Prophage-encoded Hm-oscar gene recapitulates Wolbachia-induced male-killing in the tea tortrix moth Homona magnanima
- National Agriculture and Food Research Organization, Japan
- United Graduate School of Agricultural Science, Tokyo University of Agriculture and Technology, Japan
- Department of Agricultural and Environmental Biology, Graduate School of Agricultural and Life Sciences, The University of Tokyo, Japan
- Crop Environment Section, Tea and Beverage Research Station, Ministry of Agriculture, Taiwan
eLife Assessment
Hardly anything is known about the genetic basis and mechanism of male-killing. Recently, a gene called oscar, in the bacterium Wolbachia, was implicated in killing male corn borer moths by interfering with moth genes that control sex determination and proper dosage of sex-specific genes. In this paper, the authors show that a distantly related oscar gene in another strain of Wolbachia kills male tea tortrix moths in a similar mechanism. This valuable study cements our understanding of the sophisticated way that Wolbachia kills male moths and butterflies (Lepidoptera) so early in their development. The conclusions are supported by solid evidence.
https://doi.org/10.7554/eLife.101101.4.sa0Significance of the findings:
Valuable: Findings that have theoretical or practical implications for a subfield
- Landmark
- Fundamental
- Important
- Valuable
- Useful
Strength of evidence:
Solid: Methods, data and analyses broadly support the claims with only minor weaknesses
- Exceptional
- Compelling
- Convincing
- Solid
- Incomplete
- Inadequate
During the peer-review process the editor and reviewers write an eLife Assessment that summarises the significance of the findings reported in the article (on a scale ranging from landmark to useful) and the strength of the evidence (on a scale ranging from exceptional to inadequate). Learn more about eLife Assessments
Abstract
Wolbachia are maternally transmitted bacterial symbionts that are ubiquitous among arthropods. They can hijack host reproduction in various ways, including male-killing (MK), where the sons of infected mothers are killed during development. The recent discovery of MK-associated Wolbachia genes, i.e., oscar in Ostrinia moths and wmk in Drosophila flies, stimulates our interest in the diversity and commonality of MK mechanisms, which remain largely unclear. We recently discovered that a Wolbachia symbiont of the moth Homona magnanima carries an MK-associated prophage region encoding homologs of oscar (Hm-oscar) and wmk (wmk-1–4). Here, we investigated the effects of these genes in the native host. Upon transient overexpression, Hm-oscar, but not wmk, induced male lethality in H. magnanima, in contrast to our observations in Drosophila, where the wmk homologs, but not Hm-oscar, killed the males. Hm-oscar disrupted sex determination in male embryos by inducing a female-type doublesex splicing and impaired dosage compensation, recapitulating the Wolbachia phenotype. Cell-based transfection assays confirmed that Hm-oscar suppressed the function of masculinizer, the primary male sex determinant involved in lepidopteran dosage compensation. Our study highlights the conserved roles of oscar homologs in Wolbachia-induced lepidopteran MK and argues that Wolbachia have evolved multiple MK mechanisms in insects.
eLife digest
Bacteria, viruses and other microbes are often thought of as external threats – but some also live inside animals, where they can be passed from mother to offspring. These types of microbes are commonly carried by insects and have various strategies for manipulating reproduction so they can spread through a population more easily.
One striking example is male killing, in which microbes selectively kill the male offspring of infected mothers during development. This results in a predominately female population, which can continue transmitting the microbe to future generations. Here, Arai et al. investigate whether Wolbachia – a group of bacteria that infect more than half of all insect species – employ a universal male-killing strategy, or whether they have evolved different mechanisms tailored to specific hosts.
The team focused on a region of the Wolbachia genome that encodes multiple candidate genes involved in male killing. First, Arai et al. explored whether a Wolbachia gene called Hm-oscar is responsible for male killing in the moth Homona magnanima. They found that over activating Hm-oscar in embryos not infected with Wolbachia increased male mortality, reproducing the effects of male killing, leading to a female-biased sex ratio. Further experiments confirmed that Hm-oscar exerts this effect by inhibiting masculinizer, a key gene necessary for male development.
Next, Arai et al. tested another gene called wmk, which Wolbachia use to kill the male offspring of fruit flies. However, manipulating this gene in H. magnanima had no impact on male survival, suggesting that Wolbachia use different male-killing genes in different insect species.
Understanding how Wolbachia affect insect reproduction sheds light on how bacteria interact with different host species. This knowledge also has practical applications. For instance, male-killing strategies could be leveraged for pest control to reduce populations of harmful insects without using pesticides. Future research may uncover additional Wolbachia genes involved in reproductive manipulation and explore their potential use in managing insect populations.
Introduction
Arthropods commonly carry microbial symbionts that are passed from mother to offspring (Hurst and Frost, 2015; Werren et al., 2008; Hurst, 2017). The maternally transmitted bacteria, belonging to the genus Wolbachia (Alphaproteobacteria), are estimated to be present in at least 40% of all insect species, making them one of the most widespread endosymbiont genera on the planet (Werren et al., 2008; Zug and Hammerstein, 2012). Wolbachia have achieved evolutionary success by manipulating host reproduction through various means that enhance endosymbiont transmission (Werren et al., 2008). Such manipulation of the host’s reproduction includes cytoplasmic incompatibility (CI), parthenogenesis, feminization, and MK, each of which affects the biological features, distribution, and evolution of the host. Among these strategies, MK directly skews the sex ratio of the host population toward females by killing male offspring of infected mothers during development. The lack of symbiont transmission through male hosts often leads to the evolution of MK. Wolbachia have been shown to induce MK in a wide range of insect taxa (e.g. dipterans, lepidopterans, and coleopterans) and other arthropods (Hurst et al., 1999; Kageyama and Traut, 2004; Hurst et al., 2000). Furthermore, various bacteria, viruses, and microsporidia induce MK phenotypes (Kageyama et al., 2012; Fujita et al., 2020; Kageyama et al., 2023; Nagamine et al., 2023), and recent studies have postulated that these microbes have evolved their MK ability independently (Harumoto and Lemaitre, 2018; Kageyama et al., 2023; Nagamine et al., 2023; Arai et al., 2023b).
The evolution and molecular mechanisms underlying Wolbachia-induced MK, observed across diverse insect taxa, have attracted considerable attention for decades. MK mechanisms are hypothesized to be linked to sex determination cascades regulating male and female differentiation in insects (Hornett et al., 2022). Indeed, some MK-inducing Wolbachia strains disrupt the sex-determination system in host moths (Sugimoto and Ishikawa, 2012; Sugimoto et al., 2015; Fukui et al., 2015; Arai et al., 2023b). Lepidopteran insects (moths and butterflies) generally have a female heterogametic sex chromosome system (e.g. WZ in females, ZZ in males) and employ dosage compensation, which equalizes the Z-chromosome-linked gene dose between sexes (Kiuchi et al., 2014). Dosage compensation is regulated by the masculinizer gene (masc), which is critical for male development. The lepidopteran sex determination system consists of multiple transcriptional regulators, some of which exhibit sex-linked expression and/or splicing isoforms. The primary male sex determinant, masc, also regulates the downstream master sex determination gene, doublesex (dsx), which exhibits sex-dependent splicing isoforms (dsxF in females and dsxM in males) (Kiuchi et al., 2014). In Ostrinia and Homona male moths, MK-inducing Wolbachia strains impair dosage compensation by disrupting the expression of masc. Furthermore, they disrupt sex determination in male moths by inducing the ‘female’ isoform of dsx (dsxF), leading to a ‘mismatch’ between genetic sex (male: ZZ sex chromosome constitution) and phenotypic sex (female: based on dsxF), ultimately resulting in male death (Arai et al., 2023b; Fukui et al., 2015; Sugimoto et al., 2015; Sugimoto and Ishikawa, 2012).
More recently, a Wolbachia protein named Oscar (Osugoroshi protein containing CifB C-terminus-like domain and many Ankyrin Repeats; Osugoroshi translates to male-killing in Japanese) was shown to recapitulate the Wolbachia wFur-induced MK phenotype in the native host moth Ostrinia furnacalis (Katsuma et al., 2022). The Oscar protein interacts with and degrades Masc protein, leading to the failure of dosage compensation and the production of female-type dsx isoforms in Ostrinia male moths (Fukui et al., 2024; Katsuma et al., 2022). Although oscar homologs are common in MK Wolbachia strains from Lepidoptera, MK strains from Diptera (Drosophila) and from several lepidopteran hosts lacked the gene (Arai et al., 2023a; Arai et al., 2024a). Furthermore, oscar homologs are evolutionarily dynamic, with highly variable sequences and structures (Arai et al., 2024a), making it difficult to assess their functional relevance. In addition to oscar, the helix-turn-helix domain-containing putative transcriptional regulator wmk is widely conserved among Wolbachia strains and induces various toxicities in Drosophila flies, ranging from no effect, weak male lethality (30% mortality), strong male lethality (90–100% mortality), or complete lethality of both sexes (Perlmutter et al., 2019; Perlmutter et al., 2020; Perlmutter et al., 2021; Arai et al., 2023a). Although the mechanisms underlying the wmk-induced toxicities and their connection to sex determination systems remain unclear, these findings suggest that Wolbachia strains carry multiple factors that cause male lethality. However, the diversity and commonality of these functions in insects remain largely unknown, partly due to the technical challenges in validating gene functions in non-model insects.
