Shugoshin biases chromosomes for biorientation through condensin recruitment to the pericentromere
- University of Edinburgh, United Kingdom
- Institute of Bioanalytics, Technische Universität Berlin, Germany
Figures
Figure 1 with 1 supplement
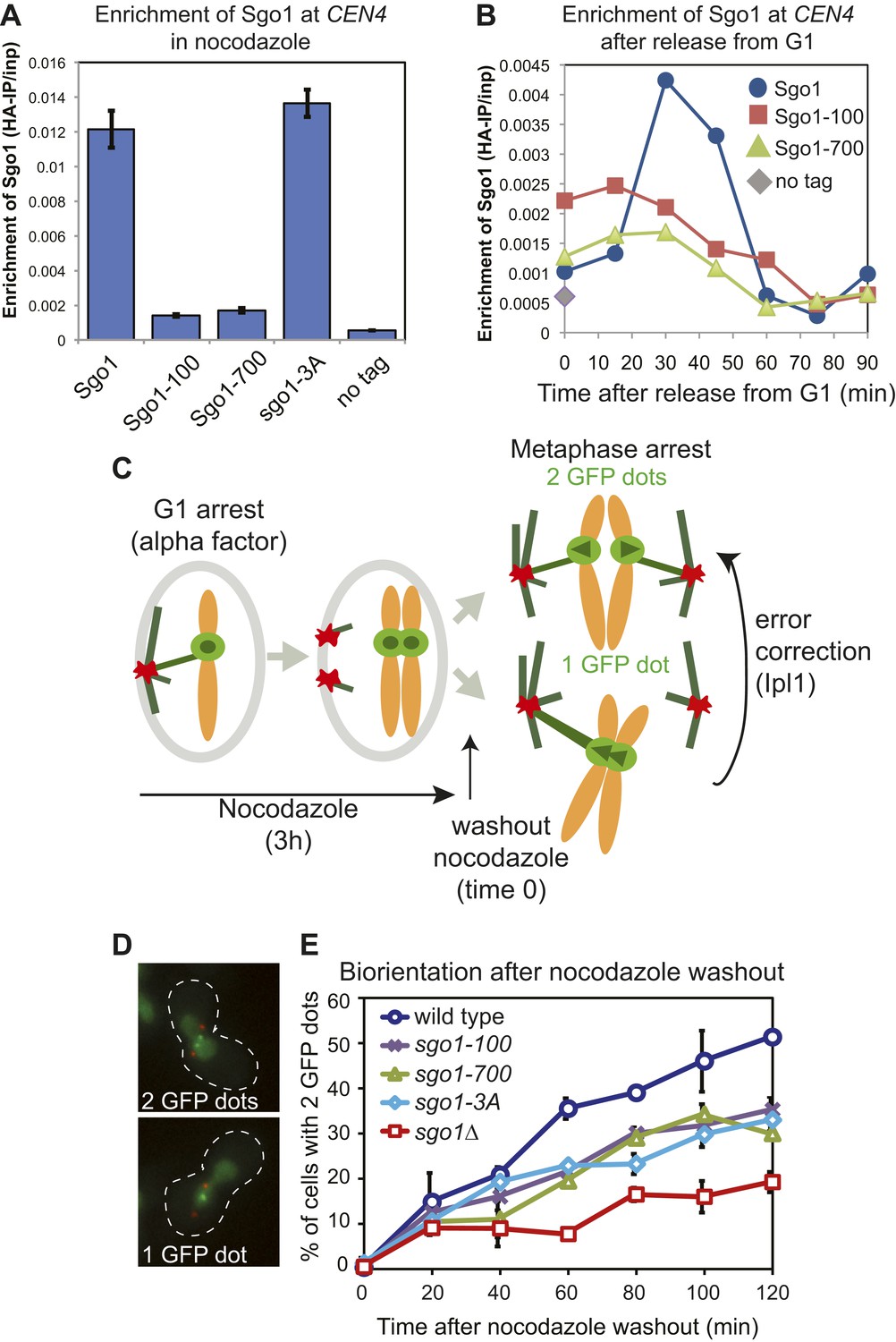
Sgo1 alleles that affect biorientation.
(A) Sgo1-3A, but not Sgo1-100 or Sgo1-700 are maintained at the centromere in cells arrested in mitosis by treatment with nocodazole. Cells carrying SGO1-6HA (AM906), SGO1-100-6HA (AM6956), SGO1-700-6HA (AM6957), SGO1-3A-6HA (AM10011) and a no tag control (AM1176) were arrested in mitosis by treating with nocodozole for 3 hr. Cells were harvested for anti-HA ChIP and the levels of each Sgo1 protein at CEN4 were analyzed by qPCR. The mean of three independent experiments is shown with error bars representing standard error. (B) Sgo1-100 and Sgo1-700 proteins are initially recruited to centromeres but fail to be maintained there. Strains as in (A) were arrested in G1 by treatment with alpha factor. Samples were extracted for analysis by anti-HA ChIP at 15 min intervals after release from G1. The levels of Sgo1-6HA at CEN4 at the indicated times after release from G1 are shown for a representative experiment. (C–E) SGO1 mutants are impaired in biorientation. Wild-type (AM4643), sgo1-100 (AM8924), sgo1-700 (AM8925), sgo1-3A (AM8923) and sgo1Δ (AM6117) cells carrying SPB (Spc42-tdTomato) and CEN4 (CEN4-GFP) markers were released from a G1 arrest into medium containing nocodazole (to depolymerize microtubules) and methionine (to deplete CDC20). After 3 hr, nocodazole was washed out, and the number of GFP dots was scored in the metaphase-arrested cells as shown in the schematic diagram (C). (D) Representative images of cells with one and two GFP dots are shown. (E) The percentage of visibly separated centromeres was determined at the indicated intervals after nocodazole washout (t = 0). Error bars indicate range (n = 2).
Figure 1—figure supplement 1

The sgo1-100 and sgo1-700 mutations do not affect the timing of cell cycle entry.
Strains as in Figure 1A were released from G1 as described in Figure 1B and DNA content was measured at the indicated times by FACS.
Figure 2
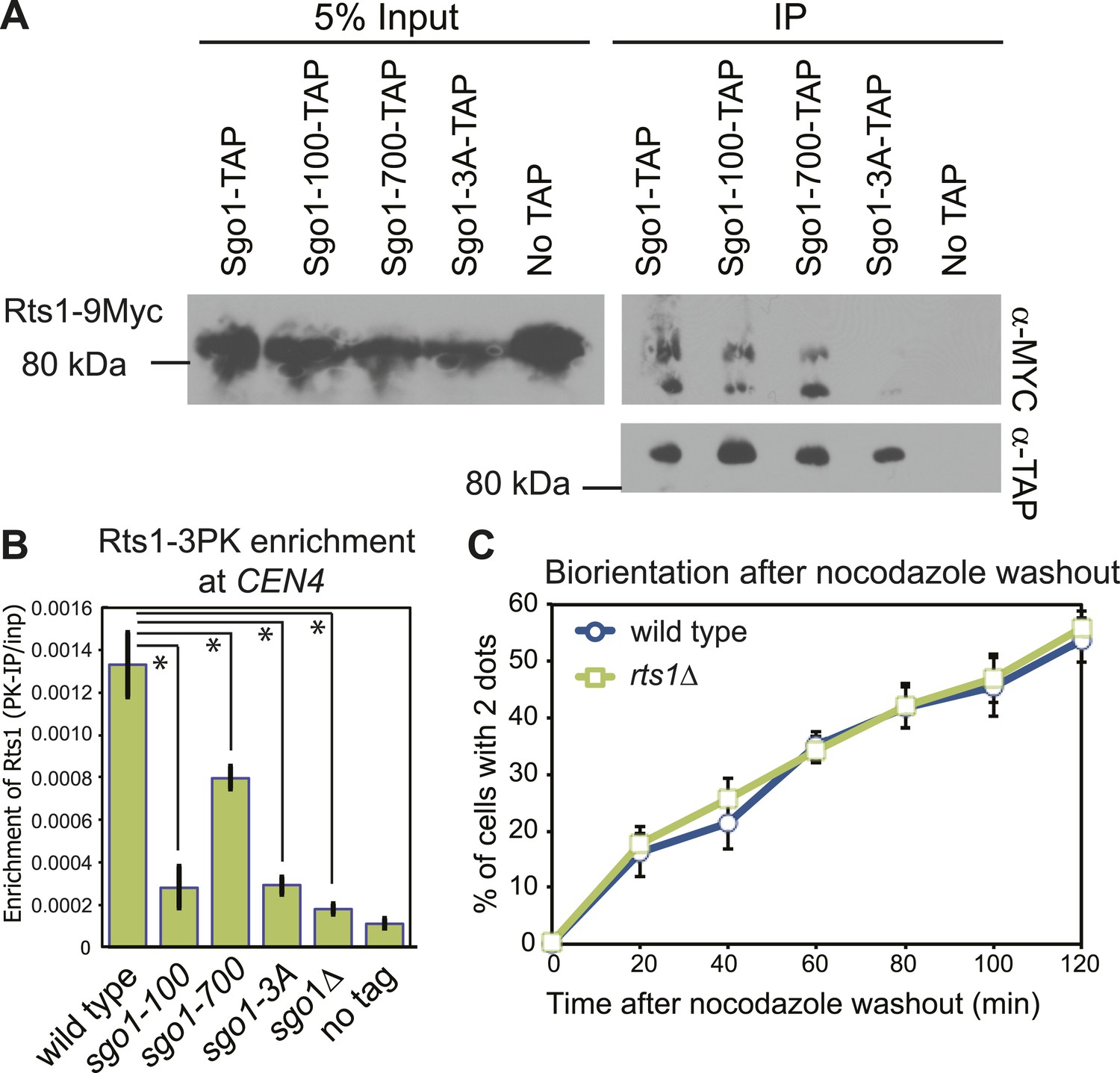
PP2A-Rts1 recruitment to the centromere by Sgo1 is not required for biorientation.
(A) Sgo1-100 and Sgo1-700, but not Sgo1-3A, associate with Rts1. Cells carrying RTS1-9MYC and SGO1-SZZ(TAP) (AM9144), sgo1-100-SZZ(TAP) (AM9272), sgo1-700-SZZ(TAP) (AM9142), sgo1-3A-SZZ(TAP) (AM9145) or no TAP (AM4721) were arrested in nocodazole for 2 hr and treated with the cross-linking reagent dithiobis(succunimidylpropionate) (DSP) before extract preparation as described in ‘Materials and methods’. Extracts were incubated with IgG-coupled beads and immunoprecipitates analyzed with the indicated antibodies. (B) Sgo1 mutants affect the centromeric localization of Rts1. Wild-type (AM8895), sgo1-100 (AM9439), sgo1-700 (AM9323), sgo1-3A (AM9293) and sgo1Δ (AM9624) cells carrying RTS1-3PK, as well as a no tag control (AM1176), were treated with nocodazole for 3 hr before harvesting for anti-PK ChIP. The mean level of Rts1-3PK enrichment at CEN4 from three experimental repeats, determined by qPCR, is shown with bars indicating standard error (*p<0.05, paired t test). (C) Sister kinetochore biorientation after microtubule depolymerization was measured in wild-type (AM4643) and rts1Δ (AM5823) cells as in Figure 1 (C). The mean of three experimental repeats with error bars representing standard deviation are shown.
