATP? If so, it is difficult to understand how conversion of ADP to ATP by ATP synthase would alter metal sequestration without concomitant burst in ADP levels.
We sincerely thank the reviewer for raising this insightful question. Indeed, ADP and AMP can also form complexes with divalent metal ions; however, these complexes tend to be less stable. According to the existing literature, ATP-metal ion complexes exhibit a higher formation constant compared to ADP or AMP complexes. This has been attributed to the polyphosphate chain of ATP, which acts as an active site, forming a highly stable tridentate structure (Khan et al., 1962; Distefano et al., 1953). An antibiotic-induced increase in ATP levels, irrespective of any changes in ADP levels or a total pool size of purine nucleotides, could still result in the formation of more stable complexes with metal ions, potentially leading to metal ion depletion. Although recent studies indicate that antibiotic treatment stimulates purine biosynthesis (Lobritz MA et al., 2022; Yang JH et al., 2019), thereby imposing energy demands and enhancing ATP production, and therefore, the possibility of a corresponding increase in total purine nucleotide levels (ADP+ATP) exist (is mentioned in discussion section). However, this hypothesis requires further investigation.
Khan MMT, Martell AE. Metal Chelates of Adenosine Triphosphate. Journal of Physical Chemistry (US). 1962 Jan 1;Vol: 66(1):10–5
Distefano v, Neuman wf. Calcium complexes of adenosinetriphosphate and adenosinediphosphate and their significance in calcification in vitro. Journal of Biological Chemistry. 1953 Feb 1;200(2):759–63
Lobritz MA, Andrews IW, Braff D, Porter CBM, Gutierrez A, Furuta Y, et al. Increased energy demand from anabolic-catabolic processes drives β-lactam antibiotic lethality. Cell Chem Biol [Internet]. 2022 Feb 17.
Yang JH, Wright SN, Hamblin M, McCloskey D, Alcantar MA, Schrübbers L, et al. A White-Box Machine Learning Approach for Revealing Antibiotic Mechanisms of Action. Cell [Internet]. 2019 May 30
Reviewer #1 (Recommendations for the authors):
(1) Some of the results in the paper diverge from what has been previously reported by some of the referenced literature. These discrepancies should be clarified.
We apologize for any confusion, but we are uncertain about the specific discrepancies the reviewer is referring. In the discussion section, we have addressed and analysed our results within the broader context of the existing literature, regardless of whether our findings align with or differ from previous studies.
(a) CCCP, nigericin, BDQ, and the atpD mutant all appear to affect M. smegmatis growth (Figures S6C, S7C, S7D-E, and Figure 1B from reference 41). Could depressed growth contribute to the rescue effects of these compounds?
We concur with the reviewer that the reagents we used (CCCP, Nigericin, and BDQ) to suppress the ATP burst in the presence of antibiotics do affect bacterial growth. This growth sub-inhibitory effect is expected given their roles in either uncoupling the electron transport chain from oxidative phosphorylation or directly inhibiting ATP synthase, leading to reduced ATP production compared to the untreated control. However, we chose concentrations that reduces the antibiotic-induced surge in ATP levels without significantly depriving the bacteria of the ATP essential for their survival, thereby avoiding cell death.
Consequently, all three reagents (as shown in Figures S6C, S7C, and S7D-E) were employed at non-lethal concentrations. We would like to emphasize, however, that it was not feasible to select a reagent concentration that had no impact on growth yet still suppressed the antibiotic-induced ATP burst. We recognize the possibility that growth retardation may have contributed to the observed rescue effects. To address this concern, we used multiple orthogonal methods (CCCP, Nigericin, and BDQ), each with distinct mechanisms having a common effect of reducing the ATP surge, to minimize off-target effects and support our findings.
Also, the authors report no growth phenotype for atpD mutant (Figure S8) but only carry out the growth curve to an OD of 2, which is approximately where the growth curve from ref 41 begins to diverge.
