A Meier-Gorlin syndrome mutation in a conserved C-terminal helix of Orc6 impedes origin recognition complex formation
- University of California, Berkeley, United States
- University of Alabama at Birmingham School of Medicine, United States
- Lawrence Berkeley National Laboratory, United States
- Howard Hughes Medical Institute, University of California, Berkeley, United States
Figures
Figure 1 with 3 supplements
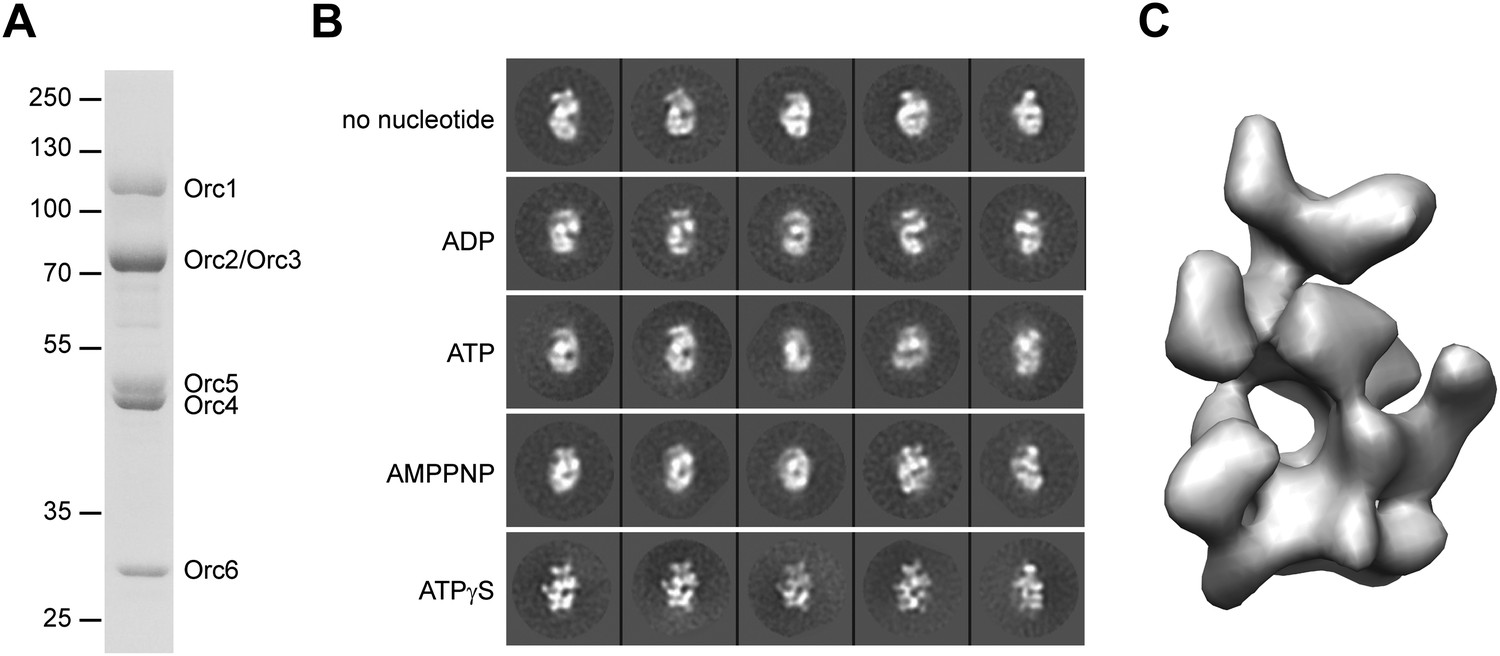
3D structure of metazoan ORC.
(A) Coomassie-stained SDS-PAGE gel of purified recombinant Drosophila ORC used for EM studies. Molecular weight markers and ORC subunits are indicated. (B) Nucleotides have different effects on the conformation of ORC. Drosophila ORC was incubated with 1 mM ADP, ATP, AMPPNP or ATPγS and analyzed by electron microscopy and compared to apo-ORC. Representative 2D class averages are shown. (C) Single-particle EM reconstruction of Drosophila ORC corrected for the absolute hand of ORC.
Figure 1—figure supplement 1
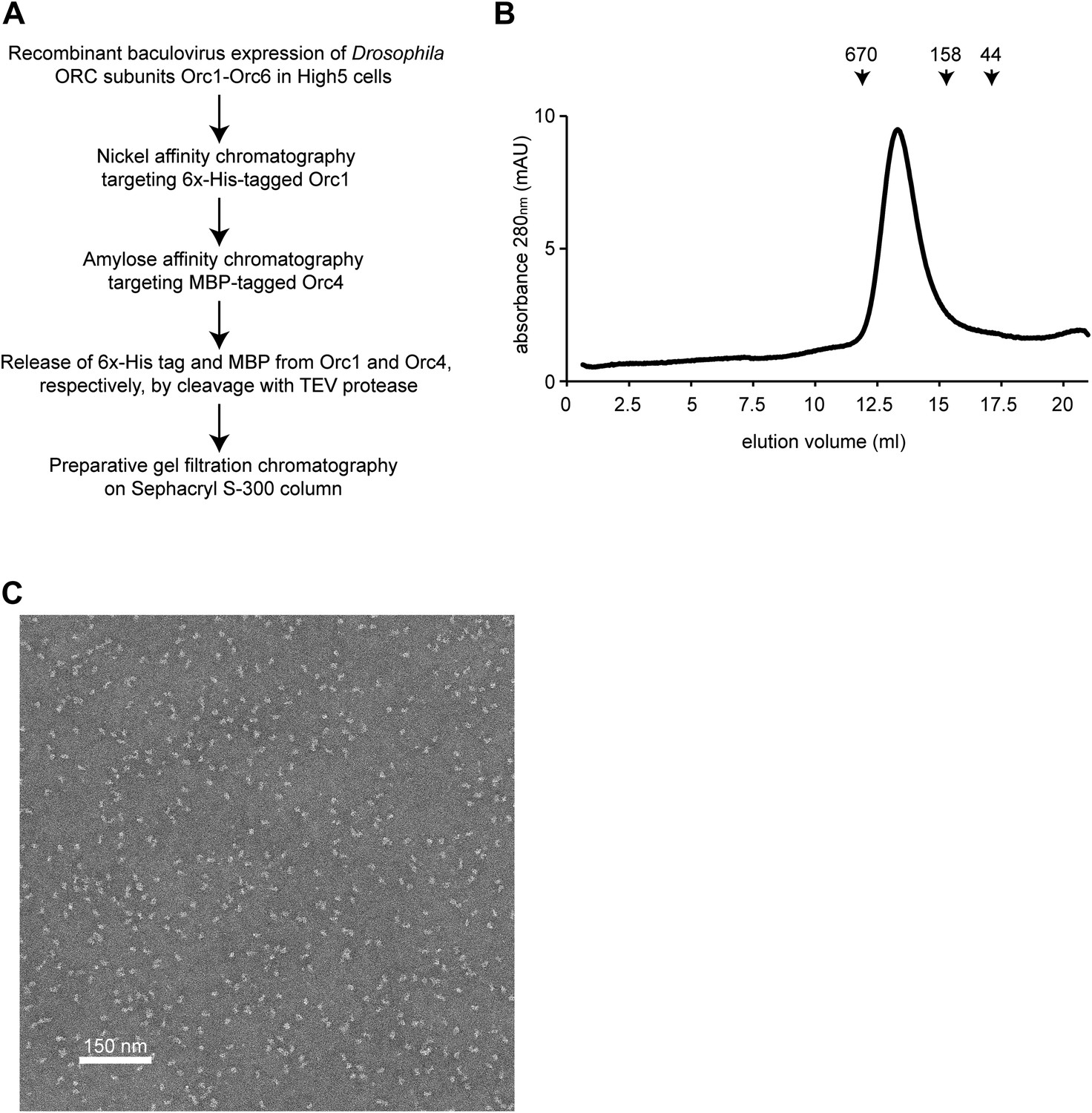
Purification of Drosophila ORC.
(A) Overview of recombinant expression and purification strategy used for ORC. (B) Analytical gel filtration chromatography of purified Drosophila ORC on a Superose 6 column demonstrates that Drosophila ORC elutes as a hexamer with the predicted molecular weight of ∼390 kDa. Elution volume of the molecular weight markers thyroglobulin (670 kDa), γ-globulin (158 kDa) and ovalbumin (44 kDa) are indicated above the UV trace. (C) Electron micrograph of negatively stained Drosophila ORC showing a monodisperse population of particles.
Figure 1—figure supplement 2
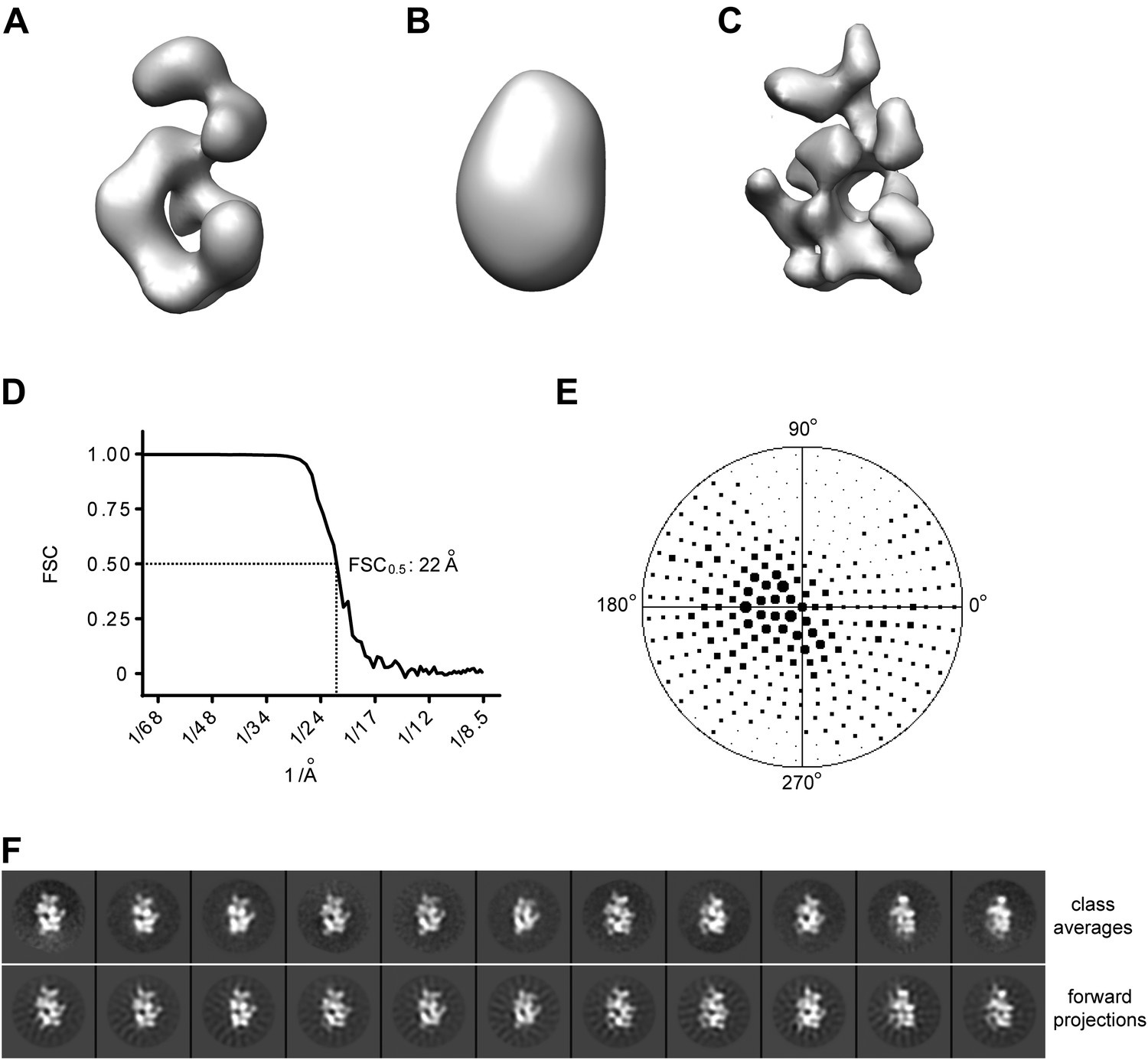
Single particle 3D EM reconstruction of Drosophila ORC.
