Contraction-induced endocardial id2b plays a dual role in regulating myocardial contractility and valve formation
- Department of Cardiology, Center for Genetic Medicine, The Fourth Affiliated Hospital of School of Medicine, and International School of Medicine, International Institutes of Medicine, Zhejiang University, China
- Division of Medical Genetics and Genomics, The Children's Hospital, Zhejiang University School of Medicine, National Clinical Research Center for Child Health, China
- Institute of Genetics, Zhejiang University School of Medicine, China
- TaiKang Medical School (School of Basic Medical Sciences), Wuhan University, China
- Hubei Provincial Key Laboratory of Developmentally Originated Disease, China
- Department of Biophysics, and Kidney Disease Center of the First Affiliated Hospital, Zhejiang University School of Medicine, China
- Liangzhu Laboratory, Zhejiang University Medical Center, China
- Zhejiang Provincial Key Laboratory for Cancer Molecular Cell Biology, Life Sciences Institute, Zhejiang University, China
eLife Assessment
This study presents a valuable finding that the biomechanical force of heart contractility is required for robust endocardial id2b expression, which in return promotes valve development and myocardial function through upregulation of Neuregulin 1. The data were collected and analyzed using solid methodology and can be used as a starting point for deeper mechanistic insights into the genetic programs regulating endocardial-myocardial crosstalk during heart development.
https://doi.org/10.7554/eLife.101151.3.sa0Significance of the findings:
Valuable: Findings that have theoretical or practical implications for a subfield
- Landmark
- Fundamental
- Important
- Valuable
- Useful
Strength of evidence:
Solid: Methods, data and analyses broadly support the claims with only minor weaknesses
- Exceptional
- Compelling
- Convincing
- Solid
- Incomplete
- Inadequate
During the peer-review process the editor and reviewers write an eLife Assessment that summarises the significance of the findings reported in the article (on a scale ranging from landmark to useful) and the strength of the evidence (on a scale ranging from exceptional to inadequate). Learn more about eLife Assessments
Abstract
Biomechanical cues play an essential role in sculpting organ formation. Comprehending how cardiac cells perceive and respond to biomechanical forces is a biological process with significant medical implications that remains poorly understood. Here, we show that biomechanical forces activate endocardial id2b (inhibitor of DNA-binding 2b) expression, thereby promoting cardiac contractility and valve formation in zebrafish. Taking advantage of the unique strengths of zebrafish, particularly the viability of embryos lacking heartbeats, we systematically compared the transcriptomes of hearts with impaired contractility to those of control hearts. This comparison identified id2b as a gene sensitive to blood flow. By generating a knock-in reporter line, our results unveiled the presence of id2b in the endocardium, and its expression is sensitive to both pharmacological and genetic perturbations of contraction. Furthermore, id2b loss-of-function resulted in progressive heart malformation and early lethality. Combining RNA-seq analysis, electrophysiology, calcium imaging, and echocardiography, we discovered profound impairment in atrioventricular (AV) valve formation and defective excitation-contraction coupling in id2b mutants. Mechanistically, deletion of id2b reduced AV endocardial cell proliferation and led to a progressive increase in retrograde blood flow. In the myocardium, id2b directly interacted with the bHLH component tcf3b (transcription factor 3b) to restrict its activity. Inactivating id2b unleashed its inhibition on tcf3b, resulting in enhanced repressor activity of tcf3b, which subsequently suppressed the expression of nrg1 (neuregulin 1), an essential mitogen for heart development. Overall, our findings identify id2b as an endocardial cell-specific, biomechanical signaling-sensitive gene, which mediates intercellular communications between endocardium and myocardium to sculpt heart morphogenesis and function.
Introduction
The heart develops with continuous contraction, and biomechanical cues play an essential role in cardiac morphogenesis (Duchemin et al., 2019; Sidhwani and Yelon, 2019). Blood flow is directly sensed by the surrounding endocardium, which undergoes multiscale remodeling during zebrafish heart development. In the atrioventricular canal (AVC) endocardium, oscillatory flow promotes valvulogenesis through transient receptor potential (TRP) channel-mediated expression of Krüppel-like factor 2a (klf2a) (Vermot et al., 2009; Heckel et al., 2015; Gálvez-Santisteban et al., 2019). Meanwhile, mechanical forces trigger ATP-dependent activation of purinergic receptors, inducing expression of nuclear factor of activated T cells 1 (nfatc1) and subsequent valve formation (Fukui et al., 2021). In the chamber endocardium, blood flow induces endocardial cells to adopt chamber- and region-specific cell morphology during cardiac ballooning (Dietrich et al., 2014). A recent study further emphasized that blood flow is essential for endocardial cell accrual in assembling the outflow tract (OFT) (Sidhwani et al., 2020). Beyond their role in endocardial cells, proper biomechanical cues are indispensable for shaping the myocardium. For instance, in contraction-compromised tnnt2a (Staudt et al., 2014) and myh6 (Peshkovsky et al., 2011) mutants, trabeculation is markedly reduced. Moreover, apart from the tissue-scale regulatory effect, the shape changes (Auman et al., 2007) and myofibril content (Lin et al., 2012) at the single-cardiomyocyte level are also sculpted by the interplay of contractility and blood flow in the developing heart.
In ventricular myocardium morphogenesis, biomechanical forces coordinate intra-organ communication between endocardial and myocardial cells by regulating bone morphogenetic protein (BMP), Nrg/Erbb, and Notch signaling. The Nrg-Erbb axis stands as one of the most extensively studied signaling pathways mediating cell-cell communications in the heart (Gassmann et al., 1995; Lee et al., 1995; Meyer and Birchmeier, 1995; Zhang et al., 2021). In particular, endocardial Notch activity induced by cardiac contraction promotes the expression of Nrg, which then secretes into the extracellular space, binding to Erbb2/4 receptor tyrosine kinases on cardiomyocyte and promoting their delamination (Lai et al., 2010; Rentschler et al., 2002; Liang et al., 2025). In agreement with the pivotal role of this signaling pathway in heart development, genetic mutations in zebrafish nrg2a and erbb2 result in severely compromised trabeculae formation (Liu et al., 2010; Rasouli and Stainier, 2017; Han et al., 2016).
The development of cardiomyocytes encompasses the specification of subcellular structure, metabolic state, gene expression profile, and functionality (Guo and Pu, 2020; Alvarez-Dominguez and Melton, 2022). The rhythmic contraction of cardiomyocytes relies on precisely regulated excitation-contraction coupling (E-C coupling), transducing electrical activity into contractile forces. This intricate signaling cascade involves membrane action potential, calcium signaling, and sarcomeric structure (Bers, 2002; Xu et al., 2024). Specifically, membrane depolarization triggers the opening of L-type calcium channel (LTCC), allowing calcium influx. The calcium signaling then activates the ryanodine receptor on the sarcoplasmic reticulum membrane, releasing additional calcium (Bers, 2002). E-C coupling is essential for heart development, as evidenced by the complete silence of the ventricle and reduced cardiomyocyte number in cacna1c (LTCC α1 subunit in zebrafish) mutant (Rottbauer et al., 2001). Beyond its role in modulating cardiac structure formation, previous studies indicate that Nrg-Erbb2 signaling is also necessary for cardiac function, as erbb2 mutants exhibit severely compromised fractional shortening and an immature conduction pattern (Liu et al., 2010; Samsa et al., 2015).
Id (inhibitor of DNA-binding) proteins belong to the helix-loop-helix (HLH) family and function as transcriptional repressors (Benezra et al., 1990). Notably, Id2 lacks a DNA-binding domain and forms a heterodimer with other bHLH proteins, acting in a dominant-negative manner (Wong et al., 2012). Id2 plays a crucial role in heart development, and its genetic deletion results in severe cardiac defects in mice (Jongbloed et al., 2011; Moskowitz et al., 2007; Moskowitz et al., 2011). In zebrafish, id2a and id2b are homologs of the mammalian Id2 gene. However, their expression pattern and function in the zebrafish heart remain largely unknown. In the present study, we identified that id2b is specifically expressed in endocardial cells of the developing heart, and its expression is regulated by cardiac contraction and blood flow. Genetic deletion of id2b led to impaired AV valve formation and reduced cardiac contractility. Therefore, id2b serves as a crucial mediator linking biomechanical cues to heart morphogenesis.
Results
Transcriptome analysis identifies id2b as a blood flow-sensitive gene
Blood circulation is dispensable for early embryonic development in zebrafish, presenting an ideal model to investigate biomechanical cues influencing heart morphogenesis. To identify genes affected by cardiac contraction or blood flow, we treated myl7:mCherry zebrafish embryos with tricaine to inhibit cardiac contractility from 72 hr post-fertilization (hpf) to 96 hpf. Hearts from control and tricaine-treated zebrafish embryos were manually collected under a fluorescence stereoscope as previously reported (Burns and MacRae, 2006). Subsequently, approximately 1000 hearts from each group were subjected to RNA-seq (Figure 1A). A total of 4530 genes with differential expression were identified, comprising 2013 upregulated and 2517 downregulated genes. With a specific focus on identifying key transcription factors (TFs) affected by perturbing biomechanical forces, differentially expressed genes (DEGs) encoding TFs were enriched into signaling pathways through KEGG analysis. Interestingly, our analysis identified several pathways known to be involved in heart development, including the transforming growth factor beta signaling and Notch signaling pathways (Figure 1B). In particular, the scaled expression levels of the top 6 DEGs (|log2FC|≥0.585), exhibiting up- or downregulation, were listed (Figure 1C). Intriguingly, Id2 has been shown to regulate murine AV valve formation, a process notably influenced by alterations in blood flow directionality. Moreover, loss of Id2 leads to malformation of both the arterial and venous poles of the heart and disrupts AV valve morphogenesis (Jongbloed et al., 2011; Moskowitz et al., 2011). Therefore, we interrogated the expression and function of id2b in developing embryos.
