The asymmetry of female meiosis reduces the frequency of inheritance of unpaired chromosomes
- University of California, Davis, United States
- University of Calgary, Canada
Figures
Figure 1
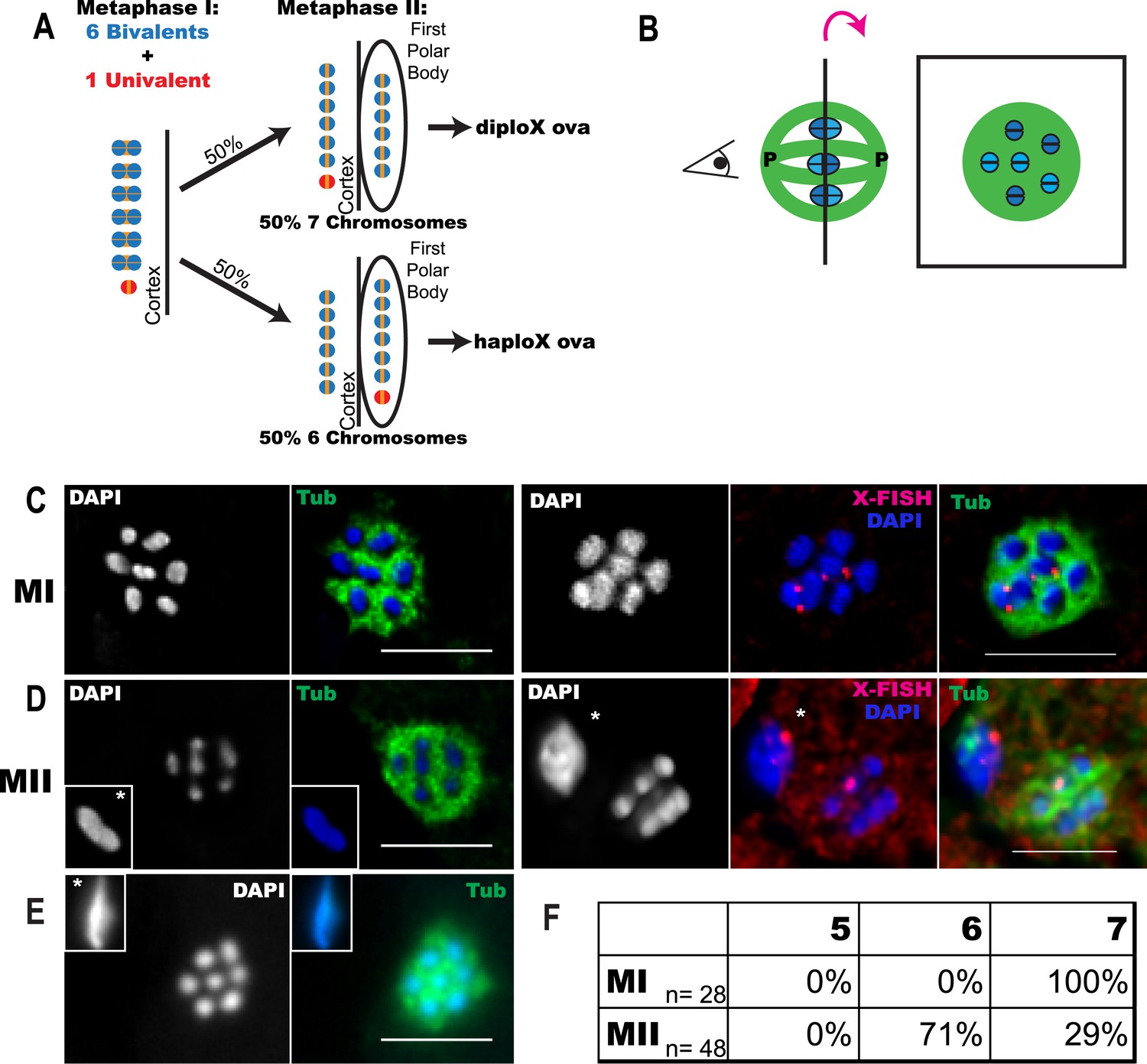
Trisomy correction during meiosis I.
(A) Illustration showing expected outcomes of female meiosis in XXX wild-type worms, assuming the extra univalent X (red) does not lose cohesion (yellow) between sister chromatids during anaphase I and assuming random segregation. (B) Illustration of a spindle with chromosomes at the metaphase plate with poles marked ‘P’ (left) and a projection of the cross-sectional view down the pole-to-pole axis at the metaphase plate (right). (C–E) Z projections of fixed meiotic embryos viewed down the pole-to-pole spindle axis. Meiotic embryos from XXX wild-type mothers were stained with DAPI and anti-tubulin antibody. (C) Metaphase I spindles with 7 chromosomes; right images of X-fluorescence in situ hybridization (FISH) show two X chromosomes on the spindle. See also Supplementary file 2. (D) Metaphase II spindles with 6 chromosomes; right images of X-FISH show one X on the spindle and 2–3 foci in the polar body. See also Supplementary file 3. (E) Metaphase II spindle with 7 chromosomes. (F) Frequency of each spindle class among the progeny of XXX wild-type mothers. Insets show polar bodies, marked by asterisks, which were used to identify metaphase II spindles. Bar = 5 μm.
Figure 2 with 1 supplement

X univalents are preferentially lost between metaphase I and metaphase II in him-8 mutants.
Z projections of living (A–E) and fixed (F–J) C. elegans meiotic embryos viewed down the pole-to-pole spindle axis at metaphase I (A, B, F, G) or metaphase II (C, D, E, H, I, J). mCherry::Histone H2B and GFP::tubulin label the chromosomes and spindle, respectively, in live embryos. Fixed embryos were stained with DAPI, anti-tubulin antibody, and a LacO FISH probe that recognizes a LacO array integrated on the X chromosome. Asterisks indicate polar bodies. Insets show polar bodies that did not fit in the image frame. In (E), ‘s’ denotes a sperm outside of the embryo. Percentages are shown for each outcome (K, L). Bar = 5 μm.