(A) Previously-determined EM structure of Drosophila ORC (EMDB-1252) (Clarey et al., 2006). (B) Initial starting model used for projection-matching refinement obtained by low-pass filtration of the EM structure in (A) using a Butterworth filter with pass band at 200 Å and stop band at 100 Å. (C) Refined Drosophila ORC EM structure determined in this study prior to hand determination and flipping (Figure 1C, Figure 1—figure supplement 3). (D) Fourier shell correlation curve for reconstructed EM volume. The resolution at 0.5 FSC is ∼22 Å. (E) Euler angle distribution of particle images used for 3D reconstruction. The points in the hemisphere of the Euler space correspond to particle images assigned to a given reference image during projection-matching refinement, with the size of the point being proportional to the number of images assigned to a given reference. (F) Comparison of experimental 2D class averages with forward projection of final 3D EM volume demonstrates good matches between the experimental data and the 3D reconstruction.
Figure 1—figure supplement 3
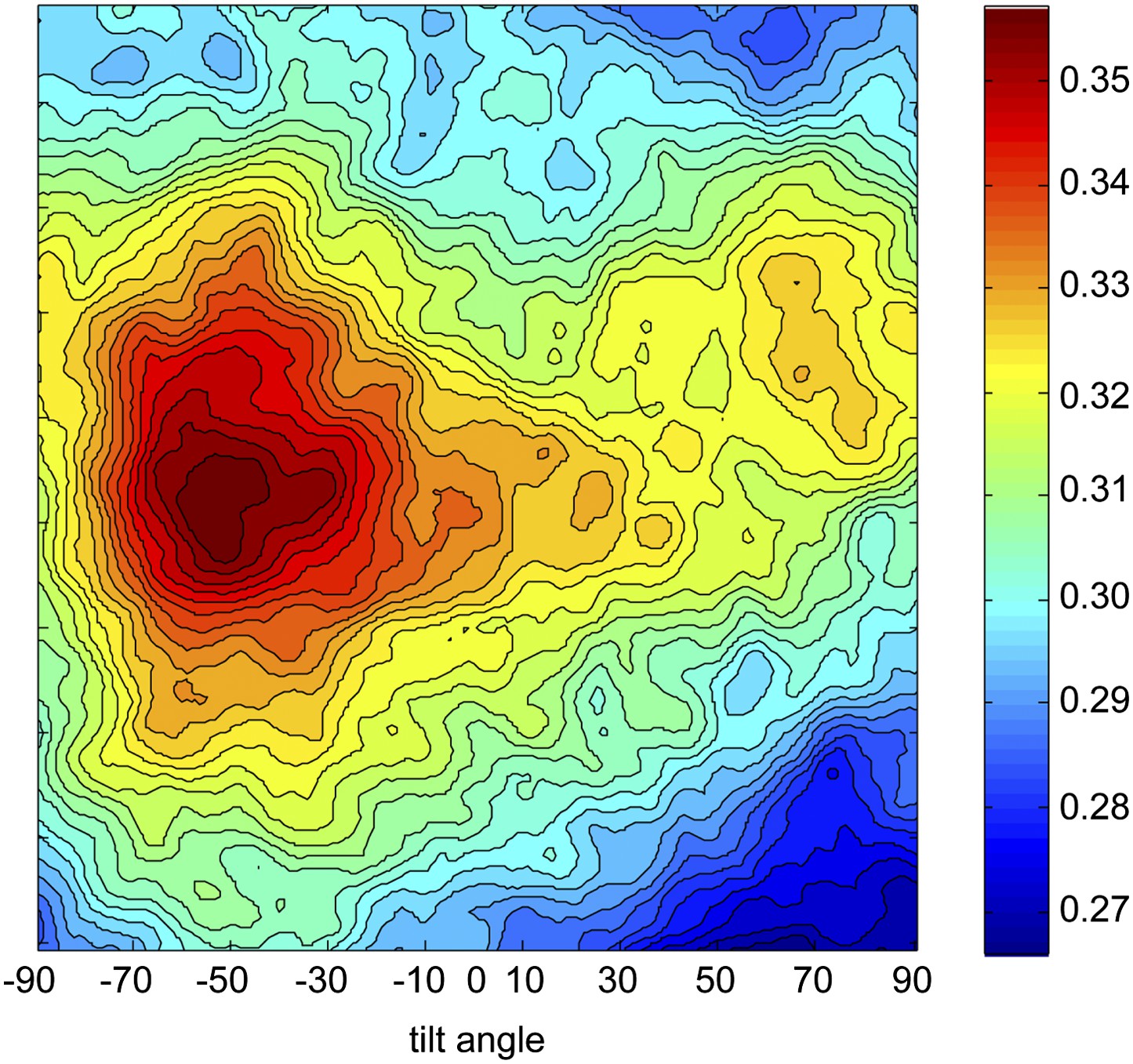
Determination of the absolute hand of ORC.
The final refined Drosophila ORC EM volume shown in Figure 1—figure supplement 2C was used in the freehand test using 0° and +50° tilt-pair images. The cross-correlation coefficients for the tilt transformations are plotted. The highest correlation is observed at a tilt angle of −50° and not +50°, indicating the absolute hand of the reconstructed ORC volume in Figure 1—figure supplement 2C is incorrect. Flipping the hand corrects the handedness of the model (Figure 1C).
Figure 2 with 1 supplement

Arrangement of subunits within metazoan ORC.
(A) Localization of AAA+ subunits Orc1–Orc5 within ORC by EM. 2D class averages of ORC containing N-terminally MBP-tagged or C-terminally GFP-tagged Orc1–5 subunits are compared to representative averages of untagged ORC. Additional densities observed that can be attributed to MBP- or GFP-tags are noted by arrowheads and mark the position of the respective subunit within ORC. 3D EM reconstructions of MBP- or GFP-tagged ORCs further pinpoint the localization of Orc1–Orc5 within ORC. With the exception of MBP–Orc2ΔN, MBP- or GFP-densities (highlighted in color) were also observed in 3D EM reconstructions of MBP- or GFP-tagged ORCs. No additional density was observed in 2D class averages for ORC containing N-terminally MBP-tagged full-length Orc1 and Orc2 nor for C-terminally GFP-tagged Orc2 and Orc4, indicating conformational flexibility of the tag (data not shown). NA—this view was not observed in 2D class averages. (B) Surface coloring of the DmORC EM volume based on subunit localization. The asterisks marks density that likely contains a domain inserted between the AAA+-like domain and the winged helix domain in Orc3 (Figure 5).
Figure 2—figure supplement 1
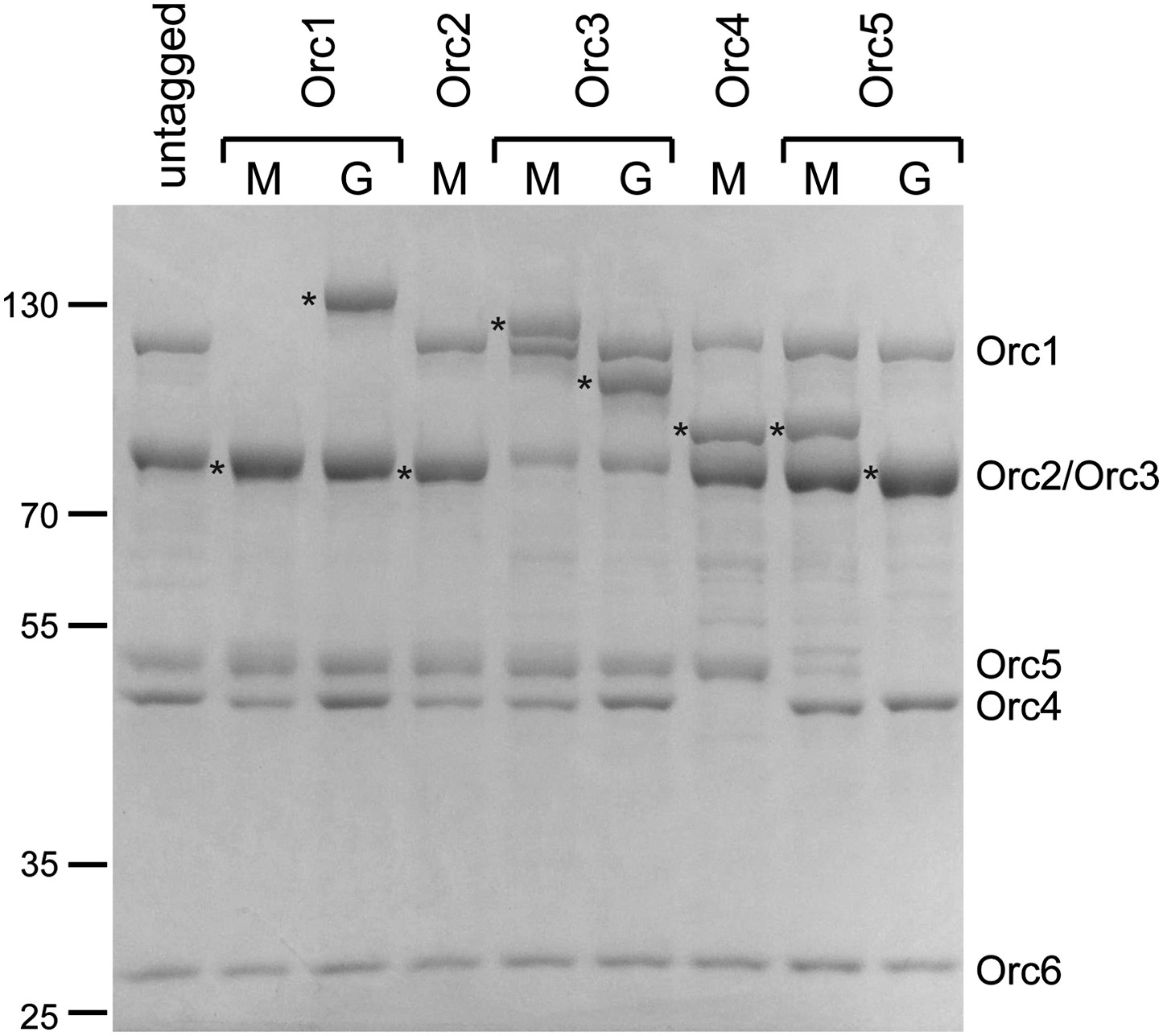
Purification of ORC with tagged Orc1–5 subunits for EM localization.