Figure 1
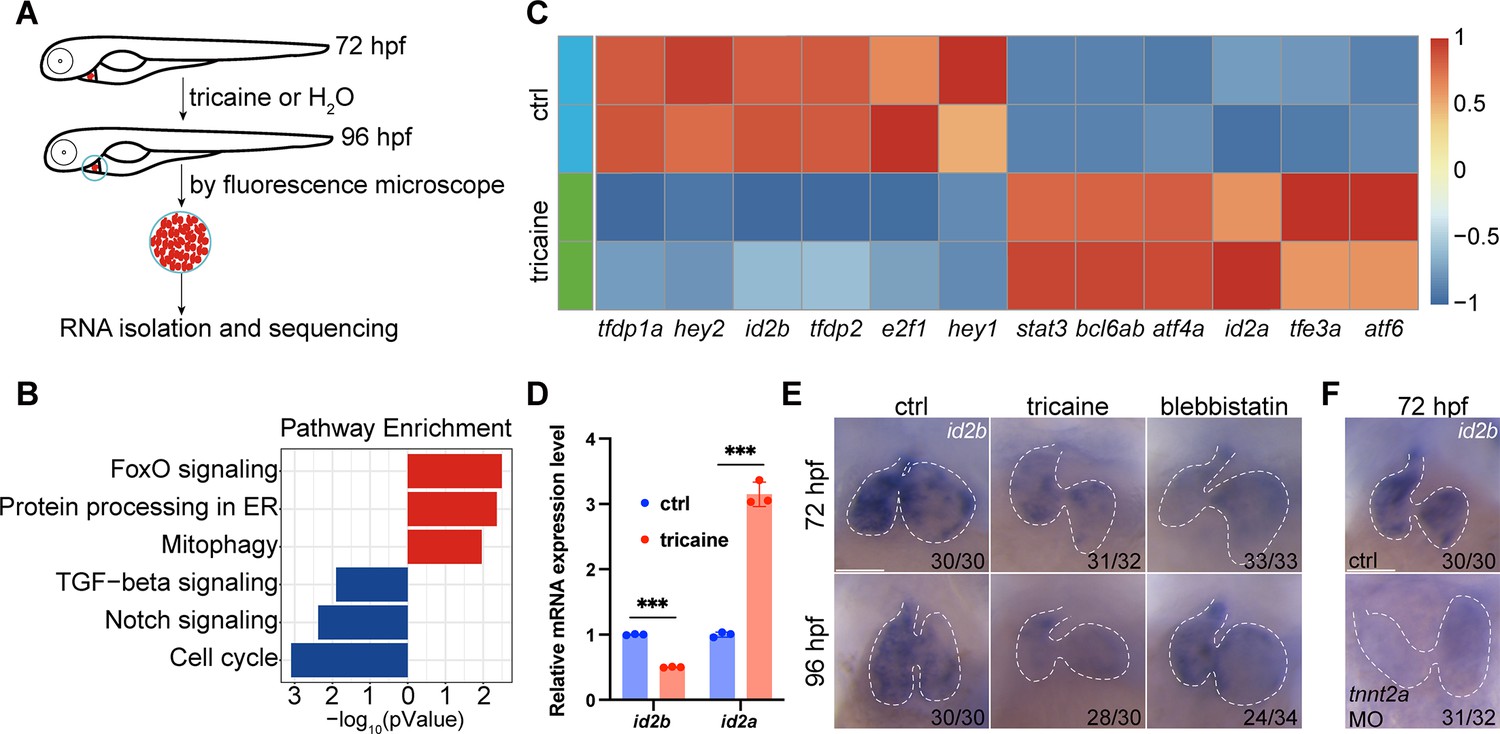
Identification of id2b as a blood flow-sensitive gene.
(A) Schematic showing the experimental procedures, including treatment, heart collection, and RNA-sequencing of zebrafish embryonic hearts. (B) KEGG enrichment analysis depicting differentially expressed genes encoding transcription factors and transcriptional regulators between control (ctrl) and tricaine-treated embryonic hearts. Red and blue rectangles represent upregulated and downregulated gene sets, respectively. |log2fold change|≥0.585, adjusted p-value<0.1. Each replicate contains approximately 1000 hearts. (C) Heatmap exhibiting representative genes from KEGG pathways mentioned in (B). (D) Quantitative real-time PCR (qRT-PCR) analysis of id2b and id2a mRNA in control and tricaine-treated embryonic hearts. Data were normalized to the expression of actb1. Each sample contains ~1000 embryonic hearts. N=3 biological replicates. (E) In situ hybridization of id2b in 72 hr post-fertilization (hpf) and 96 hpf ctrl, tricaine (1 mg/mL), and blebbistatin (10 μM)-treated embryos. Numbers at the bottom of each panel indicate the ratio of representative staining. (F) In situ hybridization showing reduced id2b expression in tnnt2a morpholino-injected embryos at 72 hpf compared to control. Data are presented as mean ± s.e.m. Unpaired two-tailed Student’s t-tests were used to determine statistical significance. ***p<0.001. Scale bars, 50 μm.
Quantitative real-time PCR (qRT-PCR) analysis of purified embryonic hearts revealed a significant reduction in id2b mRNA levels and an increase in id2a levels following tricaine treatment from 72 to 96 hpf compared to controls (Figure 1D). Furthermore, in situ hybridization was performed to visualize id2b expression under tricaine or 10 µM blebbistatin (an inhibitor of sarcomeric function and cardiac contractility) treatment from 48 to 72 or from 72 to 96 hpf as previously described (Gálvez-Santisteban et al., 2019). Consistently, our results showed a reduction in id2b signal in contraction-deficient hearts compared to the control (Figure 1E). In cardiomyocytes, tnnt2a encodes a key sarcomeric protein essential for contractility. Similarly, injection of a previously characterized tnnt2a morpholino (Sehnert et al., 2002) at the one-cell stage also led to compromised contraction and diminished expression of id2b (Figure 1F). Taken together, these results indicate that biomechanical cues are essential for activating id2b in embryonic hearts.
Visualization of the spatiotemporal expression of id2b in developing embryos
Due to technical challenges in visualizing the cell-type-specific expression of id2b in the developing heart using whole-mount in situ hybridization, we employed an intron targeting-mediated approach (Li et al., 2015) to generate a knock-in id2b:eGFP reporter line. This method allowed us to achieve specific labeling without perturbing the integrity and function of the endogenous gene (Li et al., 2015; Figure 2A). Comparison of id2b:eGFP fluorescence with in situ hybridization at 24, 48, and 72 hpf revealed that the reporter signal closely recapitulates the endogenous id2b expression pattern. The fluorescence was notably enriched in the heart, brain, retina, and notochord (Figure 2B), mirroring observations from a previously reported id2b transgenic line generated through BAC-mediated recombination (Förster et al., 2017).
Figure 2
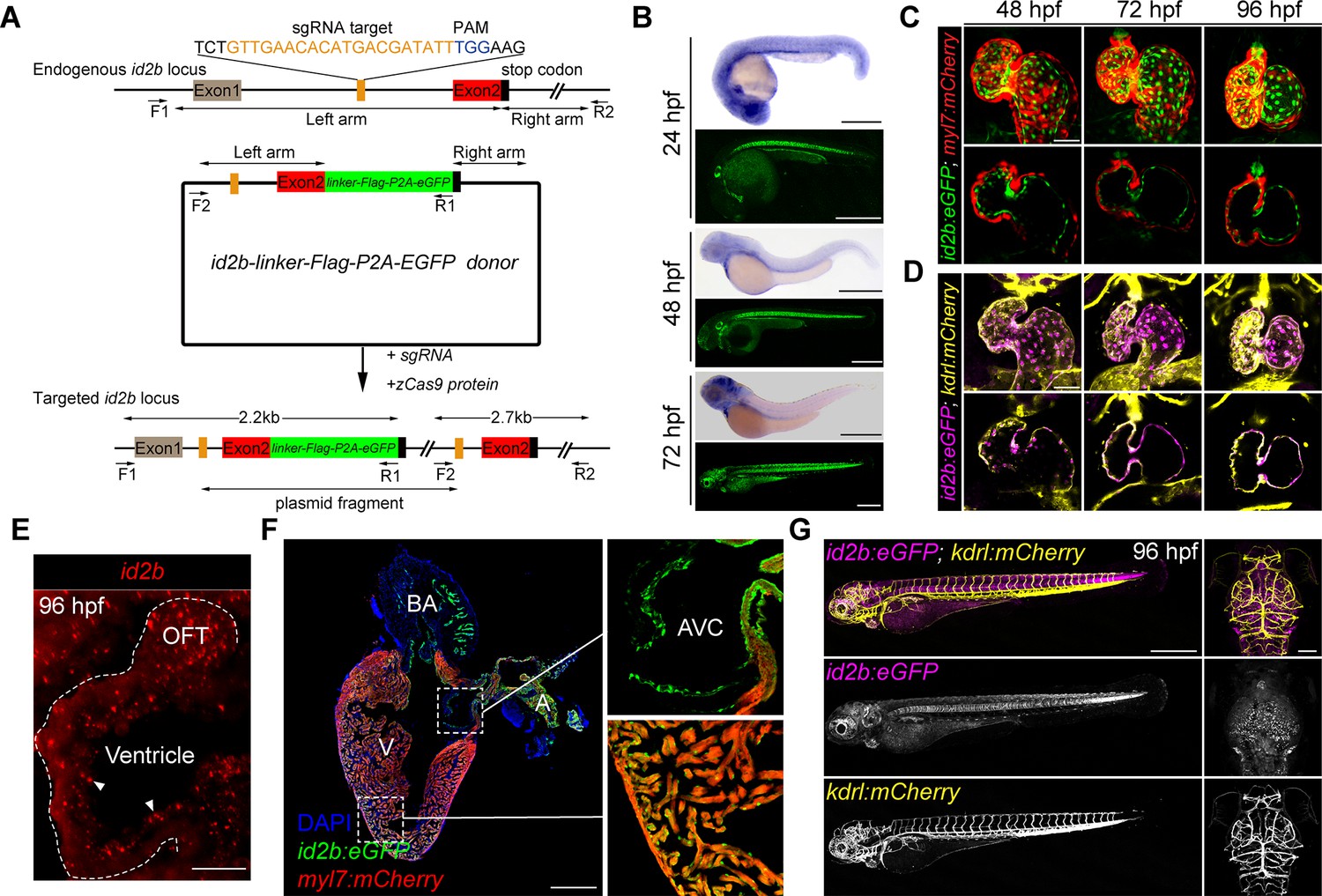
The spatiotemporal expression of id2b.
(A) Schematic of the intron targeting-mediated eGFP knock-in at the id2b locus using the CRISPR-Cas9 system. The sgRNA targeting sequence and the protospacer adjacent motif (PAM) sequence are shown in orange and blue, respectively. The donor plasmid comprises left and right arm sequences and a linker-FLAG-P2A-eGFP cassette denoted by black lines with double arrows and green box, respectively. The linker-FLAG-P2A-eGFP cassette was integrated into the id2b locus upon co-injection of the donor plasmid with sgRNA and zCas9 protein, enabling detection by PCR using two pairs of primers (F1, R1 and F2, R2) - the former length yielding a length of about 2.2 kb and the latter about 2.7 kb. (B) Zebrafish id2b expression pattern, as indicated by in situ hybridization of embryos at designated time points, was consistent with the fluorescence localization of id2b:eGFP, revealing expression in the heart, brain, retina, notochord, pronephric duct, and other tissues. (C) Maximum intensity projections (top) and confocal sections (bottom) of id2b:eGFP; Tg(myl7:mCherry) hearts at designated time points. (D) Maximum intensity projections (top) and confocal sections (bottom) of id2b:eGFP; Tg(kdrl:mCherry) embryos at specific time points. Magenta, id2b:eGFP; yellow, kdrl:mCherry. (E) RNAscope analysis of id2b in 96 hr post-fertilization (hpf) embryonic heart. White dashed line outlines the heart. OFT, outflow tract. (F) Immunofluorescence of adult id2b:eGFP; Tg(myl7:mCherry) heart section (left panel). Enlarged views of boxed areas are shown in the right panel. Green, eGFP; red, mCherry; blue, DAPI. BA, bulbus arteriosus; V, ventricle; A, atrium; AVC, atrioventricular canal. (G) Confocal z-stack maximum intensity projection of id2b:eGFP;Tg(kdrl:mCherry) embryos at 96 hpf showing the whole body (lateral view) and the head (top view). Scale bars, 500 μm (B, F, left, and G), 50 μm (C and D), 25 μm (E), 100 μm (G, right).