We recently discovered a prophage region responsible for a Wolbachia-induced MK by comparing closely related non-MK Wolbachia (wHm-c) and MK Wolbachia (wHm-t) in the tea tortrix moth Homona magnanima (Tortricidae) (Arai et al., 2023a; Arai et al., 2024b). The MK-associated prophage element encodes potential MK causative genes–four wmk homologs (wmk-1 to wmk-4) as well as an oscar homolog (Hm-oscar, encoding 1181 aa protein), which differs significantly in sequence length and structure from the wFur-encoded oscar (encoding 1830 aa protein) (Figure 1a, Figure 1—source data 1). When these MK candidate genes are transgenically overexpressed in Drosophila, wmk-1 and wmk-3 have a lethality of almost 100%, while wmk-2, wmk-4, and Hm-oscar induce no lethal effects when singly overexpressed. Furthermore, co-expression of the adjacent wmk-3 and wmk-4 has been shown to induce the death of 90% of male flies and restores female survival, suggesting their combined action (Arai et al., 2023a). However, transgenic expression of genes from a moth-derived male killer in flies does not necessarily replicate the native host–bacteria interaction, and the mechanistic links between prophage-encoded Wolbachia genes and MK in the native host Homona remain unclear.
Figure 1
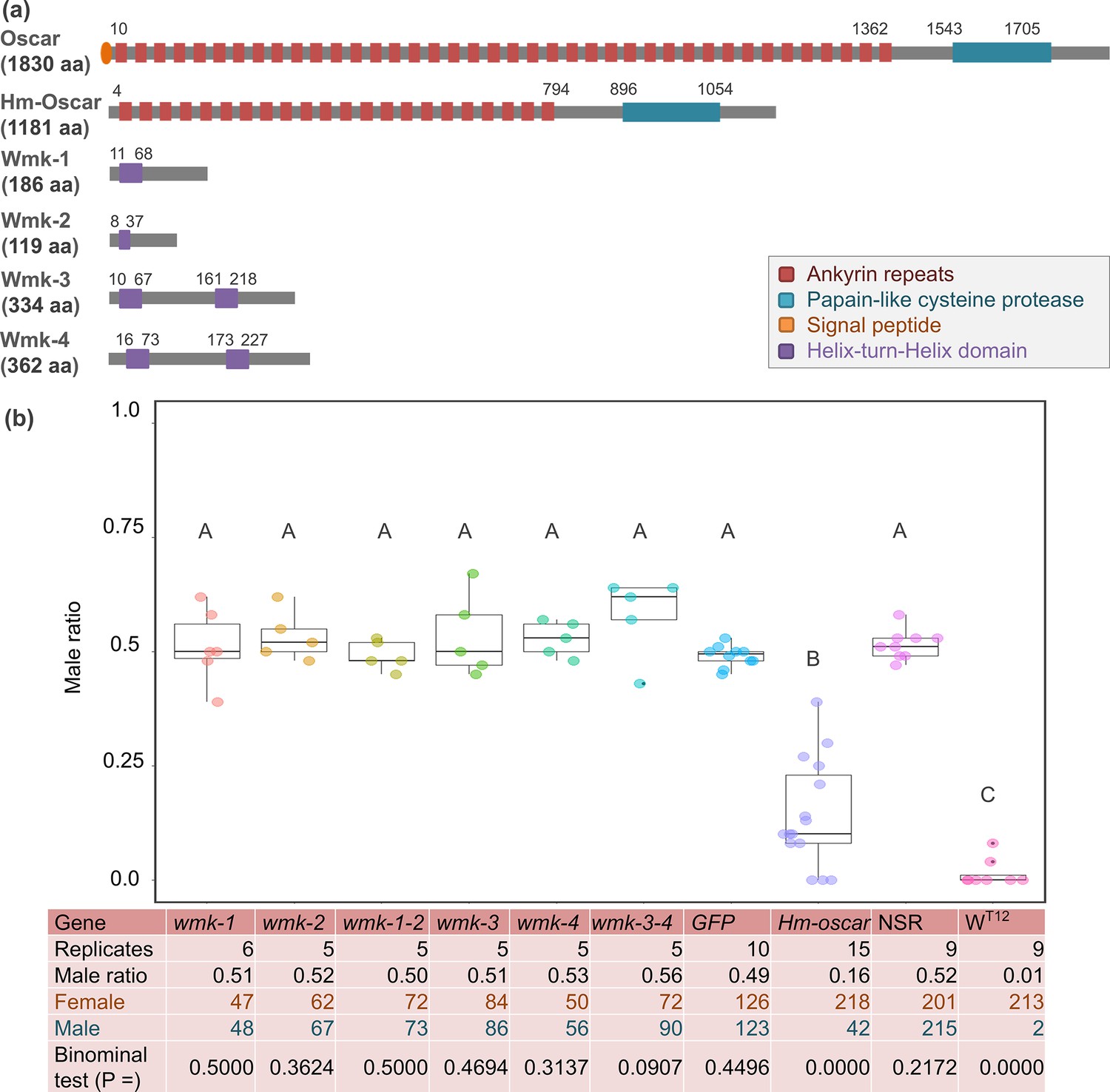
Transient expressions of the homologs of oscar (Hm-oscar) gene resulted in female-biased sex ratios.
(a) Structures of wFur-encoded Oscar and wHm-t-encoded Hm-Oscar and four Wmk proteins. (b) Male ratio of adult progeny obtained from cRNA-injected groups (wmk-1, wmk-2, T2A-bridged wmk-1 and wmk-2, wmk-3, wmk-4, T2A-bridged wmk-3 and wmk-4, GFP, and Hm-oscar; n = 5–15 independent replicates using different egg masses), the non-injected normal sex ratio (NSR) line, and the male-killing (MK) wHm-t-positive WT12 line. The total numbers of adult females and males are shown at the bottom. Different letters indicate significant differences (Steel–Dwass test, p<0.05). The dot plots show all data points individually.
-
Figure 1—source data 1
Sequence ID and data for wmk and oscar homologs displayed in Figure 1a.
- https://cdn.elifesciences.org/articles/101101/elife-101101-fig1-data1-v1.xlsx
-
Figure 1—source data 2
Number of males and females and sex ratio displayed in Figure 1b.
- https://cdn.elifesciences.org/articles/101101/elife-101101-fig1-data2-v1.xlsx
-
Figure 1—source data 3
Statistical analysis (Steel-Dwass test) data displayed in Figure 1b.
- https://cdn.elifesciences.org/articles/101101/elife-101101-fig1-data3-v1.xlsx
In this study, we showed that the prophage-encoded Hm-oscar recapitulates Wolbachia-induced MK in H. magnanima. Furthermore, we clarified the mechanistic links to host sex-determination cascades both in vivo and in vitro and discussed the underlying mechanisms of MK in Lepidoptera, arguing for the diverse evolutionary origin of Wolbachia-induced MK.
Results
Hm-oscar induces female-biased sex ratios
To achieve the transient overexpression of Hm-oscar and the four wmk genes (wmk-1, wmk-2, wmk-3, and wmk-4), constructed mRNA (cRNA) was injected into Wolbachia-free H. magnanima embryos. Subsequently, the adult moths that emerged from the cRNA-injected embryos were sexed based on their external morphology. When Hm-oscar was overexpressed, the sex ratio of adults was strongly female-biased (42 males and 218 females in total, 14.4 ± 11.1% male ratio in 15 replicates), which was in sharp contrast with the ratios observed in the GFP-injected (126 females and 123 males in total, 49.1 ± 2.08% male ratio in 10 replicates) and non-injected (NSR) (201 females and 215 males in total, 51.4 ± 3.08% male ratio in nine replicates) groups (p=0.001 and 0.002, respectively, Steel–Dwass test, Figure 1b, Figure 1—source data 2, Figure 1—source data 3). Compared with the GFP-injected group, the sex ratio was not biased by the overexpression of wmk-1 (51.1 ± 7.27% male ratio in six replicates, p=0.999), wmk-2 (53.2 ± 4.77% male ratio in 6 replicates, p=0.810), wmk-3 (53.3 ± 7.91% male ratio in five replicates, p=0.999), and wmk-4 (52.9 ± 3.47% male ratio in five replicates, p=0.773). Although the dual expression of wmk-3 and wmk-4 induces strong male lethality in Drosophila (Arai et al., 2023a), here, it did not skew the sex ratio of Homona compared with the ratio detected in the GFP-injected group (57.9 ± 7.67% male ratio in five replicates, p=0.704). Similarly, the dual expression of the tandemly arrayed wmk-1 and wmk-2 did not bias the sex ratio (49.4 ± 2.82% male ratio in five replicates) compared with the value in the GFP-injected group (p=1.000). The sex ratio data of all replicates are available on the Figure 1—source data 2.
