Essential role of lncRNA binding for WDR5 maintenance of active chromatin and embryonic stem cell pluripotency
- Howard Hughes Medical Institute, Stanford University School of Medicine, United States
- Stanford University School of Medicine, United States
- University of Michigan, United States
- Howard Hughes Medical Institute, University of Michigan, United States
Figures
Figure 1 with 1 supplement
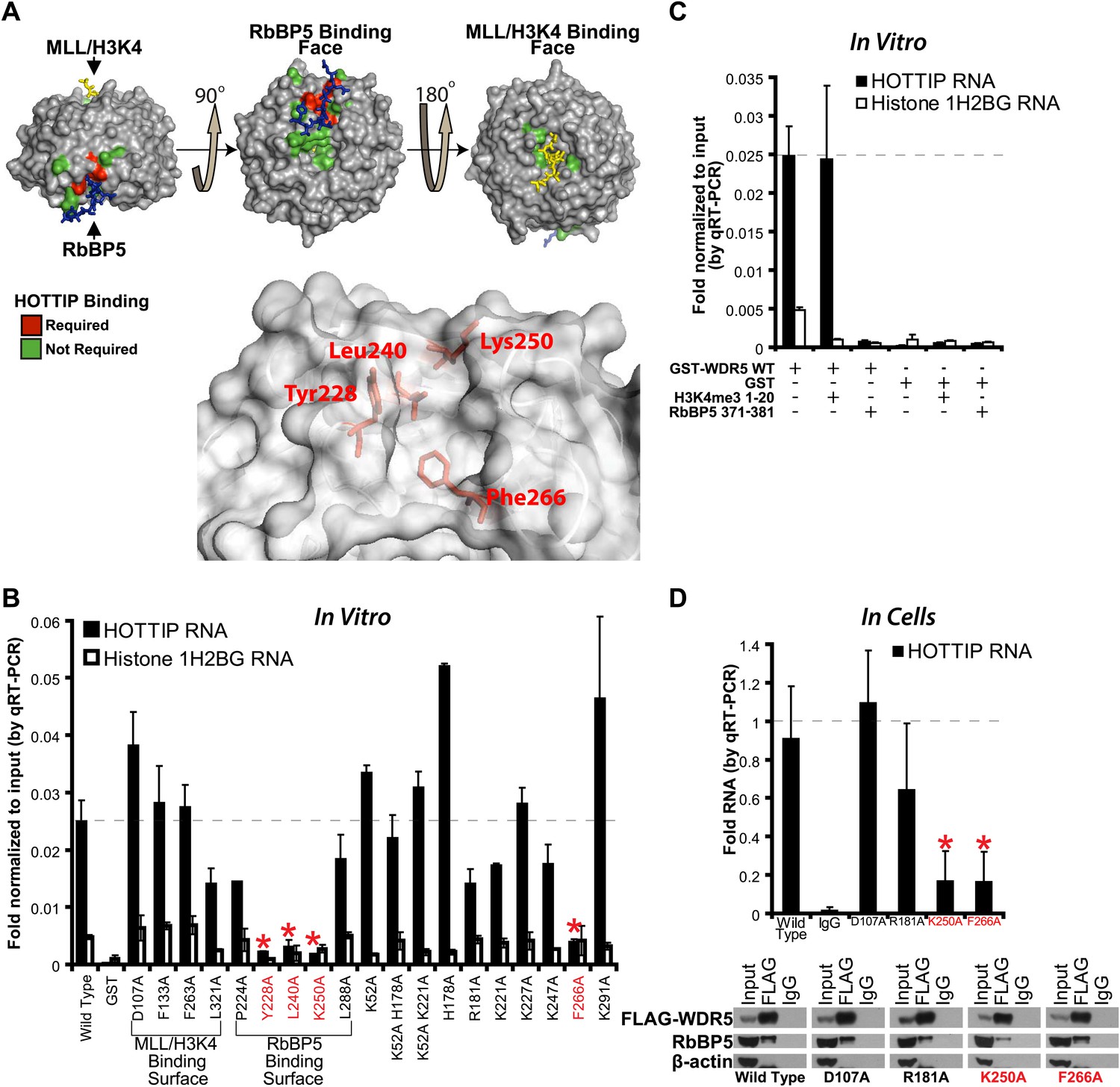
HOTTIP lncRNA binding surface overlaps with the RbBP5 binding surface on WDR5.