Figure 3 with 2 supplements
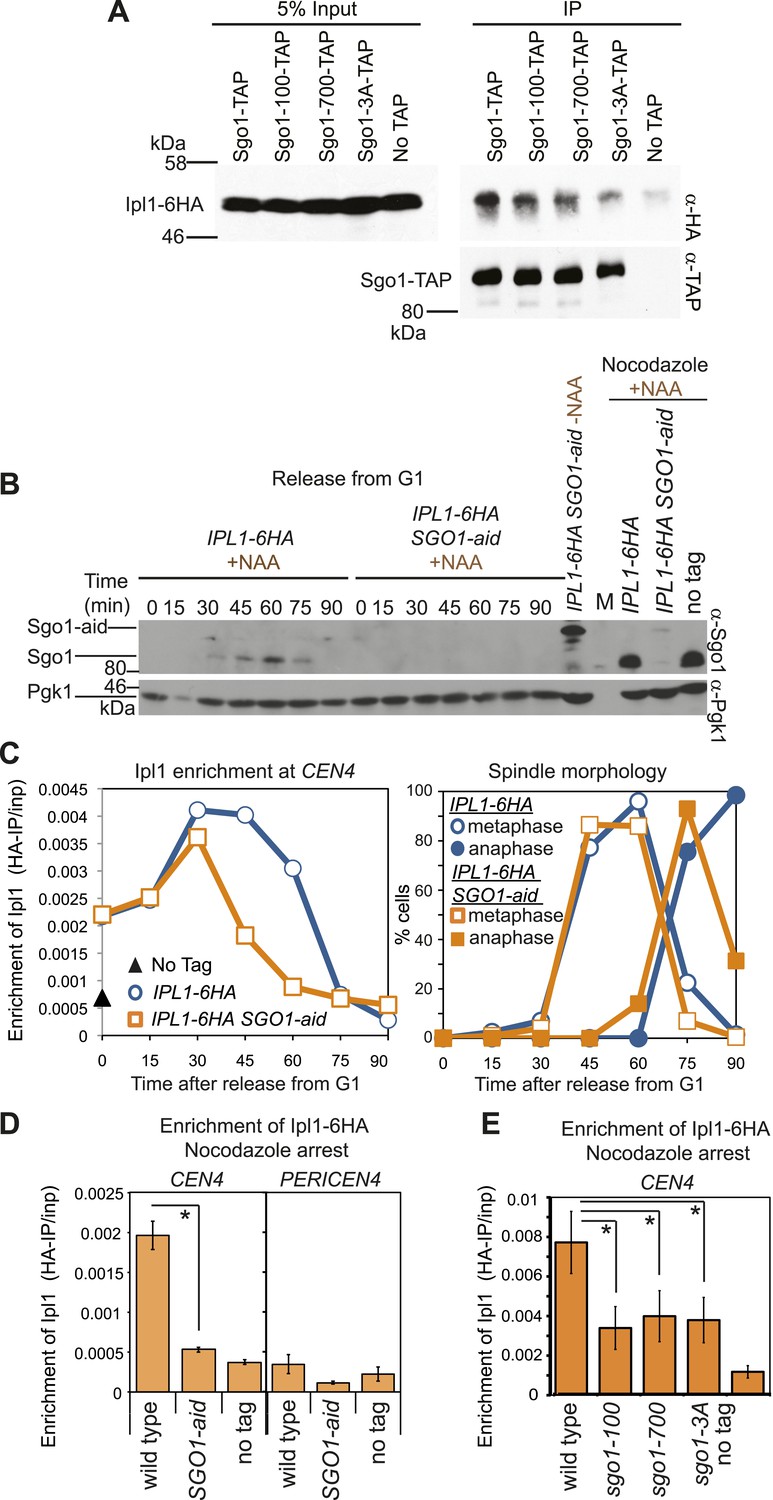
Sgo1 is required for the maintenance of Ipl1 at centromeres, but is dispensable for its initial recruitment.
(A) Ipl1/aurora B co-immunoprecipitates with Sgo1. Cells producing SZZ(TAP)-tagged Sgo1 (AM8975), Sgo1-100 (AM8971), Sgo1-700 (AM8969), Sgo1-3A (AM8973) or no TAP (AM3513) and carrying IPL1-6HA were treated with nocodazole for 2 hr before cross-linking with DSP. Extracts were prepared as described in ‘Materials and methods’, incubated with IgG-coupled beads and immunoprecipitates were analyzed by immunoblot using the indicated antibodies. (B) Degradation of Sgo1 using the auxin-inducible degron system. Representative anti-Sgo1 immunoblot for the experiments in (C and D) showing that NAA treatment leads to Sgo1 degradation. Anti-Pgk1 immunoblot is shown as a loading control. See below for experimental conditions. (C) Ipl1 is initially recruited to centromeres in the absence of Sgo1. Wild-type (AM3513) and SGO1-aid (AM9619) cells carrying IPL1-6HA were released from a G1 block in the presence of auxin (NAA) and samples harvested at 15 min intervals for measurement of Ipl1-6HA levels by anti-HA ChIP-qPCR. Also shown is a G1 sample from cells lacking IPL1-6HA (AM1176; no tag). The percentages of metaphase and anaphase spindles after anti-tubulin immunoflurescence were scored as a marker of cell cycle progression and anti-Sgo1 immunoblot confirmed Sgo1-aid degradation (shown in B). A representative experiment is shown from a total of three repeats. (D) Wild-type (AM3513) and SGO1-aid (AM9619) cells carrying IPL1-6HA together with a no tag control were arrested in G1 by alpha factor treatment and then released into medium containing NAA and nocodazole for 3 hr before harvesting for ChIP. Levels of Ipl1-6HA were determined at CEN4 and a pericentromeric site (PERICEN4) by qPCR and the mean of three experimental repeats is shown with bars representing standard error (*p<0.05, paired t test). (E) Ipl1-6HA levels at CEN4 measured by anti-HA ChIP-qPCR in wild type (AM3513), sgo1-100 (AM9090), sgo1-700 (AM9082) and sgo1-3A (AM9076) after treating directly with nocodazole for 3 hr are shown, together with a no tag (AM1176) control, treated in the same way. The mean of three independent repeats is shown with bars representing standard error (*p<0.05, paired t test). Note that levels of Ipl1-6HA at CEN4 were consistently higher in experiments where cells were directly treated with nocodazole, compared to those treated upon release from G1 (compare E with D). Presumably those cells in the population that are already in mitosis upon nocodazole addition experience an extended arrest during which Ipl1 is continually recruited.
Figure 3—figure supplement 1
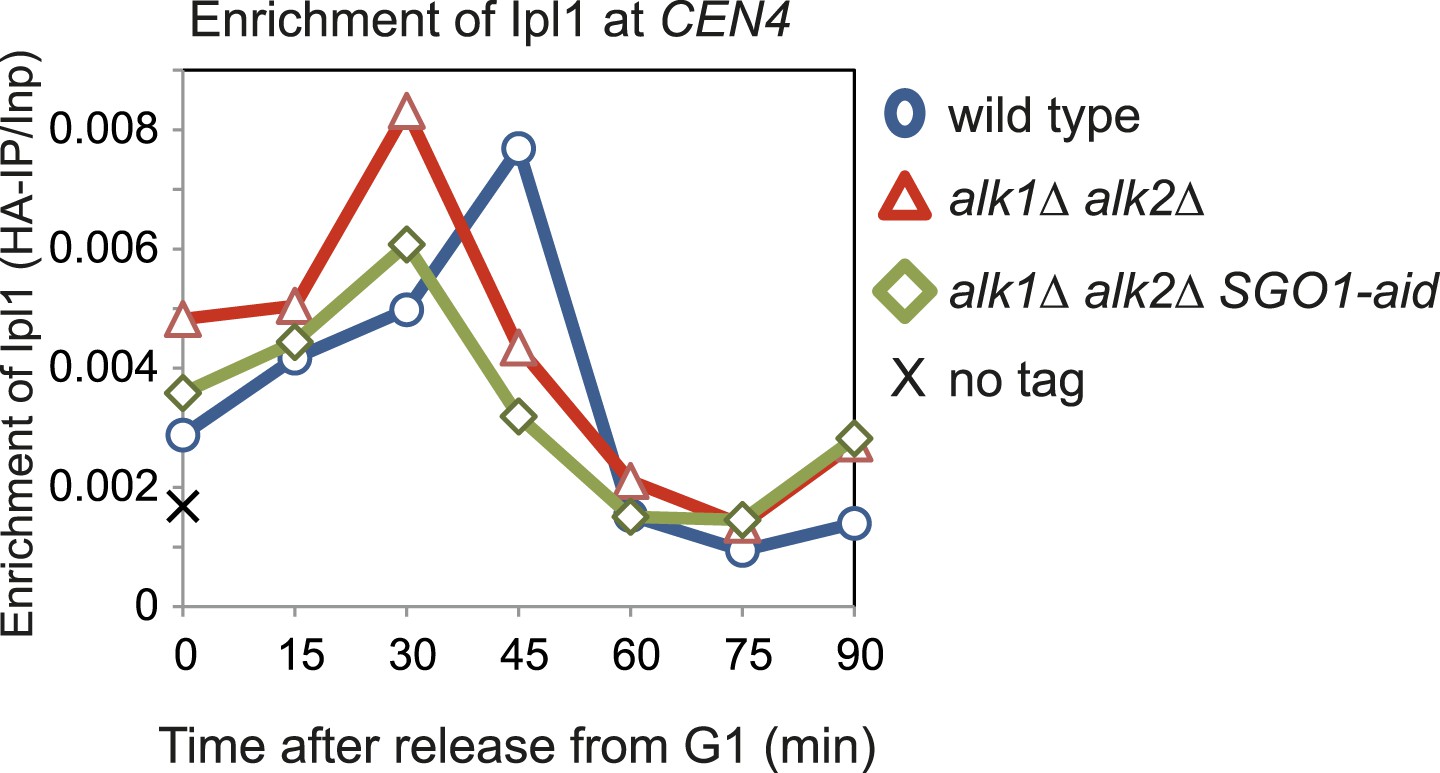
The Haspin homologs, Alk1 and Alk2, are not required for the initial recruitment of Ipl1 to centromeres.
Wild-type (AM3513), alk1Δ alk2Δ (AM10612) and alk1Δ alk2Δ SGO1-aid (AM10393) cells carrying IPl1-6HA as well as a no tag control (AM1176) were arrested in G1 by treatment with alpha factor. Samples were extracted for anti-HA ChIP at the indicated intervals after release from G1. The levels of Ipl1-6HA at CEN4 were measured by ChIP-qPCR.
Figure 3—figure supplement 2

Defective biorientation in ipl1-as mutants.
Biorientation assay showing CEN4-GFP separation in wild-type (AM4643) and ipl1-as (AM10374) cells carrying SPB (SPC42-tdTomato) markers. Cells were released from G1 into nocodazole and NAPP1, before nocodazole was washed out and GFP foci were scored in the metaphase-arrested cells as in Figure 1C.
Figure 4 with 3 supplements

Sgo1 interacts with condensin and recruits it to the pericentromere.
