Enteric pathogens deploy cell cycle inhibiting factors to block the bactericidal activity of Perforin-2
- Miller School of Medicine, University of Miami, United States
Figures
Figure 1
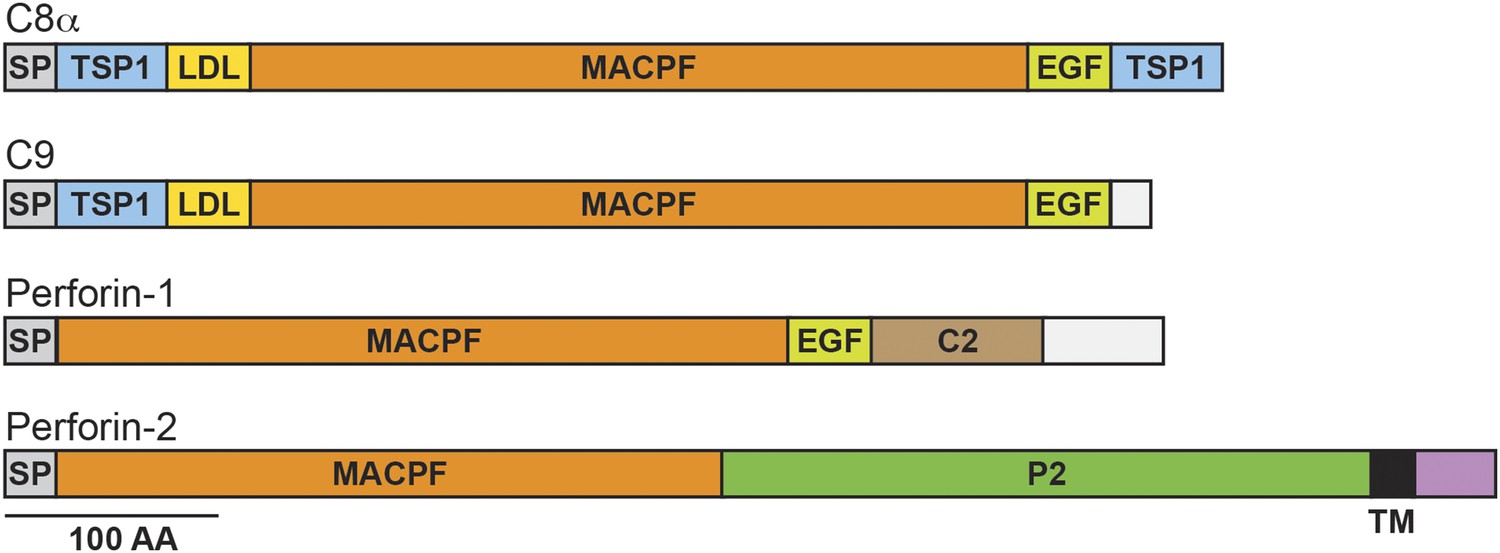
Domain organization of human C8a, C9, Perforin-1, and Perforin-2.
The pore forming components of complement (C8α and C9), Perforin-1, and Perforin-2 all contain membrane attack complex perforin (MACPF) domains and amino-terminal signal peptides (SPs). MACPF domains are also present in other components of complement (C6, C7, and C8β). The presence of a MACPF domain within Perforin-2 suggest that it is also a mediator of innate immunity. Unlike C6-C9 and Perforin-1, Perforin-2 is predicted to be an integral membrane protein because it alone contains a membrane spanning alpha helix (TM) followed by a short cytosolic tail. An additional distinguishing feature of Perforin-2 is the P2 domain which is of unknown function but conserved amongst Perforin-2 orthologs. Domain architecture was retrieved from UniProt entries P07357, P02748, P14222, and Q2M385. TSP1, thrombospondin type-1 repeat; LDL, low-density lipoprotein receptor class A repeat; EGF, epidermal growth factor-like domain; C2, calcium-dependent phospholipid binding domain.
Figure 2

Perforin-2 co-immunoprecipitates with CRL subunits and is ubiquitylated.
RAW 264.7 cells were transfected with Perforin-2-GFP or GFP expression constructs and stimulated with IFN-γ prior to immunoprecipitation with anti-GFP. (A) Western blots probed with the indicated antibodies reveal that the NEDD8 E2 ligase UBC12, CRL cullin scaffold CUL1, and substrate receptor F-box protein βTrCP specifically coimmunoprecipitate with Perforin-2. (B) Additional western blots reveal that IFN-γ and LPS, but not IFN-γ alone, stimulate ubiquitylation of Perforin-2. P2, Perforin-2; IP, immunoprecipitates; WCL, whole cell lysates.
Figure 3
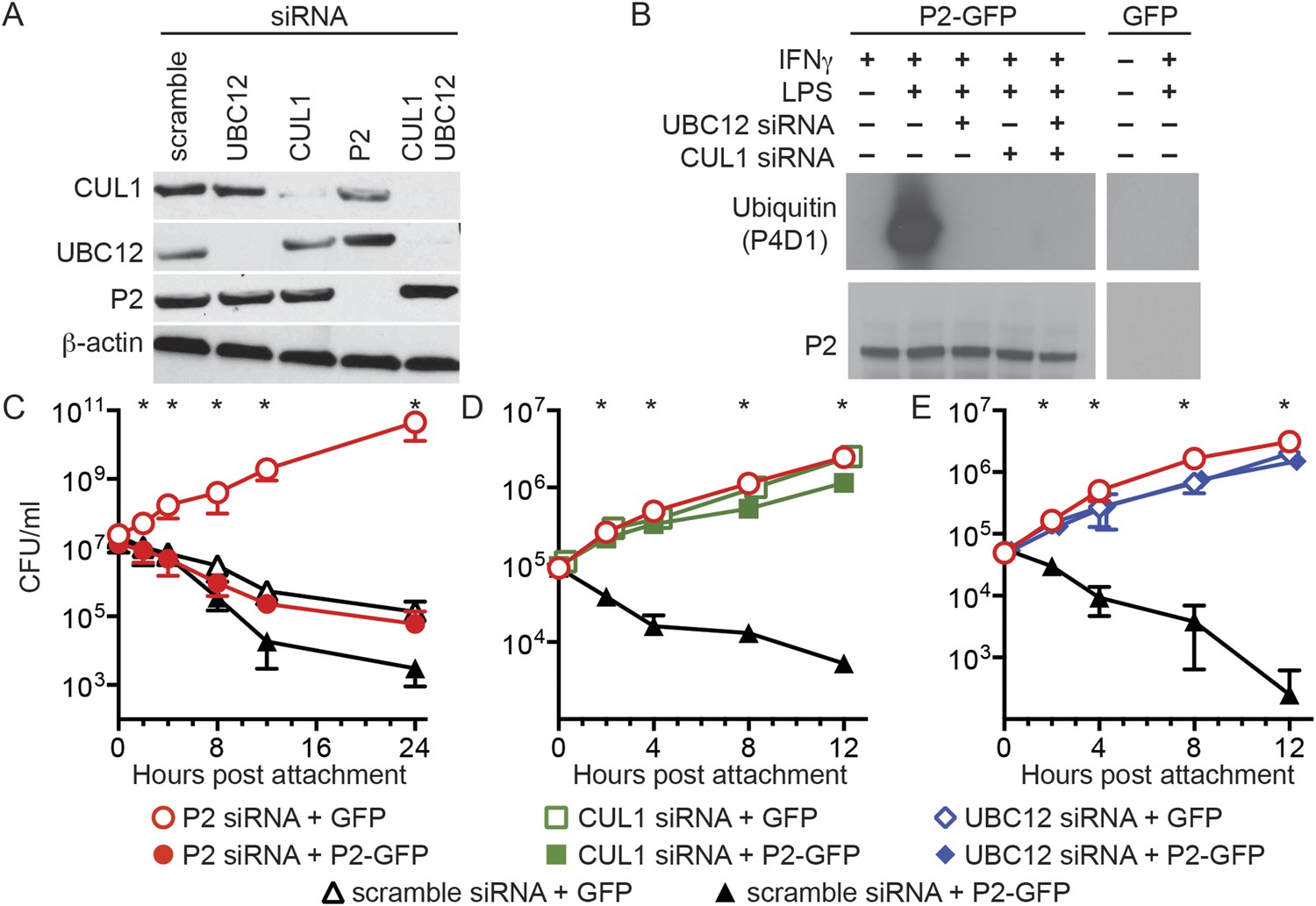
Perforin-2 activity is dependent upon its ubiquitylation by a CRL complex.