The AAA+ subunits Orc1–Orc5 were tagged at the N-terminus with MBP (M) or at the C-terminus with GFP (G) and the respective tagged ORC complexes purified. A Coomassie-stained SDS-PAGE gel of the complexes is shown. For each tagged complex, the tagged subunit is marked by an asterisk to the left of the lane. The N-terminal extensions in Drosophila Orc1 and Orc2 were deleted in MBP–Orc1 and MBP–Orc2.
Figure 3 with 1 supplement
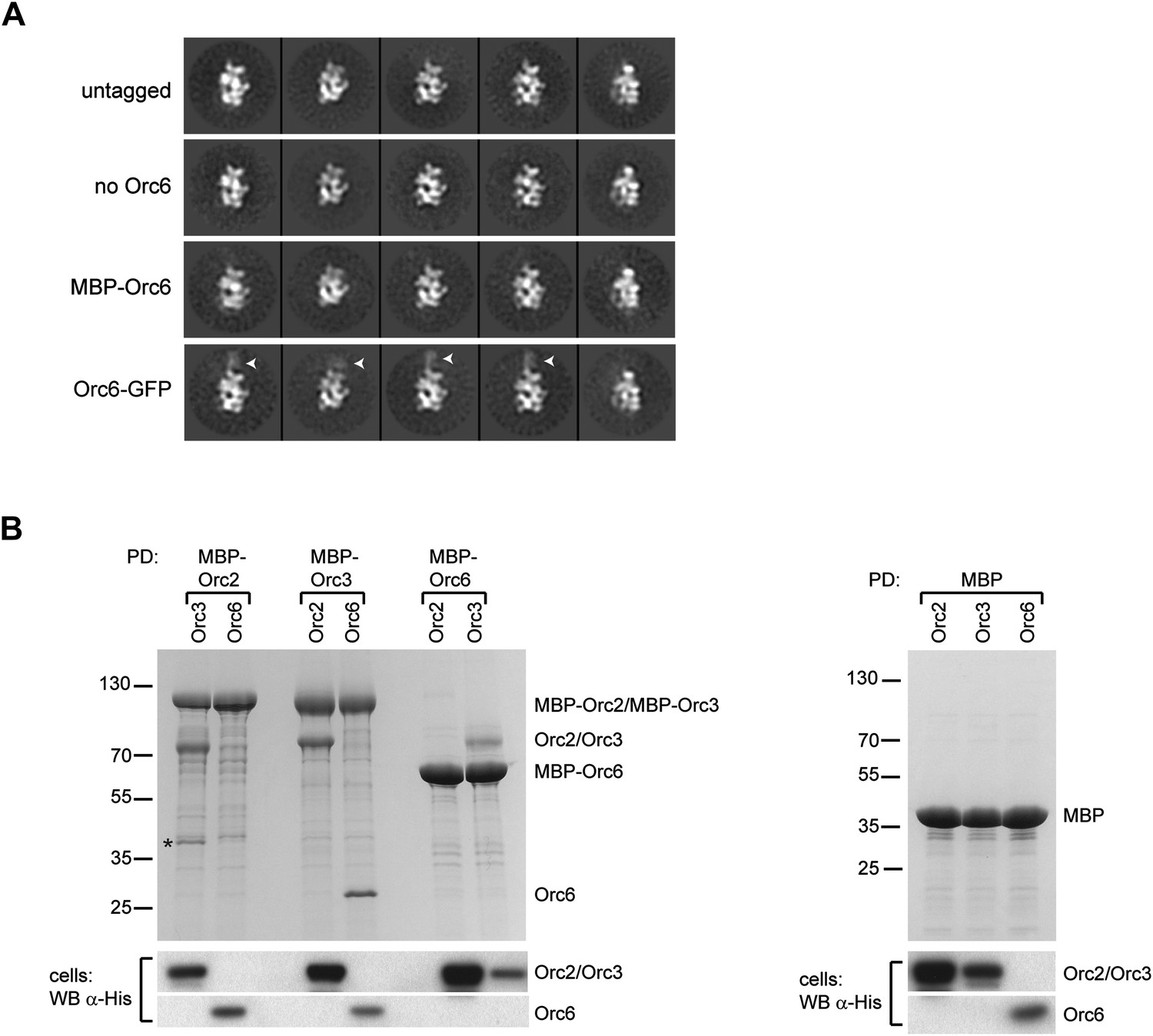
Orc6 localizes near Orc2 and Orc3 within Drosophila ORC but is conformationally flexible.
(A) ORC containing N-terminally MBP-tagged Orc6 or C-terminally GFP-tagged Orc6 was purified and subjected to EM analysis. Representative 2D class averages are shown. Drosophila ORC with untagged Orc6 is shown for comparison. Additional density corresponding to the tag (arrowheads) is only observed for the Orc6-GFP and not for MBP–Orc6. Class averages of ORC lacking Orc6 subunit reveal no difference density compared to ORC containing Orc6, indicating that Orc6 is flexibly tethered to ORC. (B) Drosophila Orc6 interacts with Orc3 but not with Orc2. MBP–Orc2, MBP–Orc3 and MBP–Orc6 fusions were co-expressed in High5 cells with 6 × His-tagged Orc2, Orc3 and Orc6 and subjected to pull-downs (PD) using amylose resin. Orc3 interacts with both Orc2 and Orc6, whereas no interaction is observed between Orc2 and Orc6 as assessed by SDS-PAGE and Coomassie staining. In addition, no interaction is observed between MBP alone and any of the ORC subunits. The asterisk denotes a degradation product of MBP–Orc2. Western blot (WB) analysis of whole cell extracts for 6 × His tagged subunits (lower panels) demonstrates that they are expressed as expected.
Figure 3—figure supplement 1
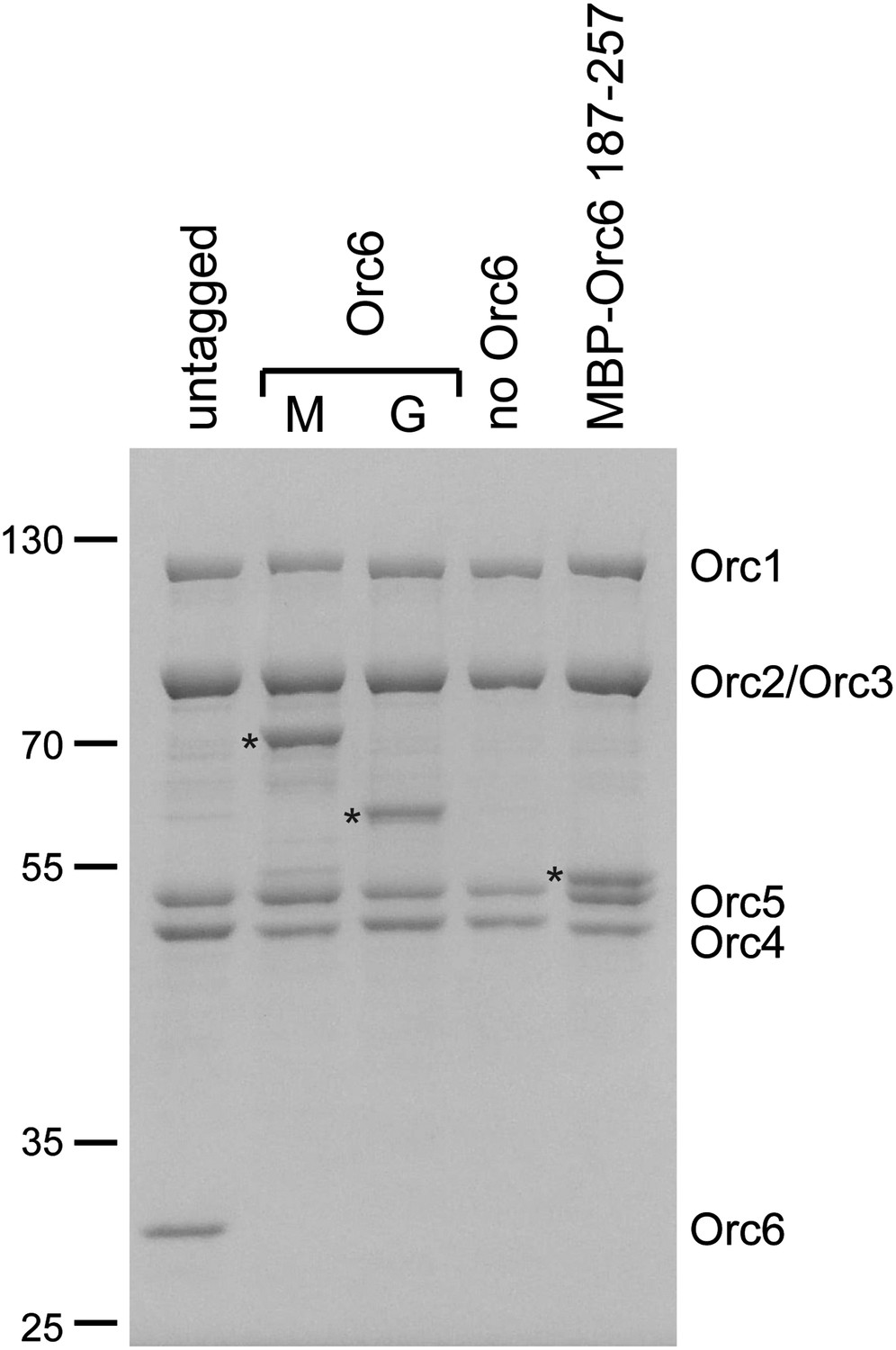
Purification of ORC complexes for EM localization of the Orc6 subunit.