(A and B) Condensin and PP2A co-purify with Sgo1. Sgo1 was purified from wild-type or protease-deficient cells that were (A) cycling or (B) arrested in mitosis using the cold-sensitive tubulin allele tub2-401 as described in ‘Materials and methods’. Comparable strains lacking TAP were used as a control for non-specific association with the beads. All cells were treated with the cross-linker, DSP, before harvesting and preparing extracts as described in ‘Materials and methods’. Extracts were incubated with IgG-coupled beads and immunoprecipitates were visualized on silver-stained SDS-PAGE gels. The table shows the number of peptides of subunits of the PP2A and condensin complexes that were identified in the Sgo1-TAP purifications after mass spectrometry. The full list of identified proteins is given in Supplementary file 1. Strains used in (A) were AM7509 (SGO1-SZZ(TAP), AM1176 (no tag), AM8226 (protease-deficient, SGO1-SZZ(TAP) and AM8184 (protease-deficient, no tag). Strains used in (B) were AM8456 (tub2-401 SGO1-SZZ(TAP), AM2730 (tub2-401, no tag), AM8455 (protease-deficient, tub2-401 SGO1-SZZ(TAP) and AM8259 (protease-deficient, tub401 no tag. (C) A schematic diagram illustrating the composition of budding yeast condensin is shown. (D and E) Cells carrying either YCS4-6HA (AM9138) (D) or BRN1-6HA (AM9266) (E) and SGO1-SZZ(TAP) or no TAP (AM5705 and AM5708) were arrested in nocodazole for 2 hr before treating with DSP. Extracts were incubated with IgG-coupled beads and immunoprecipitates were analyzed with the indicated antibodies by immunoblot. (F–H) Sgo1 is required for Brn1 association with the pericentromere. The genome-wide localization of Brn1-6HA was determined in wild-type (AM5708) and sgo1Δ (AM8834) cells by anti-HA ChIP followed by high throughput sequencing (ChIP-Seq) after arresting in mitosis by treating with nocodazole for 3 hr. (F) Brn1 enrichment along chromosome V along with a magnification of a 50 kb region including the centromere is shown. The number of reads at each position was normalized to the total number of reads for each sample (RPM: reads per million) and shown in the Integrated Genome Viewer from the Broad Institute (Robinson et al., 2011). (G) The number of reads at coordinates corresponding to the rDNA region on chromosome XII is shown for wild-type and sgo1Δ anti-HA ChIP samples normalized to the total number of reads for each sample. Brn1 enrichment at the rDNA is similar in wild-type and sgo1Δ cells. (H) Brn1 enrichment in a 50 kb domain surrounding all 16 budding yeast centromeres is shown for wild-type and sgo1Δ cells. For both wild type and sgo1Δ, the ratio of the local maximum in a 100 bp window for ChIP sample/input is calculated at the indicated distance from the centromere for all 16 chromosomes. Box plot of maximum count value for 100 bp windows for 25 kb on both sides of each centromere is shown to give a composite view of all 16 pericentromeres. (I and J) Recruitment of Brn1 to centromeres occurs coincident with, and is dependent on, Sgo1. Wild-type cells carrying SGO1-9MYC and BRN1-6HA (AM9622) as well as sgo1Δ cells (AM8834) carrying BRN1-6HA were arrested in G1 using alpha factor. Samples were extracted at 15 min intervals after release from G1 for anti-HA and anti-Myc ChIP and tubulin immunofluorescence. (I) The levels of Brn1-6HA and Sgo1-9Myc at CEN4 were measured at the indicated timepoints by anti-HA and anti-Myc ChIP-qPCR, respectively. Also shown is a G1 sample from cells lacking BRN1-6HA (no tag; AM1176). (J) The percentages of metaphase and anaphase spindles after anti-tubulin immunofluorescence were scored as a marker of cell cycle progression. Shown is a representative experiment from three repeats. (K) Schematic diagram illustrating the protein complexes (PP2A, condensin, CPC) recruited to the pericentromere by shugoshin.
Figure 4—figure supplement 1

The Sgo1-condensin interaction is not dependent on DNA or treatment with the cross-linking agent, DSP.
Cells carrying YCS4-6HA and SGO1-SZZ(TAP) (AM9138) or no TAP (AM5705) were arrested in mitosis by treatment with nocodazole for 2 hr. Cultures were either harvested and directly drop-frozen (−DSP) or treated with DSP prior to drop freezing (+DSP) as described in ‘Materials and methods’. Extracts were either treated with 25 U of the DNA degrading agent, benzonase (+benzonase) and rotated at room temperature or held on ice (−benzonase) for 30 min. All samples were incubated with IgG-coupled beads and analyzed by immunoblot with the indicated antibodies.
Figure 4—figure supplement 2
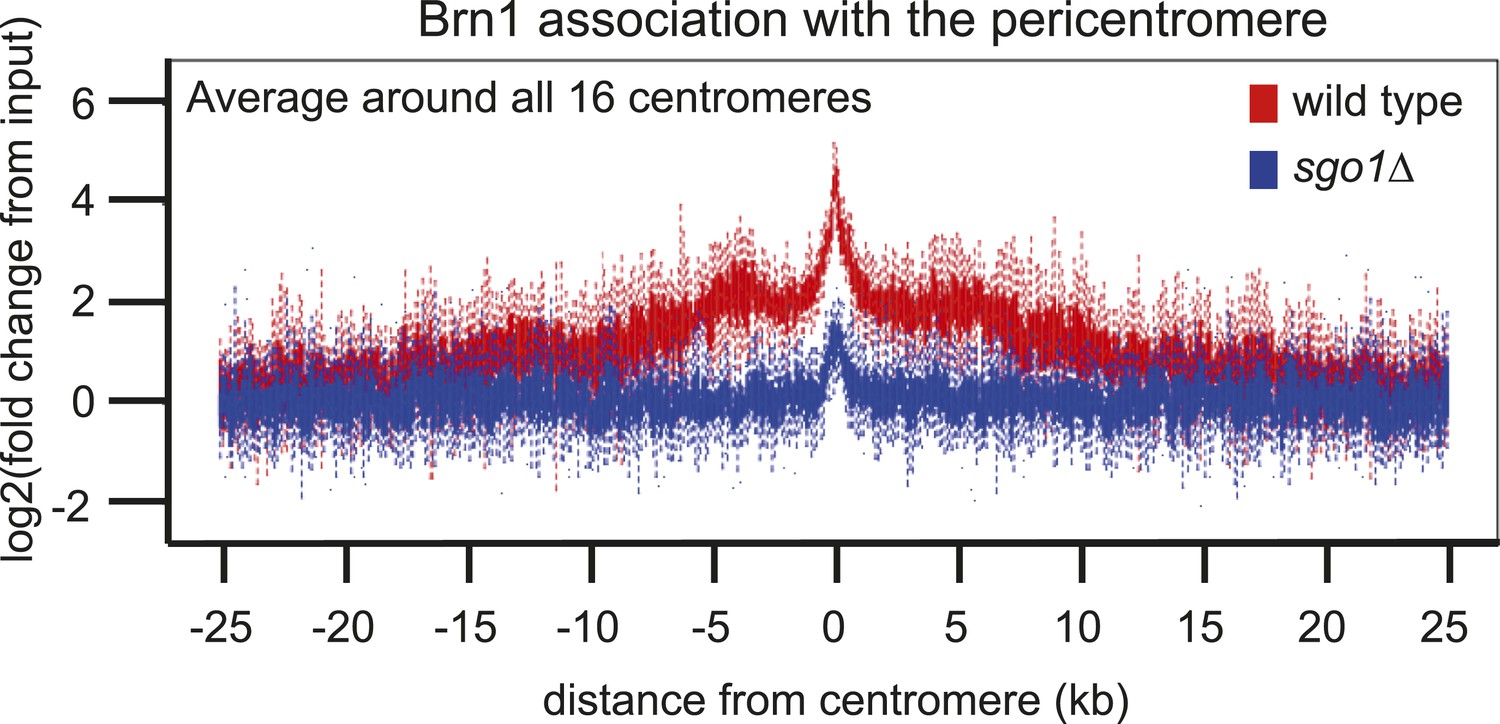
Removal of PCR duplicates does not alter the conclusion that Sgo1 is important for Brn1 enrichment in the pericentromere.
This is the same analysis as in Figure 3H, except that only unique reads are included (therefore eliminating duplicate samples generated during the PCR amplification step). Both methods of analysis lead to the conclusion that Brn1 levels in the pericentromere are greatly reduced in sgo1Δ cells. Note that the centromeric peak may be an artifact as centromeric sequences were over-represented in the sgo1Δ sample compared to wild type (where they were under-represented compared to the rest of the genome). Potentially, altered pericentromeric structure in sgo1Δ cells could enable more efficient recovery of these sequences during the purification procedure.
Figure 4—figure supplement 3
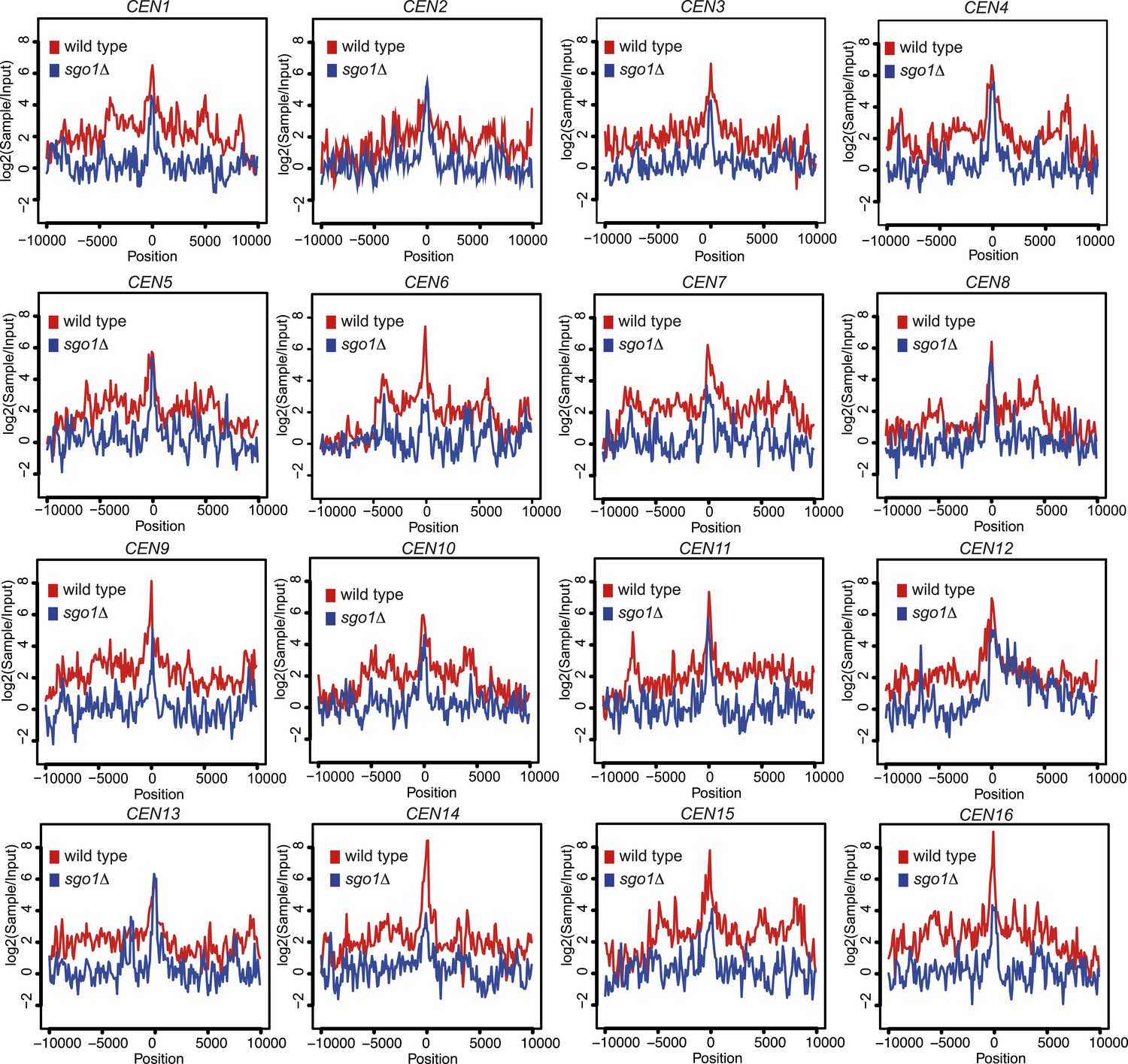
Brn1 is reduced around all 16 individual centromeres in sgo1Δ cells.
Brn1 enrichment in a 20 kb region surrounding all 16 individual centromeres in wild type and sgo1Δ cells is shown for the experiment described in Figure 4F–H. All reads were included in this analysis.