(A) Crystal structure of WDR5 (PDB:3P4F, visualized with PyMol) reveal that mutations abrogating HOTTIP RNA binding align along a cleft between blades 5 and 6, opposite the H3K4/MLL binding site. This binding surface overlaps the site for RbBP5. Top: schematic of tested and HOTTIP binding mutations. Bottom: magnification of HOTTIP binding cleft. Yellow: MLL peptide. Blue: RbBP5 peptide. (B) qRT-PCR results of indicated GST-WDR5 mutants tested by in vitro assay for binding to HOTTIP RNA or control Histone 1H2BG RNA. (C) RbBP5 peptide competition assay indicates that RbBP5 amino acids 371–381 can fully inhibit HOTTIP binding to WDR5. Wild type data same as in Figure 1B. (D) Average fold pulldown of full length HOTTIP by cell-based native RNA immunoprecipitation of select WDR5 mutants. Top: qRT-PCR results of RNA immunoprecipitation of FLAG-WDR5 mutants. All values are normalized to input, then to FLAG pulldown of wild type and positive control D107A. Negative RNA controls (U1, HOTAIR) and reaction without reverse transcription (-RT) show minimal enrichment (Figure 1—figure supplement 1, data not shown). Bottom: representative western blots of immunoprecipitations.
Figure 1—figure supplement 1
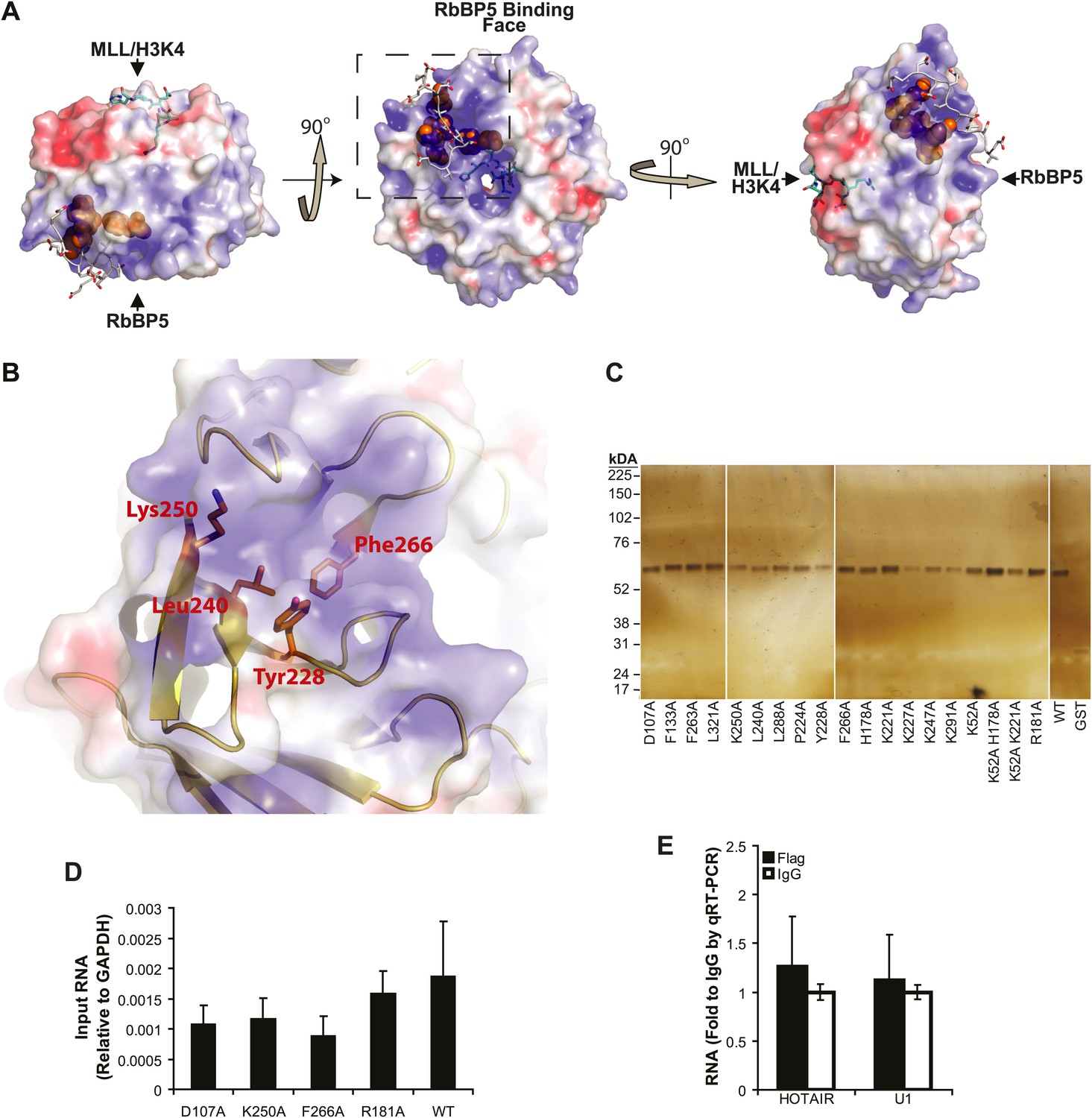
lncRNA binding surface of WDR5 and WDR5 RIP-qRT-PCR standards.
(A–B) Electrostatic surface contouring of the WDR5 crystal structure (PDB:3P4F, visualized with PyMol) reveal that mutations abrogating HOTTIP RNA binding de_ne an electropositively charged region. Orange: WDR5 residues important for HOTTIP binding. Cyan: MLL peptide. Grey: RbBP5 peptide. (C) Silver stain of post-assay glutathione-bound GST-WDR5 mutants indicates no protein degradation or loading bias occurred during in vitro binding assay. (D) Average levels of input HOTTIP RNA for cell-based native RNA immunoprecipitation assays of WDR5 mutants by qRT-PCR, relative to GAPDH. (E) Average fold pulldown of HOTAIR or U1 control RNAs by cell-based native RNA immunoprecipitation of WDR5, demonstrating no enrichment compared with IgG control.