(A) siRNA efficiently and specifically knocks down expression of CUL1, UBC12, and Perforin-2 in IFN-γ stimulated CMT93 cells. (B) Western blots of immunoprecipitates from transfected CMT93 cells probed with the indicated antibodies demonstrate that Perforin-2 is ubiquitylated in cells stimulated with IFN-γ and LPS and that ubiquitylation is dependent upon UBC12 and CUL1. (C–E) In contrast to scramble siRNA, siRNA knockdown of Perforin-2, CUL1, or UBC12 in CMT93 cells promotes survival of extracellular E2348/69, an enteropathogenic Escherichia coli (EPEC) strain that does not express cycle inhibiting factor (Cif). Solid and open symbols denote cotransfection with Perforin-2-GFP or GFP expression plasmids, respectively. *Statistically significant (p < 0.05) differences between (C) Perforin-2 siRNA + GFP and the other three conditions, (D, E) scramble siRNA + Perforin-2-GFP and the other three conditions by one-way ANOVA with Bonferroni multiple-comparisons post-hoc test; n ≥ 3. P2, Perforin-2.
Figure 4
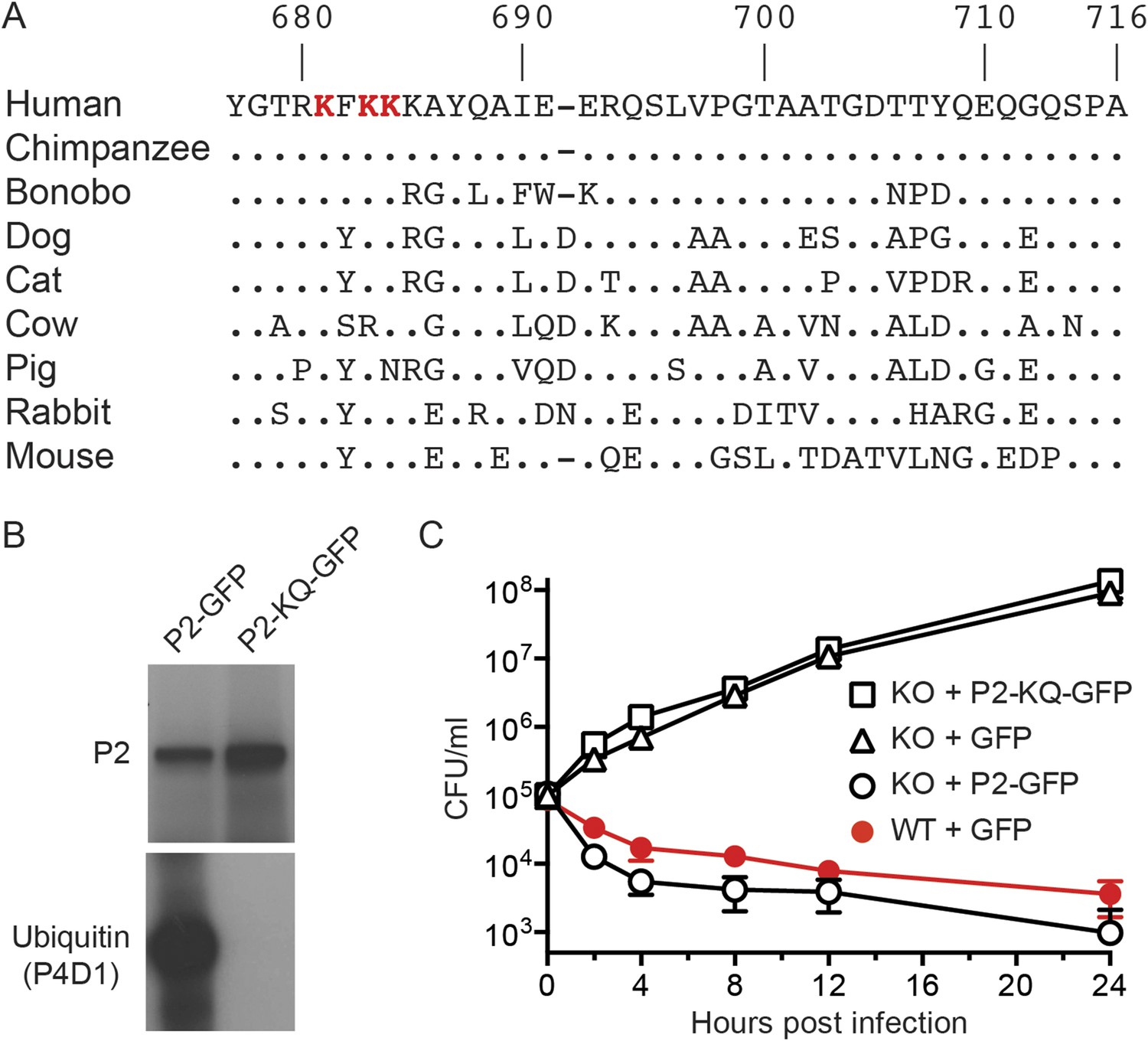
Conserved lysines within the carboxy-terminal domain of Perforin-2 are required for its ubiquitylation and bactericidal activity.
(A) Alignment of the predicted cytosolic domains of Perforin-2 (P2) molecules from select mammalian species. Dots denote identity to human Perforin-2. The three most conserved lysine residues are highlighted in red. Numbering is relative to human Perforin-2, GenBank accession number AAI12231. (B) Site-directed mutagenesis was used to mutate the three conserved lysines to glutamine residues in murine Perforin-2. Western blots of Perforin-2-GFP or Perforin-2-KQ-GFP expressed in CMT93 cells stimulated with IFN-γ and LPS demonstrate that both fusion proteins are expressed. However, only Perforin-2-GFP is ubiquitylated. (C) Murine embryonic fibroblasts (MEFs) isolated from wild-type and Perforin-2 −/− (KO) embryos were transfected with GFP, Perforin-2-GFP, or Perforin-2-KQ-GFP expression plasmids. Transfected cells were induced with IFN-γ ca. 24 hr before infection with Salmonella typhimurium. The results demonstrate that Perforin-2-GFP, but not the K-to-Q mutant nor GFP, restored killing in KO MEFs to wild-type levels. The differences between KO MEFs transfected with Perforin-2-GFP and Perforin-2-KQ-GFP or GFP are statistically significant, p < 0.05, at hours 2 through 24 as determined by one-way ANOVA with Bonferroni post-hoc multiple comparisons; n ≥ 3.
Figure 5 with 1 supplement
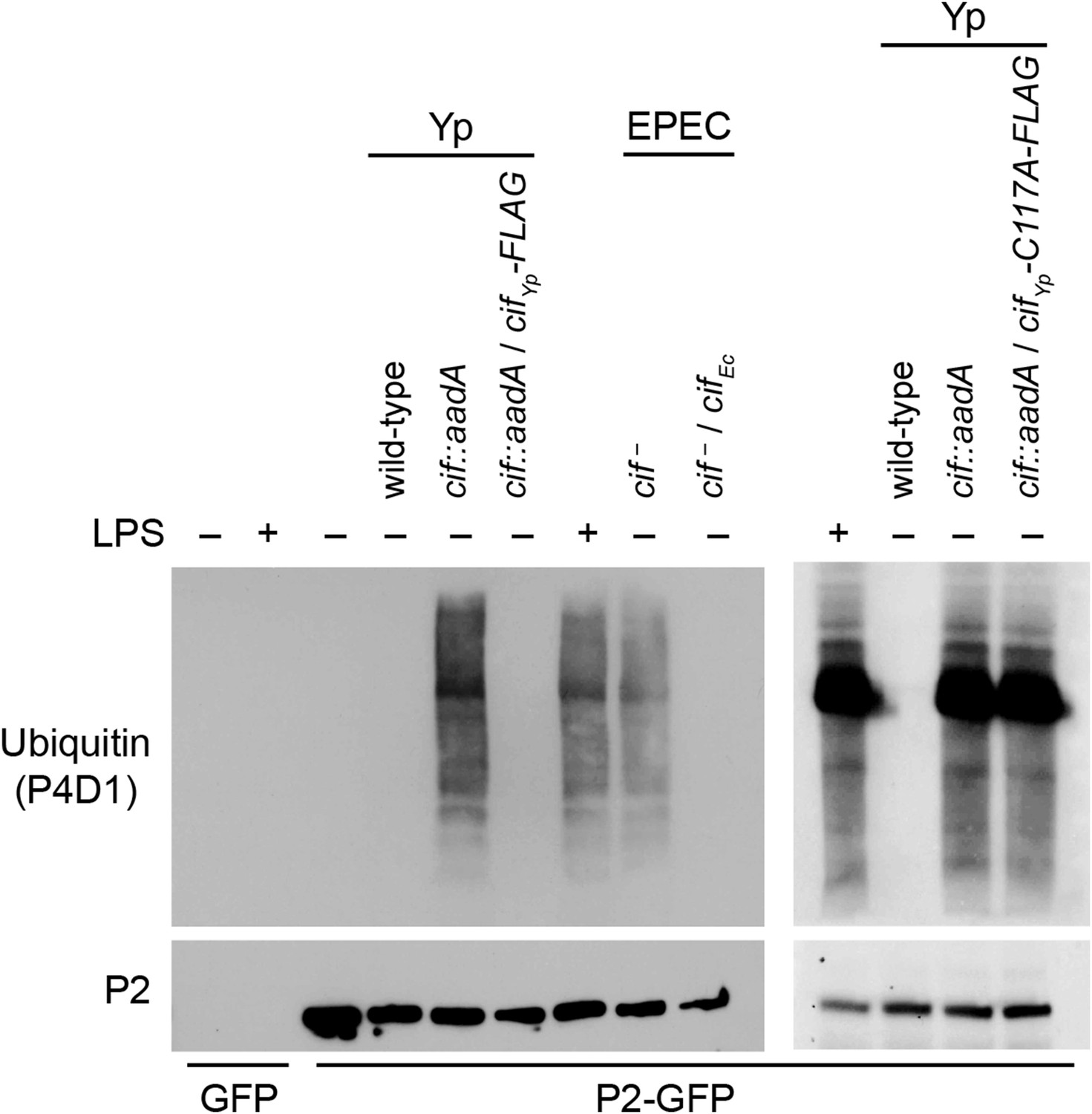
Cif blocks ubiquitylation of Perforin-2.