ORC containing N-terminally MBP-tagged (M) Orc6, C-terminally GFP-tagged (G) Orc6, or the N-terminally MBP-tagged C-terminal domain of Orc6 (amino acids 187–257) and ORC lacking the Orc6 subunit were purified. A Coomassie-stained SDS-PAGE gel of respective complexes is shown. Tagged Orc6 is marked by an asterisk to the left of the lane.
Figure 4 with 2 supplements
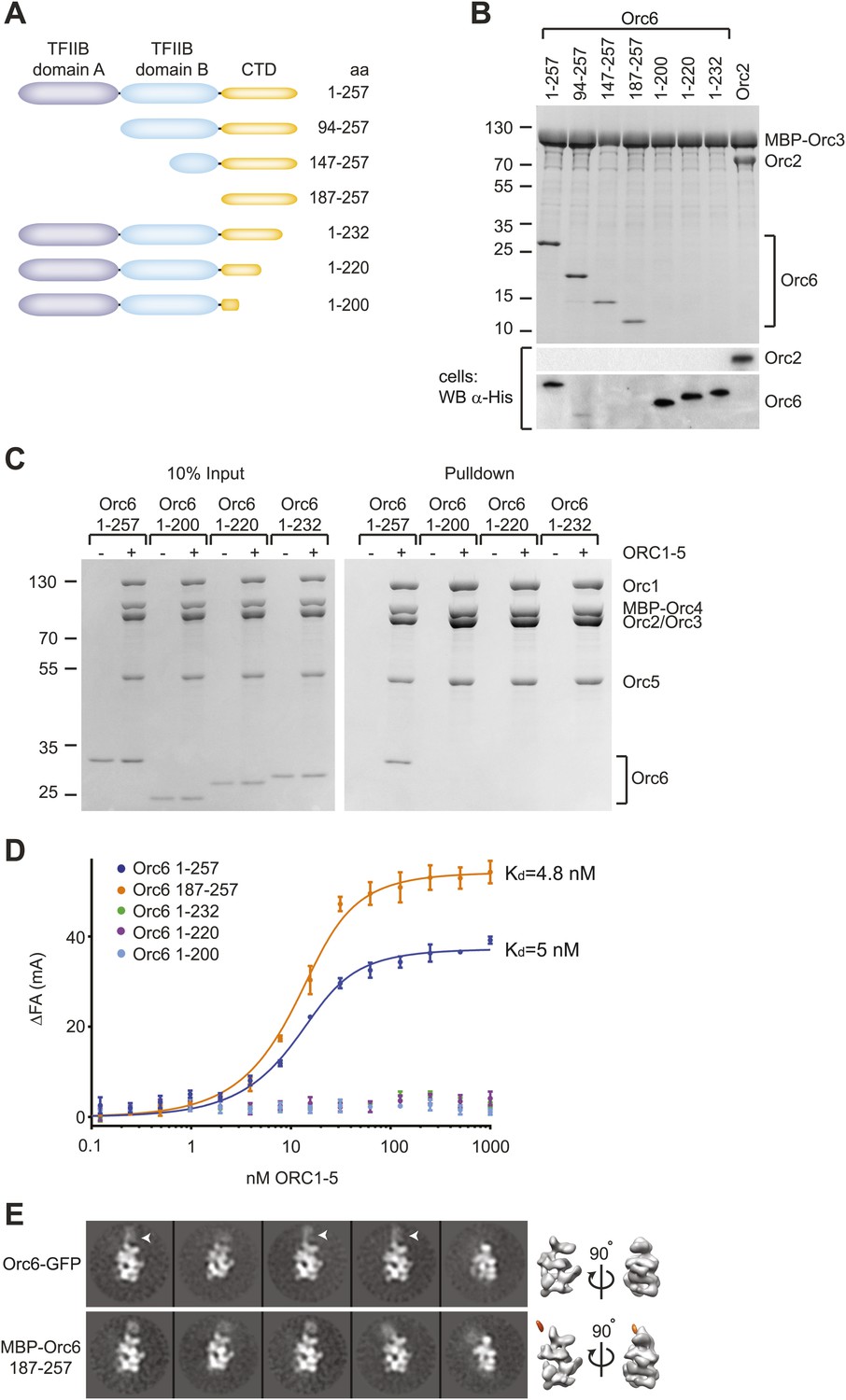
Drosophila Orc6 is recruited into ORC via interaction of its C-terminal domain with Orc3.
(A) Schematic of domain organization of Orc6 and of the N- and C-terminal deletion constructs of Orc6 used in this study. (B) The C-terminal domain (CTD) of Orc6 is essential and sufficient for the interaction of Orc6 with Orc3. His-tagged full-length Orc6 (1–257) and Orc6 deletion constructs were co-expressed with MBP-tagged Orc3 in High5 cells and analyzed for interactions by pull-downs using amylose resin. Bound proteins were analyzed by SDS-PAGE and Coomassie-staining. Western blot analysis of whole cell extracts (lower panel) shows that C-terminally truncated Orc6 proteins are expressed at similar levels to full-length wild type Orc6. Despite the clear pull-down results for the N-terminally truncated Orc6 proteins, we were not able to detect two of the proteins (Orc6 147–257 and 187–257) in whole cell extracts, possibly due to expression levels too low to be detected with our Western blot conditions. (C and D) The C-terminal domain of Orc6 is essential and sufficient for recruitment of Orc6 into ORC. Purified ORC1–5 (lacking Orc6 subunit) was incubated in vitro with purified full-length Orc6, Orc6 lacking C-terminal residues (Orc6 1–232, Orc6 1–220 and Orc6 1–200), and the isolated Orc6 C-terminal domain (Orc6 187–257). Formation of ORC containing all six subunits was tested in pull-downs using (C) MBP-tagged Orc4 as bait and by (D) fluorescence anisotropy after N-terminally labeling Orc6 proteins with Alexa Fluor-488. C-terminal deletions of Orc6 abolish recruitment of Orc6 into ORC in vitro, whereas the C-terminal domain of Orc6 binds to ORC1–5 with nanomolar affinity. Binding data of full-length Orc6 and the Orc6 C-terminal domain (Orc6 187–257) were fit to the quadratic equation describing single-site binding under ligand depletion. (E) The localization of the C-terminal domain of Orc6 is indistinguishable from the localization of full-length Orc6 within ORC. Purified ORC containing full-length Orc6-GFP or MBP–Orc6 187–257 was analyzed by negative stain EM. 2D image analysis revealed extra densities (arrow heads) in similar positions that correspond to GFP and MBP tags on the respective Orc6 constructs. Density corresponding to the affinity tag was observed in the 3D volume of ORC containing MBP–Orc6 187–257, further pinpointing the localization of Orc6 within ORC.
Figure 4—figure supplement 1
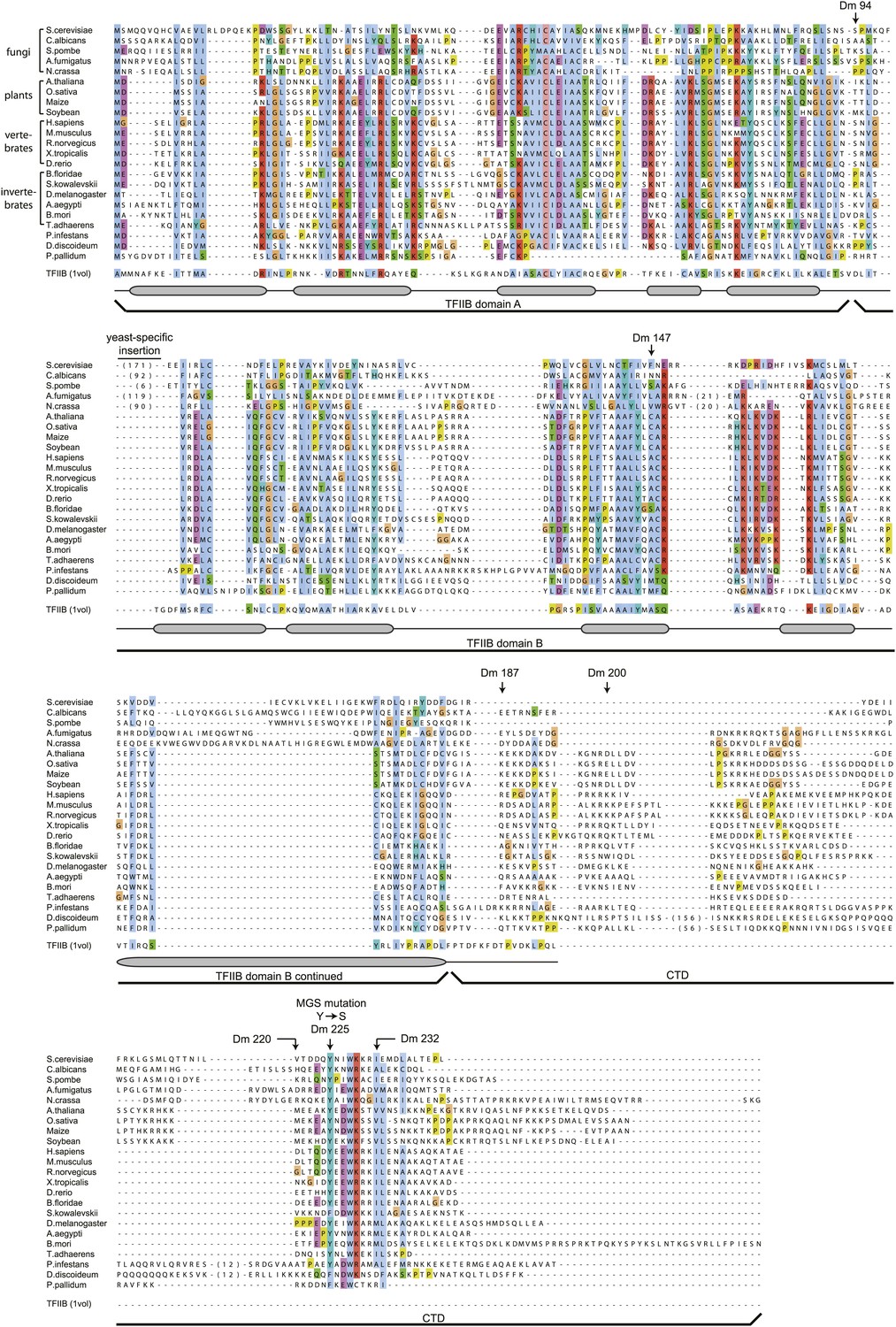
The Orc6 TFIIB-like domains as well as residues in the C-terminal domain containing the Meier–Gorlin syndrome mutation in human Orc6 are conserved across eukaryotes, including fungi.
Orc6 protein sequences from representative eukaryotes and human transcription factor TFIIB (PDB code 1vol, chain A) (Nikolov et al., 1995) were aligned using MAFFT (Katoh et al., 2005; Katoh and Toh, 2008). Conserved domains and secondary structure elements as observed in the crystal structure of human TFIIB are outlined below the alignment. Amino acid residues in Drosophila melanogaster (Dm) Orc6 that demarcate truncations as well as the residue mutated in Meier-Gorlin syndrome (MGS) are indicated above the alignment. Accession numbers of Orc6 protein sequences used in the alignment can be found in the ‘Materials and methods’.