Figure 2
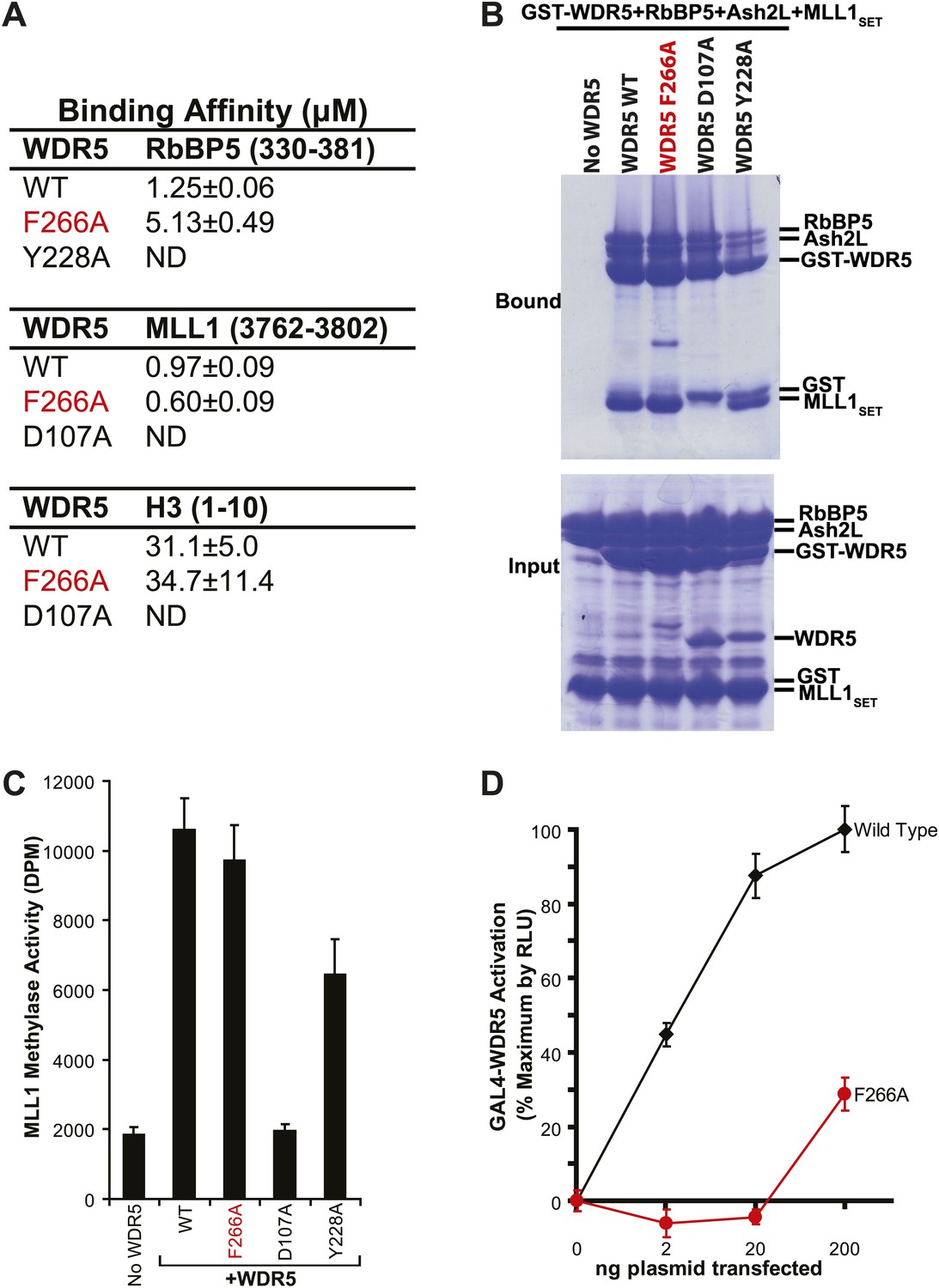
WDR5 F266A lncRNA binding mutant does not affect MLL complex formation or catalytic function, but shows decreased ability to activate target genes in 293T cells.
(A) WDR5 binding affinity to RbBP5, MLL1, or H3 peptides by isothermal calorimetry. ND, not detectable. (B) WDR5 F266A does not have decreased binding to RbBP5, Ash2L and MLL1SET domain proteins in GST protein pull down assays. (C) WDR5 F266A does not affect histone methylase activity of the MLL1 complex. (D) GAL4-WDR5 F266 is defective in activating luciferase expression, as seen in luciferase titration tests.