Additionally, to further confirm that bacterial rescue was not due to growth retardation caused by these reagents, we utilized the atpD mutant. All experiments, including those involving the atpD mutant, were conducted when the OD600nm reached 0.8 (during the exponential phase). We specifically ensured that the growth of the atpD mutant was not compromised during this phase (Figure S8) and restricted our growth curve to the early stationary phase (OD600 between 1.5 and 2). While it is possible that the atpD mutant may exhibit slower growth compared to wild-type bacteria in stationary phase at an OD600nm of 4 (as shown in ref 41), however, this does not impact our observations.
(b) Reference 41 also reports that the atpD mutant is more sensitive to some antibiotics (Figure 6). This includes isoniazid, which references 34 and 35 have both reported caused an ATP burst.
We acknowledge the reviewer’s query regarding the phenotype of the atpD mutant against isoniazid (Reference 41). However, the cited reference does not provide clarity on why the M. smegmatis atpD mutant exhibits increased sensitivity to isoniazid and other antibiotics, nor does it explain whether this sensitivity is due to reduced ATP levels or altered cell wall properties, such as enhanced drug uptake, as observed with Nile red and ethidium bromide.
While references 34 and 35 reported an ATP burst following isoniazid treatment in slow-growing M. bovis BCG and M. tuberculosis, it remains to be tested whether isoniazid acts similarly in the fast-growing M. smegmatis, where it is bacteriostatic rather than being bactericidal as observed in M. bovis BCG and M. tuberculosis.
(2) The statistics require some attention. First, the wording for almost all of the figures is something like "data points represent the mean of at least three independent replicates," is that correct? CFUs are notoriously messy so it is surprising (impressive?) that the variability between replicates is so low. Second, t-tests are not appropriate for multiple comparisons.
We thank the reviewer for raising this important query. It is correct that all our experiments included at least three independent replicates, and many of our results exhibit a high degree of variability, as indicated by the large error bars. We would like to clarify that we did not perform multiple comparisons on our results. For all analyses, an unpaired t-test was conducted between the control group and one experimental group at a time. Consequently, statistical data were generated for each pair of results, and the comparisons were displayed on the graph relative to the control data points, as mentioned in the Methods section under the heading “Statistical analysis”
(3) Figures 9 and 10A-B seem tangential to the main point of the paper and, in the case of Figure 10A-B, preliminary.
In this study, our aim was to comprehensively investigate the nature of antibiotic-induced stresses (i.e., mechanisms of action from T = 15 hrs) and leverage these insights to enhance our understanding of bacterial adaptation mechanisms, particularly antibiotic tolerance (from T = 25 hrs). While a significant portion of the manuscript focuses on the secondary consequences of antibiotic exposure, we also sought to assess the bacteria's ability to counteract these stresses, contributing to our understanding of how antibiotic tolerance phenotypes develop.
The results presented in Figure 9 clearly demonstrate that bacteria attempt to reduce respiration by decreasing flux through the complete TCA cycle, thereby mitigating ROS and ATP production in response to antibiotics. These findings not only uncovers potential metabolic pathways to downregulate respiration but also validate our observations regarding the role of increased respiration, ROS generation, and subsequent ATP production in antibiotic action.
Importantly, bacterial responses to antibiotics were not limited to metabolic adaptations. They also included the upregulation of the intrinsic drug resistance determinant Eis (Figure 10A) and an increase in mutation frequency (Figure 10B), both of which indicate a greater likelihood of these bacteria developing antibiotic tolerance and resistance. Therefore, the data presented in Figures 9 and 10A-B are not peripheral to the central theme of the paper. Rather, they complement and strengthen it by providing a comprehensive understanding of the consequences of antibiotic exposure, which aligns with the primary objectives of our study.
Do the various perturbations used here (especially streptomycin) effect expression and/or turnover of the genetically-encoded sensors Mrx1-roGFP2 or Peredox-mCherry?