Figure 5 with 4 supplements
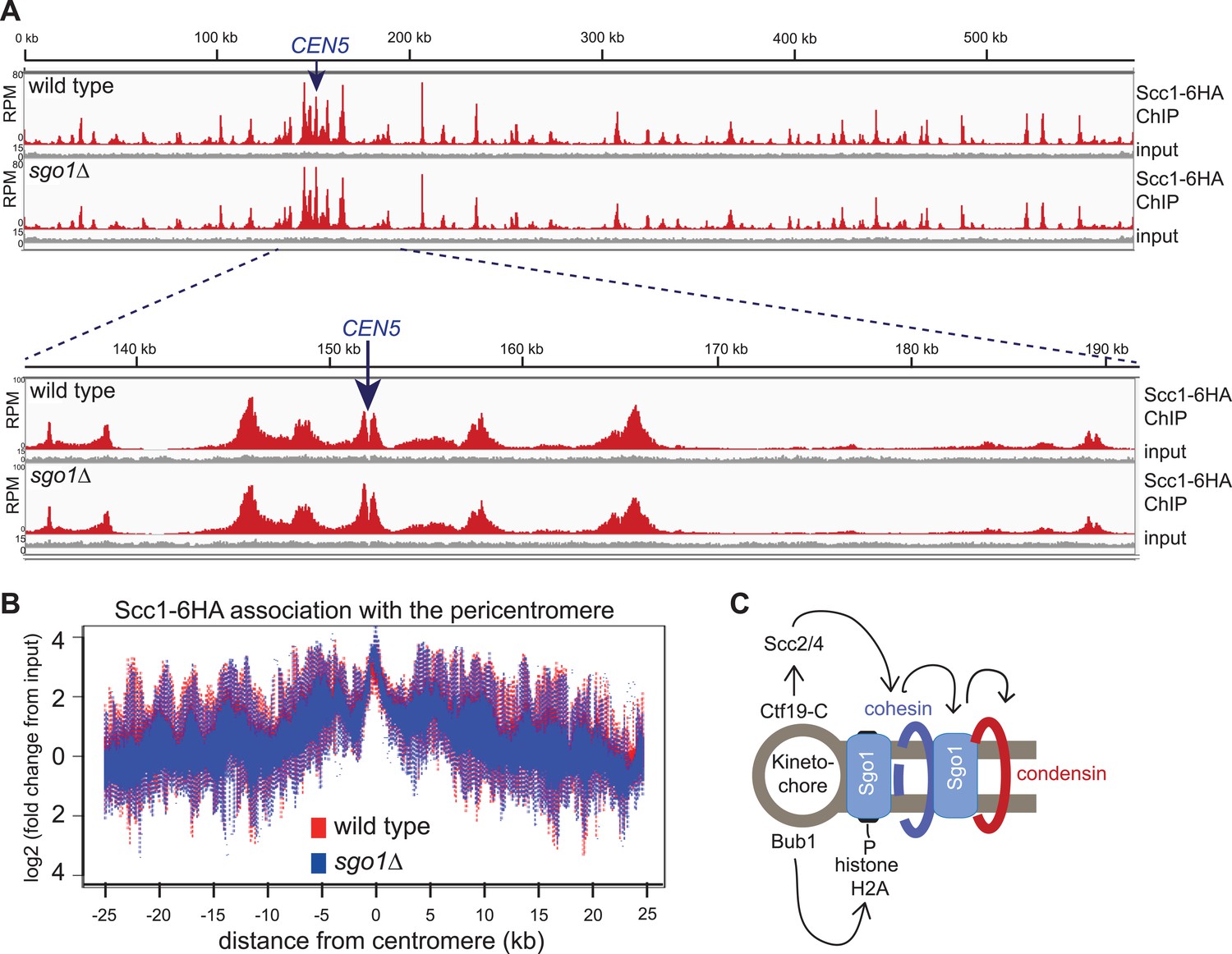
Sgo1 is not required for cohesin association with chromosomes.
Wild-type (AM1145) and sgo1Δ (AM1474) cells carrying SCC1-6HA were arrested in mitosis by treatment with nocodazole for 3 hr. Samples were harvested, anti-HA ChIP was performed and both input and IP samples were sequenced for both strains. (A) Scc1-6HA enrichment along chromosome V along with a magnification of a 50 kb region including the centromere is shown. The number of reads at each position were normalized to the total number of reads for each sample and displayed using the Integrated Genome Viewer from the Broad Institute (Robinson et al., 2011). (B) Box plot of maximum count value for 100 bp windows for 25 kb on both sides of each centromere is shown to give a composite view of all 16 pericentromeres. All reads are included. (C) Schematic diagram indicating hierarchy of factors required for condensin association with the pericentromere.
Figure 5—figure supplement 1
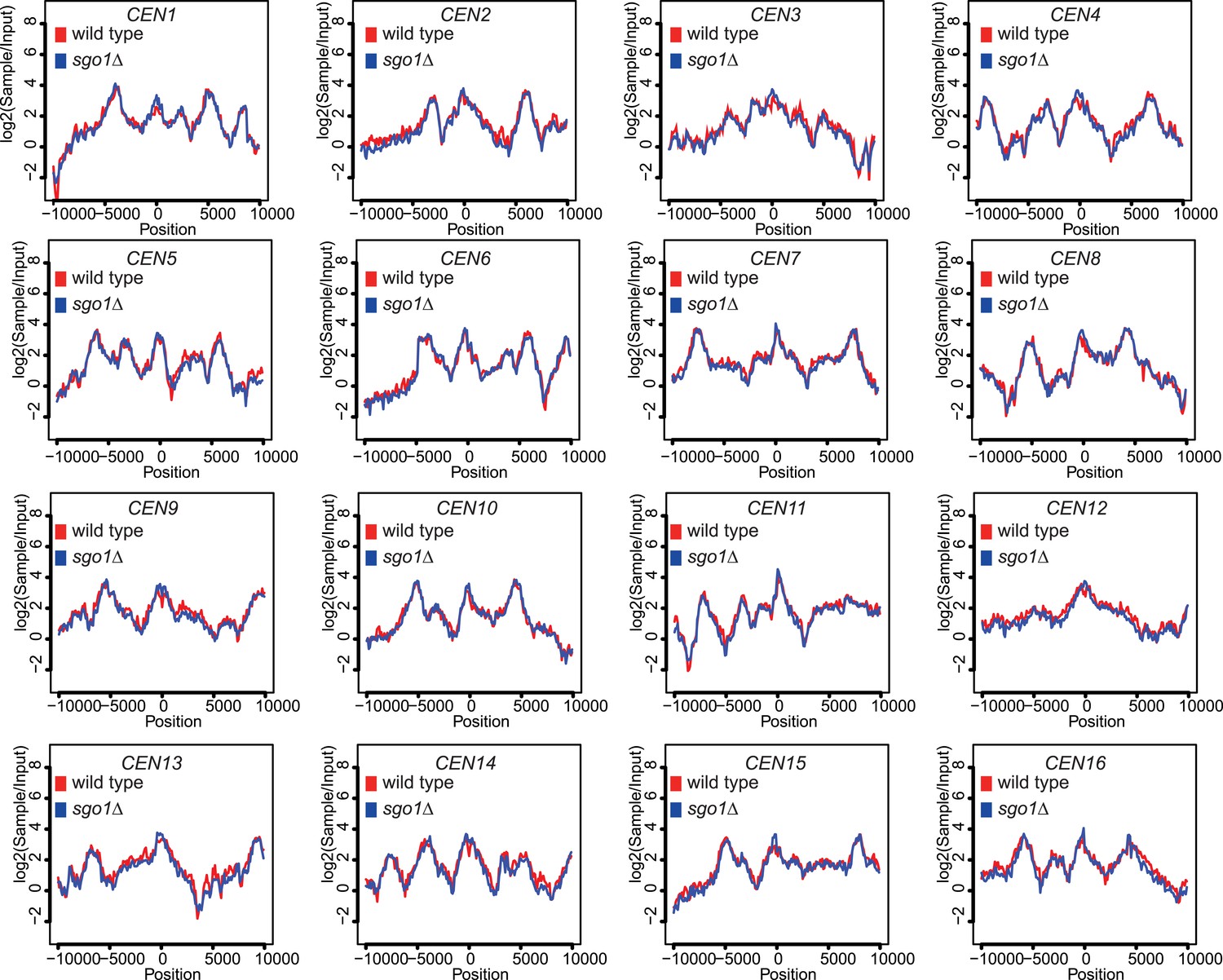
Scc1 association with all 16 centromeres is unaffected by SGO1 deletion.
Scc1 enrichment in a 20 kb region surrounding all 16 individual centromeres in wild type and sgo1Δ cells is shown from the experiment in Figure 5. All reads were included in this analysis.
Figure 5—figure supplement 2
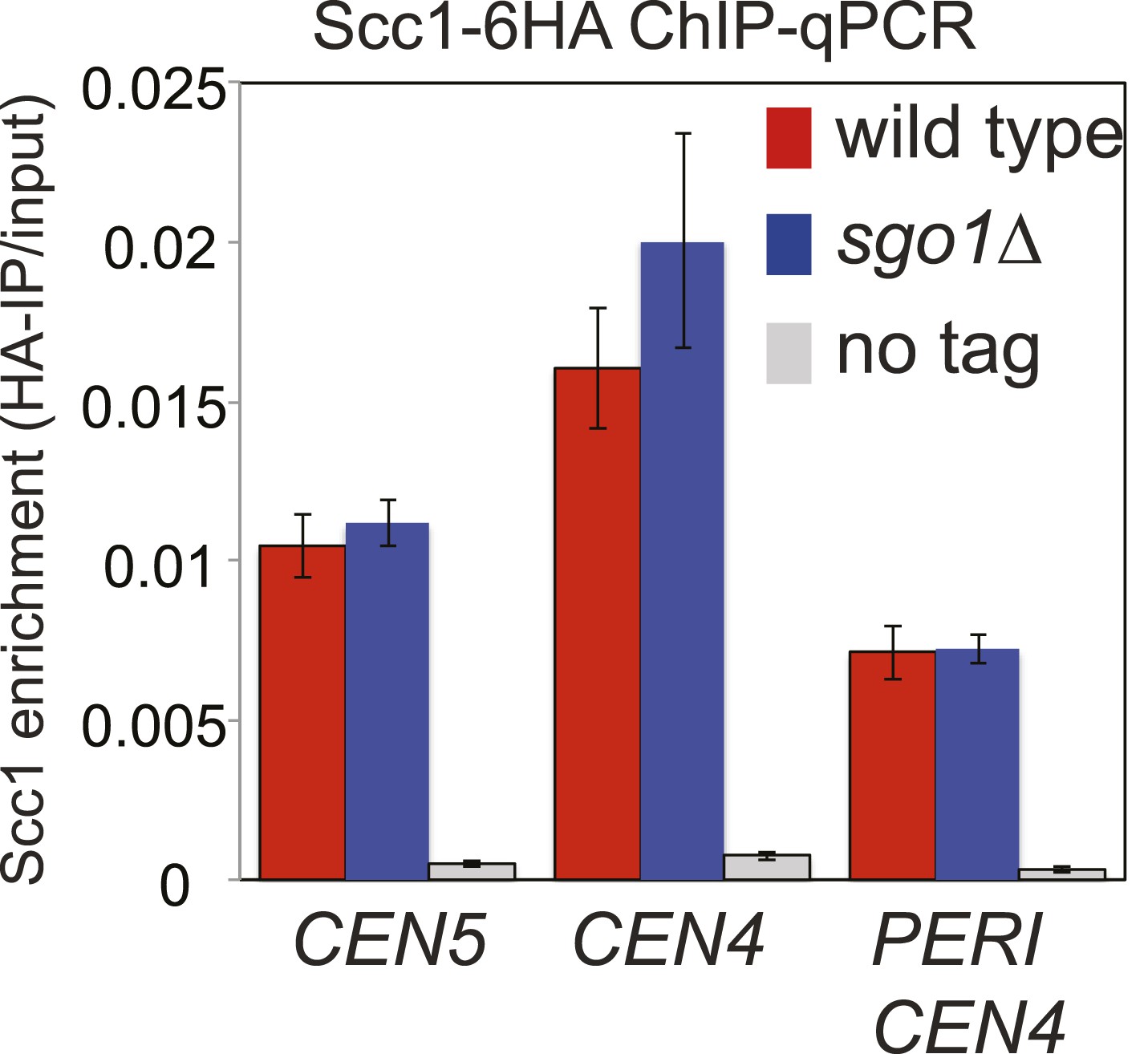
ChIP-qPCR analysis showing Scc1-6HA levels at the centromere and pericentromere.