Figure 2—figure supplement 1
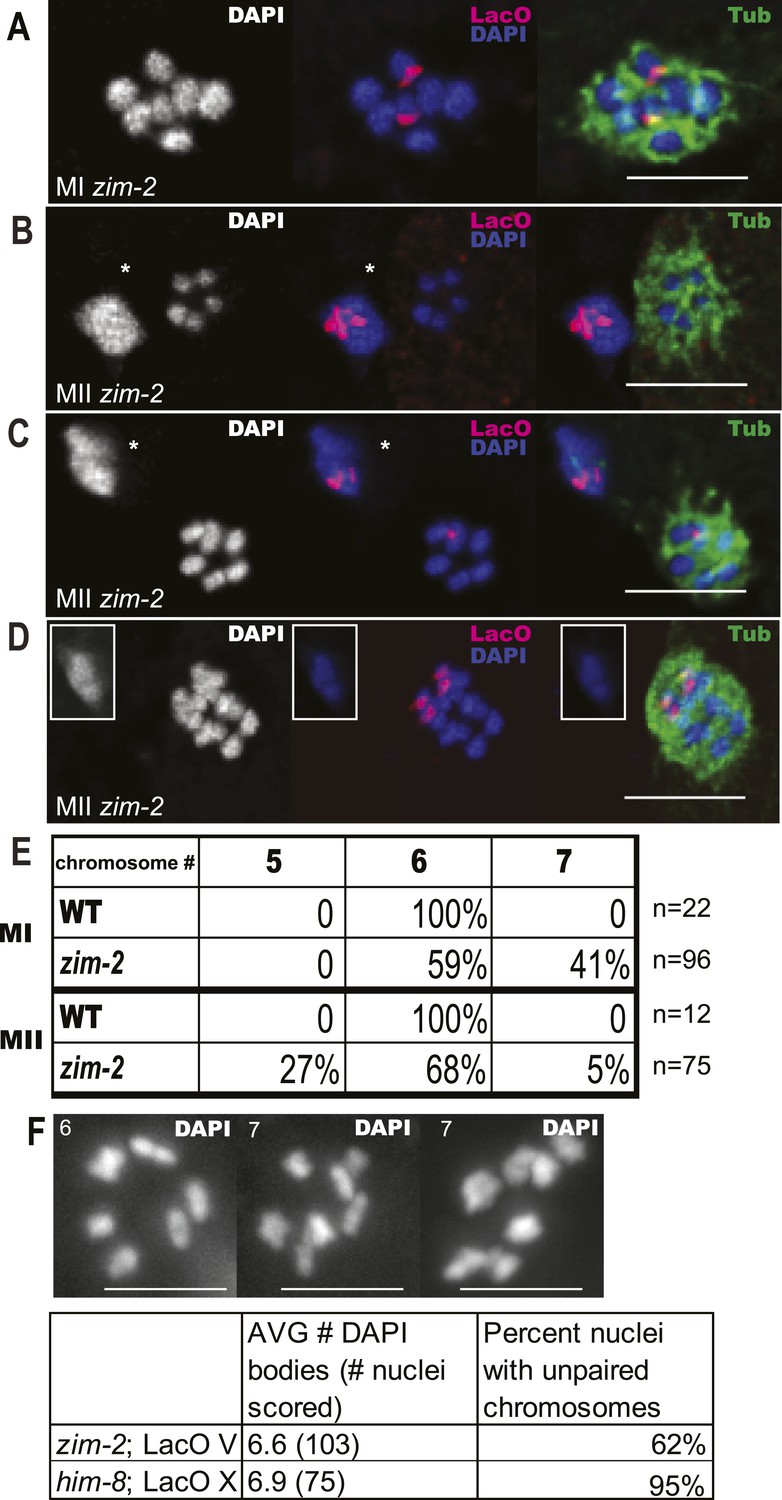
zim-2 embryos also deposit unpaired chromosome V univalents into the first polar body.
Z projections of fixed meiotic embryos viewed down the pole-to-pole spindle axis. Embryos were stained with DAPI, anti-tubulin antibody, and a LacO FISH probe that recognizes a LacO array integrated on chromosome V. This array is larger than the array inserted on X; thus, the foci are larger than those shown in Figure 2. (A) Metaphase I embryo with 7 DAPI chromosomes and two LacO (V) univalents. (B) Metaphase II embryo with 5 DAPI chromosomes and no LacO (V) chromosome on the spindle and two in the first polar body. (C) Metaphase II embryo with 6 DAPI chromosomes and one LacO (V) chromosome on the spindle and one in the first polar body. (D) Early anaphase II spindle with 2 LacO (V) chromosomes in the spindle and none in the first polar body. In (B) and (C), polar bodies are marked by asterisks. In (D), the polar body is shown as an inset because it was in a distant focal plane. (E) Quantification of the frequencies of each class. (F) Diakinesis chromosome counts from the zim-2 and him-8 strains bearing Lac operator arrays. Left panel shows representative zim-2 diakinesis nucleus with 6 chromosomes, right two panels show two examples of zim-2 diakinesis nuclei with 7 chromosomes. Bar = 5 µm.