Figure 4—figure supplement 2
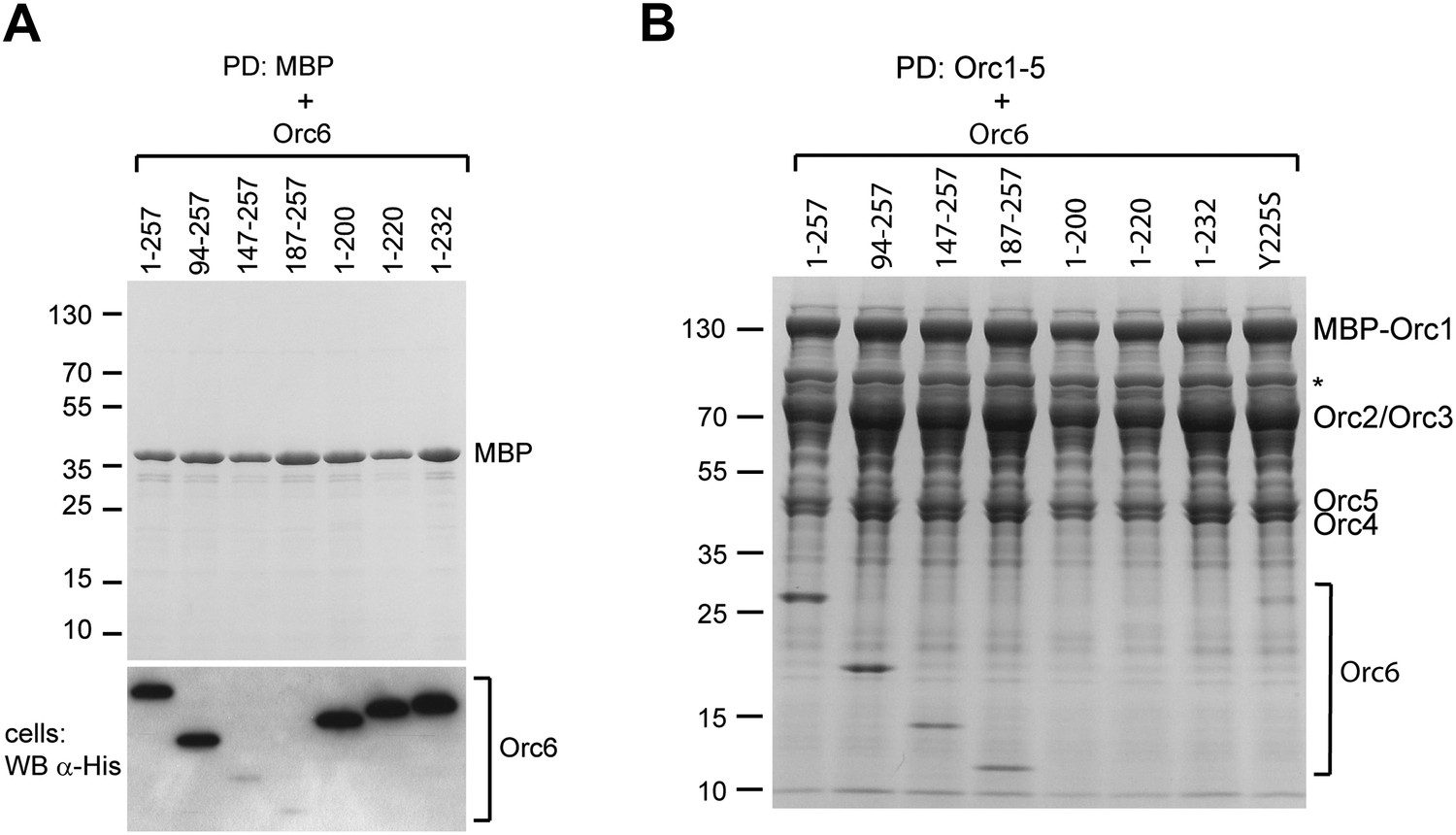
C-terminal truncations of Drosophila Orc6 do not associate with ORC1–5 when all subunits are co-expressed in insect cells.
(A) Orc6 proteins do not interact with MBP nor do they bind unspecifically to amylose beads. MBP and different Orc6 constructs were co-expressed in High5 cells and pull-downs (PD) were performed using amylose beads. Only MBP is recovered from beads. Western blots (WB) show that Orc6 proteins are expressed in insect cells. We were not able to detect Orc6 147–257 and Orc6 187–257 in Western blots of whole-cell lysates; however, the two proteins efficiently co-purified with MBP–Orc3 (Figure 4B), MBP–Orc3INS (Figure 5C), and ORC1–5 (panel B in this figure), indicating they are expressed. (B) The C-terminal domain of Orc6 is essential for hexameric ORC formation in insect cells. ORC subunits and Orc6 truncated proteins were co-expressed in insect cells and extracts were subjected to amylose pull-downs (PD) targeting MBP-tagged Orc1. Whereas N-terminal deletions of Orc6 co-purify with ORC1–5, C-terminal deletions of Orc6 abrogate the interaction with ORC1–5. The asterisk marks a protein contaminant. Coomassie-stained SDS-PAGE gels are shown in (A and B).
Figure 5 with 1 supplement
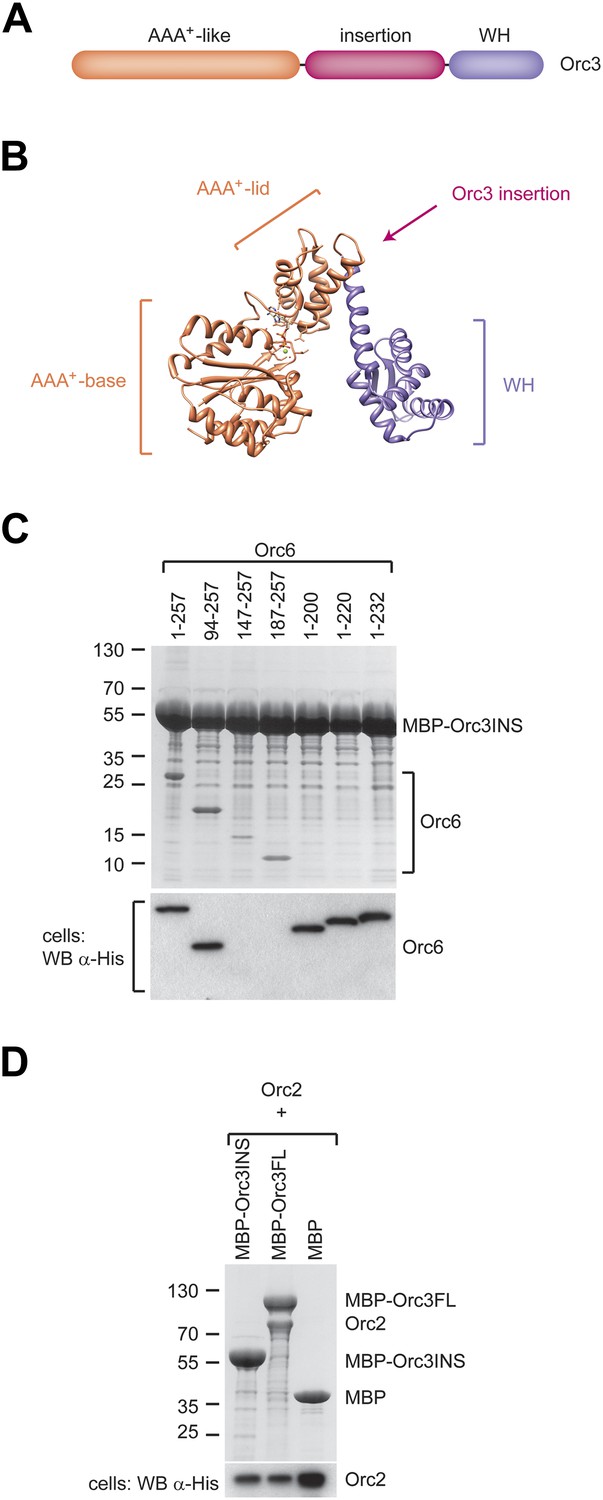
A conserved domain inserted between the AAA+-like domain and the winged helix (WH) domain of Orc3 binds the C-terminal domain of Orc6 and recruits Orc6 into ORC.
(A) Schematic domain organization of Orc3. (B) The position of the Orc3 insertion maps between the AAA+-like domain and the winged helix domain in the crystal structure of archaeal Orc1–1 (PDB code 2qby chain A) (Dueber et al., 2007). (C) The Orc3 insertion is sufficient for Orc6 binding. Drosophila Orc3 containing residues 371–559 (Orc3INS) was co-expressed as an N-terminal MBP-fusion protein with His-tagged full-length and truncated versions of Drosophila Orc6 in High5 cells and their interaction probed by pull-downs using amylose resin. (D) The insertion in Orc3 does not interact with Orc2. N-terminal MBP-fusion proteins of Drosophila full-length Orc3 (Orc3FL), the Orc3 insertion (Orc3INS) or MBP alone were co-expressed with 6 × His-tagged Drosophila Orc2 in High5 cells and subjected to amylose pull-downs. Coomassie-stained SDS-PAGE gels are shown in (C and D). Western blot analysis of whole-cell extract using an anti-His antibody (lower panels in C and D) demonstrates that lack of Orc2 or Orc6 pull-down is not due to lack of their expression. As in Figure 4B, we were not able to detect Orc6 147-257 and Orc6 187-257 in cell extracts despite being clearly visible in pull-downs, likely because of low expression levels.