To further elucidate the spatiotemporal expression of id2b in developing hearts, we crossed id2b:eGFP with myl7:mCherry or kdrl:mCherry, labeling cardiomyocytes or endocardial cells, respectively. Confocal images revealed minimal, if any, presence of id2b:eGFP in myl7:mCherry+ cardiomyocytes (Figure 2C). In sharp contrast, clear co-localization between id2b:eGFP and kdrl:mCherry was evident at 48, 72, and 96 hpf (Figure 2D). Endocardial localization of id2b was further confirmed by RNAscope analysis (Figure 2E). In adult hearts, id2b:eGFP fluorescence was enriched in the chamber endocardium and the endothelium lining AVC, OFT, and bulbus arteriosus (Figure 2F). Interestingly, there was an absence of id2b:eGFP signal in kdrl:mCherry+ endothelial cells in trunk blood vessel and brain vasculature (Figure 2G). Collectively, these results indicate that id2b is expressed in endocardial cells across different developmental stages.
BMP signaling and cardiac contraction regulate id2b expression
Taking advantage of live imaging on developing embryos, we explored the in vivo dynamics of id2b in response to biomechanical force at single-cell resolution. When embryos were treated with tricaine or blebbistatin, the intensity of id2b:eGFP in atrial and ventricular endocardium was significantly reduced (Figure 3A and B). Similarly, injection of tnnt2a morpholino also markedly suppressed id2b:eGFP signal (Figure 3A and B), in agreement with the results obtained from in situ hybridization. Strikingly, the reduction in fluorescence intensity was particularly pronounced in AVC endothelial cells (Figure 3A and B, arrowheads).
Figure 3 with 1 supplement see all
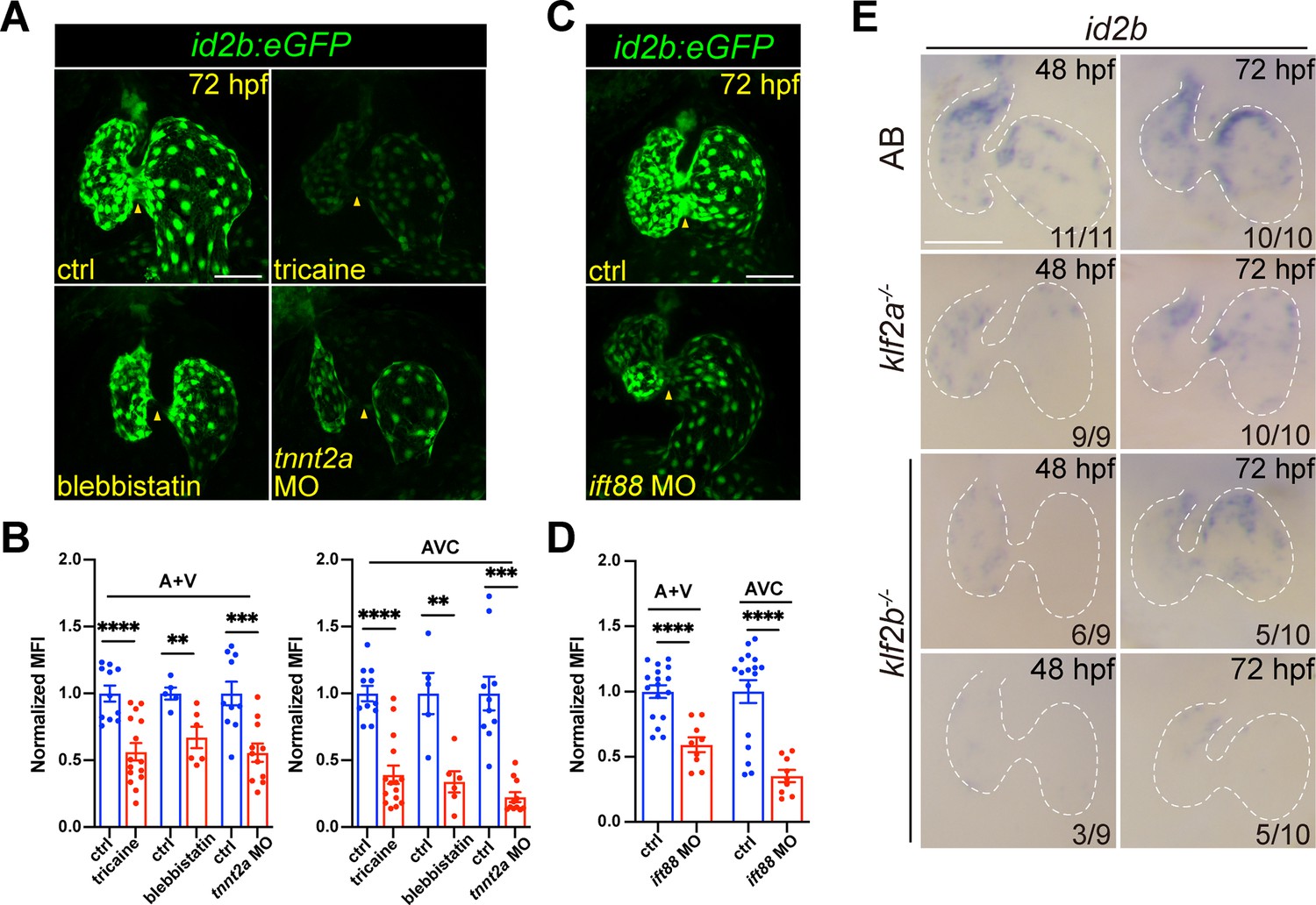
Cardiac contraction promotes endocardial id2b expression through primary cilia.
(A) Representative confocal z-stack (maximal intensity projection) of id2b:eGFP embryos under different conditions: control, tricaine-treated, blebbistatin-treated, and tnnt2a morpholino-injected. Images were captured using the same magnification. (B) Quantification of mean fluorescence intensity (MFI) of id2b:eGFP in the working myocardium (atrium and ventricle, A+V) and atrioventricular canal (AVC) in (A). Data normalized to the MFI of control hearts. n=(11, 15) (ctrl versus tricaine); n=(5, 6) (ctrl versus blebbistatin); n=(10,11) (ctrl versus tnnt2a MO). (C) Representative confocal z-stack (maximal intensity projection) of control and ift88 morpholino-injected id2b:eGFP embryos. (D) Normalized MFI of id2b:eGFP in the working myocardium (A+V) and AVC in (C). n=(17, 9). (E) Whole-mount in situ hybridization showing id2b expression in control, klf2a-/-, and klf2b-/- embryos at 48 hr post-fertilization (hpf) and 72 hpf. Numbers at the bottom of each panel indicate the ratio of representative staining. Data are presented as mean ± s.e.m. Unpaired two-tailed Student’s t-tests were used to determine statistical significance. **p<0.01, ***p<0.001, ****p<0.0001. Scale bars, 50 μm.
We then explored how cardiac contraction modulated id2b expression. Given that endocardial cells can sense blood flow through primary cilia (Li et al., 2020; Nauli et al., 2008), we used a characterized morpholino (Li et al., 2020) to knock down ift88, an intraflagellar transporter essential for primary cilia formation. Previously, work demonstrated a complete loss of primary cilia in endocardial cells upon ift88 knockdown (Li et al., 2020). As expected, a significant decrease in id2b:eGFP intensity was observed in the chamber and AVC endocardium of ift88 morphants compared to control (Figure 3C and D), suggesting that biomechanical forces promote the expression of id2b via primary cilia. In the developing heart, a central hub for mediating biomechanical cues is the Klf2 gene, which includes the klf2a and klf2b paralogues in zebrafish (Vermot et al., 2009; Heckel et al., 2015; Gálvez-Santisteban et al., 2019; Li et al., 2020; Rasouli et al., 2018). Previous studies in mammals and zebrafish have highlighted the essential role of Klf2 TF activity in cardiac valve and myocardial wall formation (Vermot et al., 2009; Rasouli et al., 2018). As a flow-responsive gene, klf2a expression has been observed throughout the entire endocardium, evidenced by mRNA expression and transgenic studies (Vermot et al., 2009; Heckel et al., 2015). Interestingly, in situ hybridization on 48 and 72 hpf klf2a-/- and klf2b-/- embryos unveiled a drastic decrease in id2b expression compared with wild-type zebrafish (Figure 3E), supporting the notion that klf2-mediated biomechanical signaling is essential for activating id2b expression.
Given that id2b has been reported as a target gene of BMP signaling, we explored whether BMP also played a role in regulating id2b expression. To this end, we knocked down bmp2b, bmp4, and bmp7a in one-cell stage embryos. Live imaging at 24 hpf revealed a significant reduction in id2b:eGFP fluorescence signal in morpholino-injected hearts compared to controls (Figure 3—figure supplement 1A and B), suggesting that id2b is a target gene of BMP signaling during early embryonic development. Similarly, treatment with the BMP inhibitor Dorsomorphin from 10 to 24 hpf resulted in a marked decrease in id2b:eGFP signal (Figure 3—figure supplement 1C and D). Considering that heartbeats in zebrafish commence at approximately 22 hpf, we treated embryos with Dorsomorphin from 24 to 48 hpf or from 36 to 60 hpf. While the number of endocardial cells was slightly reduced upon Dorsomorphin exposure as previously reported (Dietrich et al., 2014), surprisingly, quantification of the average id2b:eGFP fluorescence intensity in individual endocardial cells revealed no significant differences between Dorsomorphin and DMSO-treated controls (Figure 3—figure supplement 1C and D).