Figure 3 with 2 supplements
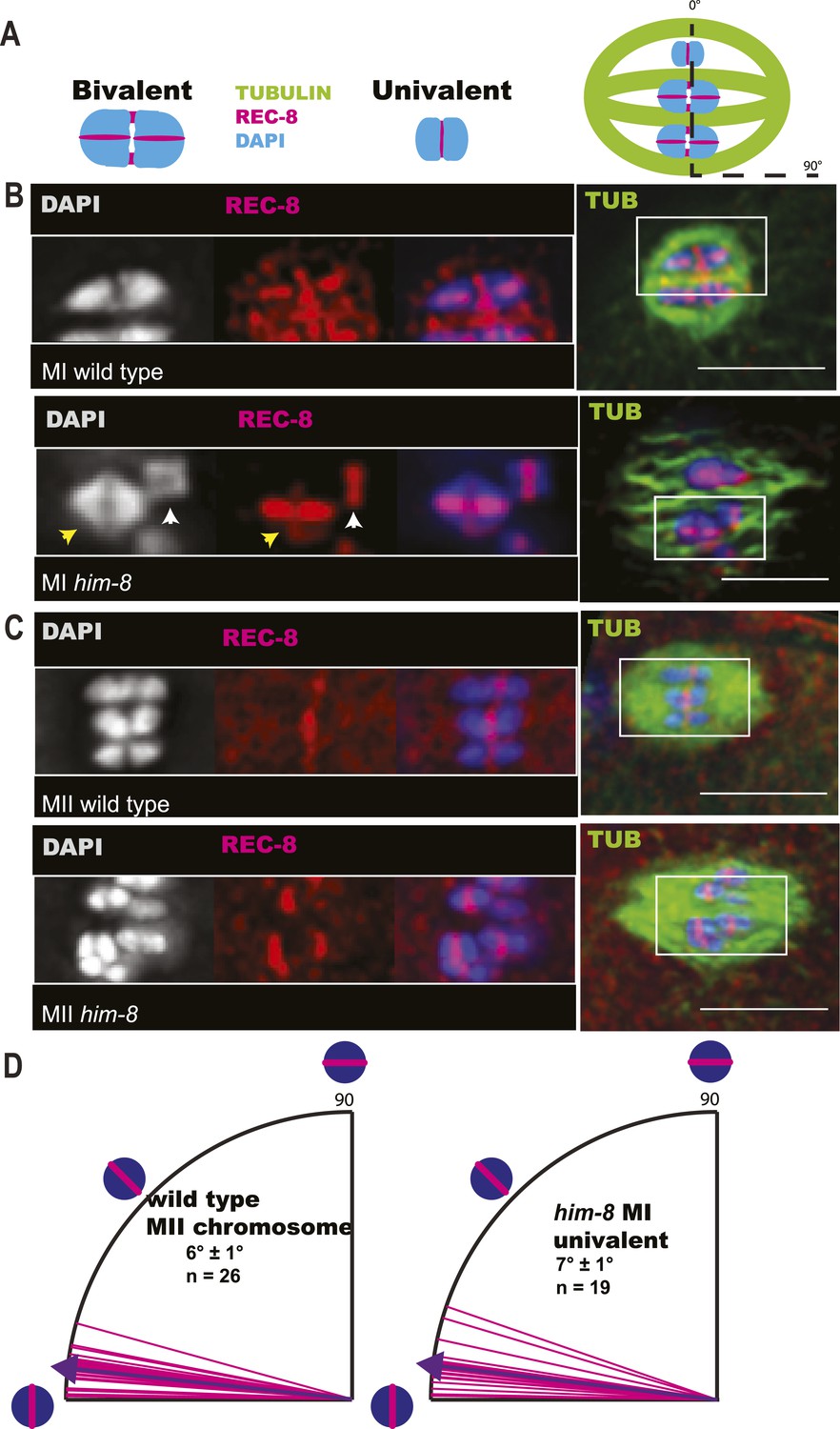
X univalents biorient at metaphase I in him-8 embryos.
(A) Cartoon diagram of REC-8 staining on bivalents and univalents. (B and C) Anti-REC-8 staining of metaphase I and metaphase II embryos with bivalents (yellow arrow head) and univalents (white arrow head). In him-8 embryos, univalents at metaphase I have a single band of REC-8 with the same orientation seen on normal chromosomes at metaphase II. (D) Quantification of the orientation of univalents by offset angle from the metaphase plate, 0° corresponds to perfect biorientation and 90° corresponds to perfect mono-orientation. Cortical pole is on the left in all images. Bar = 5 μm.
Figure 3—figure supplement 1
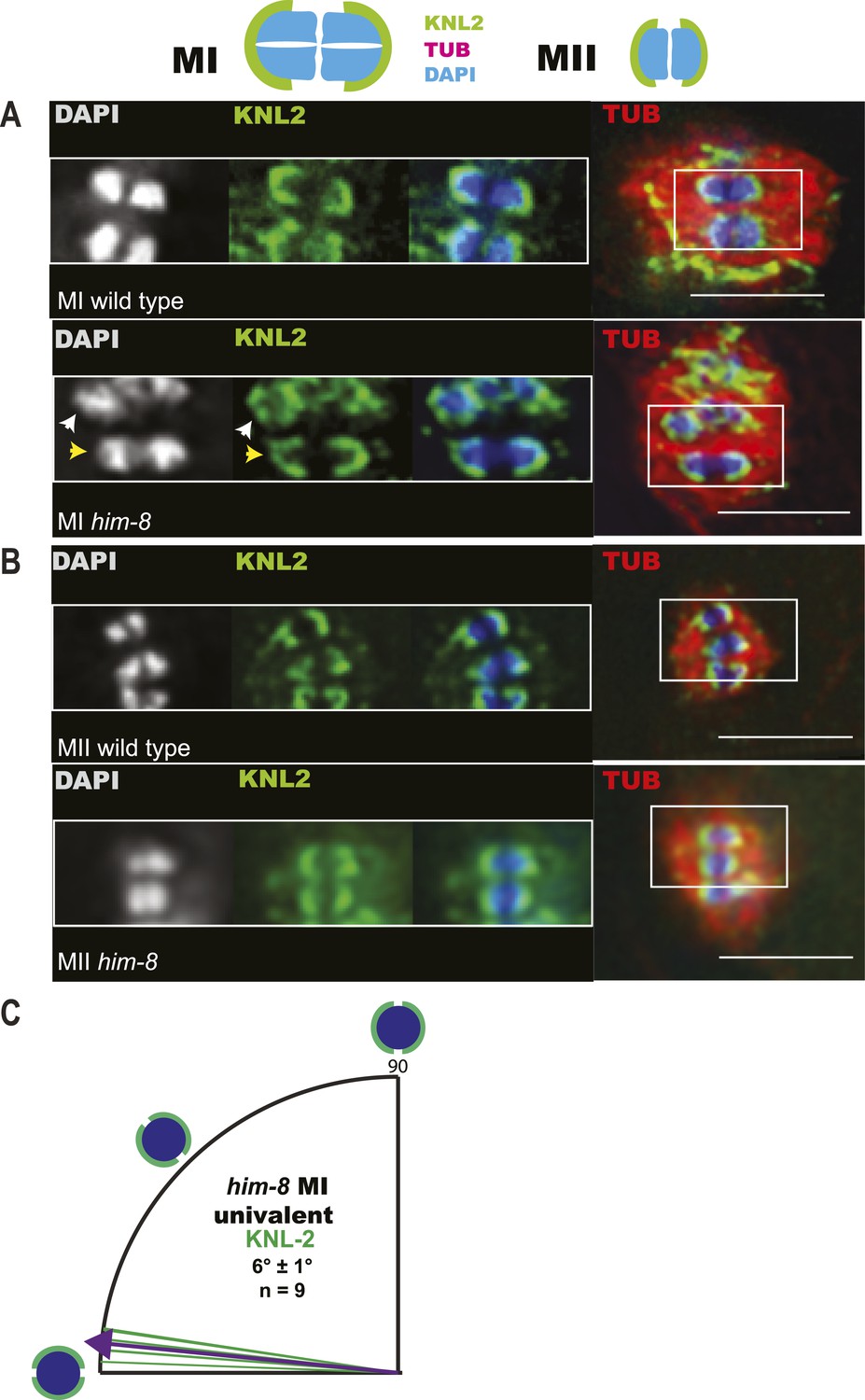
Imaging of GFP::KNL-2 demonstrates that him-8 univalent chromosomes biorient at metaphase of meiosis I.