CMT93 cells were transfected with GFP or Perforin-2-GFP expression plasmids and stimulated with IFN-γ. As indicated, some transfected cells were also stimulated with LPS, infected with Yersinia pseudotuberculosis (Yp), or EPEC strain E2348/69. Unlike Y. pseudotuberculosis or other EPEC strains, E2348/69 harbors a defective cif locus. Species specific expression plasmids were used to complement cif mutants. Immunoprecipitates were then separated by SDS-PAGE and probed with the indicated antibodies. CifYp-C117A-FLAG denotes a point mutation within the enzyme's catalytic triad, conserved amongst Cif proteins, that is essential for deamidation of NEDD8. The inability of CifYp-C117A-FLAG to block ubiquitylation of Perforin-2 is not due to a lack of expression (Figure 5—figure supplement 1).
Figure 5—figure supplement 1
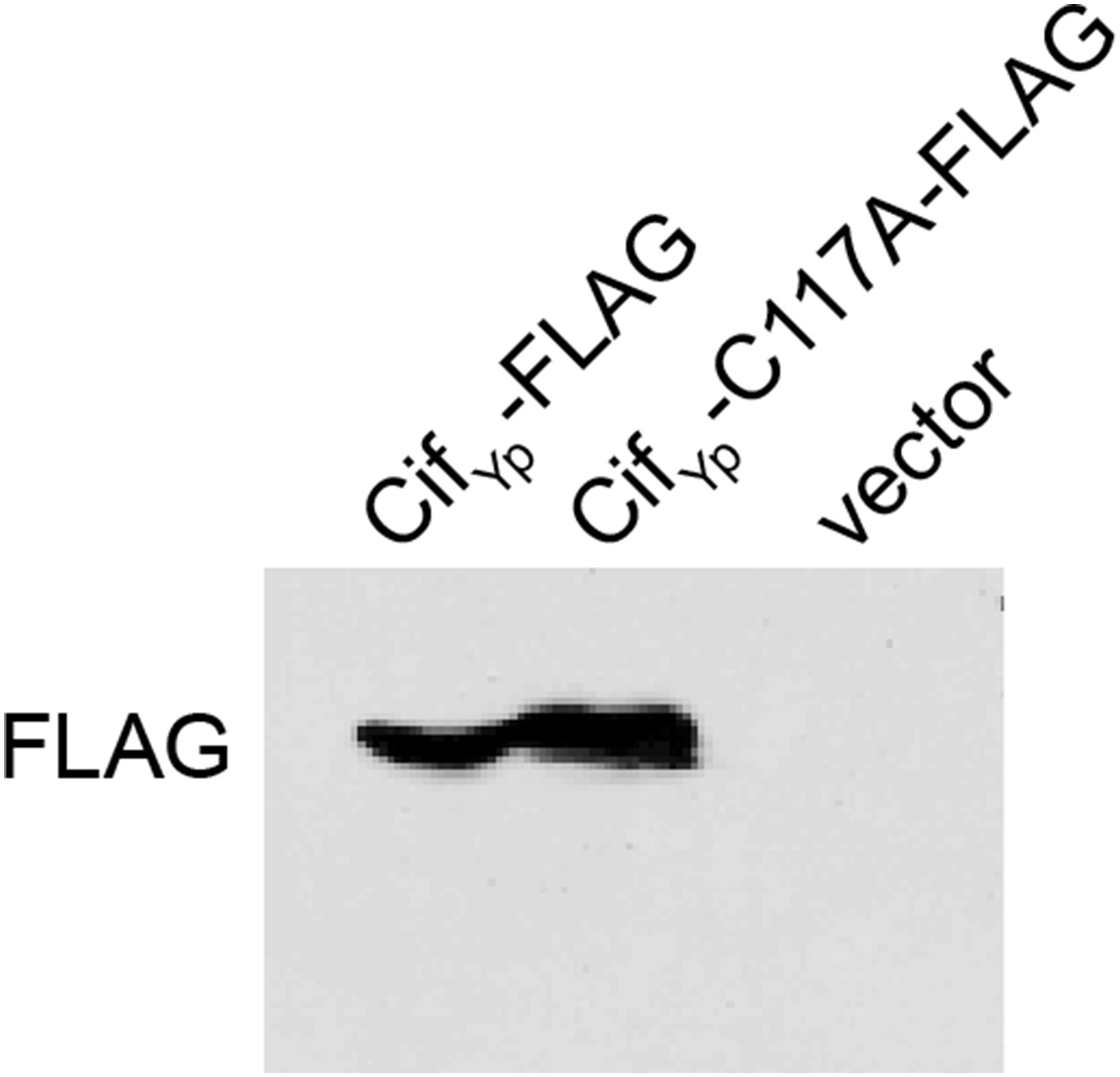
CifYp expression.
Western blot demonstrating that both CifYp-FLAG and CifYp-C117A-FLAG are expressed in Y. pseudotuberculosis cif::aadA. Cloning vector pFLAG-CTC was included as a negative control.
Figure 6

The bacterial effector protein Cif blocks the bactericidal activity of Perforin-2.
Caco-2 cells, a human intestinal epithelial cell line, were transfected with Perforin-2 specific or scramble siRNAs. The cells were subsequently infected with EPEC strain E2348/69, which carries a naturally disrupted cif gene, transformed with a (A) CifEc expression plasmid or (B) vector control. Alternatively CMT93 cells, a murine epithelial cell line, were cotransfected with the indicated siRNA and RFP or Perforin-2-RFP expression plasmids. Transfected cells were subsequently infected with (C) wild-type Y. pseudotuberculosis, (D) a cif mutant, (E) the mutant transformed with a CifYp-FLAG expression plasmid, or (F) vector control. All mammalian cells were activated with IFN-γ for 24 hr prior to infection. *p < 0.05 by Student's t-test, n ≥ 3.
Figure 7

Enzymatically inactive Cif cannot block Perforin-2-dependent killing.
CMT93 cells were transfected with the indicated siRNAs and RFP or Perforin-2-RFP expression plasmids. Following stimulation with IFN-γ, transfected cells were infected with a Y. pseudotuberculosis cif::aadA mutant transformed with a CifYp-C117A-FLAG expression plasmid. The C117A point mutation within the enzyme's conserved catalytic triad abolishes its ability to block ubiquitylation of Perforin-2. *Statistically significant (p < 0.05) difference between Perforin-2 siRNA + RFP and scramble siRNA + RFP or Perforin-2 siRNA + Perforin-2-RFP by one-way ANOVA with Bonferroni multiple-comparisons post-hoc test; n ≥ 3.