Wild type and sgo1Δ cells were treated as described in Figure 5 except that ChIP samples were analyzed by qPCR using primer sets at CEN4, CEN5 and a site in the pericentromere of chromosome IV. The mean of three independent experiments is shown with bars indicating standard error.
Figure 5—figure supplement 3
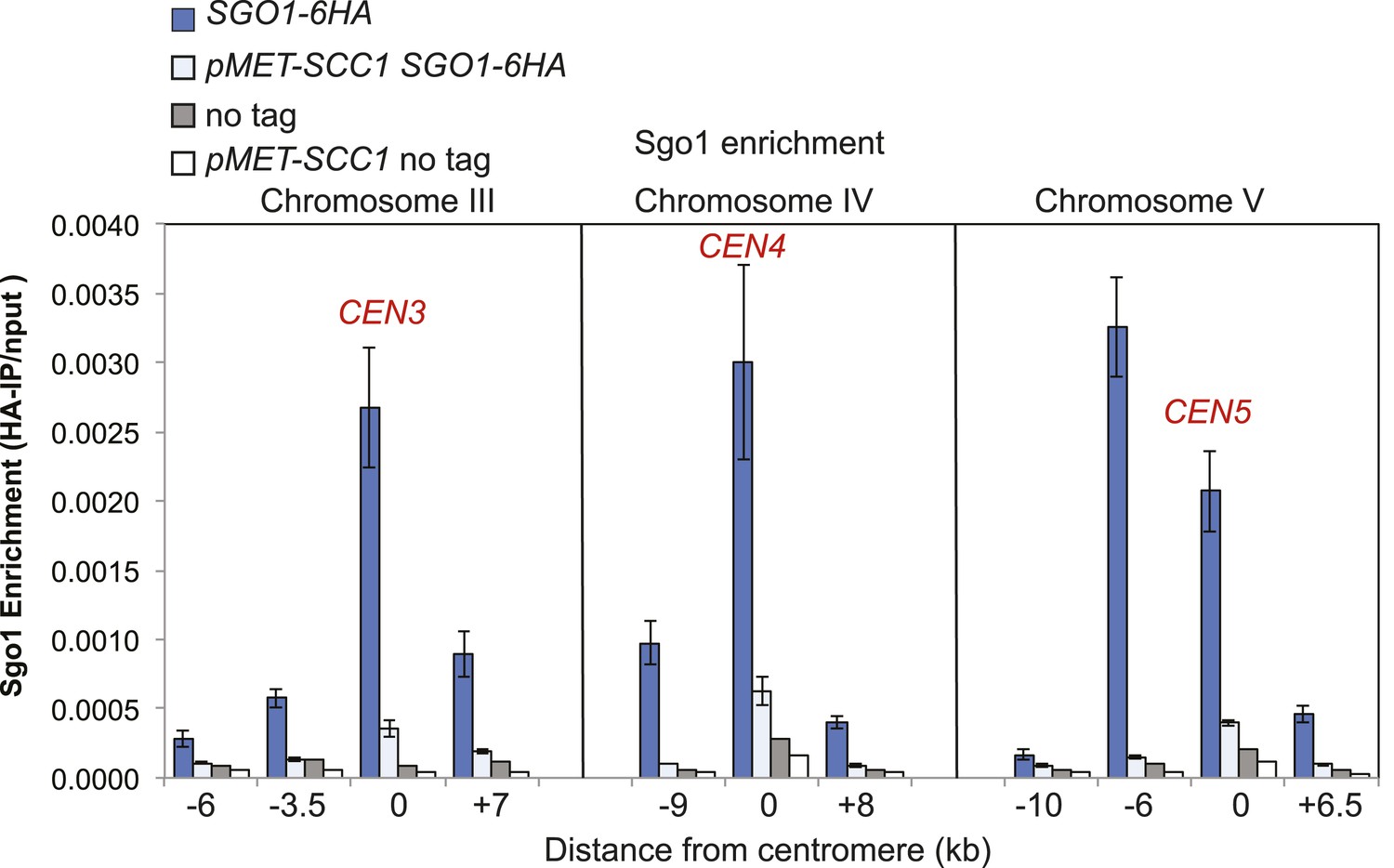
Cohesin is required for normal Sgo1 association with the pericentromere.
Wild type (AM906) and pMET-SCC1 (AM6673) strains carrying SGO1-6HA together with no tag wild type (AM1176) and pMET-SCC1 controls (AM1599) were arrested in G1 using alpha factor in the presence of methionine (to deplete Scc1). Strains were released into medium containing nocodazole and methionine for 3 hr and levels of Sgo1-6HA at the indicated sites were measured by ChIP-qPCR.
Figure 5—figure supplement 4
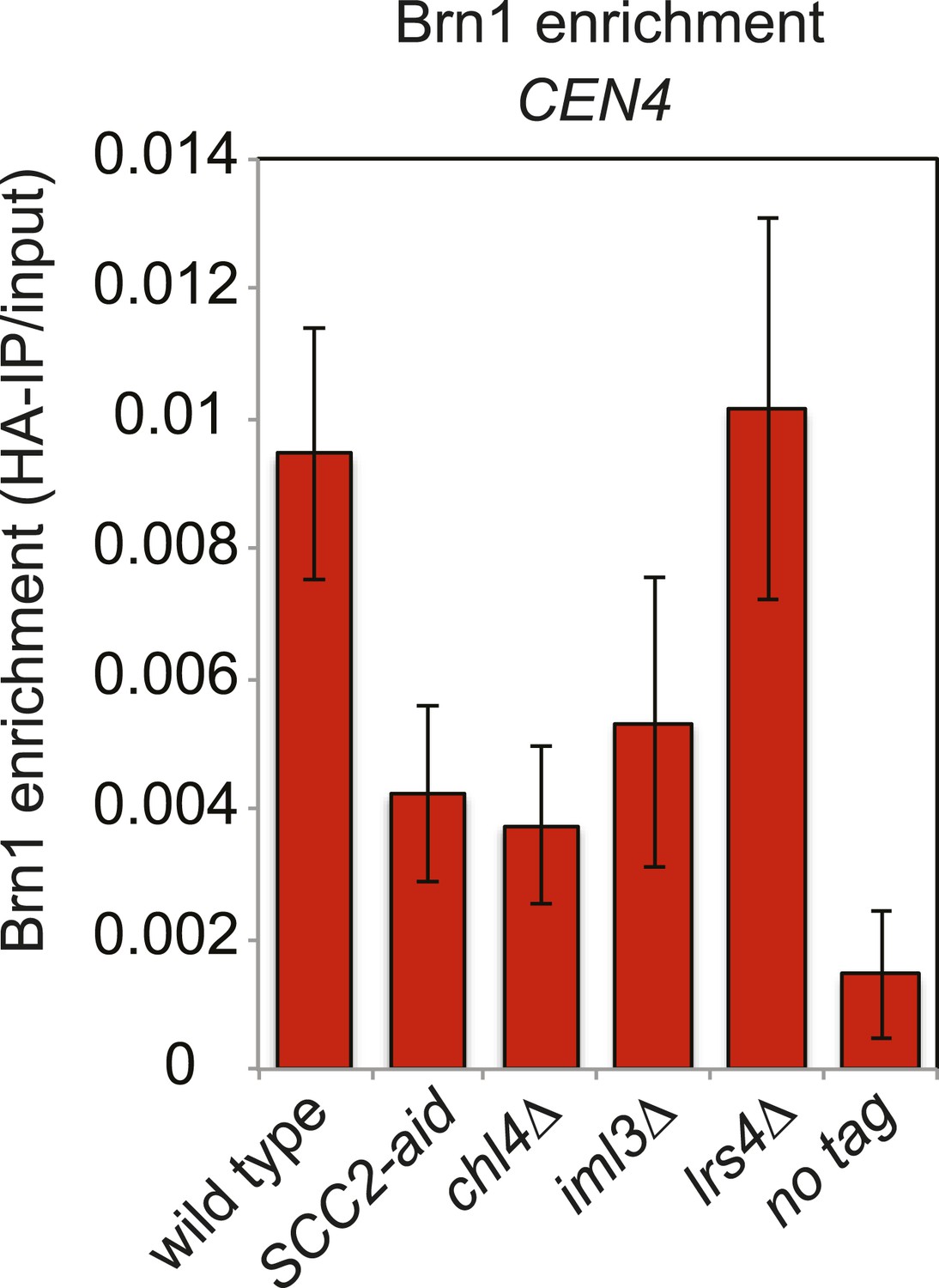
Cohesin loading factors, but not monopolin, are important for proper condensin association with the centromere.
Wild type (AM5708), SCC2-aid (AM8918), chl4Δ (AM8885), iml3Δ (AM5710) and lrs4Δ (AM9766) strains carrying BRN1-6HA, as well as a no tag control (AM1176) were treated with nocodazole and NAA (to degrade Scc2-aid) for 3 hr before harvesting for ChIP and measuring the levels of Brn1-6HA at CEN4 by ChIP-qPCR.
Figure 6 with 1 supplement
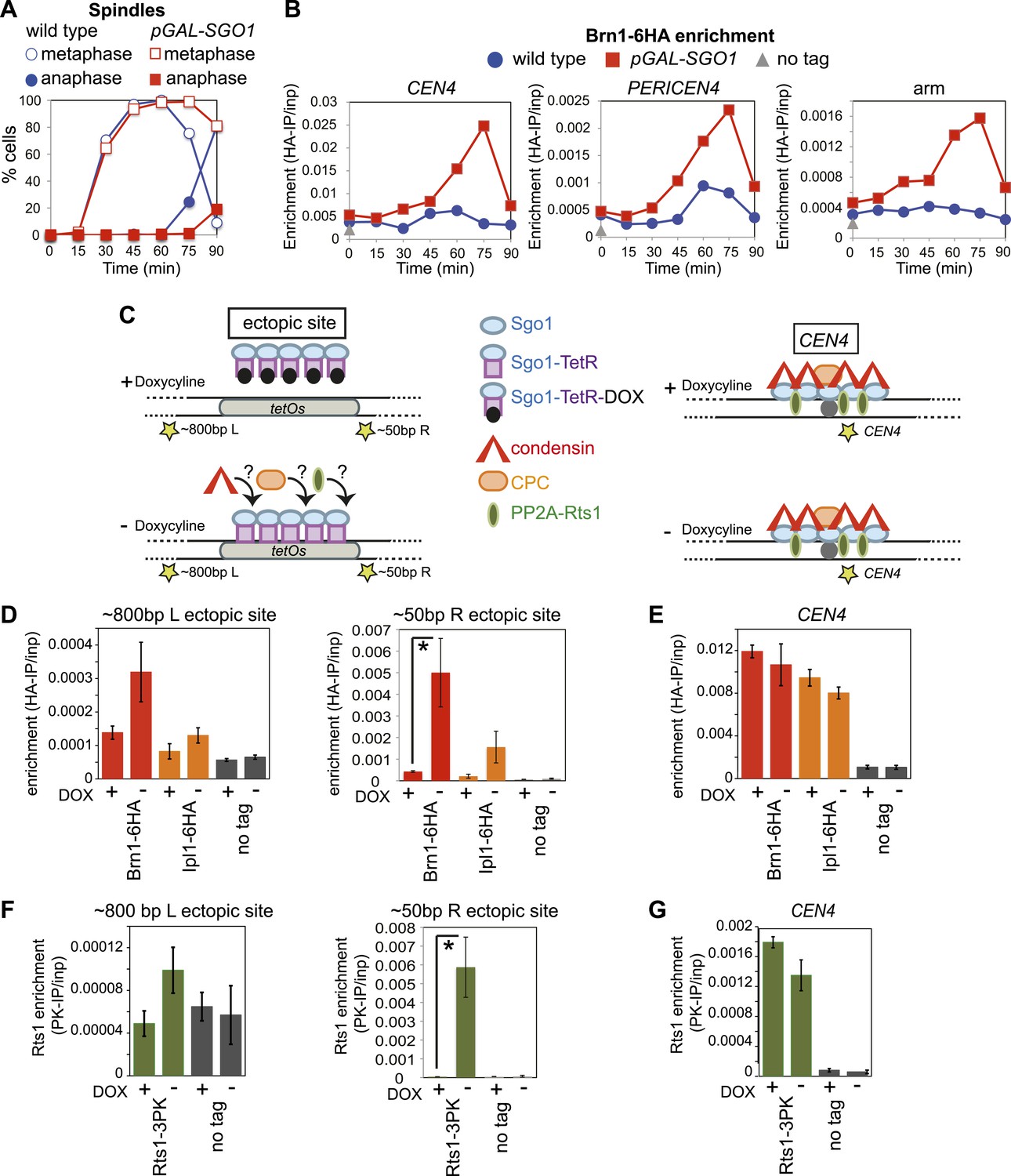
Sgo1 is sufficient for condensin recruitment.