We further visualized BMP activity using the BRE:d2GFP reporter line. Confocal images revealed strong fluorescence in the myocardium at 72 hpf, with minimal signal present in the endocardium except for the AVC endothelium (Figure 3—figure supplement 1E). Moreover, after tricaine treatment, endocardial BRE:d2GFP slightly increased (Figure 3—figure supplement 1E), as opposed to the reduced id2b:eGFP signal (Figure 3A and B). Likewise, endocardial BRE:d2GFP intensity was barely affected after completely blocking contraction with tnnt2a MO injection (Figure 3—figure supplement 1E). These observations align with previous work using pSmad-1/5/8 as a readout of BMP activity, indicating that endocardial BMP signaling is independent of blood flow (Dietrich et al., 2014). Collectively, these results suggest that id2b expression is regulated by both BMP and biomechanical signaling, with the relative contribution of each pathway varying across developmental stages.
Compromised AV valve formation in id2b mutants
To investigate the role of the contractility-id2b axis in zebrafish heart development, we generated a loss-of-function mutant line using CRISPR/Cas9. A pair of sgRNAs designed to target exon 1 was injected with zCas9 protein into one-cell stage embryos. Consequently, we identified a mutant allele with a 157 bp truncation, leading to the generation of a premature stop codon (Figure 4A, left). In id2b mutants (id2b-/-), the expression levels of id2b were significantly decreased, while id2a expression levels were increased compared to id2b+/+ siblings (Figure 4A, right). The overall morphology of id2b-/- remained unaltered at 72 and 96 hpf (Figure 4B). However, id2b-/- zebrafish experienced early lethality starting around 31 weeks post-fertilization (Figure 4C). Strikingly, pericardial edema was observed in 20% (9/45) of adult id2b-/- zebrafish (Figure 4D, top). Upon dissecting hearts from these id2b-/- zebrafish, a prominent enlargement in the atrium with a smaller ventricle was detected (Figure 4D, bottom), which has been characterized as cardiomyopathy in zebrafish (Weeks et al., 2024; Kamel et al., 2021a). Histological analysis further revealed malformation in the AV valves of these id2b-/- mutants compared to controls (Figure 4E, right). Specifically, we noted that the superior and inferior leaflets were significantly thinner, comprising only one to two layers of cells in id2b-/- zebrafish with an enlarged atrium. This was in sharp contrast to id2b+/+ zebrafish, which exhibited multilayers of cells (Figure 4E, left). Subsequent examination of the remaining 80% of id2b-/- zebrafish (36/45) that did not display prominent pericardial edema also revealed AV valve malformation, albeit to a lesser extent (Figure 4E, middle).
Figure 4 with 1 supplement see all
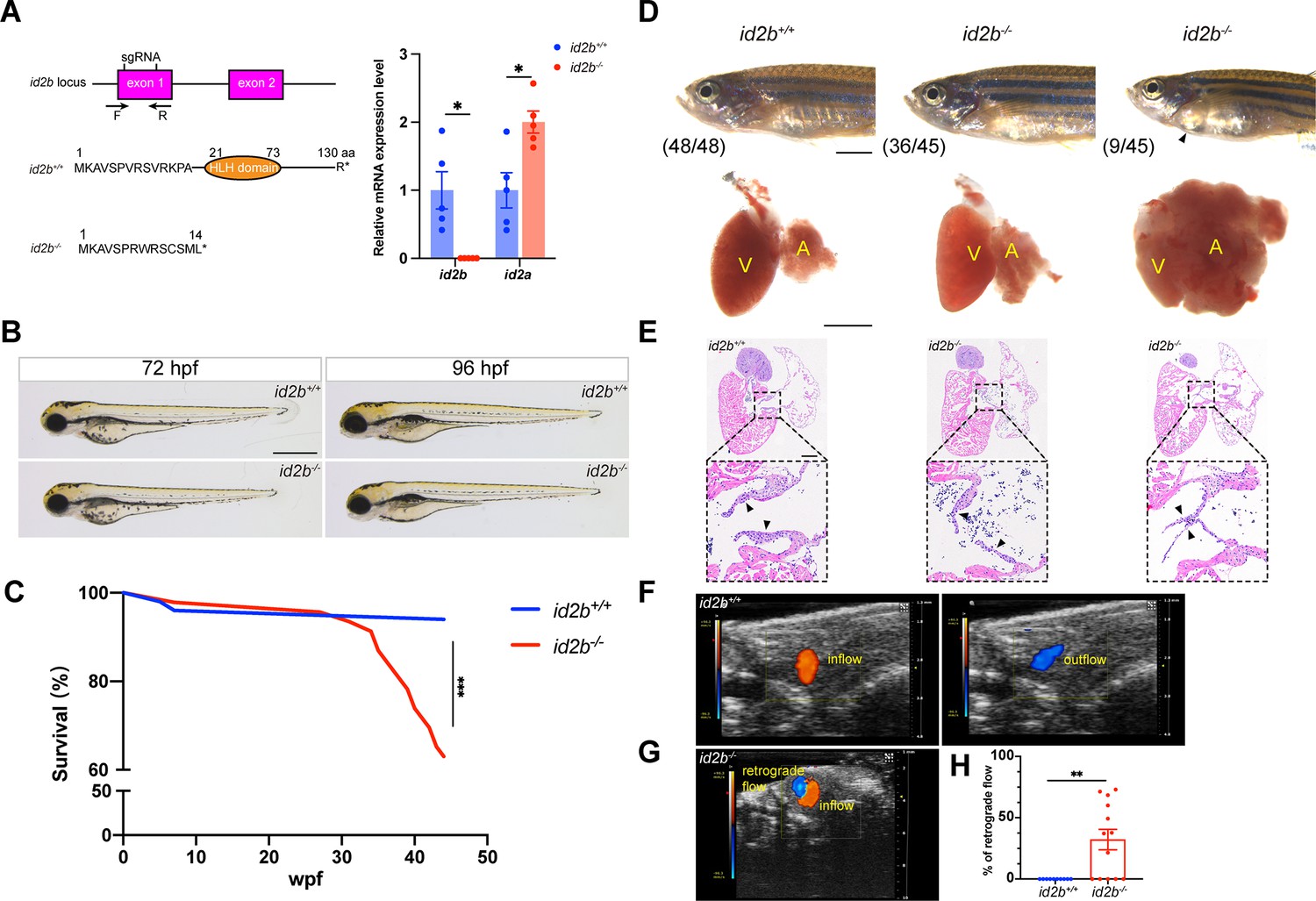
id2b-/- adults exhibit thinner atrioventricular valve leaflets and prominent retrograde blood flow.
(A) Two sgRNAs, represented by short vertical lines, were designed to create id2b-/- mutants. Co-injection of the two sgRNAs with zCas9 protein induces a 157 bp truncation in the exon 1 of id2b, which can be detected by genotyping primers marked with arrows. This genetic modification leads to the formation of a premature stop codon and the subsequent loss of the helix-loop-helix (HLH) domain. Right, quantitative real-time PCR (qRT-PCR) analysis of id2b and id2a mRNA levels in id2b+/+ and id2b-/- adult hearts. (B) No discernible morphological differences were observed between id2b+/+ and id2b-/- larvae at both 72 hr post-fertilization (hpf) and 96 hpf. (C) Kaplan-Meier survival curve analysis and log-rank test of id2b+/+ (n=50) and id2b-/- (n=46). Wpf, weeks post-fertilization. (D) Pericardial edema and an enlarged atrium are evident in a subset of id2b-/- adults. V, ventricle; A, atrium. (E) id2b-/- adults developed thinner atrioventricular valve leaflets (denoted by arrowheads) compared to id2b+/+. Enlarged views of boxed areas are shown in the bottom panels. (F, G) Echocardiograms of adult id2b+/+ (F) and id2b-/- (G) hearts. Unidirectional blood flow was observed in the id2b+/+ heart, while retrograde blood flow was evident in the id2b-/- heart. (H) Ratio of retrograde flow area over inflow area shows a significant increase in retrograde flow in id2b-/- (n=13) compared to id2b+/+ (n=10). Data are presented as mean ± s.e.m. Unpaired two-tailed Student’s t-tests were used to determine statistical significance. *p<0.05, **p<0.01, ***p<0.001. Scale bars, 500 μm (B and D, bottom), 2 mm (D, top), 200 μm (E).
To further interrogate the effect of id2b inactivation on AV valve formation and function, we analyzed the number of AVC endothelial cells using kdrl:nucGFP. At 96 hpf, a reduced number of kdrl:nucGFP+ cells were detected in the AVC region of id2b-/- embryos compared with id2b+/+ (Figure 4—figure supplement 1A and B). In contrast, the number of atrial and ventricular endocardial cells did not differ between id2b-/- and id2b+/+ (Figure 4—figure supplement 1C and D). Subsequently, we assessed hemodynamic flow by conducting time-lapse imaging of red blood cells labeled by gata1:dsred. Surprisingly, the pattern of hemodynamics was largely preserved in id2b-/- embryos compared to id2b+/+ siblings at 96 hpf (Figure 4—figure supplement 1E, Videos 1 and 2), suggesting that the reduced number of endocardial cells in the AVC region was not sufficient to induce functional defects. Additionally, we performed echocardiography to analyze blood flow in adult zebrafish as previously described (Gunawan et al., 2020). In id2b-/- hearts, prominent retrograde blood flow was detected in the AVC region (8/13) (Figure 4G), while unidirectional blood flow was observed in id2b+/+ (10/10) (Figure 4F). Quantification analysis showed ~32% retrograde blood flow in id2b-/-, compared to 0% in id2b+/+ zebrafish (Figure 4H). Consistently, the superior and inferior leaflets were much thinner in id2b-/- exhibiting retrograde flow compared with control fish (Figure 4—figure supplement 1F). Overall, these histological and functional analyses indicate that id2b deletion leads to progressive defects in AV valve morphology and hemodynamic flow.
Video 1
96 hr post-fertilization (hpf) id2b+/+ larvae displayed unidirectional blood flow in the atrioventricular (AV) canal.
Scale bar, 50 μm.
Video 2
96 hr post-fertilization (hpf) id2b-/- larvae displayed unidirectional blood flow in the atrioventricular (AV) canal.