Figure 5—figure supplement 1
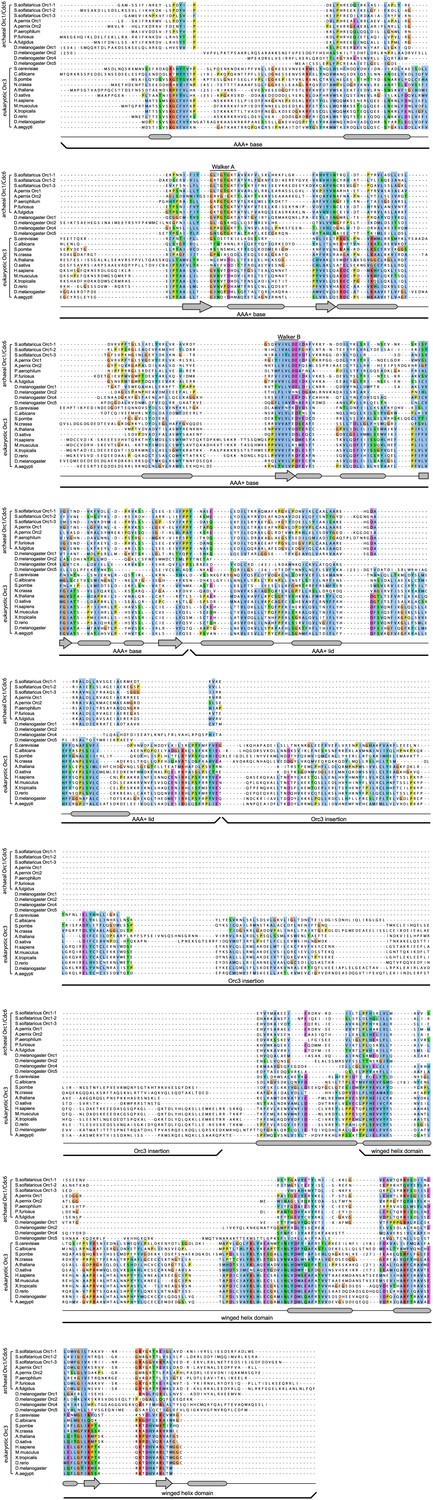
Eukaryotic Orc3 contains a domain inserted between its AAA+-like and winged helix domains that is not present in archaeal Orc1/Cdc6, nor in eukaryotic Orc1, Orc2, Orc4 or Orc5.
Protein sequences of archaeal Orc1/Cdc6, various eukaryotic Orc3 proteins, and Drosophila melanogaster Orc1, Orc2, Orc4, and Orc5 were aligned using MAFFT (Katoh et al., 2005; Katoh and Toh, 2008). Secondary structural elements as seen in the crystal structure of archaeal Orc1–1 (PDB code 2qby, chain A) (Dueber et al., 2007) are shown below the alignment. Accession numbers for protein sequences can be found in the ‘Materials and methods’.
Figure 6 with 1 supplement
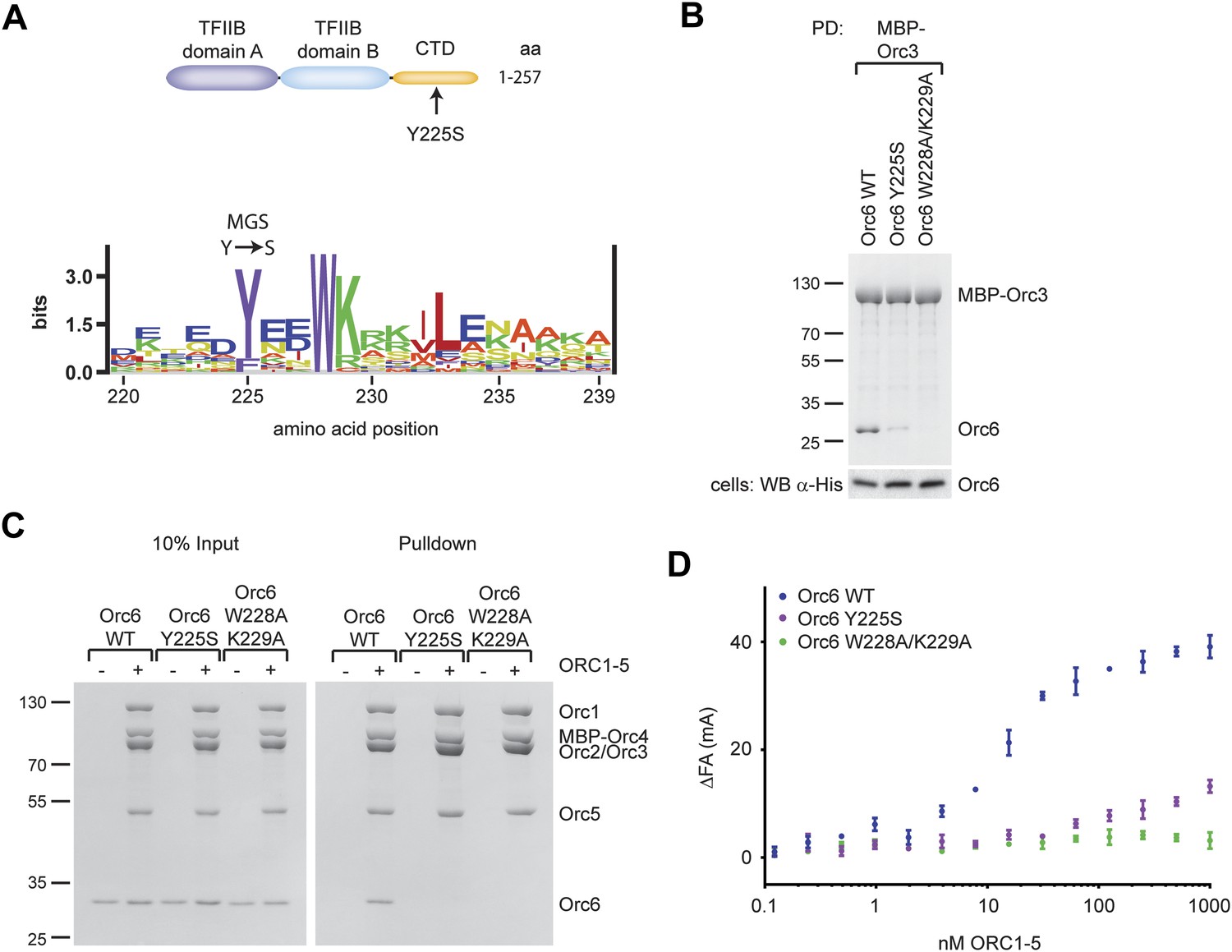
A Drosophila Orc6 mutation corresponding to the Meier-Gorlin-Syndrome mutation in human Orc6 weakens the interaction of Orc6 with Orc3 and diminishes its recruitment into ORC.
(A) The tyrosine mutated in Meier-Gorlin syndrome (MGS) is a well conserved residue and maps to the C-terminal domain of Orc6. A sequence LOGO (Workman et al., 2005) of the Orc6 C-terminus is shown. The amino acid residue corresponding to the human MGS mutation was mutated in Drosophila Orc6 (Y225S). (B) Mutations in the C-terminus of Drosophila Orc6, including the MGS mutation, reduce the affinity of Orc6 for Orc3. Wild-type (WT) Orc6, the Y225S mutant or the W228A/K229A double-mutant Orc6 were co-expressed with MBP–Orc3 in High5 cells and tested for interaction with Orc3 by pull-down analysis using amylose beads and MBP–Orc3 as bait. Coomassie-stained SDS-PAGE gels are shown for pull-downs and a Western blot (lower panel) is shown to confirm Orc6 expression in insect cells. (C and D) C-terminal mutations in Drosophila Orc6 interfere with recruitment of Orc6 into ORC in vitro. Wild-type Orc6, Orc6-Y225S and Orc6-W228A/K229A were purified and tested for association with ORC1–5 into heterohexameric ORC using pull-downs targeting MBP–Orc4 (C) or by fluorescence anisotropy using N-terminally Alexa Fluor-488 labeled Orc6 (D).
Figure 6—figure supplement 1
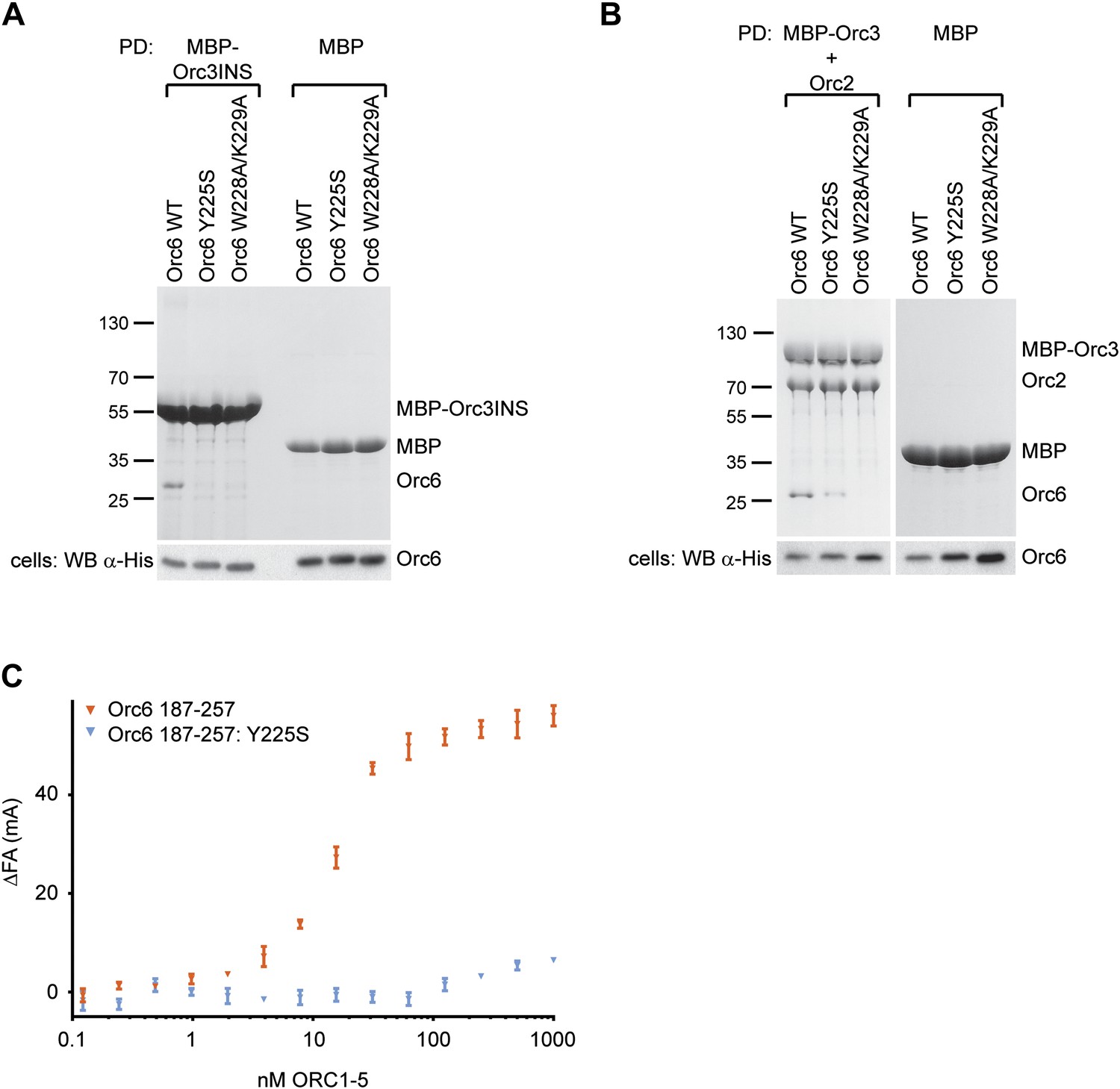
Mutations in the C-terminal domain of Drosophila Orc6, including the MGS-like mutation, abrogate binding of Orc6 to the Orc2–Orc3 subcomplex, to the Orc3 insertion (Orc3INS) and to ORC1–5.