Males are killed mainly at the embryonic stage
The sex of hatched larvae (neonates) and the remaining unhatched embryos was determined by the presence or absence of W chromatin, a condensed structure of the female-specific W chromosome observed during interphase. MK Wolbachia wHm-t kills Homona males during embryogenesis, resulting in a female-biased sex ratio in hatched larvae (neonates) and a male-biased sex ratio in unhatched embryos (Arai et al., 2023a; Arai et al., 2020). In line with this finding, the sex ratios of hatched larvae (34 females and 6 males) and unhatched embryos (27 females and 47 males) of the MK wHm-t infected line (WT12) were also female- and male-biased, respectively (p=0.013 and 0, respectively). In the Hm-oscar-injected group, the sex ratio of the hatched larvae (neonates) was strongly female-biased (21 females and 3 males, p=0.0001 in the binomial test) (Figure 2a, Figure 2—source data 1). In contrast, the unhatched mature embryos showed a male-biased ratio (18 females and 38 males, p=0.005), suggesting that male mortality primarily occurred during the embryonic stage, reflecting the wHm-t-induced MK phenotype. In the non-injected group (NSR), the sex ratios of hatched larvae (28 females and 28 males) and unhatched embryos (23 females and 25 males) were not biased (p=0.553 and 0.443, respectively).
Figure 2
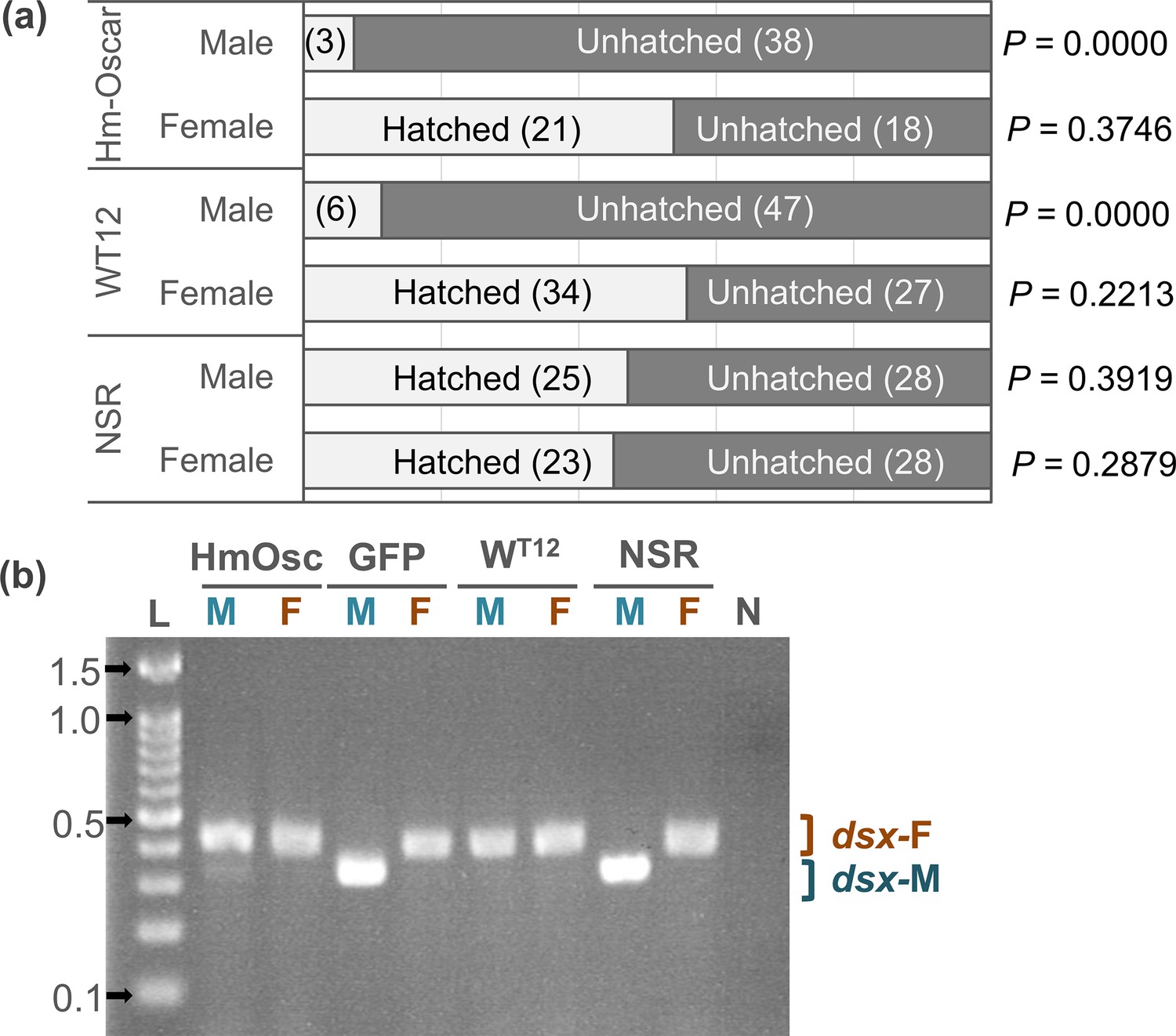
Homologs of oscar (Hm-Oscar) induced lethality in male embryos with the female-type sex determination.
(a) Sex ratio of the hatched larvae and unhatched embryos in the Hm-oscar-expressed, wHm-t-infected, and non-infected/expressed groups. Females and males were discriminated based on the presence or absence of W chromatin. The number of individuals is indicated in brackets. (b) Splicing patterns of the downstream sex-determining gene dsx of H. magnanima embryos (5 d post oviposition). Abbreviations: HmOsc, Hm-oscar injected group; GFP, GFP-injected group; WT12, wHm-t-infected line; NSR, non-infected/injected line. M and F indicate W chromatin-negative (ZZ: male genotype) and W chromatin-positive (ZW: female genotype) mature embryos, respectively. dsx-F and dsx-M represent female and male-specific splicing variants, respectively. L: 100 bp DNA ladder (ExcelBand 100 bp DNA Ladder, SMOBIO Technology, Inc, Hsinchu, Taiwan). 0.1, 0.5, 1.0, and 1.5 kb markers are indicated with arrows. N: negative control (water).
-
Figure 2—source data 1
Number of males and females displayed in Figure 2a.
- https://cdn.elifesciences.org/articles/101101/elife-101101-fig2-data1-v1.xlsx
-
Figure 2—source data 2
Original gel image for band patterns of dsx displayed in Figure 2b.
- https://cdn.elifesciences.org/articles/101101/elife-101101-fig2-data2-v1.zip
-
Figure 2—source data 3
PDF file containing an original gel image for Figure 2b, indicating the relevant bands and treatments.
- https://cdn.elifesciences.org/articles/101101/elife-101101-fig2-data3-v1.pdf
Female-type sex determination in male embryos that are destined to die
Splicing patterns of the downstream sex determinant dsx were assessed in each of the H. magnanima embryos that were Hm-oscar-expressed, GFP-expressed, wHm-t-infected, or non-expressed (i.e. non-injected, NSR). In GFP-expressed and NSR embryos, females and males (which were identified by the presence and absence of W chromatin, respectively) exhibited a female- and male-type splicing variant of dsx, respectively. However, both wHm-t-infected and Hm-oscar-expressed embryos induced female-type dsx splicing regardless of the presence or absence of W chromatin (Figure 2b, Figure 2—source data 2, Figure 2—source data 3). Notably, male embryos expressing Hm-oscar also exhibited weak male-type dsx splicing in addition to the female-type splicing, resembling the previously observed pattern in male embryos infected with low-titer wHm-t (Arai et al., 2023b).