Figure 8
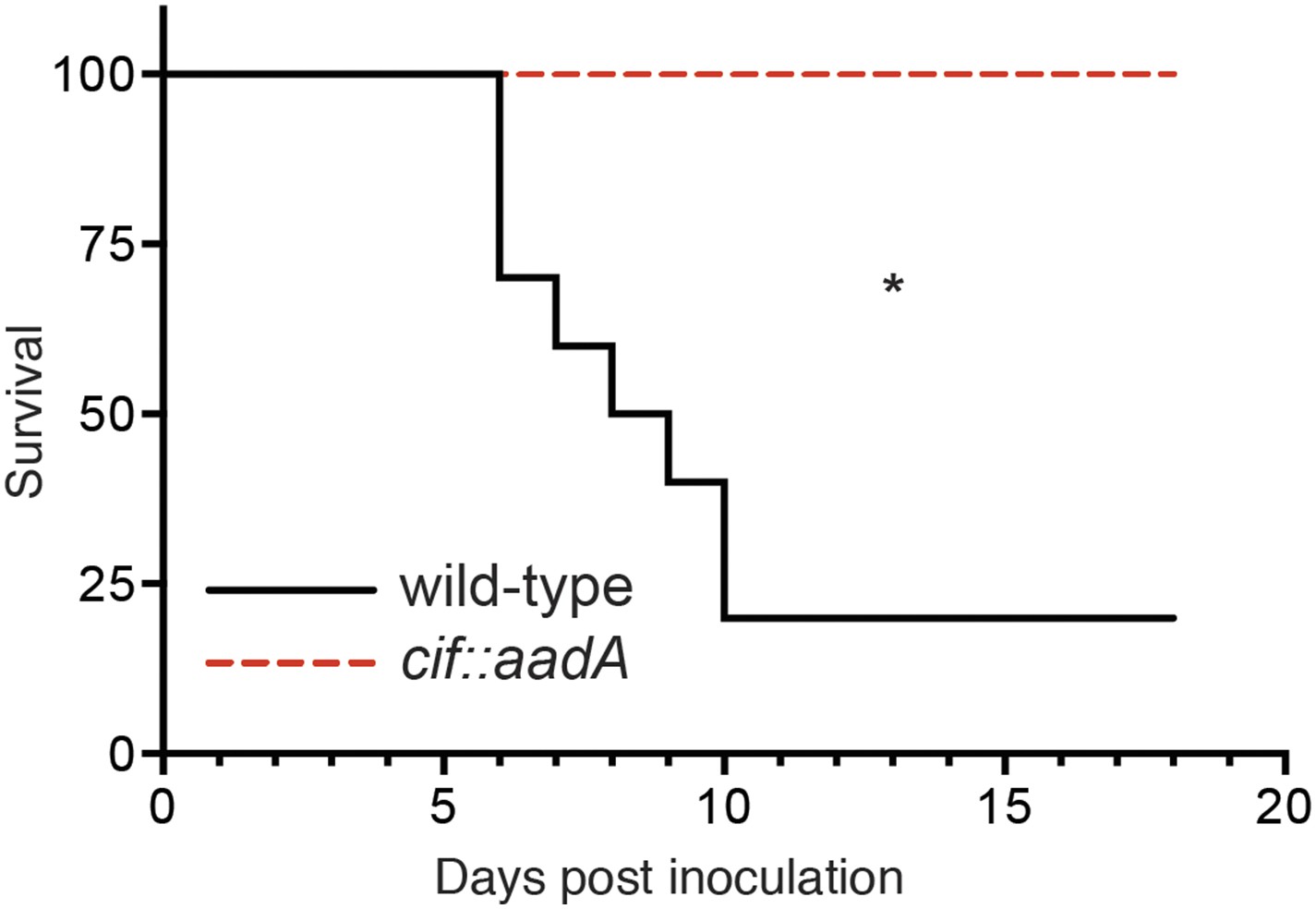
Cif enhances pathogenicity in vivo.
Survival curves of C57Bl/6 mice inoculated orogastrically with 108 CFU of wild-type Y. pseudotuberculosis or an isogenic cif::aadA mutant. Animals were weighed daily and euthanized if weight loss exceeded 20%. *p < 0.05 by log-rank (Mantel–Cox) test, n = 9–10 mice per group.
Figure 9
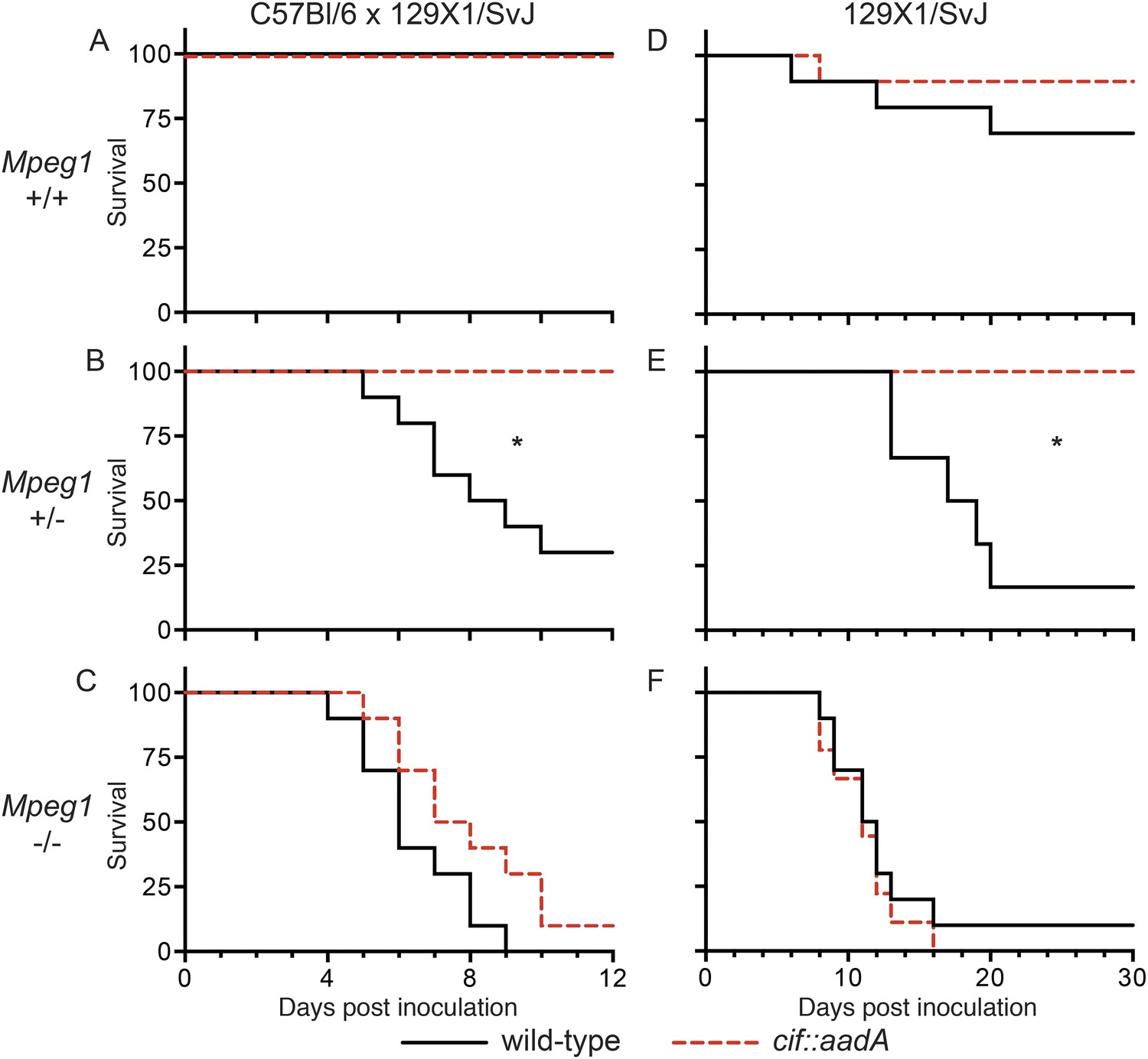
Bacterial inhibitors of neddylation abolish the bactericidal activity of Perforin-2 in vivo.
Survival curves for two different lineages of Mpeg1 +/+, +/−, and −/− mice following orogastric inoculation with 106 CFU of wild-type Y. pseudotuberculosis or an isogenic cif::aadA mutant. (A–C) n = 6–10 mice per group. (D–F) n = 22–28 mice per group. *p < 0.05 by log-rank (Mantel–Cox) test.