Figure 3
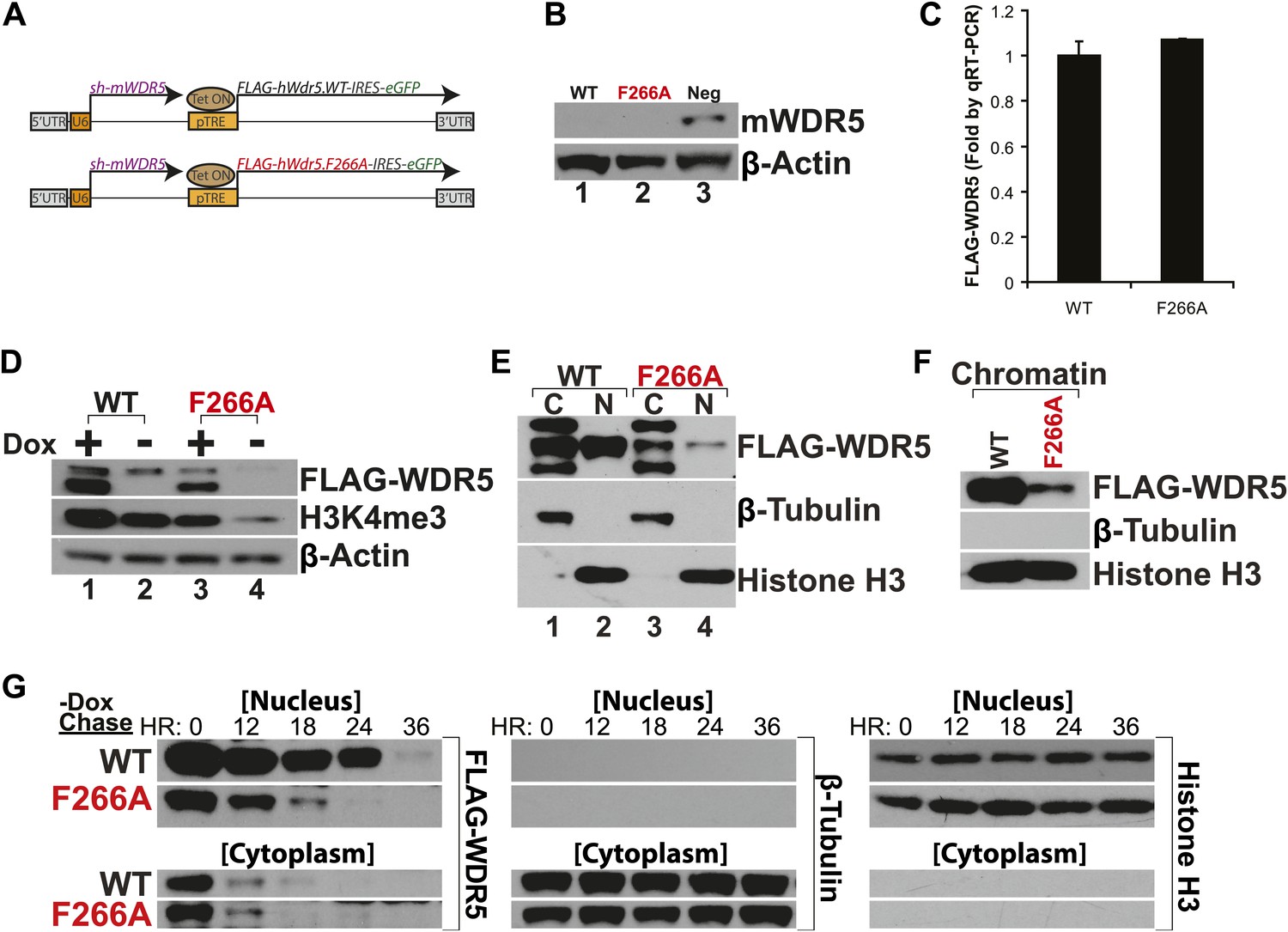
WDR5 F266A mutation decreases protein stability and localization to chromatin.