Scale bar, 50 μm.
id2b deletion perturbs calcium signaling and contractile function in the myocardium
Although similar defects in AV valve formation have been reported in both klf2a and nfatc1 mutants, they do not display noticeable pericardial edema at the adult stage, nor do they experience early lethality (Vermot et al., 2009; Li et al., 2020; Rasouli et al., 2018; Gunawan et al., 2020; Novodvorsky et al., 2015). Therefore, we sought to investigate whether other cardiac properties have also been affected by id2b loss-of-function. To this end, we employed RNA-seq analysis on purified embryonic id2b-/- and id2b+/+ hearts (Figure 5—figure supplement 1). As expected, enrichment analysis of DEGs demonstrated that the top-ranked anatomical structures affected by id2b deletion included the heart valve, the compact layer of ventricle, and the AVC (Figure 5—figure supplement 1A). Interestingly, id2b inactivation also impacted phenotypes such as cardiac muscle contraction and heart contraction (Figure 5—figure supplement 1B). Therefore, we investigated cardiac contractile function through time-lapse imaging on the myl7:mCherry background. At 72 and 120 hpf, a significant decrease in cardiac function was observed in id2b-/- compared with id2b+/+ (Figure 5A–C, Figure 5—figure supplement 2A–C). Similarly, echocardiography analysis showed that the contractile function in adult id2b-/- heart was dramatically reduced compared with age-matched id2b+/+ (Figure 5D and F). These functional defects in id2b-deleted hearts could not be attributed to differences in cardiomyocyte number, as we counted cardiomyocytes using the myl7:H2A-mCherry line and found no apparent changes between id2b-/- and id2b+/+ embryos at 72 and 120 hpf (Figure 5—figure supplement 2D and E). Similarly, id2b-/- also developed regular trabecular structures (Figure 5—figure supplement 2F). Through α-actinin immunostaining, we observed similar sarcomeric structures in id2b-/- and control cardiomyocytes at 72 hpf and adult stages (115 dpf) (Figure 5—figure supplement 2G), corroborating that the reduced contractility in id2b-depleted heart was independent of structural defects.
Figure 5 with 2 supplements see all
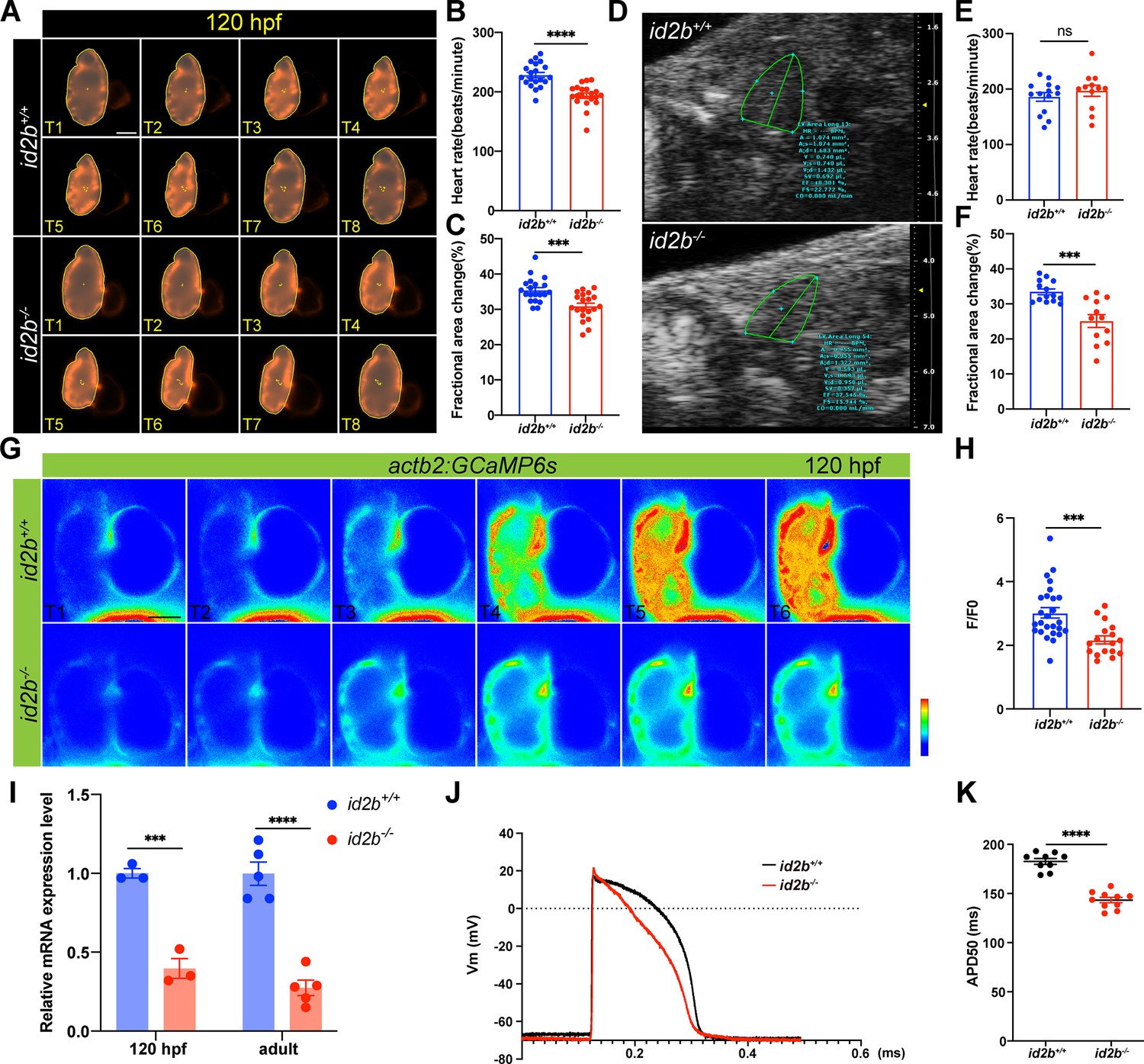
Reduced cardiac contractile function and compromised calcium handling in id2b-/- mutants.
(A) Time-lapse imaging (from T1 to T8) illustrates the cardiac contraction-relaxation cycle of 120 hr post-fertilization (hpf) id2b+/+ and id2b-/- hearts carrying myl7:mCherry. (B and C) id2b-/- larvae (n=20) display a significant decrease in heart rate and fractional area change compared to id2b+/+ (n=20). (D) Echocardiograms of adult id2b+/+ and id2b-/- hearts. (E and F) id2b-/- fish (n=12) exhibit reduced cardiac contractile function with preserved heart rate compared to id2b+/+ (n=14). (G) Time-lapse imaging illustrates the calcium dynamics of 120 hpf id2b+/+ and id2b-/- hearts carrying actb2:GCaMP6s. (H) Ratio of maximal fluorescence intensity (F) over basal fluorescence intensity (F0) of GCaMP6s signal. n=(26, 17). (I) Quantitative real-time PCR (qRT-PCR) analysis of cacnα1c mRNA in id2b+/+ and id2b-/- hearts at 120 hpf (N=3 biological replicates, with each sample containing 500–1000 embryonic hearts) and adult stage (N=5 biological replicates). Data were normalized to the expression of actb1. (J) The action potential of ventricular cardiomyocytes in adult id2b+/+ (n=9) and id2b-/- (n=9) hearts. (K) Statistical data showed a notable difference between id2b+/+ and id2b-/- accordingly. Data are presented as mean ± s.e.m. p-values were calculated by unpaired two-tailed Student’s t-tests. ***p<0.001, ****p<0.0001, ns, not significant. Scale bars, 50 μm.
The key functional unit that transmits electrical activity to contractile function is E-C coupling. Because id2b-/- displayed reduced cardiac function, we visualized calcium signaling in the developing heart using actb2:GCaMP6s zebrafish (Figure 5G). Compared to id2b+/+ controls, id2b-/- embryos exhibited markedly decreased calcium transient amplitude (Figure 5H), consistent with compromised calcium handling observed in other zebrafish cardiomyopathy models (Kamel et al., 2021a; Kamel et al., 2021b). In cardiomyocyte, the entry of extracellular calcium is mainly mediated through the LTCC. As previously reported, a defect in zebrafish LTCC pore-forming α1 subunit cacna1c leads to compromised cardiac function (Rottbauer et al., 2001). We collected hearts from 72 hpf and 5 months post-fertilization zebrafish and detected downregulated cacna1c in id2b-/- compared to id2b+/+ (Figure 5I). In addition, we measured cardiac action potential using intracellular recording (Zhang et al., 2013). Compared to id2b+/+ zebrafish, the duration of the action potential in id2b-/- was significantly shorter (Figure 5J and K), consistent with the decreased expression level of cacna1c. Together, these data indicate that id2b loss-of-function leads to compromised calcium signaling and cardiac contractile function.
Reduced expression of nrg1 mediates the compromised contractility in id2b-/-
Because the deficiency of id2b in the endocardium disrupted the function of myocardium, we speculated that the cross talk between these two types of cells was affected in id2b-/-. Interestingly, comparing the DEGs in embryonic id2b-/- and id2b+/+ hearts identified a significant reduction in the expression level of Nrg1, a key mitogen regulating the intra-organ communications between endocardial cells and cardiomyocytes (Figure 6A). Remarkably, analysis of a zebrafish single-cell database (Jiang et al., 2021) revealed enriched expression of nrg1 in endocardial cells (Figure 6—figure supplement 1). However, attempts to detect nrg1 expression through in situ hybridization were unsuccessful, likely due to its low abundance in the heart. Alternatively, qRT-PCR analysis of purified 120 hpf embryonic hearts validated decreased nrg1 levels in id2b-/- compared to control (Figure 6B). Previous studies have demonstrated that perturbations in Nrg-Erbb2 signaling, as seen in zebrafish erbb2 mutants, result in dysfunctional cardiac contractility (Liu et al., 2010). Consistently, a decrease in heart rate was observed in embryos treated with the erbb2 inhibitor AG1478 (Figure 6C).