(A–B) Z-projections through fixed GFP::KNL-2 embryos stained with DAPI and anti-tubulin antibody. (A) Metaphase I wild-type and him-8 embryos showing the distinct KNL-2 cups around bivalents (yellow arrow head) and univalents (white arrow head). (B) Metaphase II wild-type and him-8 embryos showing the characteristic KNL-2 cups around bioriented chromosomes at the metaphase plate. (C) Quantification of the orientation of chromosomes by offset angle from the metaphase plate, 0° corresponds to perfect biorientation and 90° corresponds to perfect mono-orientation. Cortical pole is on the left in all images. Bar = 5 μm.
Figure 3—figure supplement 2
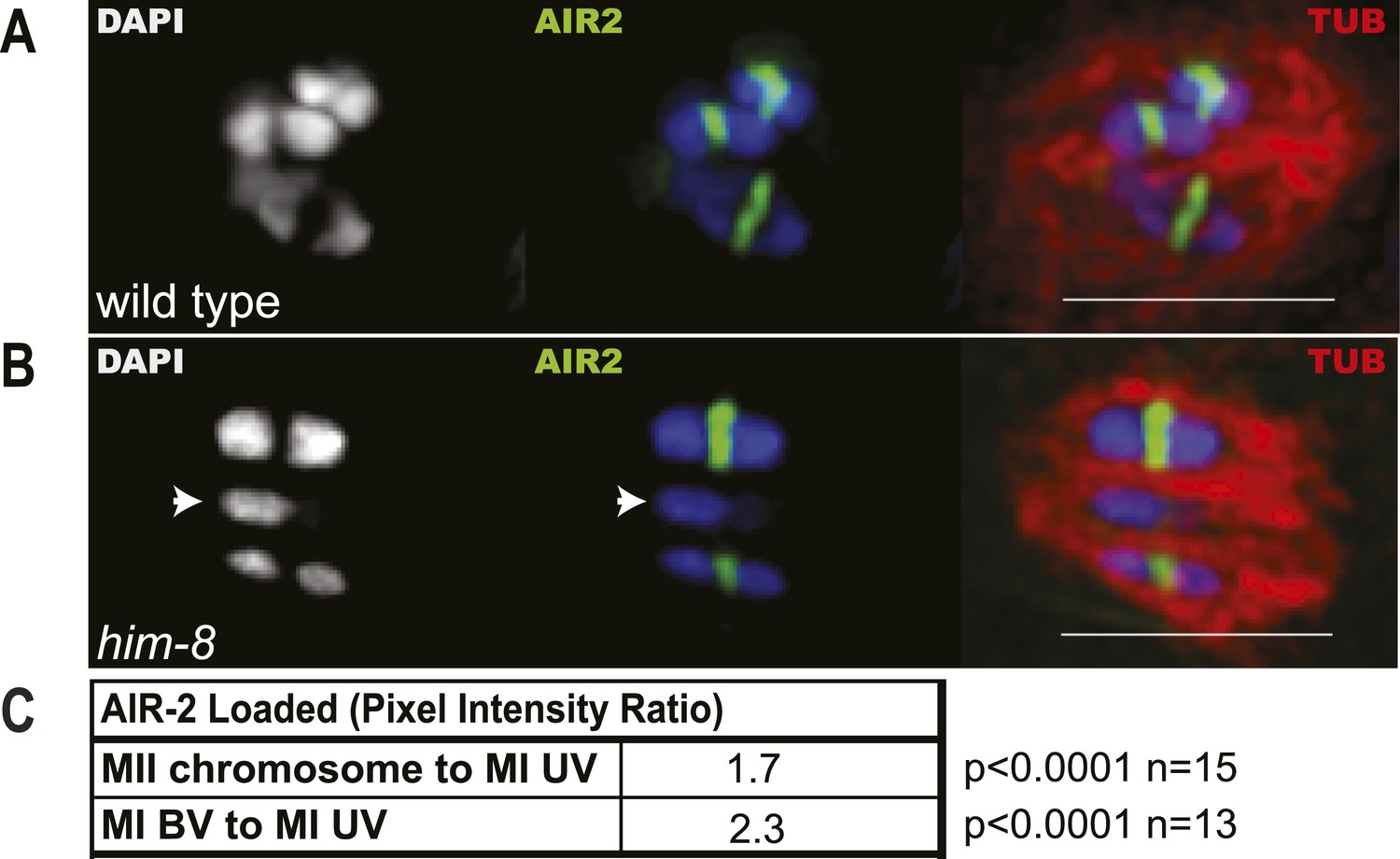
Reduced levels of AIR-2 are loaded on him-8 X univalents at meiosis I.