(A) Schematic of lentiviral vectors, modified from (Ang et al., 2011). (B) Western blot demonstrating successful mouse WDR5 knockdown. (C) qRT-PCR results demonstrating equal RNA expression of human WDR5 WT and WDR5 F266A. (D) Western blot of WDR5 WT and WDR5 F266A protein expression, also with 4 days after doxycycline removal. (E) WDR5 F266A is defective in nuclear accumulation, compared with WDR5 WT. (F) WDR5 F266A reduces chromatin association, as seen in chromatin isolation experiments. (G) WDR5 F266A mutation decreases protein stability in the nucleus after doxycycline withdrawal. Both WDR5 WT and WDR5 F266A are similarly unstable in the cytoplasmic fraction.
Figure 4 with 1 supplement
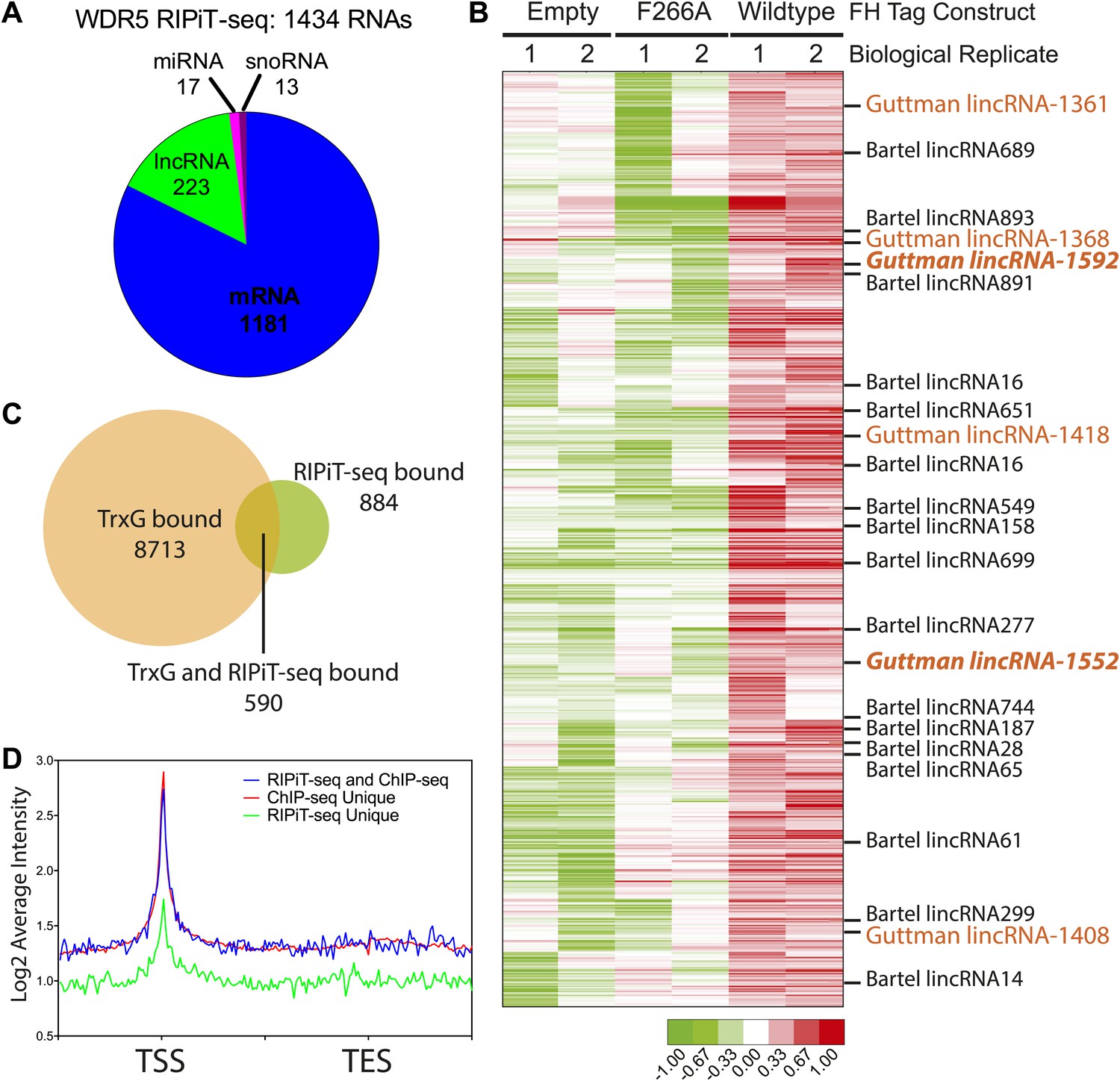
WDR5 binds to a family of ESC RNAs.