Figure 6 with 1 supplement see all
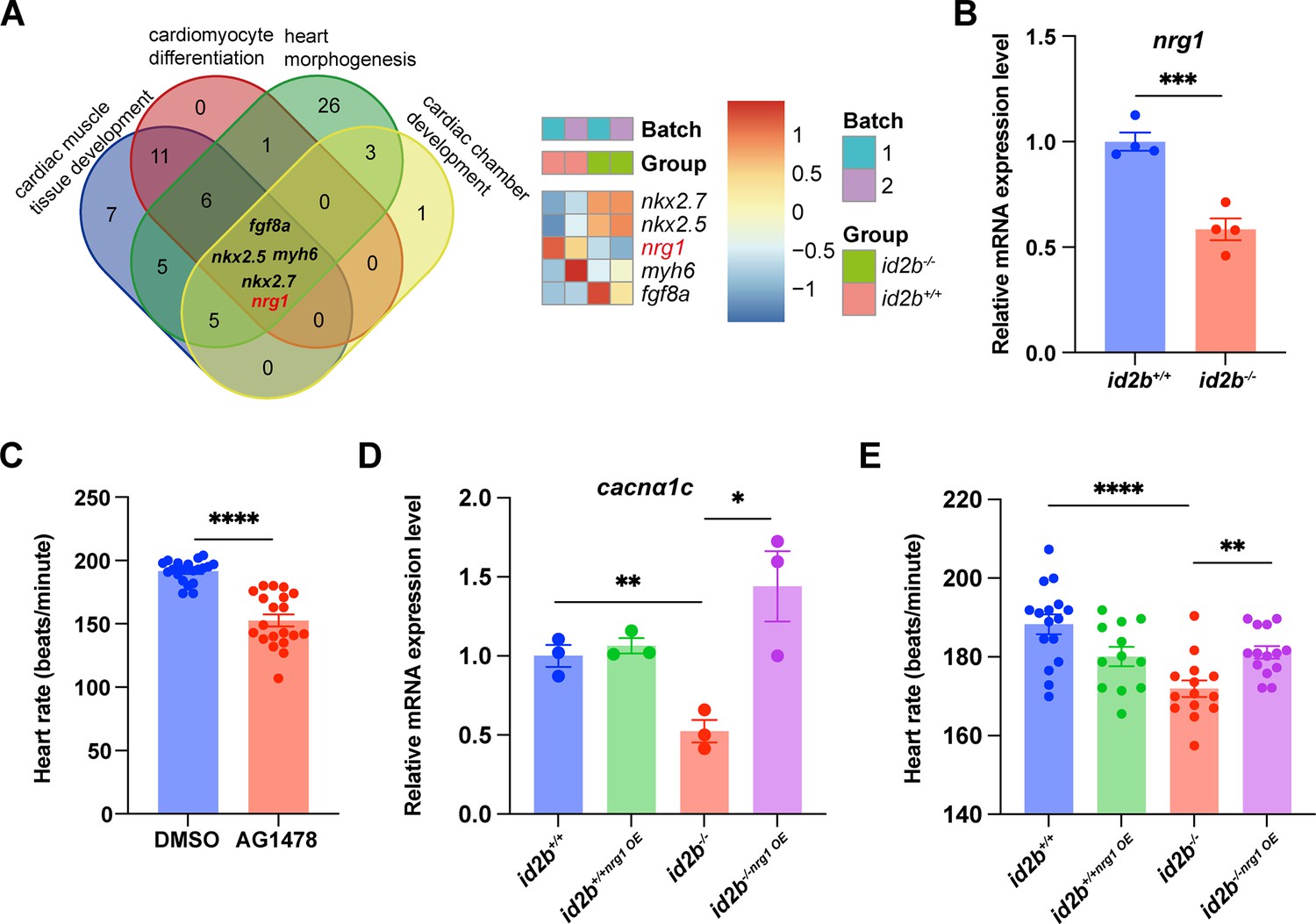
Nrg1 serves as a pivotal mitogen mediating the function of id2b.
(A) Identification of genes (fgf8a, nkx2.5, myh6, nrg1, and nkx2.7) associated with four distinct heart development processes: cardiac muscle tissue development, cardiomyocyte differentiation, heart morphogenesis, and cardiac chamber development. The heatmap illustrates scaled-normalized expression values for the mentioned genes. (B) Quantitative real-time PCR (qRT-PCR) analysis of nrg1 mRNA in 120 hr post-fertilization (hpf) id2b+/+ and id2b-/- embryonic hearts. Data were normalized to the expression of actb1. N=4 biological replicates, with each sample containing 500–1000 embryonic hearts. (C) Heart rate in 120 hpf larvae treated with AG1478 (n=20) and DMSO (n=20). (D) id2b+/+ and id2b-/- larvae were injected with nrg1 mRNA at the one-cell stage, followed by qRT-PCR analysis of cacnα1c mRNA at 72 hpf. Data were normalized to the expression of actb1. N=3 biological replicates, with each sample containing 100–200 embryonic hearts. (E) The heart rate of 72 hpf id2b+/+ and id2b-/- larvae injected with nrg1 mRNA at one-cell stage. n=(16, 12, 14, 14). Data are presented as mean ± s.e.m. p-values were calculated by unpaired two-tailed Student’s t-tests. *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001.
Remarkably, injecting nrg1 mRNA at the one-cell stage not only rescued the reduced expression of cacna1c in id2b-/- hearts (Figure 6D) but also restored the diminished heart rate (Figure 6E). This is consistent with prior studies showing that Nrg1 administration can restore LTCC expression and calcium current in failing mammalian cardiomyocytes (Wang et al., 2019). Overall, our data suggest that endocardial id2b promotes Nrg1 synthesis, thereby enhancing cardiomyocyte contractile function.
Id2b interacts with Tcf3b to limit its repressor activity on nrg1 expression
We further interrogated how id2b promotes the expression of nrg1. As an HLH factor lacking a DNA-binding motif, Id2b has been reported to form a heterodimer with Tcf3b to limit its function as a potent transcriptional repressor (Slattery et al., 2008). Notably, we detected expression of tcf3b in endocardial cells by analyzing a zebrafish single-cell database (Jiang et al., 2021; Figure 7—figure supplement 1). To determine if zebrafish Id2b and Tcf3b interact in vitro, Flag-id2b and HA-tcf3b were co-expressed in HEK293 cells. Co-immunoprecipitation analysis confirmed their interaction (Figure 7A), although whether they interact in vivo remains to be further investigated. Subsequently, qRT-PCR analysis on purified 120 hpf embryonic hearts revealed a significant increase in the expression of socs3b and socs1a, target genes of tcf3b, in id2b-/- compared to id2b+/+ (Figure 7B). This suggests an elevation in tcf3b activity associated with the loss of id2b function. Notably, the expression levels of tcf3a and tcf3b remained consistent between id2b-/- and id2b+/+ hearts (Figure 7B).
Figure 7 with 1 supplement see all
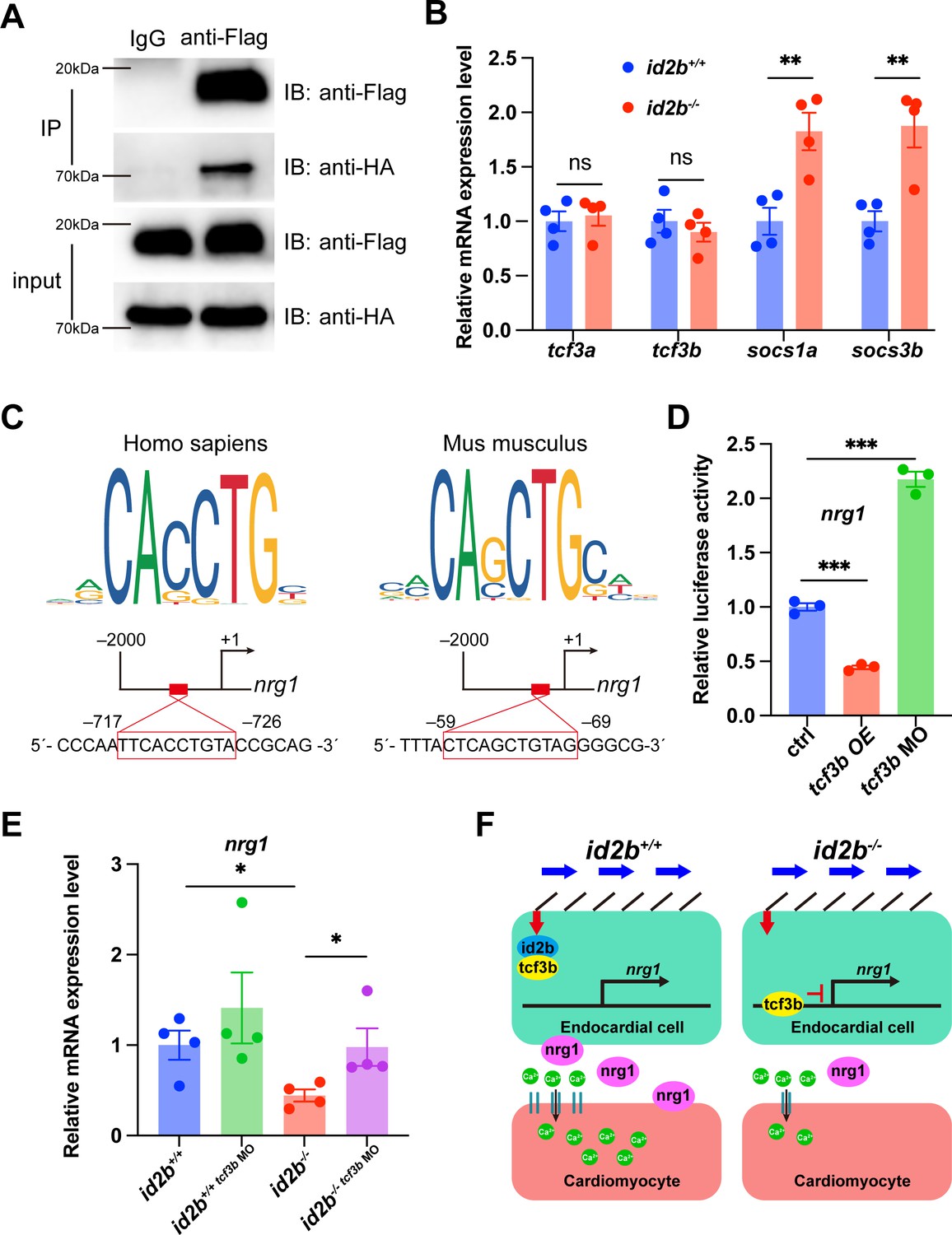
Id2b interacts with Tcf3b to restrict its inhibition on nrg1 expression.
(A) Immunoprecipitation (IP) assays of Flag-id2b and HA-tcf3b co-transfected 293T cells. (B) Quantitative real-time PCR (qRT-PCR) analysis of tcf3a, tcf3b, socs1a, and socs3b mRNA in 120 hr post-fertilization (hpf) id2b+/+ and id2b-/- embryonic hearts. Data were normalized to the expression of actb1. N=4 biological replicates, with each sample containing 500–1000 embryonic hearts. (C) Two potential Tcf3b-binding sites, with sequences corresponding to the human TCF3 (left) and mouse Tcf3 (right) binding motifs, were predicted in the 2000 bp DNA sequence upstream of the zebrafish nrg1 transcription start site using JASPAR. (D) Luciferase assay showing the expression of nrg1 in embryos with tcf3b overexpression (tcf3b OE) and morpholino-mediated tcf3b knockdown (tcf3b MO). N=3 biological replicates. (E) qRT-PCR analysis of nrg1 mRNA in 72 hpf id2b+/+ and id2b-/- embryonic hearts injected with control and tcf3b morpholino. Data were normalized to the expression of actb1. N=4 biological replicates, with each sample containing 100–200 embryonic hearts. (F) Schematic model for id2b-mediated regulation of myocardium function. During heart development, blood flow operates through primary cilia, initiating endocardial id2b expression. Subsequently, the interaction between Id2b and Tcf3b restricts the activity of Tcf3b, ensuring proper nrg1 expression, which in turn promotes L-type calcium channel (LTCC) expression (left). However, in the absence of Id2b, Tcf3b inhibits nrg1 expression. The reduced Nrg1 hinders LTCC expression in cardiomyocytes, resulting in decreased extracellular calcium entry and disruption of myocardial function. Data are presented as mean ± s.e.m. p-values were calculated by unpaired two-tailed Student’s t-tests. *p<0.05, **p<0.01, ***p<0.001. ns, not significant.