We appreciate the reviewer for raising this query. Since streptomycin treatment leads to mistranslation and eventually inhibits protein synthesis, it is possible that such treatment could impact the expression and/or turnover of the genetically encoded biosensors, Mrx1-roGFP2 (1) or Peredox-mCherry (2). However, we do not anticipate any effects on the readout as both biosensors provide ratiometric measurements of redox potential and NADH levels, respectively, which eliminates errors due to variations in protein abundance. Nevertheless, in our experiments with both drugs, we employed multiple time- and dose-dependent responses, ensuring that all meaningful conclusions were drawn from the overall trends seen in the data rather than an individual data point.
(1) Bhaskar A, Chawla M, Mehta M, Parikh P, Chandra P, Bhave D, et al. (2014) Reengineering Redox Sensitive GFP to Measure Mycothiol Redox Potential of Mycobacterium tuberculosis during Infection. PLoS Pathog 10(1): e1003902. https://doi.org/10.1371/journal.ppat.1003902
(2) Shabir A. Bhat, Iram K. Iqbal, and Ashwani Kumar*. Imaging the NADH:NAD+ Homeostasis for Understanding the Metabolic Response of Mycobacterium to Physiologically Relevant Stresses. Front Cell Infect Microbiol. 2016; 6: 145. doi: 10.3389/fcimb.2016.00145
(4) Do the antibiotics affect permeability? Especially relevant to CellROX experiments.
Antibiotics can impact, or even increase, bacterial membrane permeability, a phenomenon noticed in case of self-promoted uptake of aminoglycosides. When aminoglycosides bind to ribosomes, they induce mistranslation, including of membrane proteins, leading to the formation of membrane pores, which in turn enhances antibiotic uptake and lethality (1-2). However, whether the antibiotics used in our study (norfloxacin and streptomycin) at the concentrations applied altered membrane permeability is not known.
Experiments involving the CellROX dye are unlikely to be influenced by changes in membrane permeability, as the dye is freely permeable to the mycomembrane.
References:
(1) Davis BD Chen LL Tai PC (1986) Misread protein creates membrane channels: an essential step in the bactericidal action of aminoglycosides PNAS 83:6164–6168.
(2) Ezraty B Vergnes A Banzhaf M Duverger Y Huguenot A Brochado AR Su SY Espinosa L Loiseau L Py B Typas A Barras F (2013) Fe-S cluster biosynthesis controls uptake of aminoglycosides in a ROS-less death pathway Science 340:1583–1587.
(5) Figures 4E-H does GSH affect bacterial growth/viability on its own i.e. in the absence of a drug?
We thank the reviewer for raising this query. Indeed, the 10 mM GSH used in our experiments to mitigate and rescue cells from antibiotic-induced ROS does impact bacterial growth on its own, though it does not affect viability, likely due to GSH inducing reductive stress on bacterial physiology. For clarification, we have included the viability measurement data in the presence of 10 mM GSH alone in the revised version of the manuscript, as supplementary figure (S4E).
(6) p. 2 "...antibiotic resistance involves more complex mechanisms and manifests as genotypic resistance, antibiotic tolerance, and persistence." This reads as tolerance and persistence being a subset of resistance, which is not quite accurate. There is at least one other example of similar wording in the text.
We thank the reviewer for highlighting this point. Our intention was to convey that resistance to antibiotics can manifest in two forms: permanent or genetic resistance, and transient resilience through antibiotic tolerance and persistence.
(7) p. 3 "...and showing no visible differences in the growth rate...". It is hard to say this as all the values appear to be 0 - possible to zoom in on the CFU counts in this region? Same comment for p. 5 "...the unaffected growth rate in the early response phase...".