(A) WDR5 RIPiT-seq retrieves diverse class of RNAs. (B) 1434 RNAs bind to wild type WDR5 but not F266A mutant or vector control. Each column is a RIPiT-seq experiment; each row is a transcript. Red indicates enrichment over input and WDR5 F266A mutant. Known ESC lncRNAs are listed; those with known functions in ESC pluripotency or differentiation are highlighted in orange. Two lincRNA loci that are both bound by WDR5 on chromatin and generate lincRNAs that bind WDR5 are highlighted in orange bold font. (C) Overlap of genes retrieved by WDR5 RIPiT-seq vs ChIPseq. (D) Metagene analysis of WDR5 ChIP-seq signal in the indicated classes of genes.
Figure 4—figure supplement 1
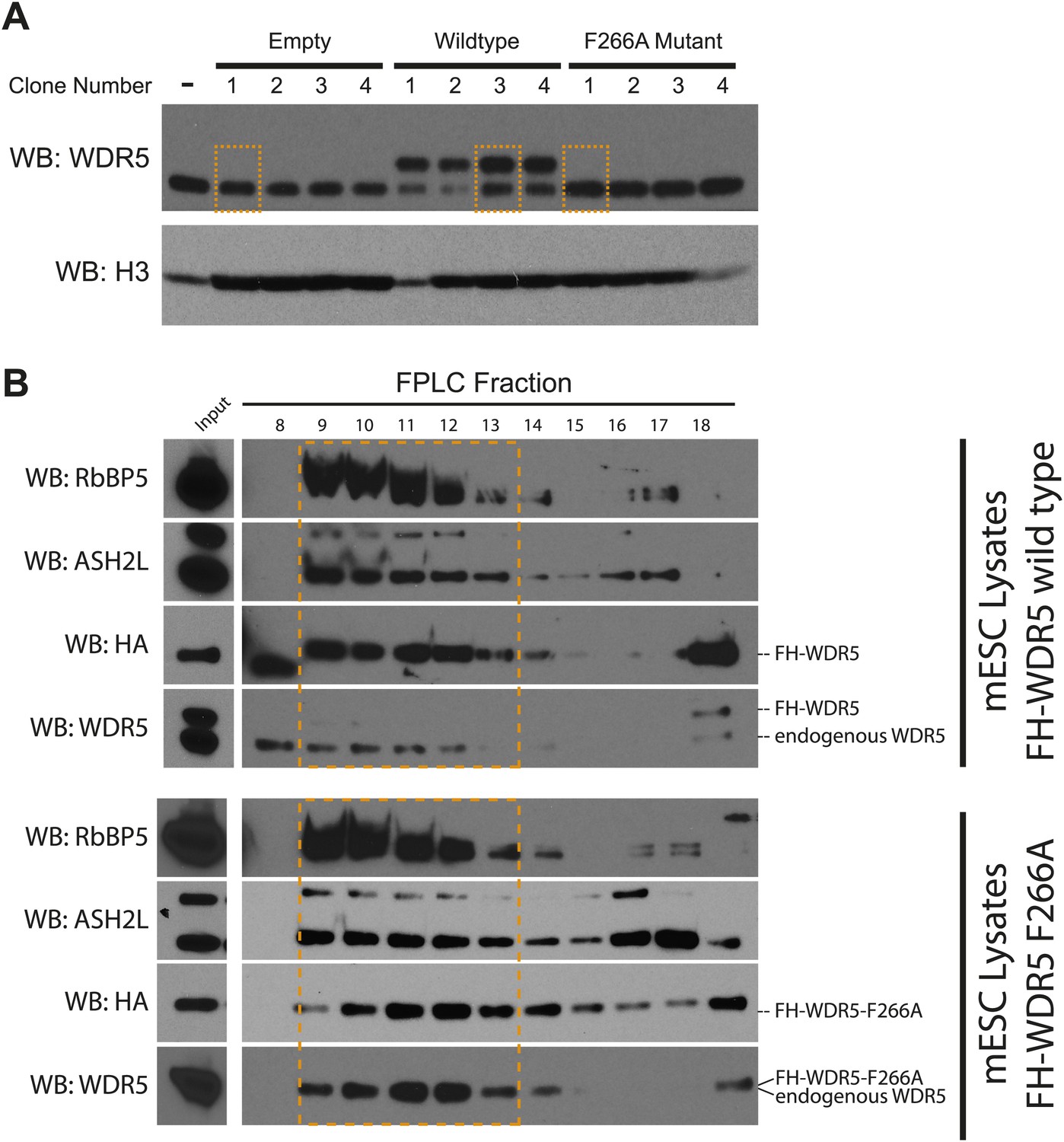
Characterization of FLAG-HA ES cell lines.
(A) Western blot analysis of individually isolated ESC clones. antiWDR5 blotting reveals the total expression level of WDR5 in mock, FH-WDR5 wild type, and FH-WDR5 F266A mutant ESCs. The FH tag adds ∼8 kDa to WDR5, resulting in the observed shift, where as the F266A sees no shift because the construct is N-terminally shorter. Orange boxes are draw around clones that were isolated and used in the RIPiT experiments due to their near endogenous expression level. (B) FPLC analysis of the selected wild type and F266A FH-WDR5 clones. Each fraction was subjected to western blot analysis for HA (tagged protein), WDR5, Ash2L, and RbBP5. Orange boxes appear around the endogenous large molecular weight MLL complex, where the majority of the tagged FH-WDR5 constructs elute.
Figure 5

WDR5 F266A mutation causes defects in ESC self renewal and increases differentiation.