(A and B) Z projections of fixed metaphase I, GFP:AIR-2 embryos stained with DAPI and anti-tubulin antibody. AIR-2 is loaded between homologs of both wild-type (A) and him-8 bivalents (B), whereas less AIR-2 was observed on him-8 univalents (arrow heads in B). (C) Relative pixel intensity ratios show that him-8 metaphase II chromosomes load 1.7 times as much AIR-2 as X univalents at metaphase I, and metaphase I bivalents load 2.3 times as much AIR-2 as X univalents at metaphase I. n for both refers to total number of embryos counted, where each metaphase II embryo bore 5–7 chromosomes and each metaphase I embryo bore 2 univalents and 5 bivalents. Bar = 5 μm.
Figure 4
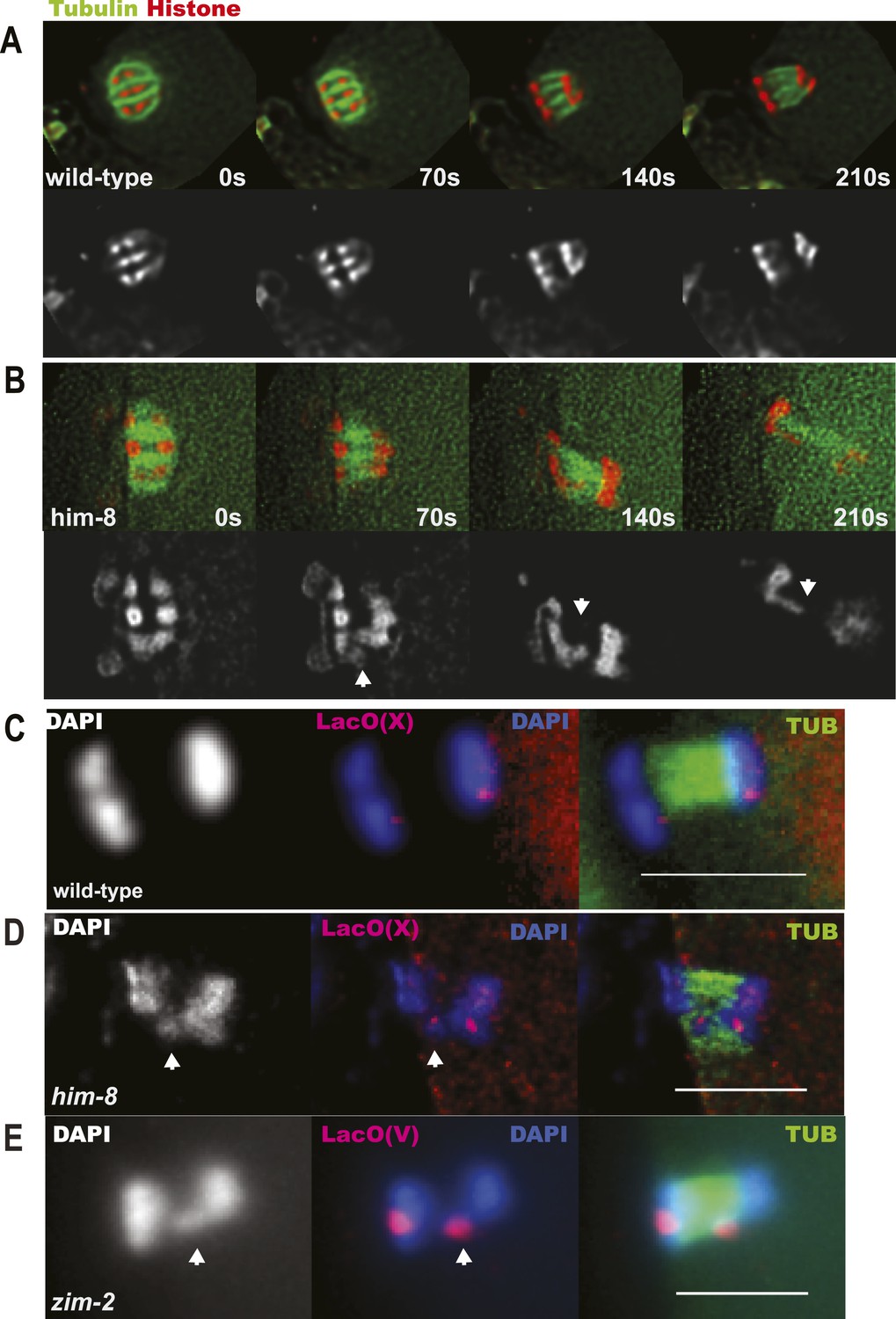
X univalents lag at anaphase I.
(A) Time-lapse images of a living wild-type embryo undergoing anaphase I show chromosomes separating as two distinct masses. (B) Time-lapse images of a living him-8 embryo show a lagging chromosome at anaphase I. (C–E) Z projections of fixed anaphase I embryos. (C) LacO FISH labeling of a wild-type strain with a LacO array integrated on the X chromosome shows normal segregation of two X homologs from one X bivalent. (D) LacO FISH shows that a lagging chromosome in him-8 is the X. (E) LacO FISH labeling of a zim-2 strain with a LacO array integrated on chromosome V showing a univalent V lagging at anaphase I. Cortical pole is to the left in all images. Bar = 5 μm.