Figure 10
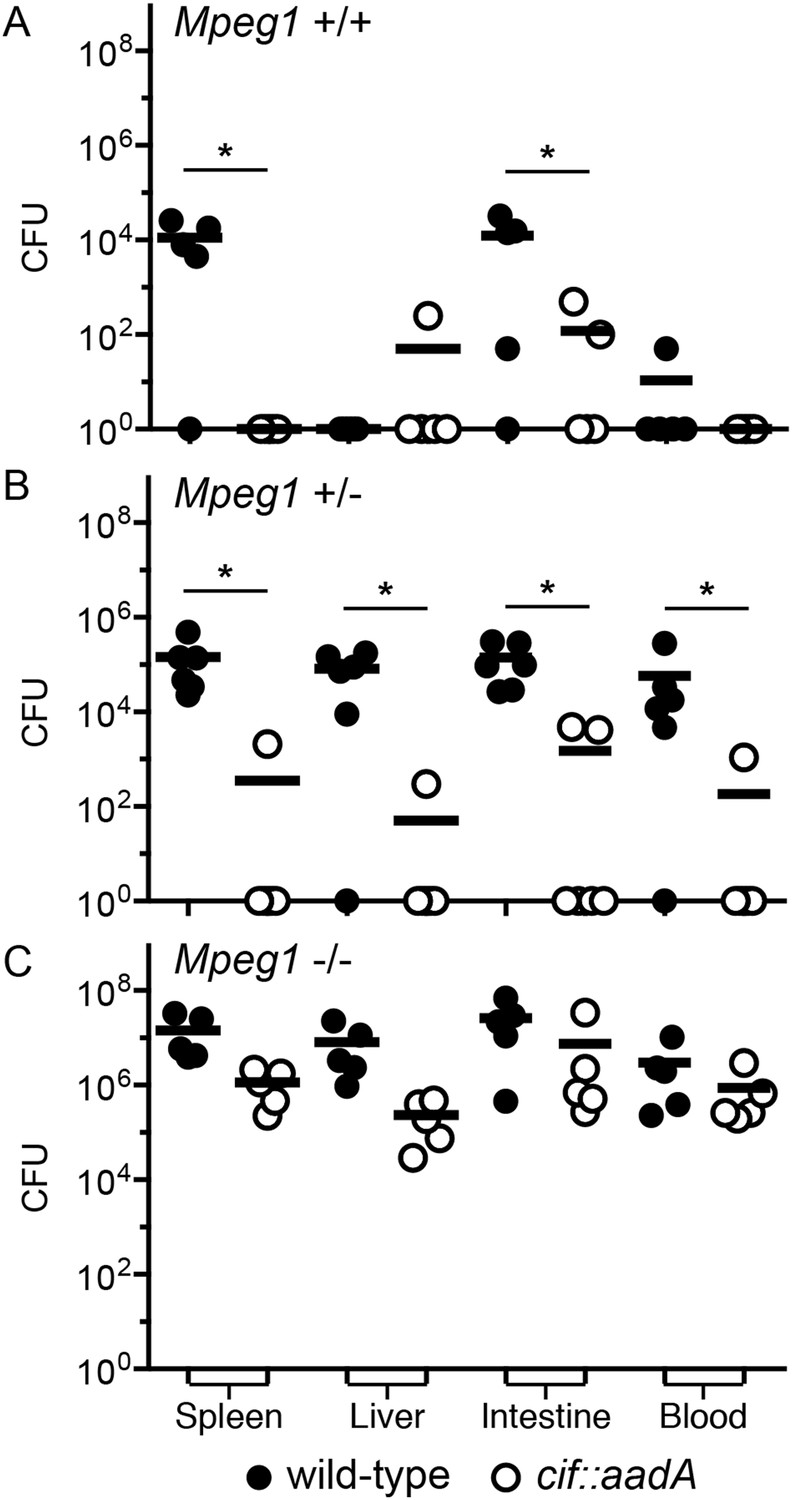
Cif diminishes the bactericidal activity of Perforin-2 in vivo.
Representative organ loads of C57Bl/6 × 129X1/SvJ Mpeg1 (A) +/+, (B) +/− and (C) −/− mice infected with Cif+ or Cif− Y. pseudotuberculosis. Animals were sacrificed 10 days after orogastric inoculation with 106 CFU. Samples were normalized by organ weight and each experiment was repeated twice. Horizontal lines represent the mean. Statistical analysis was performed by the nonparametric Kolmogorov–Smirnov test with Dunn's multiple comparison test; *p < 0.05, n ≥ 5.
Figure 11 with 1 supplement
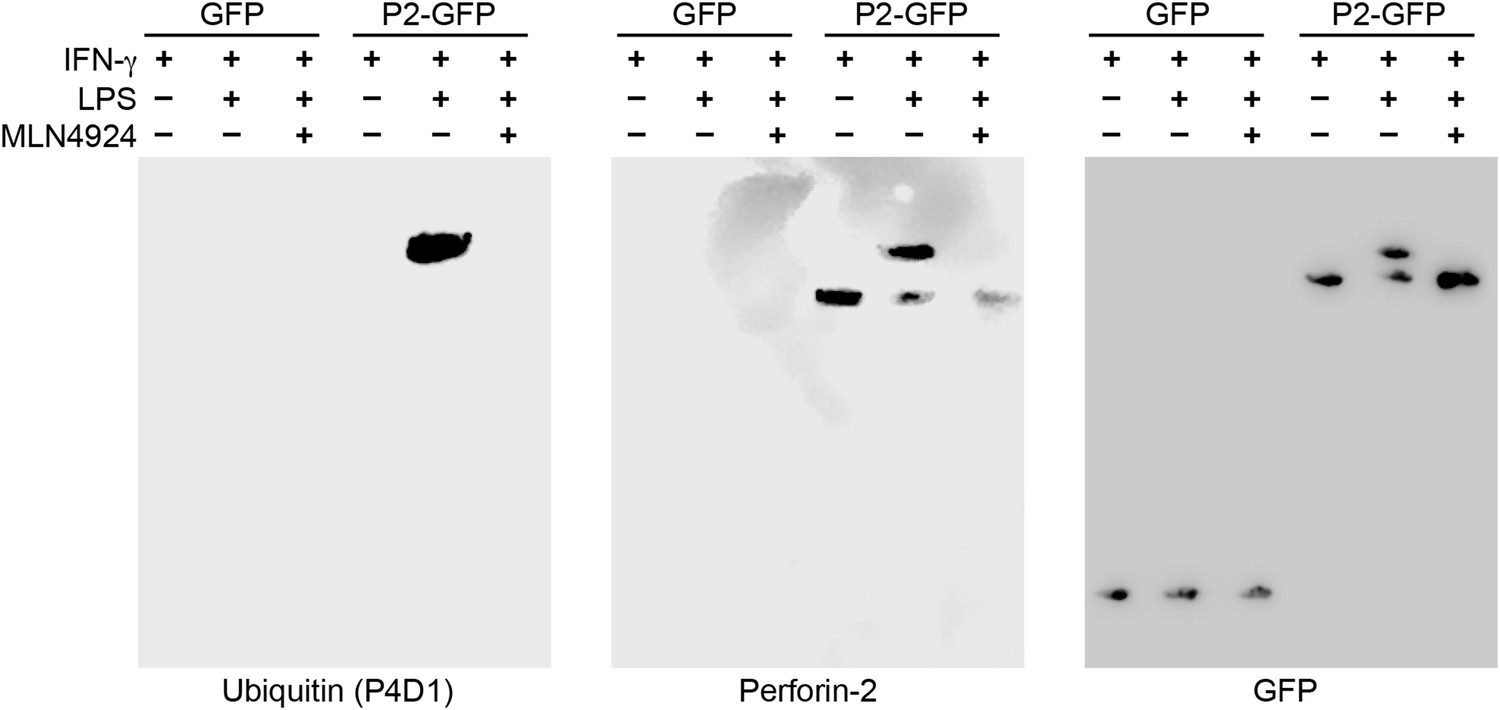
Perforin-2 is monoubiquitylated.
Mpeg1 knockout MEFs were transfected with GFP or Perforin-2-GFP expression plasmids and stimulated with IFN-γ in the presence or absence of LPS. Some cultures were also treated with MLN4924, a small molecule inhibitor of neddylation. The resulting GFP immunoprecipitates were probed with the indicated antibodies in Western blots. Consistent with previous results ubiquitylation of Perforin-2 was dependent upon LPS and neddylation when the immunoprecipitates were probed with a non-linkage specific ubiquitin antibody. When the same immunoprecipitates were probed with antibodies specific for K48 or K63 linkages, ubiquitylation was not detected. From additional Western blots we estimated that ubiquitylation increases the mass of Perforin-2-GFP by ∼10 kDa (Figure 11—figure supplement 1). These results indicate the Perforin-2 is a monoubiquitylated protein.