Hm-oscar impairs dosage compensation in male embryos
To confirm the effects of Hm-oscar on dosage compensation, we quantified the expression of Z-linked genes using RNA-seq and qPCR. Unlike other lepidopteran species, the family Tortricidae, including H. magnanima, generally possess a large Z chromosome that is homologous to B. mori chromosomes 1 (Z) and 15 (autosome). RNA-seq analysis revealed that, in Hm-oscar-injected embryos, Z-linked genes (homologs on the B. mori chromosomes 1 and 15) were more expressed in males than in females (Figure 3a, Figure 3—source data 1, Figure 3—source data 2), which was not observed in the GFP-injected group (Figure 3b, Figure 3—source data 3, Figure 3—source data 4). Similarly, as previously reported by Arai et al., 2023b, high levels of Z-linked gene expression were also observed in wHm-t-infected males, but not in NSR males (Figure 3c and d, Figure 3—source data 5, Figure 3—source data 6, Figure 3—source data 7, Figure 3—source data 8). The high (i.e. doubled) Z-linked gene expression in both Hm-oscar-expressed and wHm-t-infected males was further confirmed by quantification of the Z-linked Hmtpi gene (Figure 3c, Figure 3—source data 9, Figure 3—source data 10). These trends were statistically supported, with all data available in the source data files (i.e. Figure 3, Figure 3—source data 1, Figure 3—source data 2, Figure 3—source data 3, Figure 3—source data 4, Figure 3—source data 5, Figure 3—source data 6, Figure 3—source data 7, Figure 3—source data 8, Figure 3—source data 9, and Figure 3—source data 10).
Figure 3
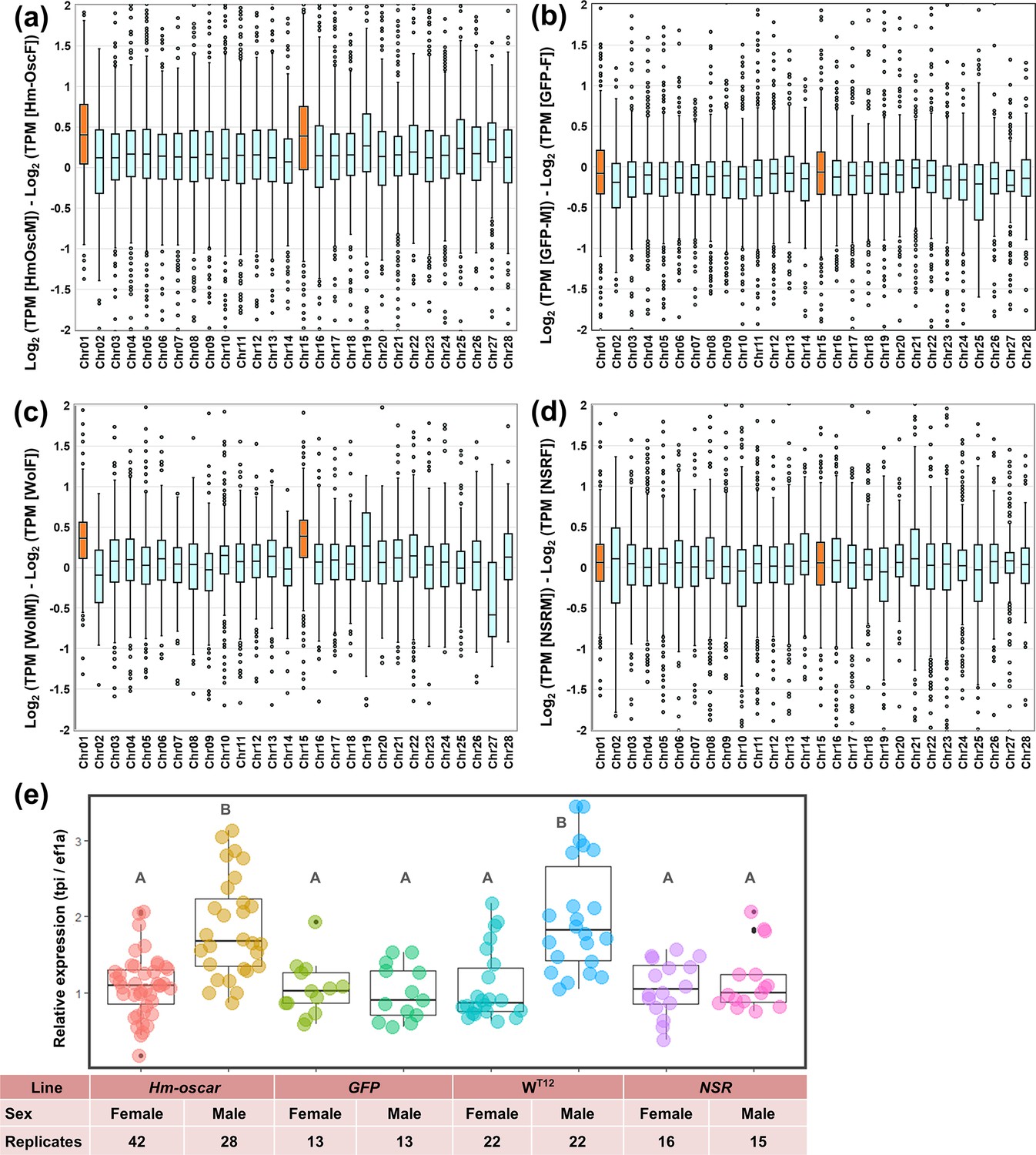
Homologs of oscar (Hm-oscar)-overexpressed male embryos showed higher levels of Z-linked gene expression.
(a–d) Normalized expression levels (TPM) and chromosomal distributions of transcripts in H. magnanima embryos. RNA-seq data of embryos (108 hpo) were used to make the following comparisons: Hm-oscar-injected males versus Hm-oscar injected females (a), GFP-injected males versus GFP-injected females (b), WT12 males versus WT12 females (c), and normal sex ratio (NSR) males versus NSR females (d). The chromosome number for each H. magnanima transcript-derived contig was assigned based on Bombyx mori gene models. The x-axis represents the chromosome number of B. mori (shown as chr01 to chr28), and chr01 and chr15 (highlighted in orange) correspond to the Z chromosome of H. magnanima (Arai et al., 2023b). (e) Relative expression of a Z-linked tpi gene in H. magnanima groups. Statistical significance is highlighted with different alphabets.
-
Figure 3—source data 1
Expression data displayed in Figure 3a.
- https://cdn.elifesciences.org/articles/101101/elife-101101-fig3-data1-v1.xlsx
-
Figure 3—source data 2
Statistical analysis (Steel-Dwass test) data displayed in Figure 3a.
- https://cdn.elifesciences.org/articles/101101/elife-101101-fig3-data2-v1.xlsx
-
Figure 3—source data 3
Expression data displayed in Figure 3b.
- https://cdn.elifesciences.org/articles/101101/elife-101101-fig3-data3-v1.xlsx
-
Figure 3—source data 4
Statistical analysis (Steel-Dwass test) data displayed in Figure 3b.
- https://cdn.elifesciences.org/articles/101101/elife-101101-fig3-data4-v1.xlsx
-
Figure 3—source data 5
Expression data displayed in Figure 3c.
- https://cdn.elifesciences.org/articles/101101/elife-101101-fig3-data5-v1.xlsx
-
Figure 3—source data 6
Statistical analysis (Steel-Dwass test) data displayed in Figure 3c.
- https://cdn.elifesciences.org/articles/101101/elife-101101-fig3-data6-v1.xlsx
-
Figure 3—source data 7
Expression data displayed in Figure 3d.
- https://cdn.elifesciences.org/articles/101101/elife-101101-fig3-data7-v1.xlsx
-
Figure 3—source data 8
Statistical analysis (Steel-Dwass test) data displayed in Figure 3d.
- https://cdn.elifesciences.org/articles/101101/elife-101101-fig3-data8-v1.xlsx
-
Figure 3—source data 9
Expression data displayed in Figure 3e.
- https://cdn.elifesciences.org/articles/101101/elife-101101-fig3-data9-v1.xlsx
-
Figure 3—source data 10
Statistical analysis (Steel-Dwass test) data displayed in Figure 3e.
- https://cdn.elifesciences.org/articles/101101/elife-101101-fig3-data10-v1.xlsx
Hm-oscar suppresses the masculinizing functions of lepidopteran masc genes
To confirm whether Hm-oscar suppresses the functions of the primary male sex determinant masc, their interactions were assessed through transfection in BmN-4 cells. These cells, derived from B. mori ovaries (Grace, 1967), exhibit female-type default sex determination (Kiuchi et al., 2014). However, overexpression of the masc gene from various lepidopteran insects, such as O. furnacalis, induces male-type sex determination by promoting the expression of BmdsxM and BmIMPM in BmN-4 cells (Fukui et al., 2015; Katsuma et al., 2019; Katsuma et al., 2022). Since BmIMPM regulates the splicing of BmdsxM and is exclusively expressed in males, we selected BmIMPM as a quantitative marker of masculinization, as described in Katsuma et al., 2022.