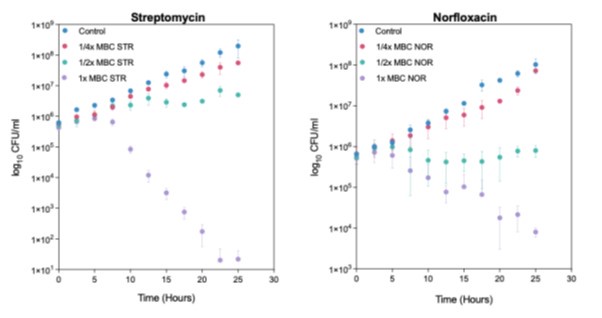
We apologize for the lack of clarity regarding the resolution of the early time points in the growth curve. Unfortunately, it was not feasible for us to zoom in on the initial time points due to the significant difference in cell viability between T=0 and T=25 hours (i.e., spanning 8 generations). For clarification in the growth phenotype at early time points, please refer to Author response image 1, where CFU counts are plotted on a logarithmic scale. The y-axis spans 6-8 orders of magnitude across different conditions, making it difficult to visualize early time points on a linear scale.
Author response image 1.
(8) p. 5 "...data for each condition were subjected to rigorous quality control analysis (S2B)." I believe that this is the case, but how Figure S2B demonstrates this fact is not clear.
Figures S2A and S2B present the quality assessment data for all six proteomics datasets. Figure S2A illustrates the consistency in the number of proteins identified across 10 samples (5 independent replicates for both control and drug treatment). The minimal variation in the number of identified proteins indicates reproducibility across the different runs. Similarly, Figure S2B displays the variability in Pearson correlation coefficient values of protein abundance (LFQ intensities) across the 10 samples. The closer and more consistent the Pearson correlation values, the greater the reproducibility of the quantitative data acquisition.
(9) p. 7 "To look for a shared mechanism of antibiotic action..." The wording implies an assumption. Perhaps "to test whether" would be more appropriate? Same comment for p. 12 "To further confirm whether enhanced respiration ...".
We appreciate the reviewer’s suggestions for both sentences and have made the necessary changes in the revised version. Thank you for bringing this to our attention.
(10) Figure S1A-B figure legend. How was this assay performed?
The experiment for Figures S1A-B was conducted using a standard REMA assay, as described in the methods section. Cells were harvested at the 25th-hour time point, and drug MICs were compared between cells grown with and without 1/4x MBC99 of the drugs. This was done to determine whether the growth recovery observed during the recovery phase was due to the presence of drug-resistant bacteria.
(11) p. 14 "...(CCCP), a protonophore, at non-toxic levels..." Figure S6C implies an effect on growth.
As clarified earlier in response to query 1(a), the CCCP reagent was used at concentrations that effectively minimize the antibiotic-induced surge in ATP levels. However, at these concentrations, CCCP reduces cellular ATP production (Figure S6A), leading to bacterial growth delay (Figure S6C). By "non-toxic levels," we intended to convey that these concentrations of CCCP are non-lethal to the bacteria, as evidenced in Figure S6C.
(12) Figure 8A y axis is this CFU/mL or OD/mL?
The y-axis for the figure 8A depicts CFU/ml as it measures the cell survival in response to increasing concentrations of bipyridyl.
Reviewer #2 (Public review):
Summary:
The authors are trying to test the hypothesis that ATP bursts are the predominant driver of antibiotic lethality of Mycobacteria.
Strengths:
This reviewer has not identified any significant strengths of the paper in its current form.
Weaknesses:
A major weakness is that M. smegmatis has a doubling time of three hours and the authors are trying to conclude that their data would reflect the physiology of M. tuberculosis which has a doubling time of 24 hours. Moreover, the authors try to compare OD measurements with CFU counts and thus observe great variabilities.
If the authors had evidence to support the conclusion that ATP burst is the predominant driver of antibiotic lethality in mycobacteria then this paper would be highly significant. However, with the way the paper is written, it is impossible to make this conclusion.
We have identified a new mechanism of antibiotic action in Mycobacterium smegmatis. However, as discussed extensively in the manuscript's discussion section, whether and to what extent this mechanism applies to other organisms still needs to be tested.
We have always drawn inferences from the CFU counts as the OD600nm is never a reliable method as reported in all of our experiments.
Reviewer #2 (Recommendations for the authors):
Figure 1 needs to have an x-axis that has intervals that have 10E5 CFU to 4 x 10E8. But even 4 x 10E8 CFU/ml is a late log and not exponentially growing cells.