Figure 11—figure supplement 1
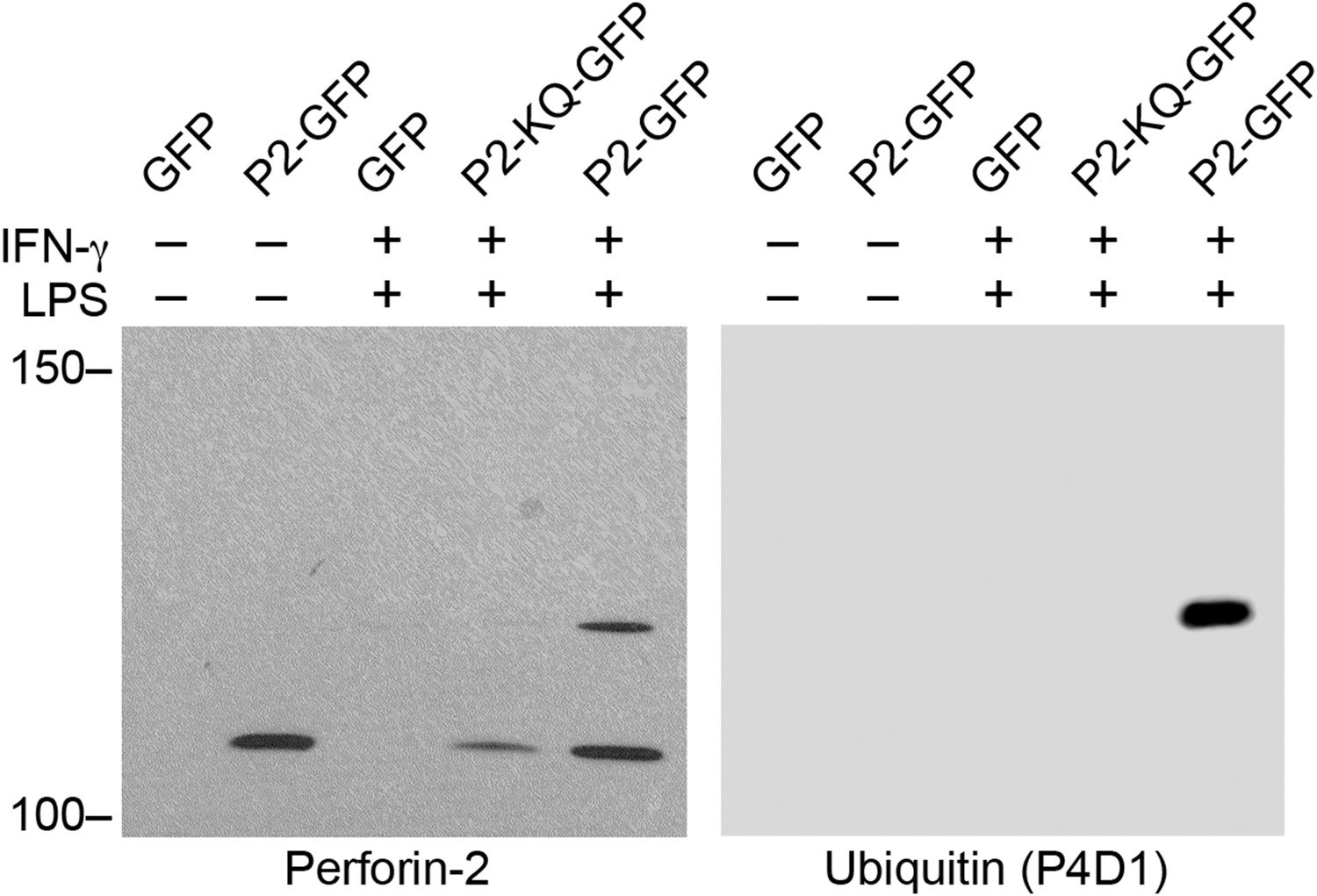
Monoubiquitylation of Perforin-2.
CMT93 cells were transfected with expression plasmids and stimulated with IFN-γ and LPS as indicated. GFP immunoprecipates were then separated by SDS-PAGE and probed with a Perforin-2 antibody. The blot was then stripped and probed for ubiquitin. Ubiquitin increases the apparent mass of Perforin-2-GFP by ∼10 kDa which, given some uncertainty in mass estimation, is consistent the addition of a ubiquitin monomer (8.5 kDa); that is, monoubiquitylation. Qualitative differences between the two images is the result of different imaging methods: The left image was collected by exposure to film while the right image was collected on a digital workstation.
Figure 12
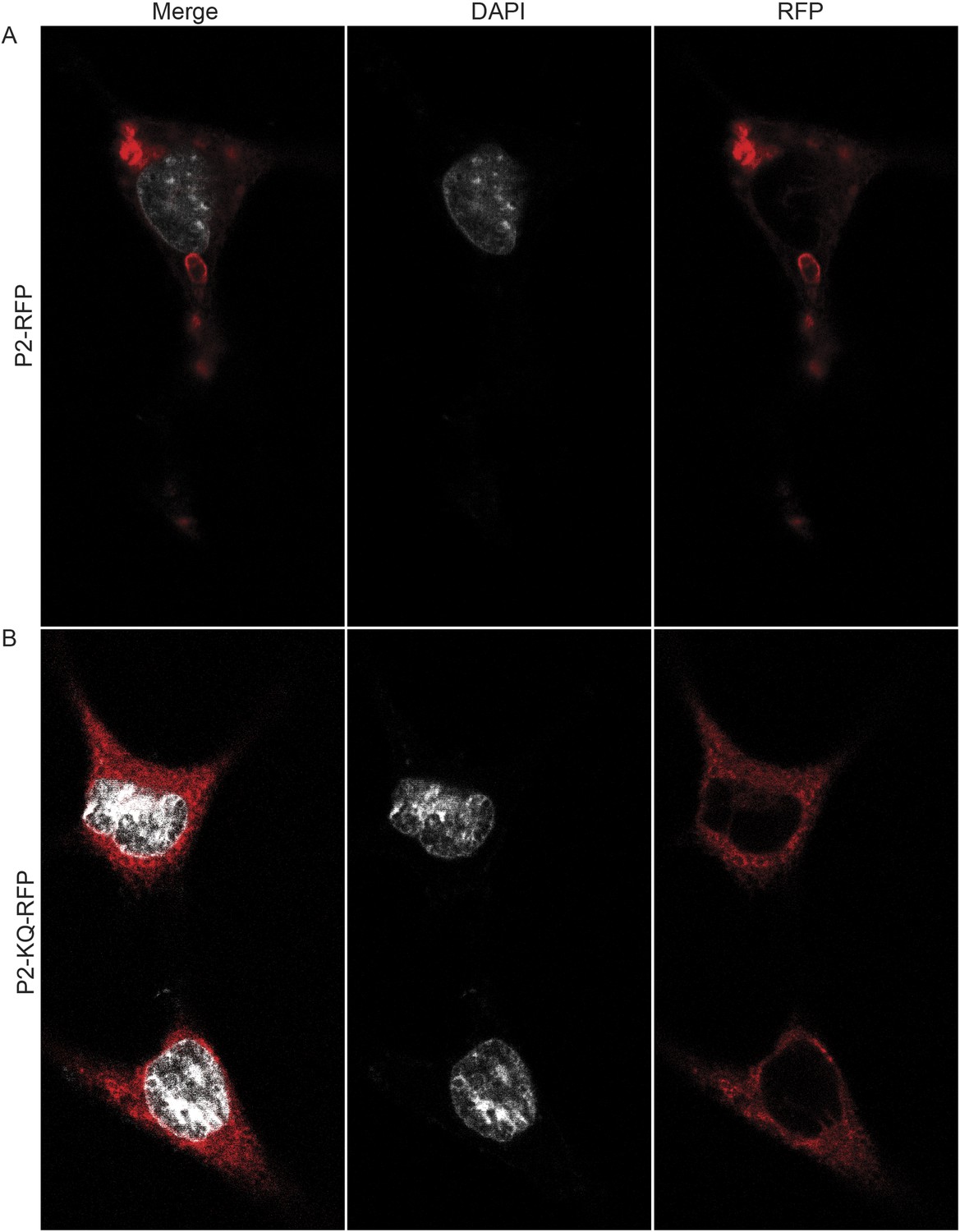
Ubiquitylation determines the subcellular distribution of Perforin-2.
Mpeg1 knockout MEFs were transfected with (A) Perforin-2-RFP or (B) Perforin-2-KQ-RFP expression plasmids, the latter of which carries K-to-Q mutations of conserved lysines in the cytosolic tail of Perforin-2 that abolish ubiquitylation. Transfected cells were stimulated with IFN-γ for 24 hr prior to addition of LPS. Cells were fixed within 15 min of LPS addition and counter stained with DAPI. Images were acquired on a Leica confocal microscope with a 63× objective.