Figure 5
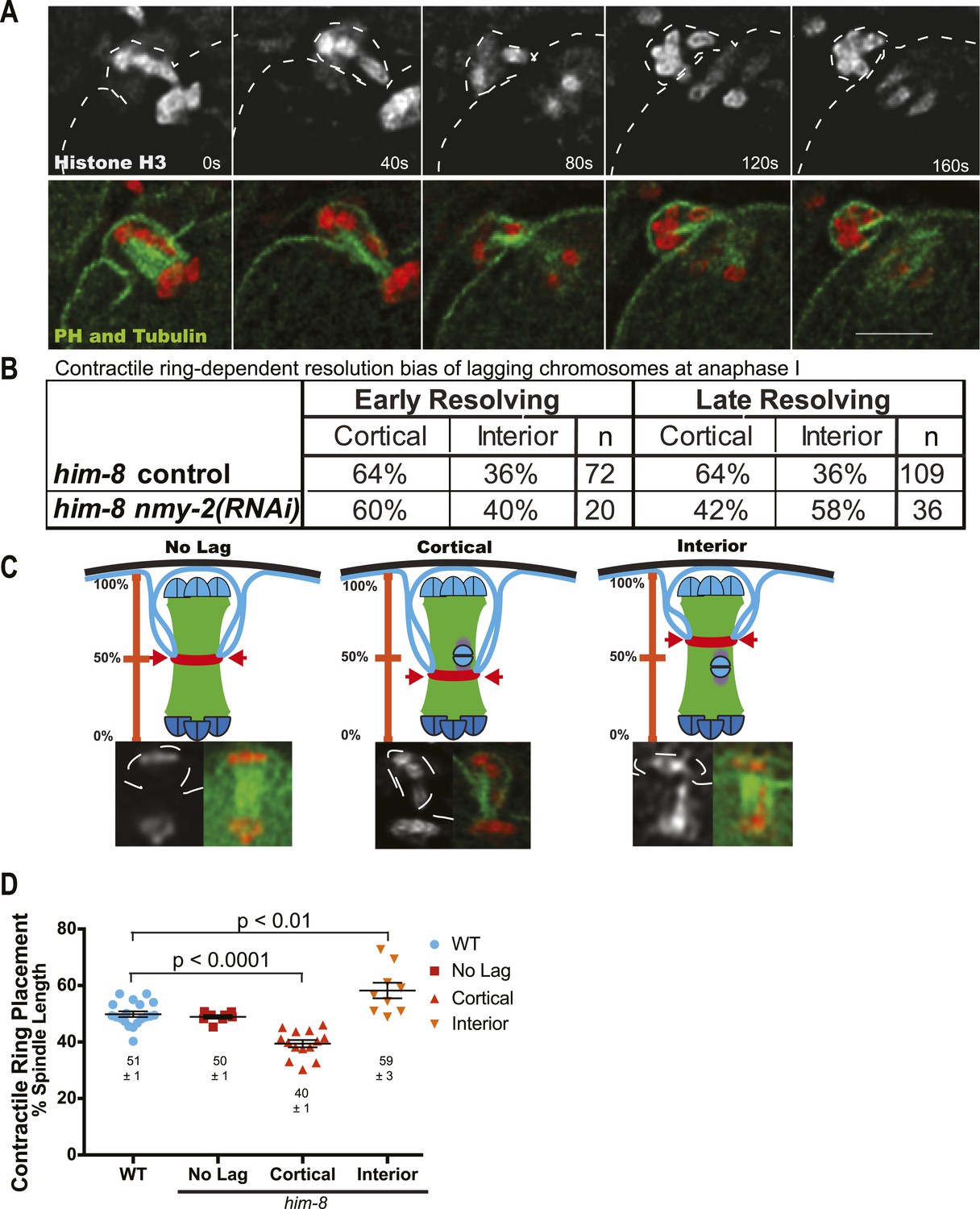
The contractile ring moves inward past the lagging chromosomes of him-8 embryos.
(A) Time-lapse sequence of anaphase I in a him-8 strain with GFP::PH, GFP::Tubulin, and mCherry::Histone H2B. The plasma membrane ingresses past the lagging chromosomes to engulf them in the polar body. (B) Fraction of him-8 anaphase I embryos in which a lagging chromosome eventually resolved toward the cortex or eventually resolved into the embryo (interior). Lagging univalents resolved more frequently toward the cortex during both early and late anaphase. Depletion of NMY-2, the myosin required for polar body formation, eliminated only the late anaphase bias. Pairwise two-tailed p values by Fisher's exact test: him-8 late vs him-8 nmy-2(RNAi) late = 0.02, him-8 early vs him-8 nmy-2(RNAi) early = 0.80, him-8 early vs him-8 late = 1.0, him-8 nmy-2(RNAi) early vs him-8 nmy-2(RNAi) late = 0.26. p values from Pearson's chi-squared test: him-8 late vs 50% = 0.003, him-8 nmy-2(RNAi) late vs 50% = 0.32, him-8 early vs 50% = 0.02, him-8 nmy-2(RNAi) early vs 50% = 0.38. (C) Top, diagram illustrating how the position of scission by the contractile ring along the pole-to-pole spindle axis was scored. Bottom, representative images from time-lapse sequences showing scission at different positions along the length of the spindle. (D) Average position of contractile ring scission along the pole-to-pole spindle axis in wild-type embryos and in him-8 embryos with no lagging chromosomes, lagging chromosomes that end up at the cortex (cortical), or lagging chromosomes that end up in the embryo (interior). Bar = 5 μm.