-
Figure 7—source data 1
Source data for western blot shown in Figure 7A.
- https://cdn.elifesciences.org/articles/101151/elife-101151-fig7-data1-v1.zip
-
Figure 7—source data 2
Original files for western blot shown in Figure 7A.
- https://cdn.elifesciences.org/articles/101151/elife-101151-fig7-data2-v1.zip
To understand how the altered interaction between id2b and tcf3b influences nrg1 expression, we analyzed the promoter region of zebrafish nrg1 using JASPAR and identified two potential tcf3b binding sites (Figure 7C). Subsequently, a DNA fragment containing the zebrafish nrg1 promoter region was subcloned into a vector carrying the luciferase reporter gene. Co-injection of this construct with tcf3b mRNA into one-cell stage embryos resulted in a significant decrease in luciferase signal. Conversely, co-injection with a previously characterized tcf3b morpholino led to enhanced luciferase intensity (Figure 7D). These results suggest a possible mechanism by which Tcf3b represses nrg1 expression in zebrafish.
Lastly, injecting tcf3b morpholino into id2b-/- embryos was performed to assess whether attenuating the overactive tcf3b in id2b-/- could restore the expression level of nrg1. qRT-PCR analysis of purified 72 hpf hearts revealed a partial restoration of the diminished nrg1 expression in id2b-/- upon tcf3b inhibition (Figure 7E). Taken together, our results indicate that biomechanical cues activate endocardial Id2b expression, leading to its interaction with Tcf3b to alleviate repression on the nrg1 promoter. Consequently, the depletion of id2b unleashes Tcf3b’s repressor activity, leading to a reduction in nrg1 expression, which further acts through erbb2 to regulate cardiomyocyte function (Figure 7F).
Discussion
Biomechanical forces play an essential role in regulating the patterning and function of the heart. At AVC, oscillatory flow promotes the expression of klf2a and nfatc1 to modulate valve morphogenesis. In chamber endocardium, blood flow induces endocardial cells to acquire distinct cell morphology. However, it still lacks a systematic analysis of the transcriptome underlying compromised heartbeats. In the present study, we analyzed embryonic zebrafish hearts without contractility and identified genes that are regulated by biomechanical forces. Specifically, our results unveiled the endocardial-specific expression of id2b, which was tightly regulated by flow-sensitive primary cilia-klf2 axis. Genetic deletion of id2b resulted in compromised valve formation and progressive atrium enlargement. In addition, a reduction in heart rate and contractile force was observed in id2b-/-, owing to decreased expression of LTCC α1 subunit cacna1c. Mechanistically, id2b interacts with bHLH TF tcf3b to limit its repressor activity. Hence, genetic deletion of id2b unleashes tcf3b activity, which further represses endocardial nrg1 expression. As a result, injection of nrg1 mRNA partially rescues the phenotype of id2b deletion. Overall, our findings identify id2b as a novel mediator that regulates the interplay between endocardium and myocardium during heart development.
In mammals, the deletion of Id2 leads to malformations in the arterial and venous poles of the heart, as well as affects AV valve morphogenesis (Jongbloed et al., 2011; Moskowitz et al., 2011). Interestingly, approximately 20% of perinatal lethality is reported in Id2 knockout mice, exhibiting AV septal defects and membranous ventricular septal defects (Moskowitz et al., 2011). Remarkably, pericardial edema is evident in 20% of adult id2b-/- zebrafish, with a prominent enlargement of the atrium. The superior and inferior leaflets of AV valves in id2b-/- mutants are significantly thinner compared to the control. Therefore, our results suggest that id2b may play a similar role in regulating AV valve formation in zebrafish as its mammalian orthologue Id2. It is proposed that the loss of Id2 in mice results in compromised endocardial proliferation and aberrant endothelial-to-mesenchymal transformation, collectively leading to defective valve morphogenesis (Moskowitz et al., 2011). Nevertheless, the mechanism by which id2b loss-of-function causes a reduction in leaflet thickness in zebrafish remains to be determined in future studies.
id2b has been recognized as a target gene of the BMP signaling pathway. As expected, knockdown of bmp2b, bmp4, and bmp7a at one-cell stage confirms that endocardial id2b expression is controlled by BMP activity during early embryonic development. Surprisingly, treatment with the BMP inhibitor Dorsomorphin at 24 and 36 hpf, when cardiac contractions have already initiated, fails to alter id2b expression in the endocardium, suggesting that BMP is dispensable for id2b activation at these stages. Instead, endocardial id2b expression is reduced upon loss-of-function of klf2a, klf2b, and ift88, suggesting an essential role of the primary cilia-klf2 axis in mediating id2b activation. In endocardial cells, Trp, Piezo, and ATP-dependent P2X/P2Y channels (Heckel et al., 2015; Fukui et al., 2021; Li et al., 2014; Nonomura et al., 2018) are well-established sensors for biomechanical stimulation. The activation of these channels further promotes the activities of Klf2 and Nfatc1 to drive heart development and valvulogenesis. However, whether these channels are also required for the activation of id2b warrants further investigation.
The Nrg-Erbb signaling plays an essential role in regulating heart morphogenesis and function. In the mammalian heart, the genetic deletion of Nrg1 or Erbb2 results in severely perturbed cardiac trabeculae formation (Gassmann et al., 1995; Lee et al., 1995; Meyer and Birchmeier, 1995). Zebrafish erbb2 mutants exhibit a similar defect in cardiomyocyte proliferation and trabeculation (Liu et al., 2010). Interestingly, nrg1 mutant zebrafish display grossly normal cardiac structure during early embryonic development (Rasouli and Stainier, 2017; Brown et al., 2018). Nevertheless, zebrafish nrg2a loss-of-function leads to defective trabeculae formation, suggesting that nrg2a is the predominant ligand secreted from endocardium, promoting ventricular morphogenesis (Rasouli and Stainier, 2017). In the adult stage, perivascular cells (Gemberling et al., 2015) or regulatory T cell-derived (Hui et al., 2017) nrg1 promotes cardiomyocyte proliferation during heart regeneration. Hence, the specific ligand/receptor and the spatiotemporal regulation of the Nrg-Erbb axis appear to be more complicated in both embryonic and adult zebrafish. Interestingly, the nrg1 mutant heart exhibits a defect in cardiac nerve expansion and heart maturation at the juvenile stage despite normal cardiac structure formation (Brown et al., 2018), suggesting its potential role in regulating cardiac function. Our findings demonstrate that the expression of nrg1 in embryonic endocardial cells is influenced by biomechanical cues and id2b activity. This signaling axis is essential for coordinating endocardium-myocardium interaction and establishing proper cardiac function.
Materials and methods
Zebrafish handling and lines
Request a detailed protocolAll animal procedures were approved by the Animal Care and Use Committee of the Zhejiang University School of Medicine (application no. 29296). Embryonic and adult fish were raised and maintained under standard conditions at 28°C on a 14/10 hr day/night cycle. The following zebrafish lines were used in this study: Tg(myl7:mCherry)sd7 (Palencia-Desai et al., 2011), Tg(myl7:H2A-mCherry)sd12 (Schumacher et al., 2013), Tg(kdrl:mCherry)S896 (Chi et al., 2008), Tg(kdrl:nucGFP)y7 (Roman et al., 2002), Tg(BRE:d2GFP)mw30 (Collery and Link, 2011), and Tg(actb2:Gcamp6s). To generate the id2b mutant, two short guide RNAs (sgRNAs) targeting exon 1 were generated using the MAXIscript T7 transcription kit (ambion, AM1314). The sgRNAs were as follows: sgRNA1: 5' - GAAGGCAGTCAGTCCGGTG - 3'; sgRNA2: 5' - GAACCGGAGCGTGAGTAAGA - 3'. The two sgRNAs, along with zCas9 protein, were co-injected into one-cell stage embryos. Embryos were raised to adulthood and crossed to wild-type zebrafish to obtain F1 progenies. Through PCR analysis, a mutant line with a 157 bp truncation was identified.
The knock-in id2b:eGFP line was generated using a previously reported method (Li et al., 2015). Briefly, three sgRNAs were designed to target the intron of id2b (sgRNA1: 5' - GAGACAAATATCTACTAGTG - 3'; sgRNA2: 5' - GTTGAACACATGACGATATT - 3'; sgRNA3: 5' - GCACAACTTAGATTTCAAGT - 3'). Co-injection of each individual sgRNA with zCas9 protein into one-cell stage zebrafish embryos yielded varying cleavage efficiency. Since sgRNA2 displayed the highest gene editing efficiency, it was selected for subsequent studies. Next, a donor plasmid containing the sgRNA targeting sequence of the intron, exon 2 of id2b, and P2A-eGFP was generated. Co-injection of sgRNA, donor plasmid, and zCas9 protein into one-cell stage embryos led to concurrent cleavage of the sgRNA targeting sites in both the zebrafish genome and the donor plasmid (Figure 2A). Accordingly, eGFP fluorescence was observed in injected 24 hpf zebrafish embryos, indicating the incorporation of the donor. The insertion of the id2b-p2A-eGFP donor into the genome was confirmed by PCR analysis with primers recognizing target site or donor sequences, respectively (Figure 2A). Embryos with mosaic eGFP expression were raised to adulthood and crossed with wild-type zebrafish to obtain F1 progenies. Overall, two founders were identified. The junction region of the F1 embryos was sequenced to determine the integration sites. Although the two founders had slightly different integration sites in the intron, the expression pattern and fluorescence intensity of eGFP were indistinguishable between the two lines.
Morpholinos
Request a detailed protocolAll morpholinos (Gene Tools) used in this study have been previously characterized. tnnt2a MO (5' - CATGTTTGCTCTGATCTGACACGCA - 3') (Sehnert et al., 2002); ift88 MO (5' - CTGGGACAAGATGCACATTCTCCAT - 3') (Li et al., 2020); bmp2b MO (5' - ACCACGGCGACCATGATCAGTCAGT - 3') (Lele et al., 2001); bmp4 MO (5' - AACAGTCCATGTTTGTCGAGAGGTG - 3') (Weber et al., 2008); bmp7a MO (5' - GCACTGGAAACATTTTTAGAGTCAT - 3') (Lele et al., 2001); tcf3b MO (5' - CGCCTCCGTTAAGCTGCGGCATGTT - 3') (Dorsky et al., 2003). For each morpholino, a 1 nL solution was injected into one-cell stage embryos at the specified concentrations: 0.5 µg/µL tnnt2a MO, 2 µg/µL ift88 MO, 0.5 µg/µL bmp2b MO, 2 µg/µL bmp4 MO, 4 µg/µL bmp7a MO, and 1 µg/µL tcf3b MO.