Figure 13 with 2 supplements
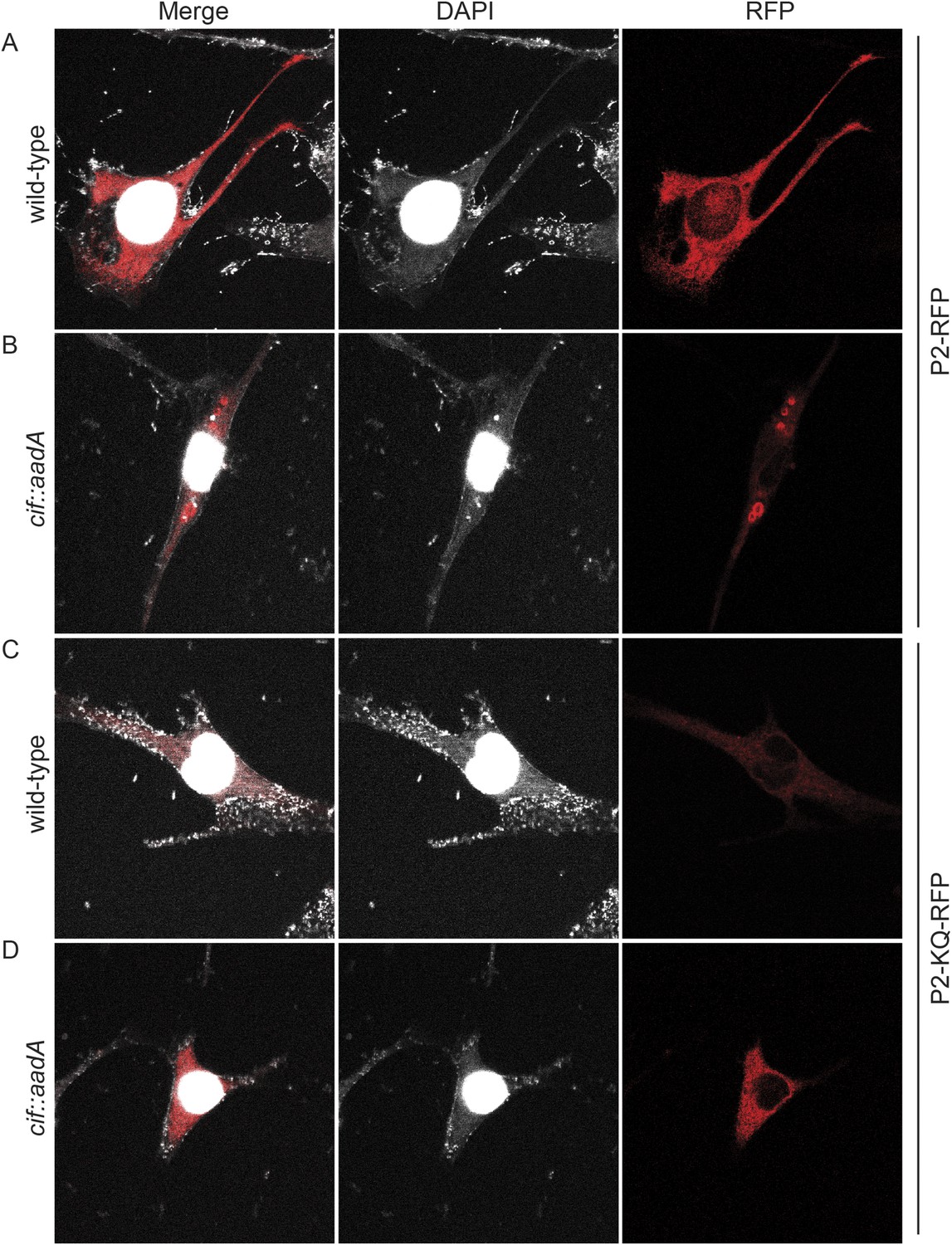
Cif blocks trafficking of Perforin-2.
Mpeg1 knockout MEFs were transfected with Perforin-2-RFP or Perforin-2-KQ-RFP expression plasmids and stimulated with IFN-γ for 24 hr prior to infection with (A, B) wild-type Y. pseudotuberculosis or (C, D) an isogenic cif::aadA mutant. Cells were fixed 15 min after infection and counter stained with DAPI. Similar results were obtained when transfected CMT93 cells were infected with Cif+ and Cif− bacteria expressing GFP (Figure 13—figure supplements 1, 2). Images were acquired on a Leica confocal microscope with a 63× objective.
Figure 13—figure supplement 1
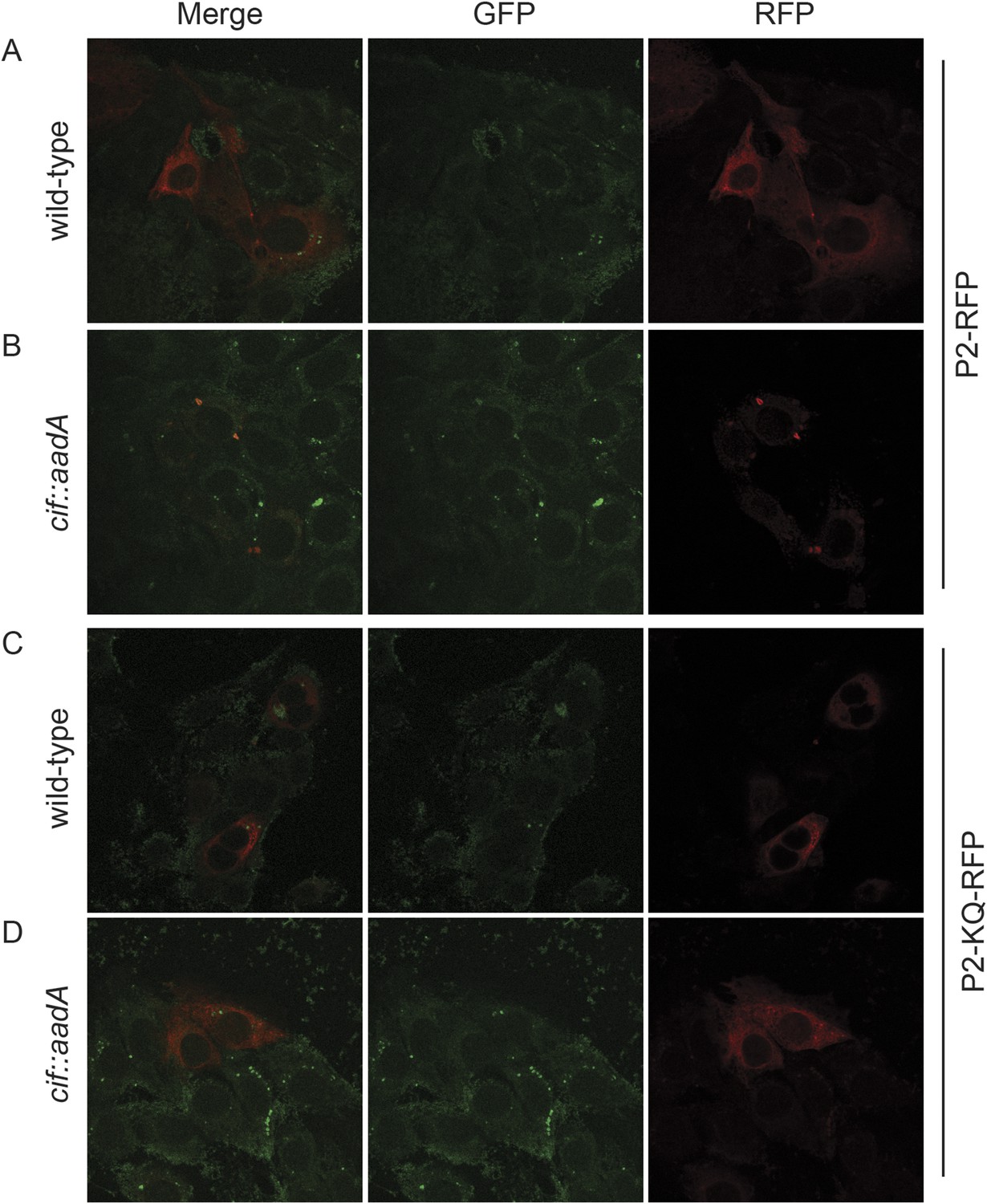
Perforin-2 relocalizes from perinuclear to punctate bodies upon ubiquitylation.
CMT93 cells, a murine rectal epithelial cell line, were transfected with murine Perforin-2-RFP or Perforin-2-KQ-RFP expression plasmids and induced with IFN-γ overnight. Cells were then infected with (A, B) wild-type Y. pseudotuberculosis or (C, D) an isogenic mutant that does not express Cif. Cells were fixed within 15 min of infection. Images were acquired on a Leica confocal microscope with a 63× objective.