Figure 1 illustrates the growth curve. We hope the reviewer meant the Y axis which represents CFU/ml on a linear scale. As mentioned in response to reviewer #1’s query no. 7, it was not feasible to include the viability (CFU/ml) values at T=0 and a few subsequent time points. Naturally, the starting cell count was not zero; we began with approximately 600,000 CFU/ml, corresponding to an OD600nm of 0.0025/ml. For clarification, we have mentioned the initial OD as well CFU/ml at T= 0 hr in the figure legend.
Carefully look at Figure 1, what were you trying to show? Your x-axis goes from 0 to 10E8, of course you did not inoculate 0 cells, but if you had measured CFUs, you might not have gotten the great variability you reported in your graph.
We assume that the reviewer is suggesting that "if we had measured OD600nm/ml instead of CFU/ml, we might not have observed the high variability we reported." While we agree with the reviewer's comment, our decision to use CFU/ml for growth measurement was to obtain more resolved and detectable data points, as an OD600nm of 0.0025/ml cannot be reliably measured with a spectrophotometer. Additionally, at around T=15 hours, where we observed an extended lag phase (referred to as the stress phase), the OD600nm was approximately 0.05, which is barely detectable. Therefore, the significant differences between the control group and the ¼ x MBC99 drug-treated group might not have been observed if we had relied on OD-based measurements. Despite the presence of high error bars and variability in the data points, we were still able to demonstrate clear differences in bacterial growth between treated and untreated samples at sub-lethal drug doses. This ultimately allowed us to capture the nature of antibiotic-induced stresses.
There is no doubt that sublethal concentrations of antibiotics will have an effect on the bacterial cells. But it is not clear how you are concluding that ATP burst is the dominant driver of lethality. M. smegmatis can be very different from Mtb.
Using a series of time- and dose-dependent experiments with plasmid and kit-based approaches, we demonstrated that both antibiotics generate and rely on ROS and ATP bursts to induce lethality in M. smegmatis. Careful monitoring of oxidative stress in cells, following specific quenching of the antibiotic-induced ATP burst (Figure 7, S9A-B), revealed that the ATP burst is the dominant driver of antibiotic lethality. In all tested experiments, surviving bacteria exhibited elevated levels of oxidative stress but were able to maintain their viability, suggesting that oxidative stress alone is not the dominant factor in antibiotic-induced lethality. Furthermore, quenching of ROS by glutathione also suppressed antibiotic-induced surge in ATP levels, thus supporting the notion that ROS alone, is not the dominant driver of antibiotic action as previously understood.
All experiments reported were conducted using fast-growing M. smegmatis, and have acknowledged the need for similar experiments in other bacterial systems, including M. tuberculosis, to assess whether our findings are applicable to other systems.
Another point, the use of a mutant in the ATP synthase is an interesting idea, but would it be better to use something where you knock out the ATP synthase activity with siRNA or a temperature-sensitive allele?
We appreciate the reviewer’s encouraging comment. Knocking out ATP synthase would completely halt oxidative phosphorylation and shut down aerobic respiration, leading to severe metabolic and growth defects. Such stressful and non-growing conditions are not suitable for testing the efficacy of antibiotics, as it is widely accepted that antibiotics are more effective against metabolically active bacteria.
Lastly, the conclusion is that norfloxacin and streptomycin have common mechanisms of action, but the authors do not explain how a DNA gyrase inhibitor shows the same mechanisms of action as a ribosome inhibitor.
The connection between antibiotic target corruption (DNA gyrase or ribosome) and the activation of respiration is indeed unclear, intriguing, and represents one of the most exciting questions in the field of antibiotic mechanisms of action. In the discussion section, we have speculated on potential pathways for this connection, including the possibility that the inhibition of cell division by both drugs may create a perception of resource scarcity (energy and biosynthetic precursors), which could subsequently trigger increased metabolism, respiration, ROS production, and ATP synthesis. However, the precise mechanisms underlying this connection require further investigation and are beyond the scope of the present study.