In contrast with the non-expressed control groups, which exhibited default female-type sex determination, cells expressing Hmmasc exhibited male-type sex determination (Figure 4a, Figure 4—source data 1). This was indicated by the increased expression of the male-specific splicing variant BmIMPM, although this increase was not statistically significant. Furthermore, the masculinizing function of Hmmasc was suppressed when Hmmasc and Hm-oscar were co-expressed, as evidenced by the low expression levels of BmImpM (Figure 4a). wFur-encoded Oscar, which differs from Hm-Oscar in structure and amino acid sequence (Figure 1a), also suppressed Hmmasc, but its function was less active than that of Hm-oscar (Figure 4a). Hm-oscar also suppressed the function of masc genes derived from various lepidopteran insects (i.e. O. furnacalis, Spodoptera frugiperda, Bombyx mori, Trilocha varians, and Papilio machaon), which are highly divergent in their amino acid sequences (Figure 4b, c, Figure 4—source data 2, Figure 4—source data 3). These results suggest that Hm-oscar has a broad spectrum of activity in Lepidoptera.
Figure 4
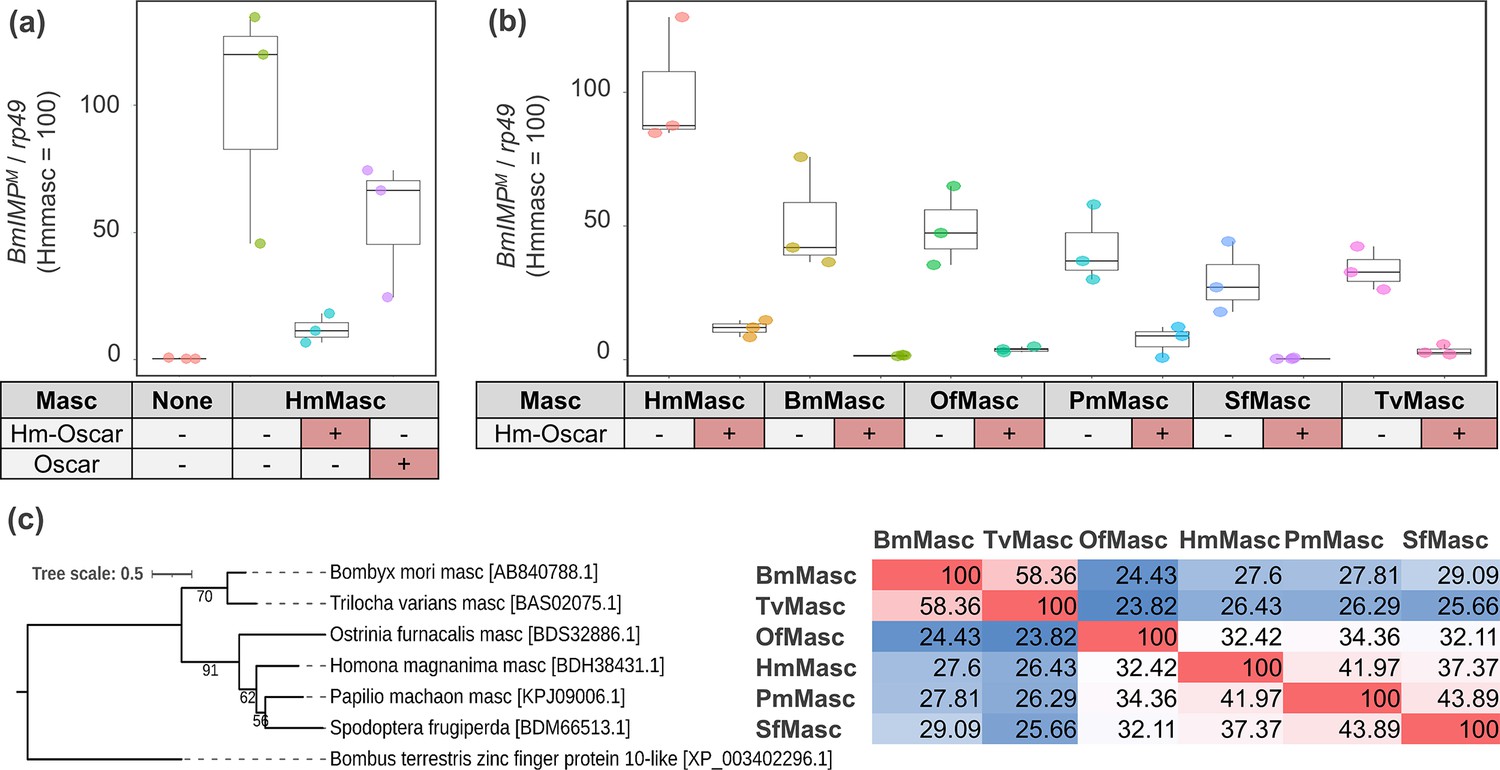
Hm-oscar suppressed the masculinizing function of lepidopteran Masc.
(a) Relative expression levels of the male-specific BmImpM variant in transfected BmN-4 cells. The relative expression of BmImpM in the control (non-inserted pIZ/V5) and Hm-oscar/oscar co-transfected groups was normalized by setting the mean in the Hmmasc single transfected group as 100. (b) Relative expression levels of the BmImpM in Hm-oscar/masc transfected BmN-4 cells. The relative expression of BmImpM in each condition was normalized by setting the mean in the Hmmasc single transfected group as 100. The dot plots show all data points individually. Each experimental condition was replicated three times. Abbreviations: Hm, Homona magnanima; Bm, Bombyx mori; Of, Ostrinia furnacalis; Pm, Papilio machaon; Sf, Spodoptera frugiperda; and Tv, Trilocha varians. (c) Phylogeny and homologies of the Masc protein sequences used in this study. The sequence homology between each pair is presented in the figure and visualized as a heatmap, with red indicating high homology (Max = 100) and blue indicating low homology (Min = 23.82).
-
Figure 4—source data 1
Expression data for IMP gene displayed in Figure 4a.
- https://cdn.elifesciences.org/articles/101101/elife-101101-fig4-data1-v1.xlsx
-
Figure 4—source data 2
Expression data for IMP gene displayed in Figure 4b.
- https://cdn.elifesciences.org/articles/101101/elife-101101-fig4-data2-v1.xlsx
-
Figure 4—source data 3
Sequence homology data displayed in Figure 4c.
- https://cdn.elifesciences.org/articles/101101/elife-101101-fig4-data3-v1.xlsx
Discussion
In this study, we showed that the phage-encoded Hm-oscar, but not wmk, induced male lethality and a female-biased sex ratio in H. magnanima. Furthermore, the overexpressed Hm-oscar impaired male sex determination in Homona, recapitulating the wHm-t-induced phenotypes. Cell-based assays confirmed that Hm-oscar suppressed the masculinizing functions of masc. These results strongly suggested that Hm-oscar underlies the wHm-t-induced MK function in H. magnanima. In light of our results and previous findings in Homona, Ostrinia, and Bombyx (Arai et al., 2023a; Katsuma et al., 2022; Kiuchi et al., 2014), we hypothesize the following molecular mechanisms for wHm-t-induced MK: The Hm-Oscar protein targets and inhibits the function of the HmMasc protein, thereby resulting in female sex determination and disrupting dosage compensation, which ultimately leads to male death. Although the full mechanisms remain unclear, unbalanced Z-linked gene expression mediated by improper dosage compensation—resulting in Z-linked gene expression levels twice as high as those in normal males—appear to be lethal for lepidopteran males (Kiuchi et al., 2014; Fukui et al., 2015; Visser et al., 2021).
The Wolbachia-encoded oscar homologs identified so far show diversity in sequence length and are classified as type I, II, and III based on their amino acid sequence homologies. Type I and II Oscar proteins, derived from MK-inducing Wolbachia, contain many ankyrin repeats and a proteinase domain (Arai et al., 2024a). Although long ankyrin repeat sequences are postulated to be critical for the function of wFur-encoded Oscar protein (1830 aa, type I) (Katsuma et al., 2022), our study revealed that the functions of Hm-Oscar protein (1181 aa, type II), which encodes fewer ankyrin repeats, were comparable with those of Oscar protein carried by wFur. Interestingly, type II oscar gene is also present in the feminizing Wolbachia wFem in Eurema butterflies (Arai et al., 2024a). oscar homologs, which inhibit the masculinizing function and induce female sex determination, may have a conserved function in Wolbachia-induced MK and feminization in Lepidoptera.