(A and B) Sgo1 overproduction leads to increased levels of Brn1 on chromosomes. Cells carrying BRN1-6HA and that were otherwise wild type (AM5708) or carrying pGAL-SGO1 (AM10859) integrated an ectopic locus, were arrested in G1 in rich medium containing raffinose and adenine using alpha factor (YEP + R + A). After 2 hr 30 min, galactose (2%) was added to induce SGO1 overexpression and 30 min later, cells were released from G1. Samples were collected at the indicated times after release from G1 for analysis of cell cycle progression by scoring spindle morphology after anti-tubulin immunofluorescence (A) or for measurement of Brn1 levels by anti-HA ChIP-qPCR (B). Sites analyzed were at CEN4, a pericentromeric site or a chromosomal arm site on chromosome IV. A representative experiment from a total of three independent repeats is shown. (C–G) Tethered Sgo1 at an ectopic site recruits Brn1, Rts1 and Ipl1. (C) Schematic diagram showing the expected effects of doxycycline at the ectopic site and at CEN4, as well as the locations of primer sets used for qPCR (yellow stars). Primer sets used were ∼800 bp left of the tethering site, ∼50 bp right of the tethering site and at CEN4. (D–G) Strains carrying Sgo1-TetR-GFP and tetOs integrated at the HIS3 locus were arrested in nocodazole for 3 hr either in the presence (+DOX) or absence (−DOX) of doxycycline and harvested for ChIP-qPCR. (D and E) Anti-HA ChIP was performed on SGO1-TetR-GFP HIS3::tetOs strains carrying either BRN1-6HA (AM9847), IPL1-6HA (AM9940) or no tag (AM9655) and levels of Brn1-6HA and Ipl1-6HA were measured by qPCR at the indicated sites adjacent to the ectopic site (D) or at CEN4 (E). (F and G) Anti-PK ChIP was performed on SGO1-TetR-GFP HIS3::tetOs strains carrying Rts1-3PK (AM9783) or no tag (AM9655) and levels of Rts1-3PK were measured by qPCR at the indicated sites adjacent to the ectopic site (F) or at CEN4 (G). In (D–G), the mean of three or four experimental repeats is shown with bars representing standard error (*p<0.05, unpaired t test).
Figure 6—figure supplement 1
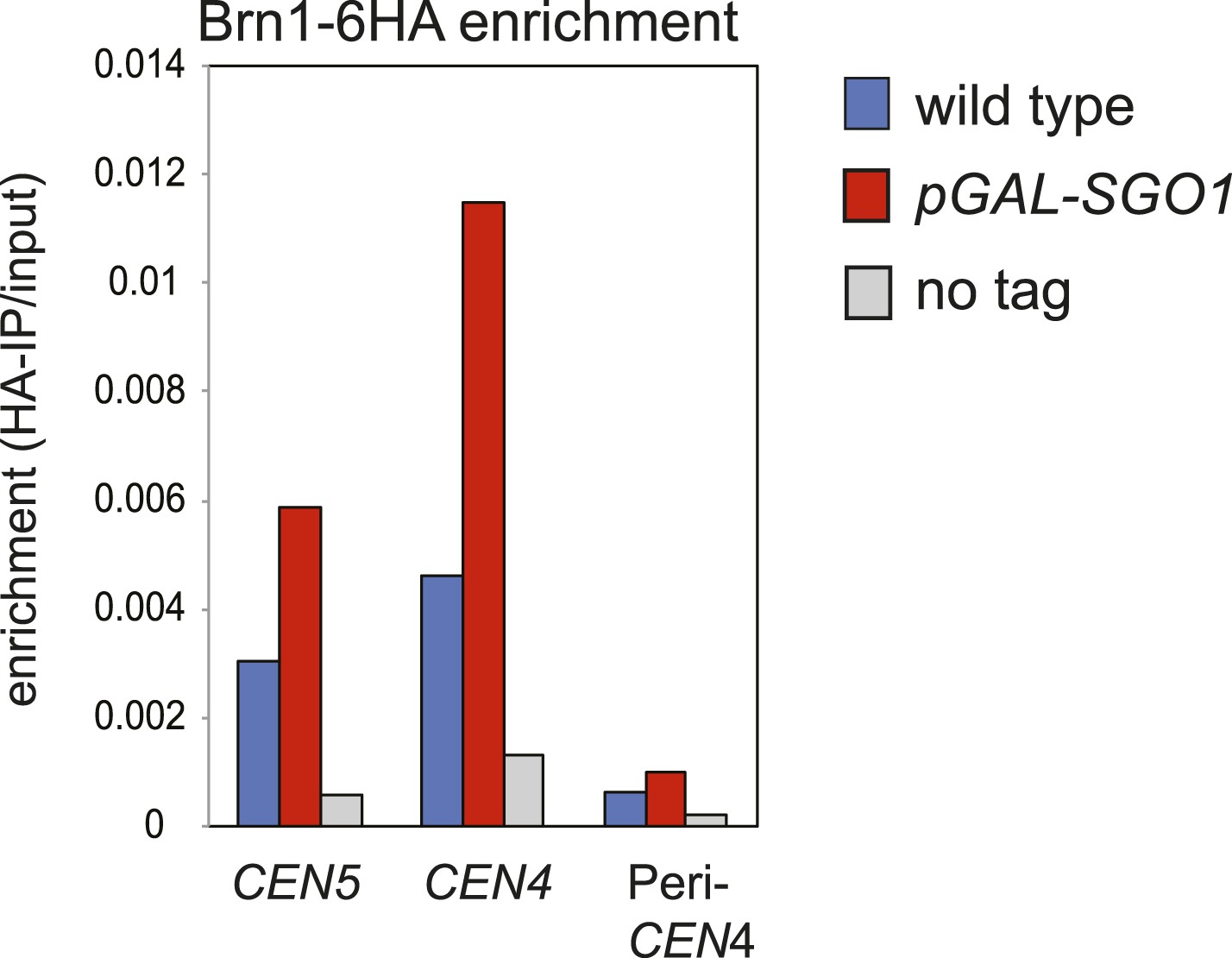
SGO1 overexpression in metaphase-arrested cells increases Brn1 association with the centromere.
Strains AM5708 (BRN1-6HA), AM10859 (BRN1-6HA pGAL-SGO1) were pre-cultured in YEP + R + Ade medium before supplementing with nocodazole and galactose (2 hr) for 3 hr and then harvesting for anti-HA ChIP. The levels of Brn1-6HA were measured at the indicated sites by qPCR.
Figure 7 with 1 supplement
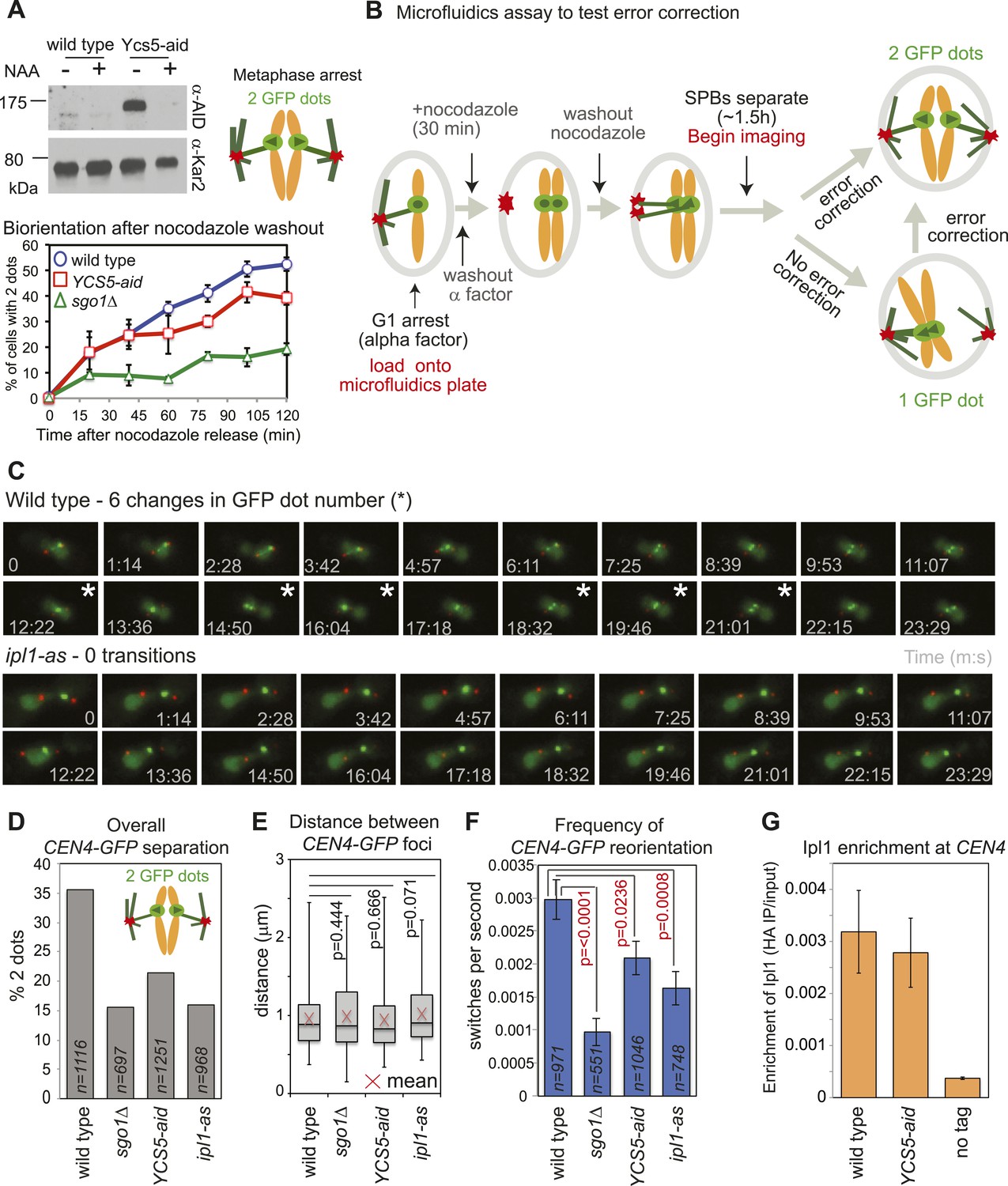
Condensin facilitates effective error correction.