(A and B) Amylose pull-downs (PD) from High5 cells co-expressing His-tagged Orc6 wild type (WT) or Orc6 containing the MGS-like (Y225S) or WK (W228A/K229A) mutation with either (A) MBP–Orc3INS or (B) MBP–Orc3 and Orc2. The results show that both mutations in the Orc6 C-terminus interfere with Orc6 binding to the insertion in Orc3 and with recruitment of Orc6 into the Orc2-Orc3-Orc6 subcomplex. Expression of prey proteins was verified in Western blots (WB) of whole cell extract (lower panels). (C) The MGS-like mutation in the context of the Orc6 C-terminal domain (CTD) alone significantly reduces the affinity of the Orc6-CTD for ORC1–5. The results of fluorescence anisotropy experiments with fluorescently labeled wild-type and Y225S Orc6-CTD are shown.
Figure 7

Mutations of conserved amino acids in the C-terminal domain of Drosophila Orc6 result in a loss of its association with ORC and in reduced MCM chromatin association in vivo in flies.
(A) Western immunoblotting analysis of ORC complexes precipitated using anti-Orc2 antibody from extracts isolated from fly ovaries expressing different GFP-tagged Orc6 mutant proteins: GFP–Orc6 D224A/Y225A, GFP–Orc6 W228A/K229A and GFP–Orc6 WT (wild type). GFP–Orc6 fusion proteins were detected with anti-GFP monoclonal antibody, while Orc3 and Orc5 subunits were detected using anti-Orc3 or anti-Orc5 antibodies. Ovary extract (In), supernatant after immunoprecipitations (S) and immunoprecipitated material (IP) are shown for each transgene. (B) Quantitation of immunoprecipitated GFP from ovary extracts containing different Orc6 mutants. The Y-axis shows fluorescence emission intensity. Mean and standard deviation from three independent experiments are plotted (p≤0.05). (C) Chromatin association of Mcm4, a member of the MCM2–7 complex, is reduced in orc6-null fly larvae expressing Orc6 mutants. Brains of homozygous, orc6-null (orc635/orc635) or heterozygous (orc635/Cy) Drosophila larvae expressing GFP–Orc6 transgenes were isolated and subjected to salt extraction to solubilize cellular proteins. Insoluble and chromatin associated proteins were pelleted by centrifugation and analyzed by Western blotting using Mcm4 polyclonal antibodies. Pnut was used as a loading control. Total brain extracts were also analyzed to ensure equal Mcm4 expression in larvae expressing Orc6 transgenes.
Figure 8
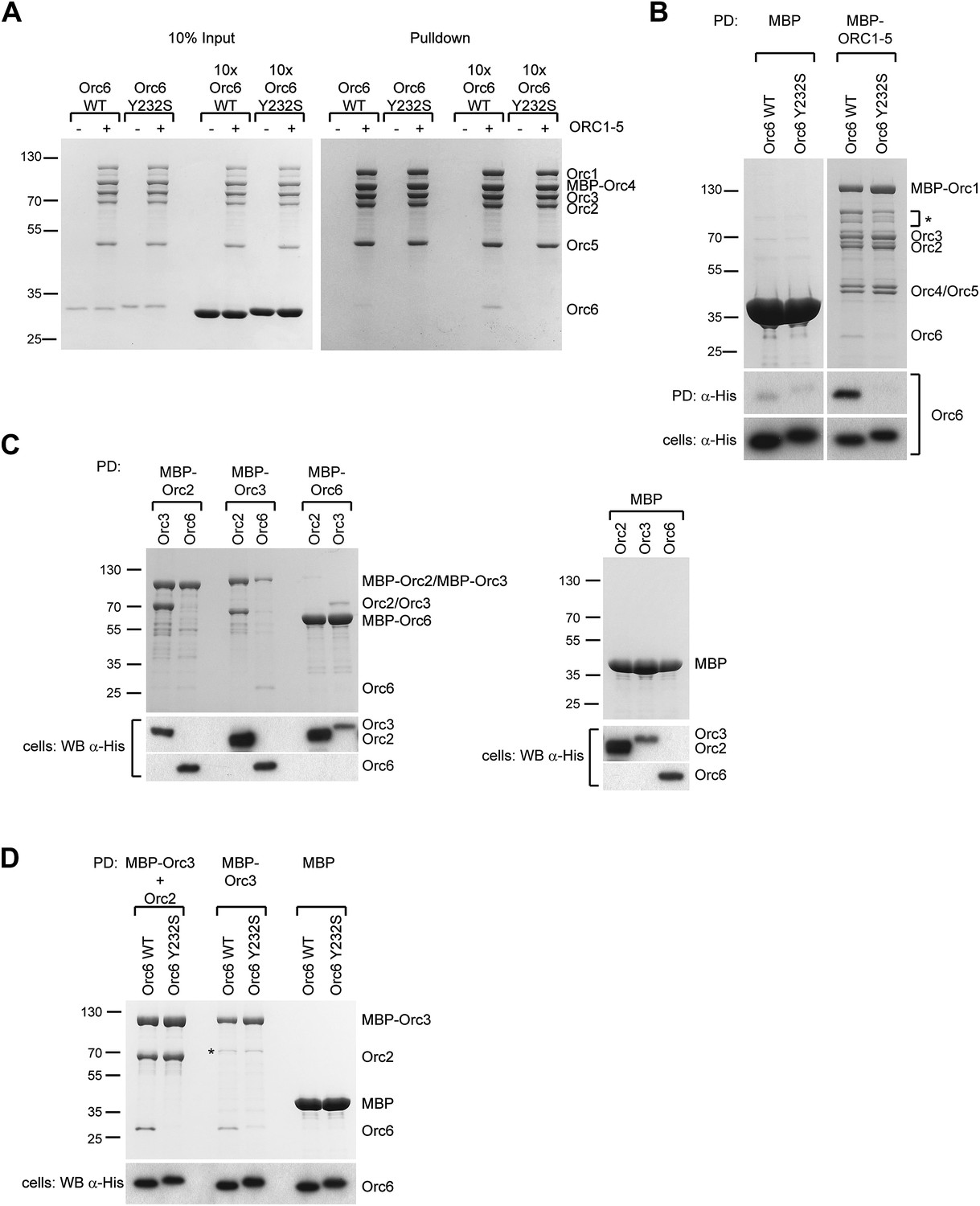
The Orc6–Orc3 interaction is conserved in humans and is affected by a mutation in human Orc6 that is found in MGS patients.
(A) Human wild-type (WT), but not MGS mutant (Y232S), Orc6 binds to human ORC1–5 in vitro. Purified Orc6 was incubated with recombinant ORC1–5 at either equimolar ratios or with a 10 × excess of Orc6. Orc6 binding to ORC1–5 was analyzed by pull-downs using amylose beads that bind MBP–Orc4. Contrary to wild-type Orc6, MGS-mutant Orc6 does not associate with ORC1–5, even when a 10-fold excess of Orc6 is used in binding reactions. (B) Human Orc6 containing the MGS mutation does not co-purify with human ORC1–5 when co-expressed in insect cells. High5 cells expressing human ORC1–5 and either Orc6-WT or Orc6-Y232S were used in pull-downs targeting MPB-Orc1. As a negative control, Orc6-WT or Orc6-Y232S were co-expressed with MBP. Only Orc6-WT is recovered from beads, and only when Orc1–Orc5 are co-expressed. The presence of Orc6 was detected by Western blotting in both pull-downs (PD) and whole-cell extracts (lower panel). The asterisk marks degradation products of MBP–Orc1. (C) Subunit interactions within the human Orc2-Orc3–Orc6 subcomplex. MBP-tagged human ORC subunits were co-expressed in High5 cells with 6 × His-tagged subunits in a pairwise manner and probed for interaction by amylose pull-downs. Interactions are observed between human Orc3 and Orc2 as well as between Orc3 and Orc6. As a control, 6 × His-tagged subunits were co-expressed with MBP alone, and no interaction is observed with MBP. Western blots (lower panels) verified that all His-tagged subunits are present in whole cell extracts. (D) The MGS mutation in human Orc6 decreases the affinity of human Orc6 for human Orc3 and reduces Orc6 recruitment into the Orc2-Orc3-Orc6 subcomplex. Human ORC subunits were co-expressed in High5 cells as indicated and analyzed by amylose pull-downs for binding. The asterisk marks a degradation product of MBP–Orc3. Western blot of cell lysate shows that wild-type and mutant Orc6 are expressed in insect cells at similar levels (lower panel). Coomassie-stained gels are shown for pull-downs in (A–D). Orc6-Y232S always migrated slightly slower than wild-type Orc6 in SDS-PAGE gels. Mass spectrometry confirmed that the molecular weight of wild type and Y232 Orc6 are as expected based on their amino acid sequence.
Figure 9 with 1 supplement

The C-terminal domain in S. cerevisiae Orc6 interacts with Orc3 and an MGS-like mutation abrogates this interaction.
(A) S. cerevisiae Orc6 binds Orc3. His-tagged Orc2, Orc3 or Orc6 were co-expressed with MBP–Orc2, MBP–Orc3 or MBP only in High5 cells and cell extracts were subjected to pull-downs (PD) using amylose beads. Orc6 and Orc3, as well as Orc2 and Orc3, interact directly with each other. MBP–Orc2 was very unstable when expressed without Orc3. The asterisk indicates a degradation product of either MBP–Orc3 or Orc2. (B) An MGS-like mutation in the C-terminal domain of S. cerevisiae Orc3 abolishes binding of Orc6 to Orc3. Wild-type (WT) Orc6 or Orc6 containing a mutation corresponding to the MGS mutation in human Orc6 (Y418S) were co-expressed with either MBP–Orc3 or MBP only in insect cells. Only wild-type S. cerevisiae Orc6, but not the Y418S mutant, bound to MBP–Orc3 in amylose pull-downs. (C) The MGS-like mutation in S. cerevisiae Orc6 allows incorporation of Orc6 into the Orc2-Orc3-Orc6 subcomplex. Orc6-WT and Orc6-Y418S were co-expressed with untagged Orc2 and MBP–Orc3 in High5 cells. In amylose pull-downs, a ternary complex containing Orc2, Orc3, and Orc6 was recovered for both Orc6-WT and Orc6-Y418S. Since Orc6-Y418S does not interact with Orc3 alone (panel B) this result suggests that a second binding site for Orc6 exists in S. cerevisiae Orc2 which is not sensitive to the MGS-like mutation in the C-terminal domain of Orc6. (D) The S. cerevisiae Orc3–Orc6 interaction is not essential for hexameric ORC formation. S. cerevisiae ORC subunits were co-expressed in High5 cells with Orc6-WT or Orc6-Y418S and cell extracts were subjected to pull-downs (PD) using amylose beads binding to MBP–Orc1. All six subunits co-purify with MBP–Orc1. Due to the similar migration of Orc6 and Orc4/Orc5, bead-bound Orc6 was detected by Western blotting using anti-His antibody. No Orc6 was recovered in control pull-downs from cells expressing Orc6 and MBP. (E) At least two different binding sites recruit S. cerevisiae Orc6 into ORC. S. cerevisiae ORC subunits were co-expressed in insect cells to reconstitute ORC containing all six ORC subunits (-), ORC lacking Orc2 (no Orc2) and ORC lacking Orc3 (no Orc3). Amylose pull-downs targeting MBP–Orc1 result in reduced Orc6 incorporation into ORC in the absence of Orc2, and no Orc6 is recovered when Orc3 is absent. Omission of Orc3 also resulted in loss of co-purification of Orc2. As in (D) Orc6 was detected by Western blotting (WB). Asterisks in (D and E) indicate a protein contaminant binding to amylose beads. In (A–E) Coomassie-stained SDS-PAGE gels are shown for pull-downs and Western blots (WB) of whole cell extract verify that all His-tagged subunits were expressed.