In contrast to the results of this study, we have previously demonstrated that the phage-encoded wmk, but not Hm-oscar, induces male lethality in D. melanogaster (Arai et al., 2023b). Although the means of expression are different (i.e. transient in Homona and transgenic in Drosophila), this finding highlighted the differences in the mode/range of action of Wolbachia genes between insect species. It has been hypothesized that microbes induce MK in insects by targeting molecular mechanisms involved in sex determination and differentiation (Hornett et al., 2022). Sex determination systems in insects are diverse; for example, Lepidoptera (including H. magnanima) and Diptera (including D. melanogaster) do not share any known sex-determining genes other than dsx (Suzuki, 2018). The different outcomes in Homona and Drosophila are probably due to their different sex determination systems. Because oscar interacts with and suppresses masc, Hm-oscar could induce mortality in Homona males that possess masc, but not in Drosophila males that lack it. Considering that oscar homologs are not present in known MK Wolbachia strains in dipteran insects (Arai et al., 2024c; Katsuma et al., 2022), the mechanisms (i.e. causative genes) of Wolbachia-induced MK probably differ between insect taxa (e.g. between Lepidoptera and Diptera). While the mechanisms underlying wmk-induced lethality remain unclear, the distinct effects associated with this gene between Homona and Drosophila may also reflect their genetic backgrounds, such as the presence or absence of host factor(s) that interact with wmk. In addition, Katsuma et al., 2022 reported that the wmk homologs encoded by wFur did not affect the masculinizing function of masc in vitro, indicating that wmk likely targets factors other than masc. Whilst we cannot rule out the possibility that wmk may work synergistically or interactively with oscar in vivo—potentially acting downstream of oscar’s impact—our results strongly suggested that Wolbachia strains have acquired multiple MK genes through evolution.
Wolbachia-induced phenotypes are known to be influenced by the genetic backgrounds of hosts (Hornett et al., 2006; Sasaki et al., 2002; Veneti et al., 2012). Our study showed that the wHm-t-encoded Hm-oscar suppresses the function of masc in H. magnanima more efficiently than the wFur-encoded oscar, suggesting that this Wolbachia factor has undergone evolutionary tuning to adapt to its natural host. However, the mere presence of oscar and wmk homologs does not ensure MK expression. For example, wCauA, bearing type I oscar, did not induce MK in Ephestia (Cadra) cautella collected around 2000, although it did induce MK when transferred to the closely related host Ephestia kuehniella (Sasaki et al., 2002). An MK phenotype, presumably induced by wCauA, was also recorded in E. cautella around the 1970s (Takahashi and Kuwahara, 1970). These findings suggest a suppressor evolution against wCauA-induced MK in E. cautella. In addition, Wolbachia typically encode a number of wmk homologs in their genomes, but their effects differ among insects and are not apparent in some species such as Homona, possibly due to the diversity in sex determination system or the evolution of suppressors. Nonetheless, given the high copy number of wmk homologs in Wolbachia genomes, they may still contribute to Wolbachia biology as transcriptional regulators and potential virulence factors. Virulence genes often undergo duplication and substitution under strong selective pressure (Hill et al., 2022; Jones and Dangl, 2006). An intense evolutionary arms race between Wolbachia and their hosts could have increased and diversified the MK-associated Wolbachia genes. Conversely, natural selection favors the rescue of males by suppressing the Wolbachia-induced reproductive manipulations (Hornett et al., 2006; Hornett et al., 2014), which may involve changes in the sex determination system because Wolbachia strains frequently hijack host reproduction systems. In this context, Wolbachia and other MK-inducing microbes may be a hidden driver for the diversification of complex insect sex determination systems.
In this study, we clarified the conserved roles of the Wolbachia-encoded oscar homologs in Lepidoptera and demonstrated that Wolbachia have evolved distinct MK mechanisms (through causative genes) in insect taxa. The diversification of phenotype/virulence-associated genes and the rampant horizontal transmission of phages carrying virulence genes between Wolbachia strains may have contributed to the outstanding success of this bacterial genus. In addition to oscar and wmk, Wolbachia may retain other uncharacterized genes that induce male lethality, and further studies on diverse Wolbachia–host systems are highly warranted. Our findings provide insights into the molecular mechanisms and evolutionary relationships between endosymbionts and their hosts, which may also contribute to the design of pest management strategies.
Materials and methods
Experimental model and subject details
Homona magnanima
Request a detailed protocolA laboratory-maintained H. magnanima line with a normal sex ratio (NSR) that was negative for Wolbachia and other endosymbionts, was used in our experiments. This line was initially collected in Tokyo, Japan, in 1999 and has been maintained inbred for over 250 generations in the laboratory. Larvae were reared using the artificial SilkMate 2 S diet (Nosan Co., Yokohama, Japan) at 25 °C under a long photoperiod (16 L:8D) (Arai et al., 2019).
The laboratory-maintained all female H. magnanima WT12 line, which was initially collected in Taiwan (Tea Research Extension Station, Taoyuan city) in 2015 (Arai et al., 2020), was also used in this study. This line was maintained for over 50 generations by crossing it with the males of the NSR line.
BmN-4 cell line
Request a detailed protocolBombyx mori BmN-4 cells (provided by Chisa Yasunaga-Aoki, Kyushu University, and maintained in our laboratory) were cultured at 26 °C in IPL-41 Insect Medium (Applichem, Darmstadt, Germany) supplemented with 10% fetal bovine serum. The cells used were authenticated by sequencing masc gene. The BmN-4 cells are of insect origin; therefore, we did not test for mycoplasma contamination, which is commonly screened in mammalian cell cultures. However, to prevent potential contamination, we used cells that were passaged in a medium containing penicillin, streptomycin, and amphotericin B.
Transient expression of MK-associated phage genes
mRNA synthesis
Request a detailed protocolCodon-optimized wmk genes (wmk-1 to wmk-4), conjugated wmk pairs using a T2A peptide (i.e. wmk-1-T2A-wmk-2 and wmk-3-T2A-wmk-4), and Hm-oscar genes synthesized by Arai et al., 2023a were used for mRNA synthesis. These synthetic genes were ligated into the plasmid pIZ/V5-His (Invitrogen, MA, USA) using the NEBuilder HiFi DNA Assembly kit (New England Biolabs, MA, USA) following the manufacturer’s protocol. The inserts of the vector (i.e. wmk-1, wmk-2, wmk-3, wmk-4, Hm-oscar, wmk-1-T2A-wmk-2, wmk-3-T2A-wmk-4, and GFP as a control gene) were amplified using KOD-one (TOYOBO, Osaka, Japan) with the primer set containing the T7 promoter described in Fukui et al., 2015 (i.e. pIZ-F-T7: 5′-TAATACGACTCACTATAGGGAGACAGTTGAACAGCATCTGTTC-3′ and pIZ-R: 5′-GACAATACAAACTAAGATTTAGTCAG-3′) under the following PCR conditions: 20 cycles at 98 °C for 10 s, 62 °C for 5 s, and 68 °C for 15 s. The amplicons were purified using the QIAquick PCR purification kit (Qiagen, Hilden, Germany), and 100–200 ng DNA was used for mRNA synthesis. The capped mRNA (cRNA) with poly(A) tail was synthesized using the mMESSAGE mMACHINE T7 Ultra kit (Invitrogen, MA, USA) following the manufacturer’s protocol with some modifications. In brief, the assembled transcription reaction (10 µL of T7 2 X NTP/ARCA, 2 µL of 10X T7 reaction buffer, 2 µL of T7 enzyme mix, and 6 µL of linear DNA template diluted with nuclease-free water) was incubated at 37 °C for at least 10 hr to maximize RNA yields. After poly(A) tailing, cRNA was purified using ISOGEN II (Nippongene, Tokyo, Japan) and dissolved in up to 10 µL of nuclease-free water to achieve an RNA concentration of approximately 1500–2000 ng/µL. The synthesized cRNA was preserved at −80 °C until further use.
Preparation of H. magnanima eggs
Request a detailed protocolA total of 15 males and 10 females of H. magnanima were mated in a plastic box (30 cm × 20 cm × 5 cm) for 3–4 d. The collection of egg masses began the day after oviposition was confirmed. In brief, newly oviposited egg masses were collected at 30 min intervals during the dark period using red light (which the moths cannot perceive). Females started to oviposit eggs at least 4 hr into the dark period and, within less than 30 min post oviposition (mpo), the egg masses were collected. The collection lasted until the start of the light period. The egg masses were then subjected to microinjection assays.
Inoculum preparation, microinjection, and maintenance of the injected embryos
Request a detailed protocolA glass needle for microinjection was prepared using glass capillary GD-1 (Narishige, Tokyo, Japan) with a PC-10 puller (Narishige). The glass capillaries were pulled at two temperatures (first stage: 75 °C, second stage: 65 °C) using two heavy weights and one light weight. The movement position during the second heating stage was set to 3 mm (range 1–10 mm).
The cRNA solution was diluted in a buffer (100 mM potassium acetate, 2 mM magnesium acetate, and 30 mM HEPES-KOH; at pH 7.4) containing 0.2% (W/V) Brilliant Blue FCF (Wako, Osaka, Japan) to obtain an RNA concentration of 1000 ng/µL. Approximately 1–4 µL of dye-containing mRNA solution was aspirated into the glass needles (capillaries), and the edge of each needle was ground using Micro Grinder EG-402 (Narishige) at an angle of 20 degrees for 2 s.