(A) Sister kinetochore biorientation is defective after nocodazole washout in cells lacking condensin. Strains carrying SPB (Spc42-tdTomato) and CEN4 (CEN4-GFP) markers were released from a G1 arrest into nocodazole and arrested in metaphase by CDC20 depletion. After 3 hr, nocodazole was washed out (t = 0) and CEN4-GFP separation scored at the indicated intervals as shown in Figure 1C. Error bars represent standard deviation (wild type and YCS5-aid; n = 3) or range (sgo1Δ; n = 2; reproduced from Figure 1E). A representative anti-aid immunoblot is shown to confirm Ycs5 degradation upon NAA addition. Samples were taken in G1 (−NAA) and 120 min after release (+NAA). Anti-Kar2 immunoblot is shown as a loading control. (B–F) Condensin contributes to efficient error correction. (B) Scheme of the live single-cell microfluidics experiment. Wild-type (AM4643), sgo1Δ (AM6117), YCS5-aid (AM9038) and ipl1-as (AM10374) cells carrying CEN4 (CEN4-GFP) and SPB markers (SPC42-tdTomato) were released from a G1 arrest into nocodazole, NAA and NAPP1 and arrested in metaphase by CDC20 depletion. After 30 min, nocodazole was washed out. When the majority of cells had 2 SPBs (∼1 hr 30 min later), we began imaging and a total of 21 frames were grabbed at approximately 74 s intervals. (C) Representative images for wild type and ipl1-as are shown. Numbers indicate time (s) each frame was grabbed and asterisks indicate a change in GFP dot number compared to the previous frame. (D) The overall percentage of separated CEN4-GFP foci was determined for cells with two visible SPBs from all frames combined. (E) The distance between CEN4-GFP foci was measured in cells with separated foci. Box boundaries represent the upper and lower quartiles, respectively. The red cross indicates the mean, the horizontal line indicates the median and error bars show the maximum and minimum values observed. n = 396 (wild type), 108 (sgo1Δ), 267 (YCS5-aid) and 154 (ipl1-as). (F) The observed frequency of switching between one and two GFP foci was calculated for cells with 2 SPBs. A student t test was used to obtain p values. (G) Ycs5 is not required for Ipl1 association with the centromere. The levels of 6HA-tagged Ipl1 in wild-type (AM3513) and YCS5-aid (AM10334) cells, grown in the presence of NAA and nocodazole for 3 hr, were measured at CEN4 by anti-HA ChIP-qPCR and compared to a no tag control (AM1176). The mean of three independent experiments is shown with bars representing standard error. This is the same experiment as shown in Figure 3D and the wild-type data is reproduced for comparison.
Figure 7—figure supplement 1

Deletion of SGO1 impairs biorientation rather than centromere cohesion.
The distance from CEN4-GFP to the nearest SPC42-tdTomato focus was measured for cells with just one visible CEN4-GFP foci from the experiment shown in Figure 7C–F. The fraction of cells with a CEN4-GFP to SPB distance greater or less than the median value for wild type (0.865 mm) is plotted for wild type, sgo1Δ, YCS5-aid and ipl1-as cells. p values were obtained using a chi square test.
Figure 8

Condensin biases chromosomes to biorient.
(A–C) Condensin and Sgo1, but not Ipl1, are required to bias sister kinetochores towards biorientation. (A) Scheme of the microfluidics assay to test sister kinetochore bias. Wild-type, sgo1Δ, YCS5-aid and ipl1-as cells as in Figure 5B were released from a G1 arrest into NAA and NAPP1 and arrested in metaphase by CDC20 depletion. SPBs were allowed to separate for 1 hr 30 min before cells were treated with nocodazole for an additional 30 min. After 2 hr total, nocodazole was washed out and frames were grabbed at approximately 94 s intervals for a total of 21 frames. (B) The percentage of cells that separated CEN4-GFP foci at least once during the observation period is shown for the indicated strains. p values indicate significance (chi-square test). (C) Representative images of wild-type and YCS5-aid cells are shown. Time of image acquisition (s) is shown. The asterisk indicates the first time GFP foci are separated.
Figure 9 with 2 supplements
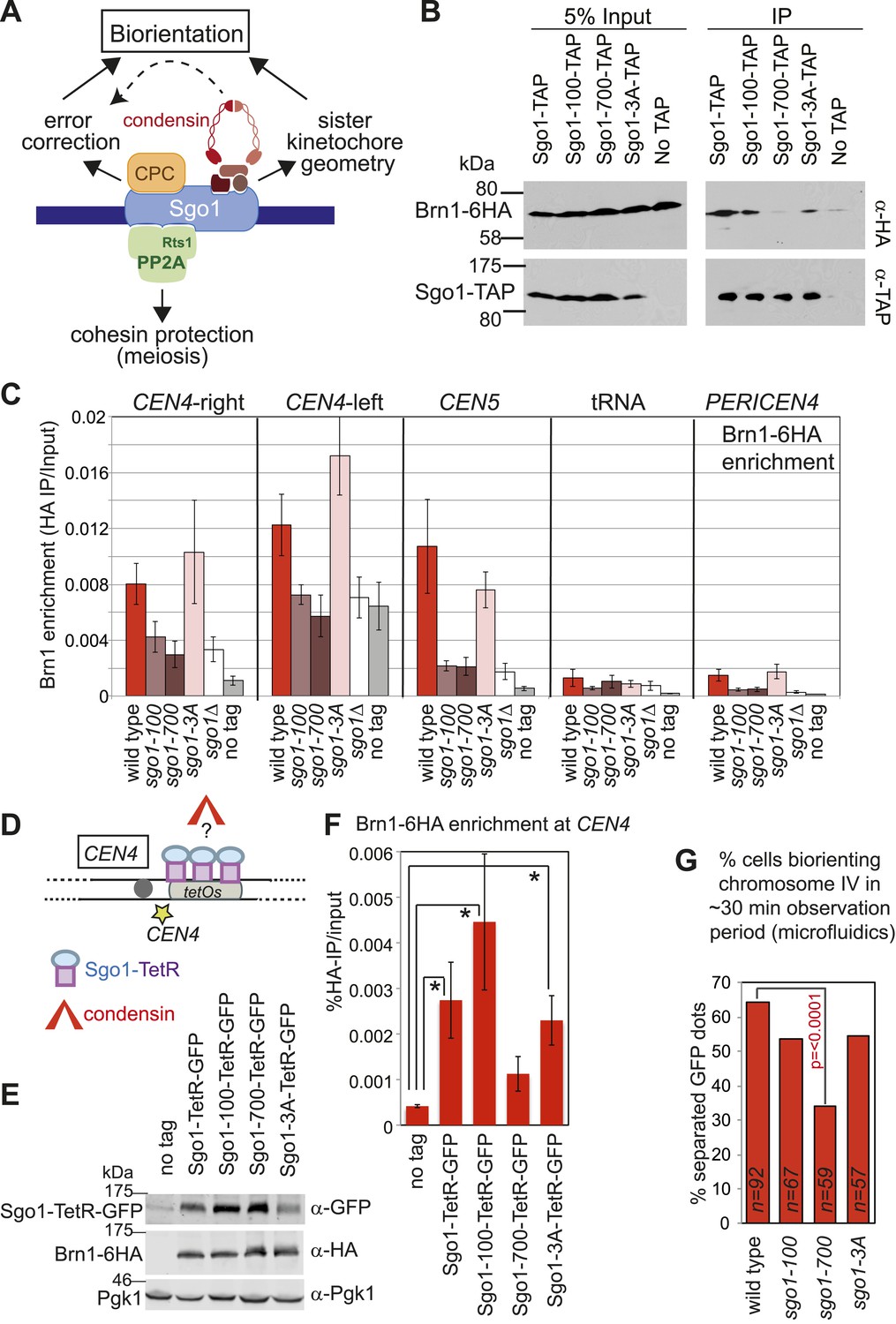
Shugoshin enables the bias towards sister kinetochore biorientation by condensin recruitment.
(A) Schematic diagram showing Sgo1-associated complexes and their functions at the pericentromere. (B) Sgo1-100 and Sgo1-3A, but not Sgo1-700, retain association with Brn1. Cells carrying BRN1-6HA and SGO1-SZZ(TAP) (AM9266), SGO1-100-SZZ(TAP) (AM9149), SGO1-700-SZZ(TAP) (AM9264), SGO1-3A-SZZ(TAP) (AM9262) or no TAP (AM5708) were arrested in nocodazole for 2 hr before treating with the cross-linker DSP. Prepared extracts were incubated with IgG-coupled beads and immunoprecipitates analyzed by immunoblotting with the indicated antibodies. (C) Brn1 is maintained at the centromere in metaphase-arrested sgo1-3A, but not sgo1-100 or sgo1-700 cells. Wild-type (AM5708), sgo1Δ (AM8834), sgo1-100 (AM9442), sgo1-700 (AM9291) and sgo1-3A (AM9276) cells carrying BRN1-6HA as well as a no tag control (AM1176) were arrested in nocodazole for 2 hr before harvesting for anti-HA ChIP. The levels of Brn1-6HA were measured at the indicated sites by qPCR. (D–F) Tethered Sgo1, Sgo1-100 or Sgo1-3A, but not Sgo1-700 can enrich Brn1-6HA at CEN4 in otherwise sgo1Δ cells. SGO1-tetR-GFP (AM14012), sgo1-100-tetR-GFP (AM13902), sgo1-700-tetR-GFP (AM13907), and sgo1-3A-tetR-GFP (AM13904) were introduced into cells carrying tetOs integrated at CEN4, producing Brn1-6HA and with SGO1 deleted from its endogenous locus. A strain carrying just tetOs integrated at CEN4 but otherwise wild type was used as a no tag control (AM11060). All strains were arrested in mitosis by treatment with nocodazole for 3 hr before harvesting for ChIP and immunoblotting. (D) Schematic diagram of the tethering locus. (E) Levels of Brn1 recruited adjacent to the tethering site (CEN4) when the indicated proteins are fused to TetR-GFP, as measured by ChIP-qPCR. The mean of four independent repeats is shown except for sgo1-3A-tetR-GFP where six repeats are included. Error bars are standard error, significance was calculated using the student t test (*p<0.05). (F) Total cellular levels of the Sgo1-tetR-GFP fusion proteins, Brn1-6HA and Pgk1 (loading control) were analyzed by immunoblot using the indicated antibodies. (G) The bias to sister kinetochore biorientation is absent in sgo1-700 cells. Wild-type (AM4643), sgo1-100 (AM8924), sgo1-700 (AM8925) and sgo1-3A (AM8923) cells were released from G1 and treated with nocodazole after SPB separation as in Figure 7A. The percentage of cells that separated CEN4-GFP foci at least once during the observation period is shown for the indicated strains. p values indicate significance (chi-square test).
Figure 9—figure supplement 1
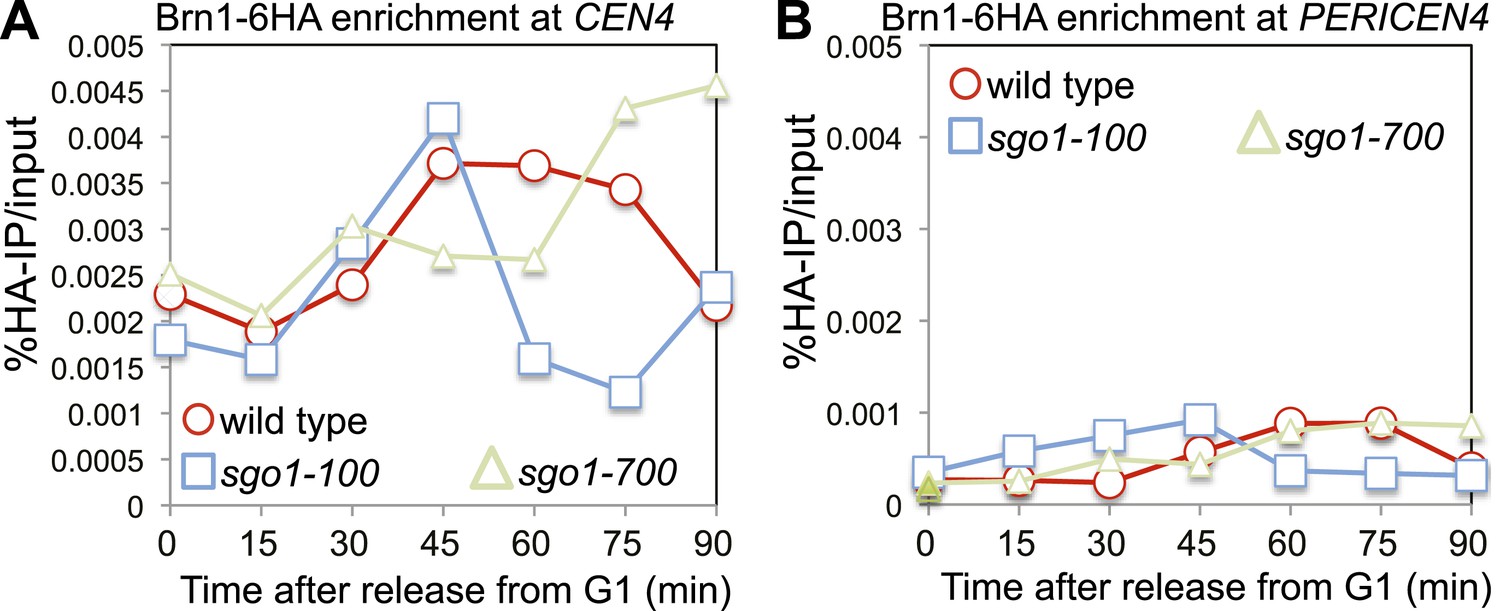
The Sgo1-100 protein can recruit condensin to kinetochores.