Figure 9—figure supplement 1
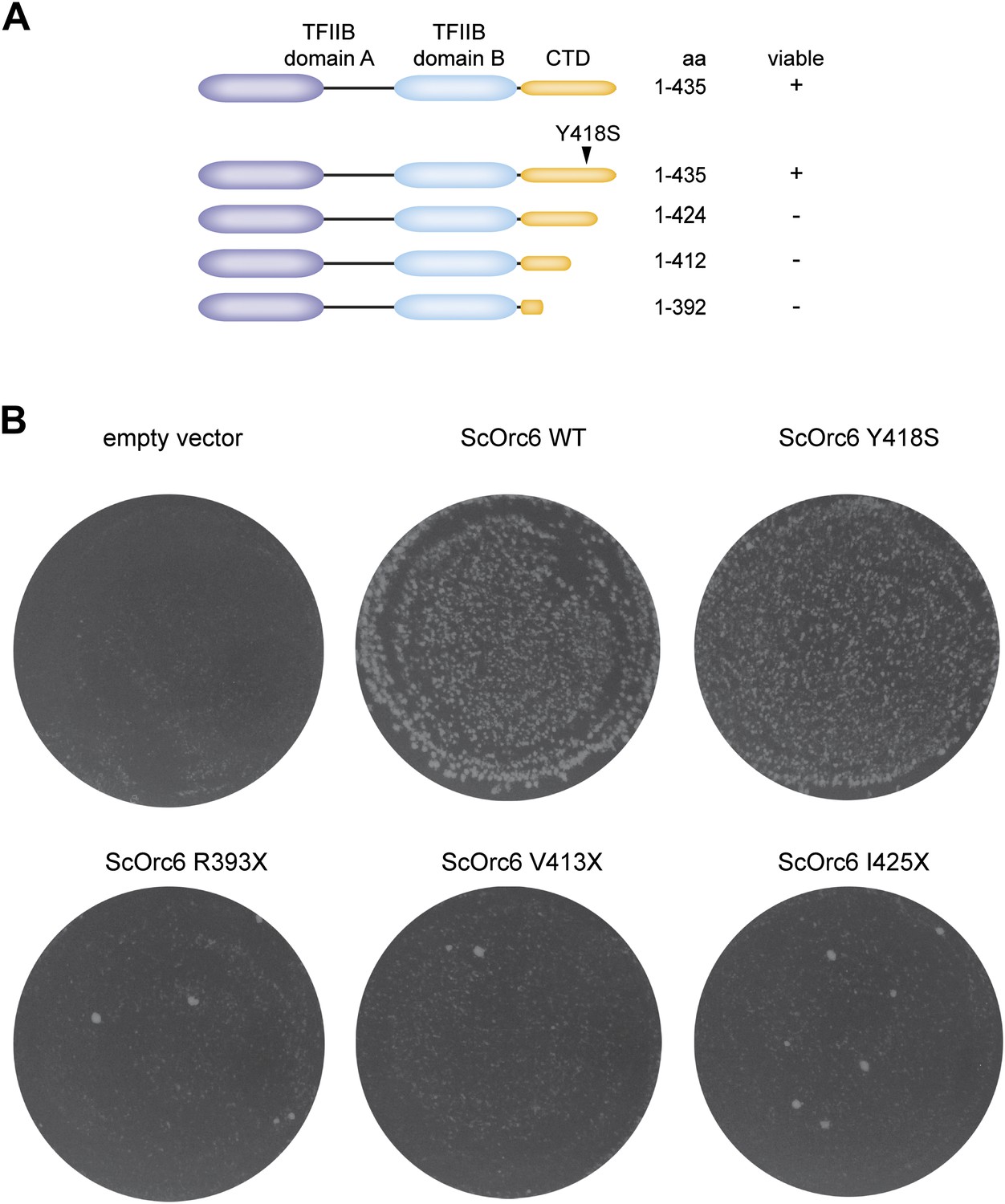
The C-terminus of S. cerevisiae Orc6 is essential for yeast growth.
(A) Schematic of Orc6 constructs tested for complementation. Orc6 lacking C-terminal amino acid residues was generated by introducing stop codons at the appropriate positions in the plasmid expressing Orc6. The results of the complementation analysis are summarized. (B) Complementation analysis of S. cerevisiae Orc6. Plasmids expressing no Orc6 (empty vector), Orc6 wild-type (WT) or mutated Orc6 as indicated were transformed into strain ySC166 carrying a degron fusion of the chromosomal copy of Orc6 (Chen et al., 2007). Transformants were selected for the plasmid in permissive growth conditions, and then replica-plated and grown in non-permissive conditions. The number of viable colonies is significantly reduced in cells expressing Orc6 lacking parts of the conserved C-terminal domain.
Figure 10
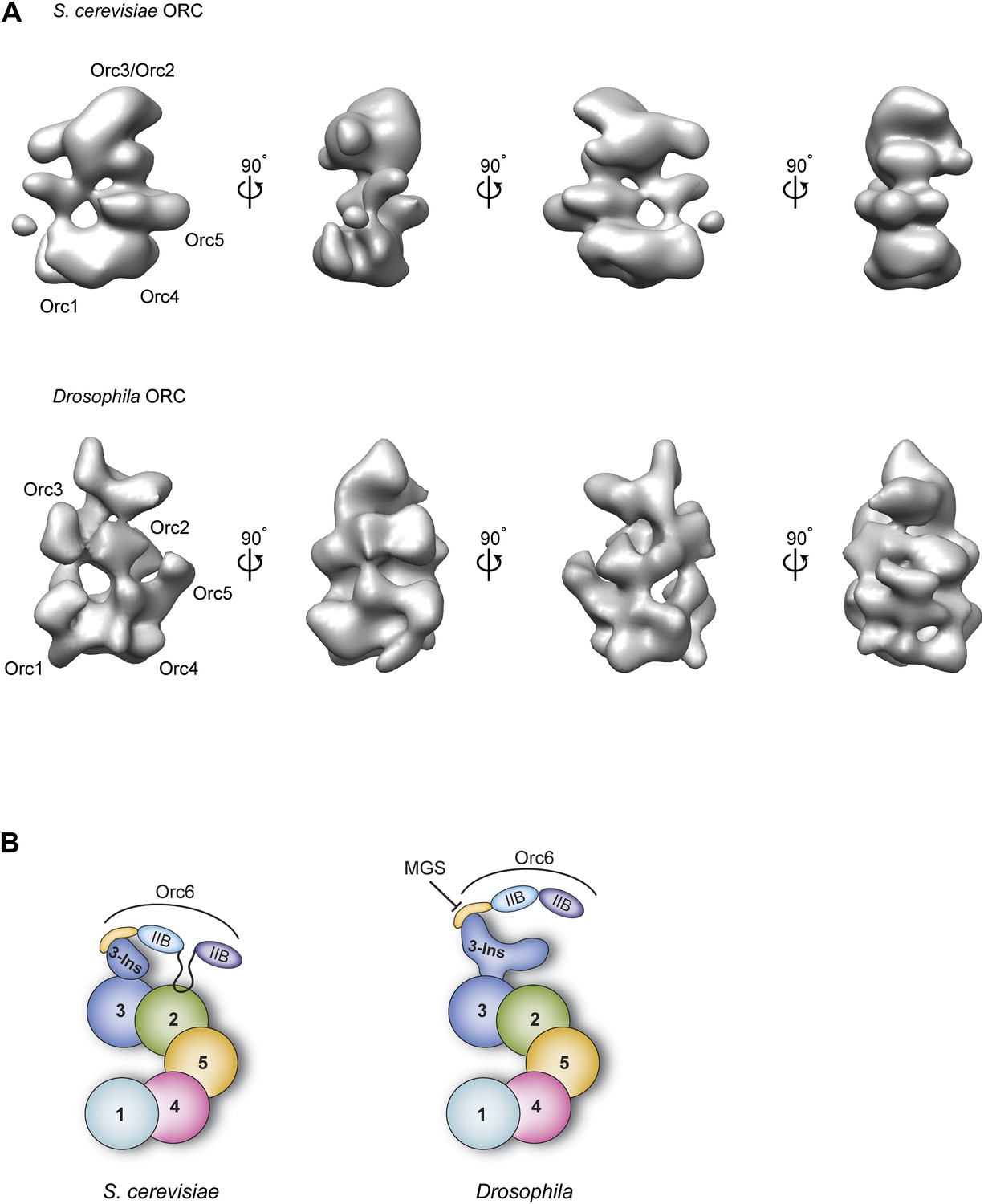
Comparison of metazoan and S. cerevisiae ORC architectures and model of how the MGS mutation in Orc6 impedes ORC formation.
(A) Different views of S. cerevisiae ORC (EMDB 1156, Speck et al., 2005) and the new Drosophila ORC structure (this study) are compared (a more recent, higher-resolution EM model of budding yeast ORC in isolation (Sun et al., 2012) has been not deposited into the EM database and so could not be used here). The localization of ORC subunits as reported for S. cerevisiae ORC (Chen et al., 2008; Sun et al., 2012) and Drosophila ORC (this study) are indicated. (B) Model for AAA+ subunit architecture and Orc6 recruitment in ScORC and DmORC. In contrast to Drosophila, yeast Orc6 also interacts with Orc2 (Sun et al., 2012). The MGS mutation in the C-terminal domain of Orc6 impedes binding of Orc6 to the Orc3 insertion and recruitment of Orc6 into metazoan ORC, likely resulting in reduced origin licensing. Numbers indicate the respective ORC subunit, ‘Ins’ the Orc3 insertion, and ‘IIB’ the TFIIB domains of Orc6.
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
A Meier-Gorlin syndrome mutation in a conserved C-terminal helix of Orc6 impedes origin recognition complex formation
eLife 2:e00882.
https://doi.org/10.7554/eLife.00882




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}