The fresh egg masses of H. magnanima (collected at less than 30 mpo as described above) were put on a double-sided sticky tape (15 mm × 5 m T4612, Nitoms, Tokyo, Japan) and placed on glass slides. Under a Nikon SMZ1270 microscope (Nicon, Tokyo, Japan), the RNA solution (30–100 fL) was injected into individual eggs using an microinjector IM-400 (Narishige) (balance pressure set at 0.010–0.030; injection pressure set at 0.050–0.100) and a QP-3JOY-2R electric micromanipulator (MicroSupport, Shizuoka, Shizuoka).
The injected egg masses were maintained in a 90 mm plastic Petri dish fitted with a slightly wet filter paper. As high humidity interfered with the development of the embryos, the injected eggs (egg mass) were first maintained at a relative humidity of 30–50% (0–3 d post-injection) and then at a higher humidity until hatching (4–6 dpo, 60% relative humidity). The hatched larvae were reared separately with 1/2 ounces of SilkMate 2 S (Nosan Co.) until eclosion. The adult H. magnanima moths that emerged from the cRNA-injected embryos were sexed based on their external morphology.
Sexing of embryos/neonates and RNA extraction
Request a detailed protocolTo verify the effect of the transient expression of Hm-oscar on sex determination in H. magnanima, Hm-oscar/GFP-expressed, non-injected, and wHm-t-infected mature embryos showing black head capsule (1 d before hatching, 5–6 d post oviposition, dpo) were dissected on glass slides using forceps, as described in Arai et al., 2022. Their Malpighian tubules were fixed with methanol/acetic acid (50% v/v) and stained with lactic acetic orcein for W chromatin observations. The remaining tissues not used for sexing were stored in ISOGEN II (Nippon Gene) at −80 °C until subsequent extraction. In total, 12 males or females (confirmed based on the presence or absence of W chromatin) were pooled and homogenized in ISOGEN II to extract RNA as described in Arai et al., 2023b. In brief, 0.4 times the volume of UltraPure distilled water (Invitrogen) was added to the ISOGEN II homogenates, which were centrifuged at 12000×g for 15 min at 4 °C to pellet proteins and DNAs. The resulting supernatant was mixed with the same volume of isopropanol to precipitate RNAs; then, the resulting solutions were transferred to EconoSpin columns (Epoch Life Science) and centrifuged at 17,900×g and 4 °C for 2 min. The RNAs captured in the column were washed twice with 80% ethanol and eluted in 20 µL of UltraPure distilled water (Invitrogen).
Hmdsx detection
Request a detailed protocolSex-specific dsx splicing variants of H. magnanima were detected as described in Arai et al., 2023b. In brief, total RNA (100–300 ng) extracted from sex-determined mature embryos was reverse-transcribed using PrimeScript II Reverse Transcriptase (TaKaRa, Shiga, Japan) at 30 °C for 10 min, 45 °C for 60 min, and 70 °C for 15 min. Then, cDNA was amplified using KOD-FX Neo (Toyobo Co., Ltd.) with the following two primers: Hmdsx_long3F (5′-TGCCTAAAGTGAAAACGCCGAGGAGCC-3′) and Hmdsx_Mrev (5′-TGGAGGTCTCTTTTCATCCGG-3′). The PCR conditions used were as follows: 94 °C for 2 min, followed by 45 cycles of 98 °C for 10 s, 66 °C for 30 s, and 68 °C for 30 s. The amplicons were subjected to electrophoresis on 2.0% agarose Tris-borate-EDTA buffer (89 mM Tris-borate, 89 mM boric acid, 2 mM EDTA) gels.
Quantification of Z chromosome-linked genes
Request a detailed protocolThe effects on dosage compensation were assessed by measuring differences in gene expression, as described in Fukui et al., 2015 and Arai et al., 2023b. A total of 1.0 µg of the total RNA extracted from Hm-oscar or GFP-overexpressed mature embryos (108 hpo, approximately 40–50 sex-determined embryos) was used to prepare mRNA-seq libraries via the NEBNext Poly (A) mRNA Magnetic Isolation Module (New England Biolabs) and the NEBNext Ultra II RNA Library Prep kit for Illumina (New England Biolabs) following the manufacturer’s protocol. The adaptor sequences and low-quality reads (Qscore <20) were removed from the generated sequence data [150 bp paired-end (PE150)] using Trimmomatic (Bolger et al., 2014). The trimmed reads were mapped to the previously assembled transcriptome database for H. magnanima (Arai et al., 2023b) using Kallisto (Bray et al., 2016) to generate the normalized read count data (transcripts per million, TPM). While we did not generate new RNA-seq data of the wHm-t-infected and uninfected groups, the expression data included in Arai et al., 2023b were re-analyzed along with the newly obtained RNA-seq data above. The binary logarithms of the TPM differences between males and females belonging to each H. magnanima group (i.e. Hm-oscar/GFP-expressed) were calculated to assess the fold-changes in gene expression levels between sexes. As described in Arai et al., 2023b, the transcriptome data of H. magnanima were annotated using the B. mori gene sets obtained from KAIKOBASE (https://kaikobase.dna.affrc.go.jp). The binary logarithms of the TPM differences between males and females in the B. mori chromosomes 1–28 were plotted.
To confirm the effects observed in the RNA-seq-based quantification of the Z chromosome genes, expression level of the Z-linked Hmtpi gene was quantified by reverse transcription-qPCR as described in Arai et al., 2023b. Briefly, total RNA (100–300 ng) extracted from pooled male or female mature embryos (108 hpo) was reverse transcribed using PrimeScript II reverse transcriptase (TaKaRa) as described above. The cDNA was used to quantify relative gene expression levels, with normalization to the control gene elongation factor 1 a (ef1a). qPCR was performed using primer sets for Hmtpi (HmTpi_F180702_212: 5′-GCTGCGAGTGGGTGATTTTG-3′; HmTpi_R180702_326: 5′-GCGATCACTTTCAGACCCGA-3′) and Hmef1a (Hmef1a_F_val1_85: 5′-TTTCCAGGGTGGTTGAGCA-3′; Hmef1a_R_val1_193: 5′-CCGTTAAGGAGCTGCGTCG-3′), and KOD SYBR qPCR Mix (Toyobo, Osaka, Japan) in a LightCycler 96 system (Roche, Basel, Switzerland). The qPCR reaction was made with 5 µl KOD SYBR (TOYOBO), 0.4 µL forward primer [10 pmol/µl], 0.4 µl reverse primer [10 pmol/µl], and 3.2 µl water. The qPCR was performed under the following conditions: 180 s at 95 °C, 40 cycles of 8 s at 98 °C, 10 s at 60 °C, and 10 s at 68 °C, followed by heating to 90 °C for melting curve analysis. Mean cycle threshold (CT) values of samples were calculated for at least 13 replicates, and both ΔCT (CT Z gene – CT ef1a) and ΔΔCT (ΔCTmale – ΔCTfem) values were calculated.
Transfection assays and quantification of BmIMPM
Request a detailed protocolThe coding sequence of Hmmasc was amplified from cDNA derived from the RNA extracted from male embryos of the NSR line using KOD-one (TOYOBO) with the following primer set: HmMasc_CDS_HindIIIF: 5′- GCAAAGCTTCAACATGATCTCTCGCCAACCACAATCAACATCA-3′ and HmMasc_CDS_BamHI R: 5′- GCAGGATCCCAACCTACTGATAAGGAGGGAAGTAAGGCTGCTG-3′. The following PCR conditions were applied: 20 cycles of 98 °C for 10 s, 62 °C for 5 s, and 68 °C for 15 s. The amplicon was purified with the QIAquick PCR purification kit (Qiagen) and cloned into the pIZ/V5-His vector using the HindIII and BamHI enzymes (New England Biolabs). The codon-optimized oscar gene of wFur (Katsuma et al., 2022) was also cloned into the pIZ/V5-His vector using the KpnI and NotI enzymes (TaKaRa).
To verify the masculinizing function of Hmmasc, BmN-4 cells (4 × 105 cells per dish, diameter 35 mm) were transfected with 1 µg of each plasmid DNA (pIZ/V5-His having Hmmasc) using FuGENE HD (Promega, WI, USA), as described in Katsuma et al., 2022. To clarify whether Hm-oscar suppressed the function of Hmmasc, 1 µg of plasmid DNA (pIZ/V5-His bearing either Hmmasc or Hm-oscar and non-inserted vector as negative control) was transfected to the BmN-4 cells using FuGENE HD (Promega). Three days after transfection, the cells were collected and subjected to RNA extraction via TRI REAGENT (Molecular Research Center Inc, USA) and cDNA construction with AMV transcriptase (TaKaRa). The degree of masculinization in the BmN-4 cells (default female-type sex determination) was verified by quantifying the expression levels of BmIMPM (Suzuki et al., 2010), which is involved in male-specific sex determination cascades, using primers BmIMP_F: 5′-ATGCGGGAAGAAGGTTTTATG-3′ and BmIMP_R: 5′-TCATCCCGCCTCAGACGATTG-3′, as described in Fukui et al., 2015. BmRp49 gene was also amplified as a control gene using primers rp49_F: 5′-CCCAACATTGGTTACGGTTC-3′ and rp49_R: 5′-GCTCTTTCCACGATCAGCTT-3′. Further interactions between Hm-oscar and masc genes derived from lepidopteran insects [i.e. Trilocha varians masc (Tvmasc), Spodoptera frugiperda masc (Sfmasc), B. mori masc (Bmmasc), O. furnacalis masc (Ofmasc), and Papilio machaon masc (Pmmasc)] (Katsuma et al., 2022) were assessed using the same procedures. The expression levels of BmIMPM were analyzed as described in Katsuma et al., 2022. In brief, the relative expression of BmIMPM (CT BmIMPM / CT BmRp49) was calculated for each experimental group. Then, relative expression of BmIMPM of each sample was normalized by setting the mean in the Hmmasc singly transfected group as 100. The normalized relative expression of BmIMPM was visualized using R v4.0 with ggplot2 (https://ggplot2.tidyverse.org/).