(A and B) Condensin is at least partially recruited to the centromere after release from G1 in sgo1-100 cells. Wild-type (AM5708), sgo1-100 (AM9442) and sgo1-700 (AM9291) cells carrying BRN1-6HA were arrested in G1 using alpha factor. Samples were extracted for anti-HA ChIP-qPCR at the indicated levels for analysis of Brn1 association with CEN4 (A) and the pericentromere of chromosome IV (B).
Figure 9—figure supplement 2
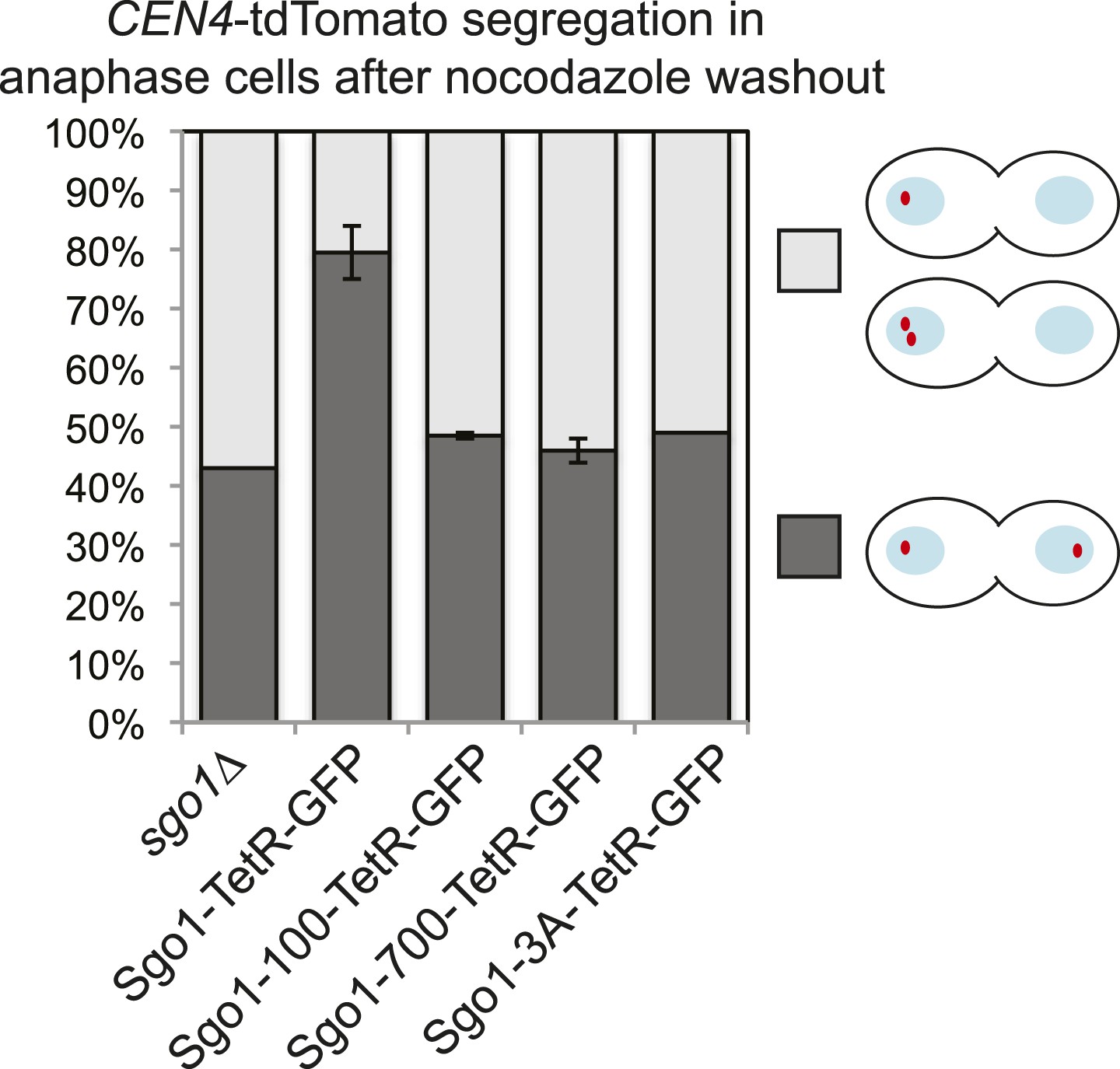
Sgo1-tetR-GFP, but not Sgo1-100-tetR-GFP, Sgo1-700-tetR-GFP or Sgo1-3A-tetR-GFP tethered to CEN4 can partially rescue the mis-segregation of chromosome IV after nocodazole washout in otherwise sgo1Δ cells.
Diploid cells carrying tetR-tdTomato and SGO1-tetR-GFP (AM14005), sgo1-100-tetR-GFP (AM14006), sgo1-700-tetR-GFP (AM14007), sgo1-3A-tetR-GFP (AM14008) or no Sgo1-TetR fusion (AM14009), with tetOs integrated at CEN4 and with SGO1 deleted from its endogenous locus were treated with nocodazole to depolymerize microtubules and arrest cells in mitosis. Nocodazole was washed out, allowing microtubules to reform and, 60 min later, the position of CEN4-tdTomato foci was scored in the anaphase cells after chromosome segregation. At least 100 cells were scored from each of two experimental repeats with error bars representing the range, except for the sgo1-3A-tetR-GFP strain, where results are shown from a single experiment.
Figure 10
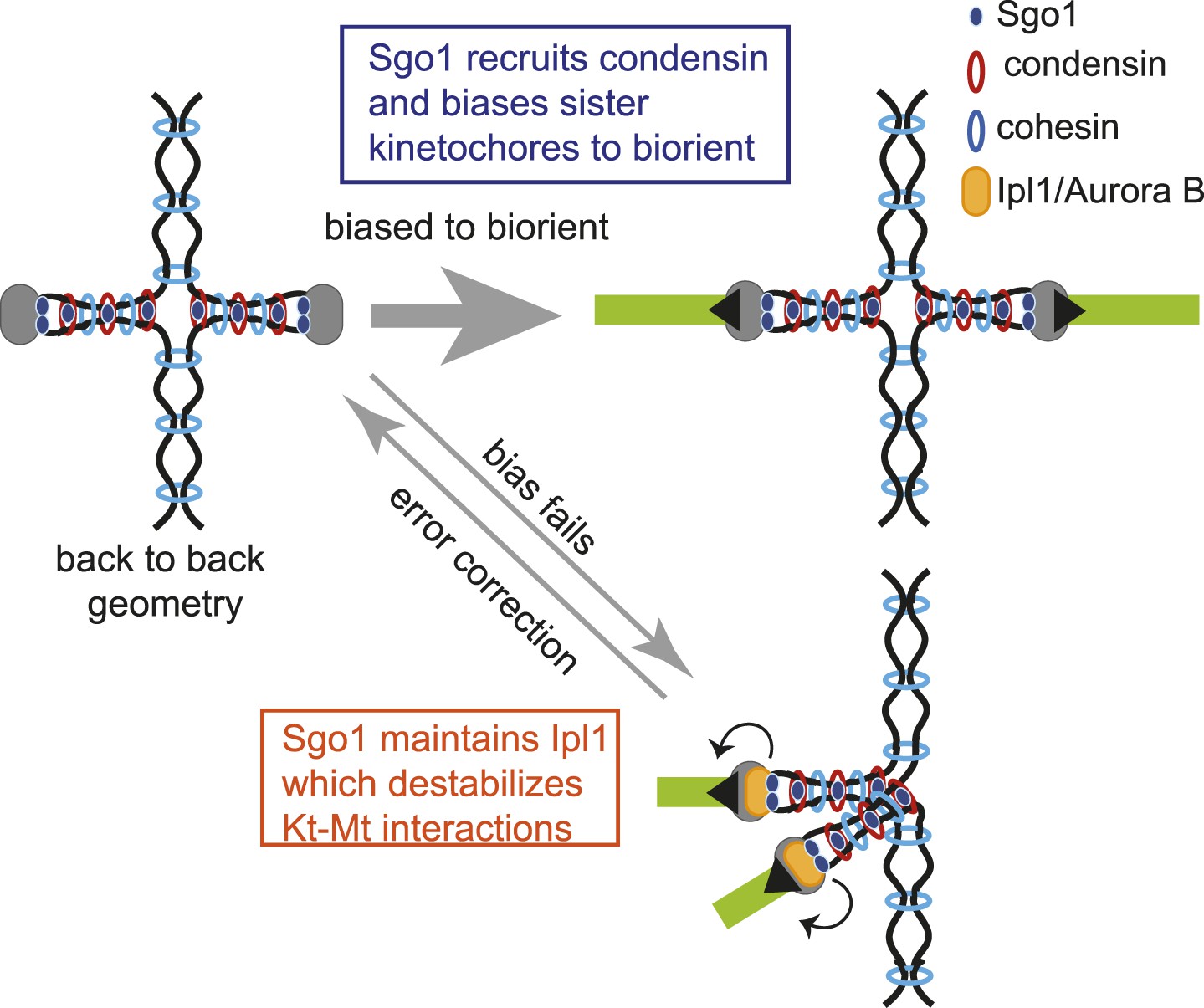
Model for dual role of Sgo1 in biorientation is shown.
Shugoshin ensures sister kinetochore biorientation through two mechanisms. First, early in the cell cycle, shugoshin mediates the enrichment of condensin within the pericentromere. We propose that condensin enables the pericentromere to adopt a geometry that favors the capture of sister kinetochores by microtubules from opposite poles, thereby biasing them to biorient. Second, shugoshin maintains aurora B at the pericentromere for those kinetochores that fail to biorient and come under tension. Aurora B destabillizes these tension-less attachments, thereby providing a further chance to for sister kinetochores to make the appropriate, tension-generating attachments. We suggest that condensin facilitates this ‘error correction’ process by conferring a rigid structure to the pericentromere upon which aurora B can act.
Videos
Video 1
Example video of a wild-type cell in the error correction assay.
The video corresponds to the image gallery in Figure 5C (upper panel).
Video 2
Example video of an ipl1-as cell in the error correction assay.
The video corresponds to the image gallery in Figure 5C (lower panel).
Video 3
Example video of a wild type cell in the assay to test for a bias towards sister kinetochore biorientation.
The video corresponds to the image gallery in Figure 6C (upper panel).
Video 4
Example video of a YCS5-aid cell in the assay to test for a bias towards sister kinetochore biorientation.
The video corresponds to the image gallery in Figure 6C (lower panel).
Additional files
-
Supplementary file 1
Complete list of peptides identified in the experiments shown in Figure 4A,B.
- https://doi.org/10.7554/eLife.01374.031
-
Supplementary file 2
(A) Yeast strains used in this study. (B) qPCR primers used in this study. (C) Genome summary table for Brn1-6HA ChIP-seq.
- https://doi.org/10.7554/eLife.01374.032
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Shugoshin biases chromosomes for biorientation through condensin recruitment to the pericentromere
eLife 3:e01374.
https://doi.org/10.7554/eLife.01374




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}