Figure 13—figure supplement 2
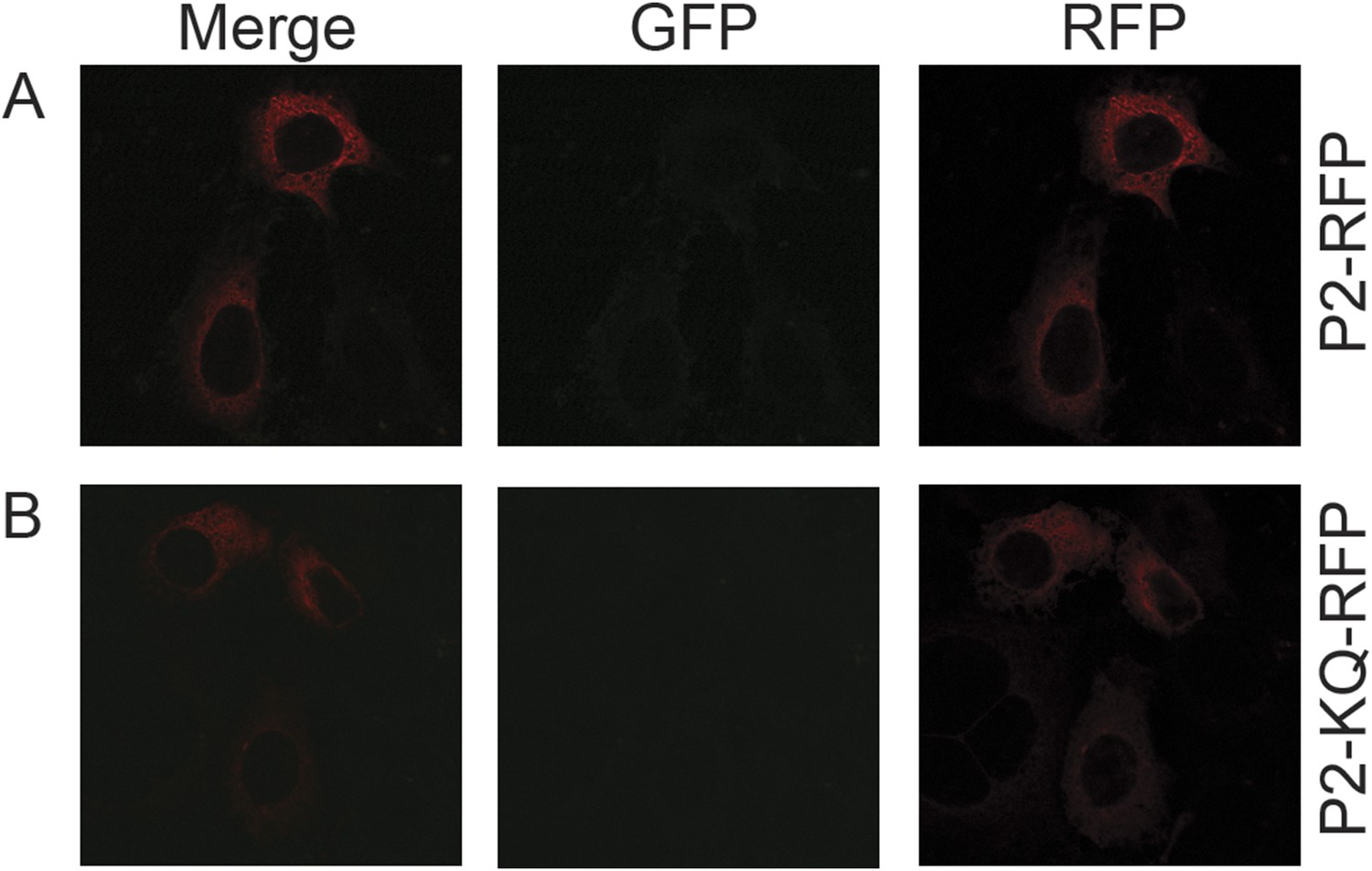
Perforin-2 perinuclear localization in noninfected CMT93s.
CMT93 cells were transfected with (A) murine Perforin-2-RFP or (B) murine Perforin-2-KQ-RFP and induced overnight with IFN-γ. Images were taken on a Leica confocal microscope with a 63× objective.
Figure 14
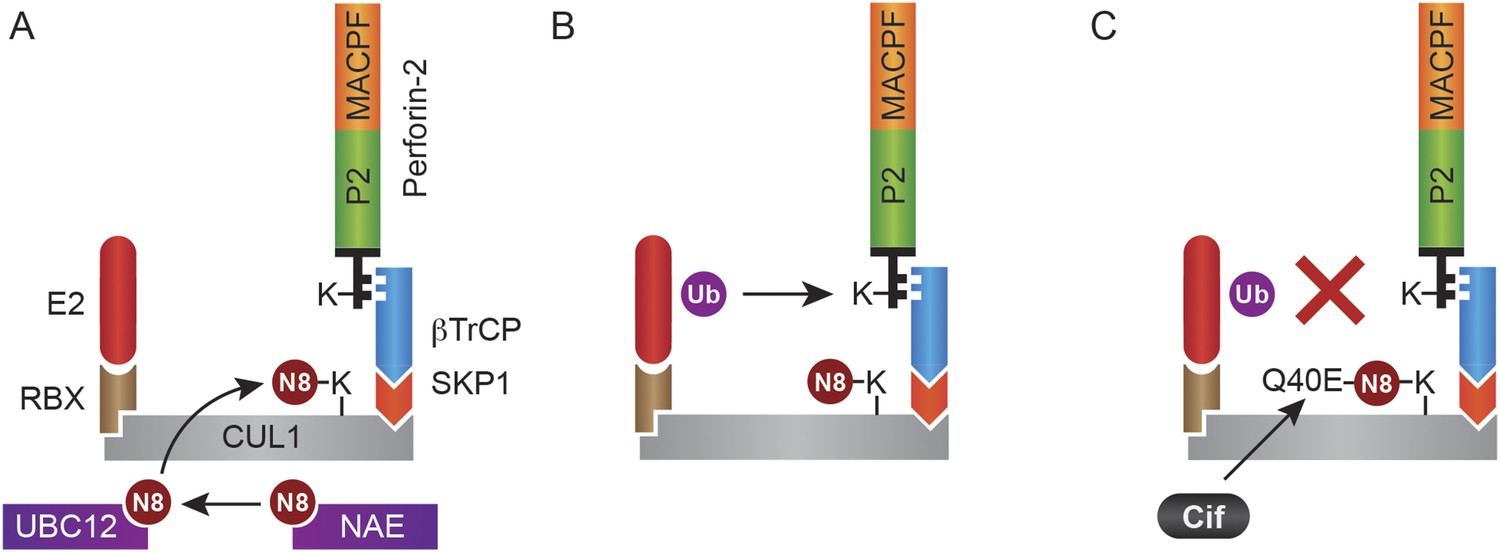
Monoubiquitylation of Perforin-2 and inhibition by Cif.
(A) CRLs are modular complexes assembled upon cullin scaffolds such as CUL1. F-box proteins such as βTrCP, which we have shown interacts with Perforin-2, provide substrate specificity within the multisubunit complex. Because βTrCP is known to recognize phosphorylated substrates, it is likely that TLR signaling activates one or more kinases that phosphorylate Perforin-2 prior to its association with the CRL. SKP1 serves as an adaptor between βTrCP and the cullin scaffold. An ubiquitin E2 ligase and a RING protein—either RBX1 or 2—dock at the opposite end of the elongated cullin. The E1 enzyme NEDD8 activating enzyme (NAE) activates NEDD8. The E2 ligase UBC12 then covalently binds NEDD8 to CUL1. (B) Neddylation of CUL1 activates the CRL complex and is essential for subsequent monoubiquitylation of Perforin-2. (C) The deamidation of NEDD8 by the bacterial effector protein Cif inhibits ubiquitylation of Perforin-2 and its subsequent trafficking to and destruction of extracellular bacteria at the plasma membrane or intracellular bacteria within an endosomal compartment. Ub, ubiquitin; N8, NEDD8; N8-Q40E, deamidated NEDD8.
Videos
Video 1
Perforin-2-RFP is located in vesicular bodies following stimulation with LPS.
Mpeg1 knockout MEFs were transfected with a Perforin-2-RFP expression plasmid and stimulated overnight with IFN-γ. After stimulation with LPS for 15 min, cells were fixed and counter stained with DAPI. Three dimensional projections of the acquired confocal Z-stacks reveal that Perforin-2 is located in punctate bodies with vesicular structure in LPS stimulated cells.
Video 2
Redistribution of Perforin-2 is dependent upon ubiquitylation.
Mpeg1 knockout MEFs were transfected with a Perforin-2-KQ-RFP expression plasmid. This fusion protein cannot be ubiquitylated because it carries three K-to-Q mutations in the carboxy-terminal tail of Perforin-2. Transfected cells were stimulated overnight with IFN-γ then LPS. Cells were fixed 15 min after exposure to LPS and counter stained with DAPI. Three dimensional projections of the acquired confocal Z-stacks reveal that Perforin-2-KQ-RFP has a diffuse, perinuclear distribution in LPS stimulated cells.
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Enteric pathogens deploy cell cycle inhibiting factors to block the bactericidal activity of Perforin-2
eLife 4:e06505.
https://doi.org/10.7554/eLife.06505




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}