Quantification and statistical analysis
Request a detailed protocolThe number of surviving H. magnanima injected with either Hm-oscar, wmks, or GFP in each replicate were counted at the adult stage. The male ratios (number of adult males/numbers of all adults) under all conditions were compared using the Steel–Dwass test in R v4.0. The sex ratio bias was also assessed based on the total numbers of (i) male and female adults and (ii) male and female embryos under each condition using the binomial test in R v4.0.
Gene expression levels between chromosomes (RNA-seq), a Z-linked Hmtpi gene expression in H. magnanima (RT-qPCR), and normalized BmIMPM expression in BmN-4 cells under all conditions (RT-qPCR) were compared using the Steel–Dwass test in R v4.0.
A phylogenetic tree of Masc proteins used in this study was constructed based on maximum likelihood with bootstrap re-sampling of 1000 replicates using MEGA7 (Kumar et al., 2016).
Data availability
High-throughput sequencing data are available under accession numbers DRA018708 and PRJDB18169 (BioProject). All data generated during this study are included in the manuscript and a supporting Excel file. Further information and requests for resources and reagents are accessible from the lead contact, Hiroshi Arai (dazai39papilio@gmail.com/HArai@liverpool.ac.uk).
-
NCBI BioProjectID PRJDB18169. Transcriptome data for Homona magnanima.
-
NCBI BioProjectID PRJDB13118. Distinct effects of the male-killing bacteria, Wolbachia and Spiroplasma, and a partiti-like virus in the tea pest moth, Homona magnanima.
References
-
Mass-rearing and molecular studies in tortricidae pest insectsJournal of Visualized Experiments 181:e63737.https://doi.org/10.3791/63737
-
Diverse molecular mechanisms underlying microbe-inducing male killing in the moth Homona magnanimaApplied and Environmental Microbiology 89:e0209522.https://doi.org/10.1128/aem.02095-22
-
Conserved infections and reproductive phenotypes of Wolbachia symbionts in Asian tortrix mothsEnvironmental Microbiology Reports 16:e13219.https://doi.org/10.1111/1758-2229.13219
-
Two male-killing Wolbachia from Drosophila birauraia that are closely related but distinct in genome structureRoyal Society Open Science 11:231502.https://doi.org/10.1098/rsos.231502
-
Trimmomatic: a flexible trimmer for illumina sequence dataBioinformatics 30:2114–2120.https://doi.org/10.1093/bioinformatics/btu170
-
Near-optimal probabilistic RNA-seq quantificationNature Biotechnology 34:525–527.https://doi.org/10.1038/nbt.3519
-
Positive selection and horizontal gene transfer in the genome of a male-killing WolbachiaMolecular Biology and Evolution 39:msab303.https://doi.org/10.1093/molbev/msab303
-
Sex determination systems as the interface between male-killing bacteria and their hostsProceedings. Biological Sciences 289:20212781.https://doi.org/10.1098/rspb.2021.2781
-
Reproductive parasitism: maternally inherited symbionts in a biparental worldCold Spring Harbor Perspectives in Biology 7:a017699.https://doi.org/10.1101/cshperspect.a017699
-
Extended genomes: symbiosis and evolutionInterface Focus 7:20170001.https://doi.org/10.1098/rsfs.2017.0001
-
Opposite sex-specific effects of Wolbachia and interference with the sex determination of its host Ostrinia scapulalisProceedings. Biological Sciences 271:251–258.https://doi.org/10.1098/rspb.2003.2604
-
A male-killing gene encoded by a symbiotic virus of DrosophilaNature Communications 14:1357.https://doi.org/10.1038/s41467-023-37145-0
-
Masc-induced dosage compensation in silkworm cultured cellsFEBS Open Bio 9:1573–1579.https://doi.org/10.1002/2211-5463.12698
-
A Wolbachia factor for male killing in lepidopteran insectsNature Communications 13:6764.https://doi.org/10.1038/s41467-022-34488-y
-
MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasetsMolecular Biology and Evolution 33:1870–1874.https://doi.org/10.1093/molbev/msw054
-
Misdirection of dosage compensation underlies bidirectional sex-specific death in Wolbachia-infected Ostrinia scapulalisInsect Biochemistry and Molecular Biology 66:72–76.https://doi.org/10.1016/j.ibmb.2015.10.001
-
BookSex determination cascade in insects: a great treasure house of alternative splicingIn: Kobayashi K, Kitano T, Iwao Y, Kondo M, editors. Reproductive and Developmental Strategies: The Continuity of Life. Springer. pp. 267–288.https://doi.org/10.1007/978-4-431-56609-0_13
-
Studies on sex pheromones of Pyralididae. III. The inheritance of the abnormal sex ratio condition in a strain of the almond moth, Cadra cautella Walker (Phycitinae)Botyu-Kagaku = Scientific Insect Control 35:11–21.
-
Wolbachia: master manipulators of invertebrate biologyNature Reviews. Microbiology 6:741–751.https://doi.org/10.1038/nrmicro1969
Article and author information
Author details
Funding
Japan Society for the Promotion of Science (19J13123)
- Hiroshi Arai
Japan Society for the Promotion of Science (21J00895)
- Hiroshi Arai
Japan Society for the Promotion of Science (22K14902)
- Hiroshi Arai
Japan Society for the Promotion of Science (23H02229)
- Daisuke Kageyama
Japan Society for the Promotion of Science (24H02293)
- Daisuke Kageyama
Japan Society for the Promotion of Science (22H00366)
- Susumu Katsuma
Japan Society for the Promotion of Science (24H02289)
- Susumu Katsuma
Japan Society for the Promotion of Science (21KK0105)
- Hiroshi Arai
- Maki N Inoue
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
We thank Greg Hurst (Institute of Infection, Veterinary and Ecological Sciences, University of Liverpool, Liverpool, UK), and Takeshi Suzuki (Graduate School of Bio-Applications and Systems Engineering, Tokyo University of Agriculture and Technology, Tokyo, Japan) for their kind advice. We acknowledge support from the Japan Society for the Promotion of Science (JSPS) Research Fellowships for Young Scientists [Grant Number 19J13123 and 21J00895 to H Arai], JSPS Grant-in-Aid for Scientific Research [Grant Number 22K14902 to H Arai, 23H02229 and 24H02293 to D Kageyama, 22H00366 and 24H02289 to S Katsuma], JSPS Fund for the Promotion of Joint International Research (Fostering Joint International Research (B)) [Grant Number 21KK0105 to H Arai and MN Inoue]. The wHm-t-infected H. magnanima was collected from Tea Research and Extension Station (Taoyuan City, Taiwan), and imported with permission from the Ministry of Agriculture, Forestry and Fisheries (No. 27 - Yokohama Shokubou 891 and No. 297 - Yokohama Shokubou 1326). All collaborators are presented as co-authors, and the results have been shared with the provider communities. Moreover, our group is committed to international scientific partnerships as well as institutional capacity building.
Version history
- Sent for peer review:
- Preprint posted:
- Reviewed Preprint version 1:
- Reviewed Preprint version 2:
- Reviewed Preprint version 3:
- Version of Record published:
Cite all versions
You can cite all versions using the DOI https://doi.org/10.7554/eLife.101101. This DOI represents all versions, and will always resolve to the latest one.
Copyright
© 2024, Arai et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 1,447
- views
-
- 100
- downloads
-
- 10
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 6
- citations for umbrella DOI https://doi.org/10.7554/eLife.101101
-
- 1
- citation for Reviewed Preprint v2 https://doi.org/10.7554/eLife.101101.2
-
- 3
- citations for Version of Record https://doi.org/10.7554/eLife.101101.4
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Prophage-encoded Hm-oscar gene recapitulates Wolbachia-induced male-killing in the tea tortrix moth Homona magnanima
eLife 13:RP101101.
https://doi.org/10.7554/eLife.101101.4




{kind=link}
{kind=link}
{kind=link}
{kind=link}