Figure 6
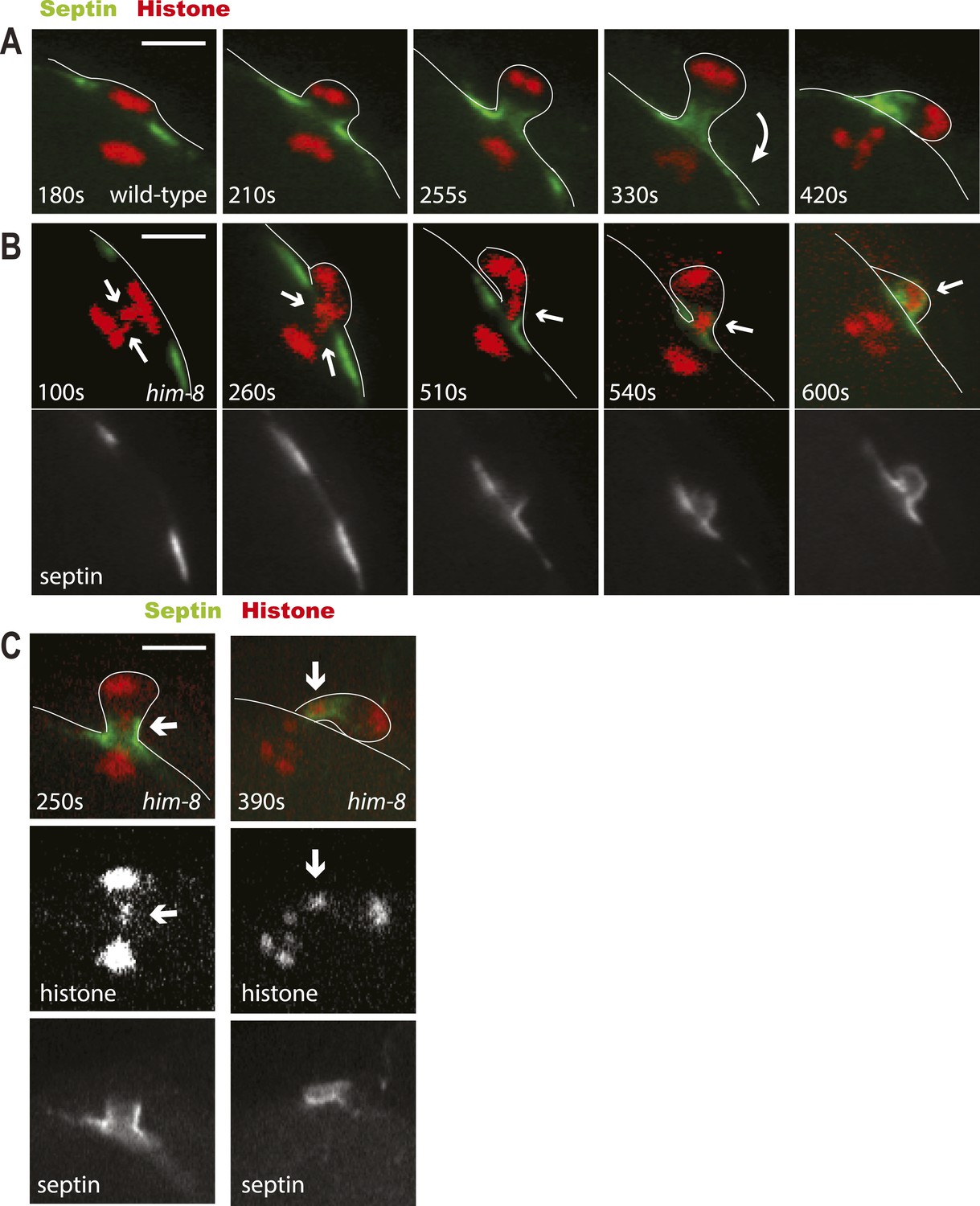
Lagging chromosomes are captured by the septin tube and expelled with polar bodies.
Time-lapse imaging of embryos expressing GFP::septin and mCherry::histone. (A) Time-lapse images of a living wild-type embryo undergoing anaphase I show the conversion of a flat washer–shaped contractile ring into a tube during formation of the first polar body. (B) Time-lapse images of a living him-8 embryo show two lagging chromosomes at anaphase I (arrows) as one moves into the polar body early on, while the second is trapped in the septin tube and is extruded with the first polar body. (C) 2 time points of a him-8 embryo during telophase I showing the lagging chromosome trapped in the septin tube. Bar = 4 μm. Times are from the onset of homolog separation.
Figure 7
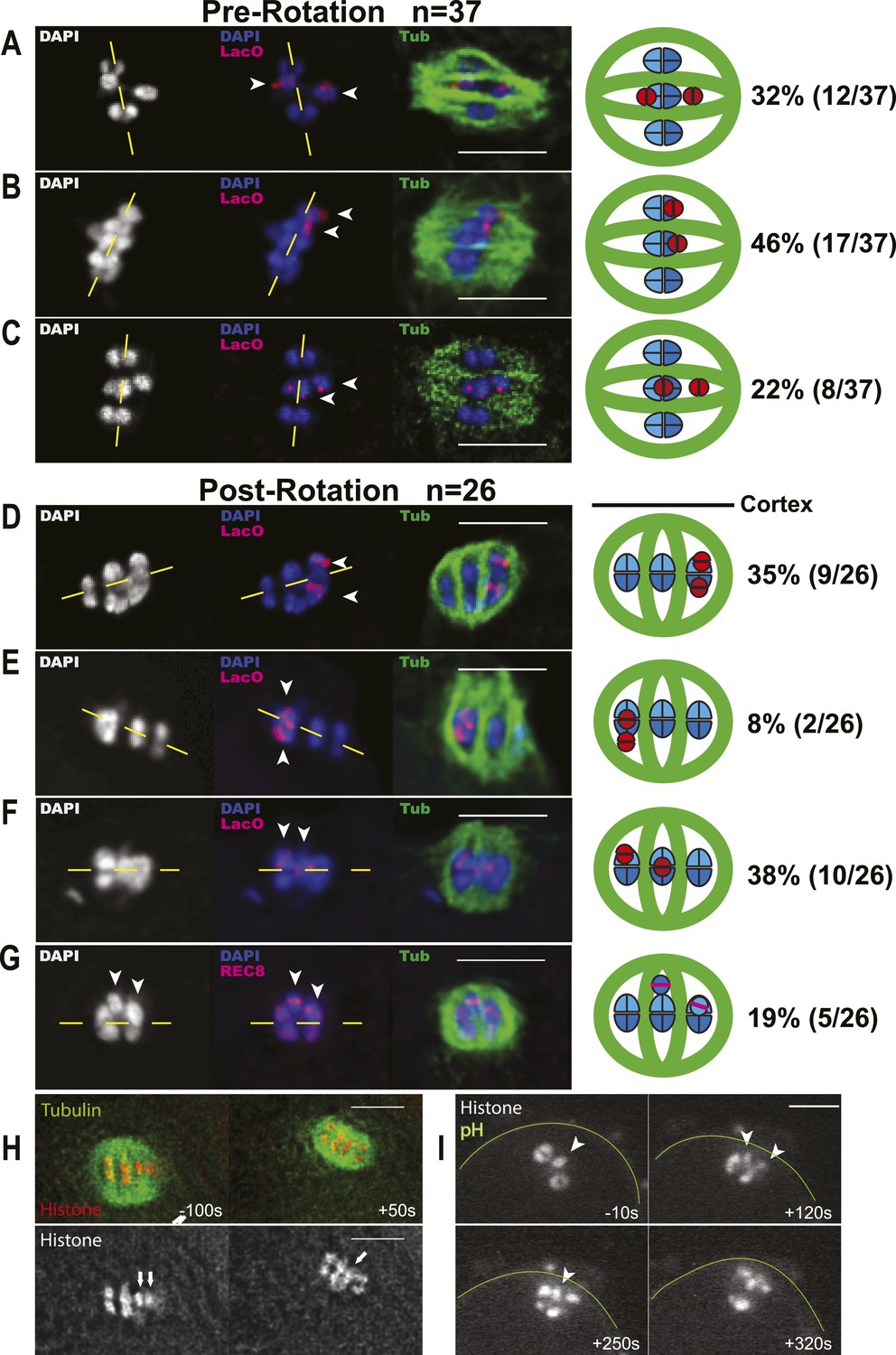
Early bias of univalent X chromosomes might occur at the metaphase to anaphase I transition.