(A and B) Alkaline phosphatase staining and morphology of ESC colonies after 6 days growth in conditions specified. WDR5 F266A demonstrates reduced alkaline phosphatase positive colonies with increased differentiation compared with wild type. (A) Representative wells and colony morphology. Scale bar represents 200 μm. (B) Quantitation of colonies per well. (C) Microarray data reveal that both WDR5 F266A +dox and WDR5 WT −dox displayed loss of self renewal genes. WDR5 F266A +dox further demonstrates increased expression of ectodermal and mesodermal markers. Mixl1 is a mesendoderm marker indicative of potential for both endoderm and mesoderm. (D and E) qRT-PCR validation of (D) reduced expression of ESC pluripotency genes and (E) ectopic expression of differentiation genes with either WDR5 F266A expression or shutoff of WDR5 WT. qRT-PCR values were first normalized to the housekeeping gene beta-actin, then normalized to WT. Mean +/− SD are shown.
Figure 6

Model for RNA-mediated switch of protein turnover to activate target genes.
Expressed lncRNAs bind the WDR5 protein to increase protein stabilization on chromatin, which facilitates MLL complex assembly and methyltransferase activity for target gene activation. Without lncRNA binding, WDR5 fails to associate effectively with chromatin and is rapidly degraded.
Additional files
-
Supplementary file 1
Protein coding genes common between RIPiT-seq Enriched RNAs and mRNAs identified from Ivanova et al. (2006).
- https://doi.org/10.7554/eLife.02046.011
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Essential role of lncRNA binding for WDR5 maintenance of active chromatin and embryonic stem cell pluripotency
eLife 3:e02046.
https://doi.org/10.7554/eLife.02046




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}