Representative cartoon diagrams and Z projections from fixed embryos stained with DAPI, anti-tubulin antibody, and LacO(X) FISH probe. Cortex is at the top. (A–C) Both X univalents on metaphase I spindles that were shortened (5.3–7.2 μm spindle length) but still parallel to the embryo cortex were frequently (46%) aligned closer to the same spindle pole. (D–G) One or both univalents on MI spindles that had rotated but homologs had not yet separated were closer to the cortex and future polar body in 38 + 19% of embryos. Both univalents were never observed closer to the interior spindle pole. Yellow dashed lines indicate the metaphase plate. (H and I) Time-lapse images of two univalents (arrows in H) or one univalent (arrowhead in I) offset from the metaphase plate just before rotation of the univalent-proximal pole to the cortex. Time zero is initiation of spindle rotation. Bar = 5 μm.
Tables
Table 1
Enhancement of the segregation bias in him-8 mutants by mutations in the myosin phosphatase, mel-11
| Self-progeny counts | |||||
|---|---|---|---|---|---|
| Genotype | Temperature (°C) | % XO male | % XX hermaphrodite | % XXX Dpy | Total progeny |
| mel-11(sb55) unc-4 | 20 | 0.2 | 99.8 | NC | 1763 |
| mel-11(sb55) unc-4; him-8 | 20 | 49* | 51 | NC | 374 |
| unc-4; him-8 | 20 | 34 | 66 | NC | 1442 |
| mel-11(it126) unc-4 | 15 | 0.6 | 99 | NC | 790 |
| mel-11(it126) unc-4; him-8 | 15 | 58* | 38.6 | 3.4 | 873 |
| Ratio of nulloX ova/diploX ova calculated from progeny of cross with lon-2 males | |||||
|---|---|---|---|---|---|
| Maternal genotype | Temperature (°C) | # NulloX (ion male progeny) | # DiploX (dpy progeny) | Nullo/diplo | Total progeny |
| mel-11(it26) unc-4 | 25 | 1 | 0 | NA | 785 |
| mel-11(it26) unc-4; him-8 | 25 | 160 | 7 | 22.9 | 595 |
| unc-4; him-8 | 25 | 98 | 31 | 3.2 | 677 |
-
mel-11 increases the frequency of male progeny from him-8 mothers. mel-11(sb55) and mel-11(it26) worms produce high frequencies of dead embryos, which cannot be scored for sex at 25°C (Wissmann et al., 1999). Percent male (XO), hermaphrodite (XX), and dumpy (XXX) progeny from self-fertilizing mel-11, him-8, or him-8 mel-11 double mutant worms were therefore scored at 15°C and 20°C. Only progeny that developed to the L4 or adult stage were counted. *Two-tailed p < 0.0001 by binomial test compared with him-8 alone. 100% of mel-11(it26) self progeny die as embryos at 25°C, but this lethality is rescued by mel-11(+) sperm (Kemphues et al., 1988). The progeny of mel-11(it26) hermaphrodites crossed with lon-2 males could therefore be scored at 25°C. When lon-2(+) hermaphrodites are crossed with lon-2 males (lon-2 is a recessive X-linked marker), 50% of the ova will be fertilized by sperm with a single lon-2 X chromosome. Fertilization of a nulloX ova by a lon-2 X sperm will result in a lon-2 male. Fertilization of a diploX ova by a lon-2 X sperm will result in a XXX dumpy worm. Random segregation of the unpaired X chromosomes in him-8 would result in a ratio of nulloX/diplo X ova of 1.0. The mel-11; him-8 double mutant showed a sevenfold increase in the ratio of nullo/diploX ova relative to him-8 alone, indicating an increased efficiency of eliminating maternal unpaired X chromosomes.
Additional files
-
Supplementary file 1
C. elegans strains used in this study.
- https://doi.org/10.7554/eLife.06056.014
-
Supplementary file 2
Z-stack of XC FISH on XXX wild-type metaphase plate in meiosis I. 16-bit 3-channel TIFF can be opened using FIJI or basic ImageJ (http://fiji.sc/Downloads). Data shown are a z-stack acquired with 300 nm steps through a meiosis I metaphase spindle. Chromosomes are shown in blue (DAPI), tubulin antibodies label the spindle in green, and the XC FISH probe labels X chromosomes (2 present) in red. Channels can be split for individual analysis using the channel splitter (Image > Colors > Split channels) or can be hidden using the channels tool (Image > Colors > Channels tool).
- https://doi.org/10.7554/eLife.06056.015
-
Supplementary file 3
Z-stack of XC FISH on XXX wild-type metaphase plate in meiosis II. 16-bit 3-channel TIFF can be opened using FIJI or basic ImageJ (http://fiji.sc/Downloads). Data shown are a z-stack acquired with 300 nm steps through a meiosis II metaphase spindle. Chromosomes and the first polar body, which is on the top, are shown in blue (DAPI), tubulin antibodies label the spindle in green, and the XC FISH probe labels X chromosomes (1 present on the spindle) in red. Channels can be split for individual analysis using the channel splitter (Image > Colors > Split channels) or can be hidden using the channels tool (Image > Colors > Channels tool).
- https://doi.org/10.7554/eLife.06056.016
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
The asymmetry of female meiosis reduces the frequency of inheritance of unpaired chromosomes
eLife 4:e06056.
https://doi.org/10.7554/eLife.06056




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}