Small molecules treatment
Request a detailed protocolTo inhibit cardiac contraction, embryos were incubated in 1 mg/mL tricaine (Sigma, A5040) or 10 µM blebbistatin (MedChemExpress, HY13441) PTU-added egg water for 12–24 hr. In order to inhibit erbb2 signaling pathway, 10 µM AG1478 (Sigma, 658552) was used to treat 4 dpf larvae. To inhibit BMP signaling pathway, 10 µM Dorsomorphin (Sigma, P5499) was used to treat 10, 24, and 36 hpf embryos.
In situ hybridization and RNAscope
Request a detailed protocolWhole-mount in situ hybridization was performed as previously described (Zhang et al., 2013). The probes were synthesized using the DIG RNA labeling kit (Roche). The primers used for obtaining the id2b probe template were as follows: Forward 5' - ATGAAGGCAGTCAGTCCGGTGAGGT - 3'; Reverse 5' - TCAACGAGACAGGGCTATGAGGTCA - 3'. RNAscope analysis was performed using the probe Dr-id2b (Advanced Cell Diagnostics, 517541) and the Multiplex Fluorescent Detection Kit version 2 (Advanced Cell Diagnostics, 323100) as previously described (Liang et al., 2025).
Embryonic heart isolation and RNA-seq analysis
Request a detailed protocolHearts were isolated from embryos carrying the Tg(myl7:mCherry) transgene following an established protocol (Burns and MacRae, 2006). A minimum of 1000 hearts for each experimental group was manually collected under a Leica M165FC fluorescence stereomicroscope and transferred into ice-cold PBS buffer. After centrifugation at 12,000×g for 2 min at 4°C, the supernatant was removed, and hearts were lysed in cold TRIzol buffer (Ambion, 15596). Total RNA was extracted for subsequent qRT-PCR or RNA-seq analysis.
Duplicate samples from control and Tricaine-treated embryonic hearts underwent RNA-seq. Raw sequencing reads were preprocessed to remove adapters and filter low-quality reads. Clean sequencing reads were then mapped to the zebrafish reference (Weinberger et al., 2020) using STAR with default parameters (Dobin et al., 2013). Subsequently, gene quantification was carried out with RSEM (Li and Dewey, 2011). The gene expected count was applied to identify DEGs, retaining only genes with counts per million of 10 in at least two samples. DESeq2 (Love et al., 2014) was employed for differential expression analysis, and p-values were adjusted using BH correction. DEGs were defined as those with |log2fold change|≥0.585 and an adjusted p-value<0.1. The primary focus was on genes related to transcription regulation, and gene enrichment analysis was conducted using ClusterProfiler (Wu et al., 2021). To analyze DEGs in id2b-/- and control embryonic hearts, we performed enrichment analysis with the R package EnrichR, dissecting the potential anatomy expression pattern and underlying phenotypes. We mainly focused on genes with the heart-related phenotypes, including cardiac muscle tissue development, cardiomyocyte differentiation, heart morphogenesis, and cardiac chamber development. All the analysis on identifying DEGs was batch-corrected.
qRT-PCR analysis
Request a detailed protocolAfter extraction from isolated embryonic hearts, 50 ng to 1 µg of mRNA was reverse-transcribed to cDNA using the PrimeScript RT Master Mix kit (Takara, RR036A). Real-time PCR was performed using the TB Green Premix Ex Taq kit (Takara, RR420A) on the Roche LightCycler 480. Expression levels of the target genes were normalized to actb1 as an internal control. All experiments were repeated three times. The following primer sets were used: id2b Forward 5' - ACCTTCAGATCGCACTGGAC - 3', Reverse 5' - CTCCACGACCGAAACACCATT - 3'; nrg1 Forward 5' - CTGCATCATGGCTGAGGTGA - 3', Reverse 5' - TTAACTTCGGTTCCGCTTGC - 3'; cacnα1c Forward 5' - GCCCTTATTGTAGTGGGTAGTG - 3', Reverse 5' - AGTGTTTTGGAGGCCCATTG - 3'; tcf3a Forward 5' - CCTCCGGTCATGAGCAACTT - 3', Reverse 5' - TTTCCCATGATGCCTTCGCT - 3'; tcf3b Forward 5' - CCTTTAATGCGCCGTGCTTC - 3', Reverse 5' - GCGTTCTTCCATTCCTGTACCA - 3'; socs1a Forward 5' - TCAGCCTGACAGGAAGCAAG - 3', Reverse 5' - GTTGCACAGGGATGCAGTCG - 3'; socs3b Forward 5' - GGGACAGTGAGTTCCTCCAA - 3', Reverse 5' - ATGGGAGCATCGTACTCCTG - 3'; actb1 Forward 5' - ACCACGGCCGAAAGAGAAAT - 3', Reverse 5' - GCAAGATTCCATACCCAGGA - 3'.
Co-IP and western blot
Request a detailed protocolZebrafish tcf3b and id2b were overexpressed in 293T cell for 48 hr. The transfected cells were then collected and lysed using IP lysis buffer (Sangon Biotech, C500035) containing protease and phosphatase inhibitors (Sangon Biotech, C510009, C500017). For the IP experiment, anti-Flag antibody (Cell Signaling Technology, 14793, 1:100) and IgG antibody (ABclonal, AC005, 1:100) were incubated with cell lysates overnight at 4°C. Pretreated magnetic beads were bound with the antigen-antibody complex for 4 hr at 4°C, followed by washing with IP lysis buffer three times. For western blot, samples were denatured at 95°C for 10 min, separated on a 5–12% gradient gel. Proteins were then transferred to a PVDF membrane (Sigma, ISEQ00010). The membrane was blocked for 1 hr with 5% nonfat milk or 5% BSA (Sangon Biotech) dissolved in TBST and then incubated with primary antibodies (anti-FLAG, Cell Signaling Technology, 14793, 1:1000; anti-HA, Sigma, H3663, 1:1000) overnight at 4°C, followed by three times 10 min TBST washes. HRP-conjugated secondary antibodies (Invitrogen, 31430, 31460) were incubated for 1 hr at room temperature, followed by three times 10 min TBST washes. The detection of immunoreactive bands was performed with a chemiluminescent substrate (Thermo Scientific, 34577) and imaged using the Azure Biosystems 400.
Immunofluorescence
Request a detailed protocolFor immunofluorescence on adult zebrafish hearts, we fixed the hearts overnight in 4% paraformaldehyde at 4°C, followed by equilibration through 15% and 30% sucrose in PBS solution. The hearts were embedded and frozen in O.C.T. compound (Epredia, 6502), and 10-μm-thick cryosections were prepared using a CryoStar NX50 cryostat. Immunofluorescence experiments were performed as previously described (Han et al., 2014). For immunofluorescence on embryonic hearts, embryos were fixed overnight in 4% paraformaldehyde at 4°C, washed twice quickly in 100% methanol, and then dehydrated overnight at –20°C in 100% methanol. Subsequently, rehydration was performed through a methanol gradient (100%, 75%, 50%, 25%, 10 min each), followed by three times washes in PBST (1% PBS/0.1% Triton X-100, 10 min each). The embryos were treated with 10 μg/mL proteinase K diluted in PBST for 20 min at room temperature, refixed in 4% paraformaldehyde for 20 min, washed in PBST, and immersed in blocking solution (PBST/1% BSA/2% goat serum) for 1 hr at room temperature. Following this, the embryos were incubated in the primary antibody diluted in blocking solution overnight at 4°C. After washing in PBST, they were incubated in the secondary antibody (1:200) for 2 hr at room temperature. The primary antibody used was anti-GFP antibody (Santa Cruz Biotechnology, sc9996, 1:200) and anti-α-actinin antibody (Sigma, A7811, 1:200). The secondary antibody used was anti-mouse IgG-Alexa 488 (Invitrogen, A11011, 1:400). DAPI was used to stain cell nuclei.
Cardiac function analysis
Request a detailed protocolTo assess cardiac function in embryonic hearts, embryos were incubated in 0.16 mg/mL tricaine (Sigma, A5040) and embedded in 1% low melting agarose. Heart contractions were recorded for 1 min using a Nikon Ti2 microscope at a rate of 25 frames per second. Fractional shortening and heart rate were measured as described previously (Zhang et al., 2013). For cardiac contractile functions in adulthood, zebrafish were fixed on a sponge soaked with system water with the belly facing up, and echocardiography was performed (Wang et al., 2017). Videos and images in color Doppler mode and B-mode were obtained using the Vevo1100 imaging system at a frequency of 50 MHz. Nikon NIS-Elements AR analysis and ImageJ software were employed for data extraction. To evaluate AV valve function, the ratio of inflow and outflow area in the same frame was quantified (Gunawan et al., 2020).
Calcium imaging
Request a detailed protocolAt 120 hpf, embryos were treated with 10 mM 2,3-butanedione monoxime (Sigma, B0753) and mounted in 1% low melting agarose. Time-lapse images were acquired using a Nikon Ti2 microscope at a rate of 50 frames per second. Data were analyzed using Nikon-NIS Elements AR analysis software.
Intracellular action potential recording
Request a detailed protocolElectrophysiology study was performed on adult zebrafish ventricles as previously described (Zhang et al., 2013). Briefly, hearts were mounted in a chamber containing Tyrode’s solution: NaCl 150 mM, KCl 5.4 mM, MgSO4 1.5 mM, NaH2PO4 0.4 mM, CaCl2 2 mM, glucose 10 mM, HEPES 10 mM, pH was adjusted to 7.4. Glass pipettes with tip resistance 30–40 MΩ were filled with 3 M KCl solution. Intracellular action potentials were recorded using an HEKA amplifier and pClamp10.3 software (Molecular Devices).
Histology and HE staining
Request a detailed protocolAdult hearts were dissected and fixed overnight at 4°C in 4% PFA, followed by three times PBS washes. Dehydration involved an ethanol gradient (70%, 80%, 95%, 100%, 100%, 30 min each), followed by three soaks in dimethylbenzene at 65°C, before embedding in paraffin. Sections of 5 µm thickness were prepared using the Leica RM2235 manual rotary microtome for hematoxylin and eosin (HE) staining.
Injection of mRNA
Request a detailed protocolThe embryonic zebrafish cDNA library was used as a template to amplify the nrg1 and tcf3b fragment, which was then subcloned into the pCS2 vector. The vector was linearized using Not I restriction endonuclease, and mRNA was transcribed in vitro using the mRNA transcription kit (Ambion, AM1340). 100 pg of purified mRNA was injected into one-cell stage embryos.
Luciferase assay
Request a detailed protocolThe LCR (luciferase reporter) plasmid was generated by subcloning the 5′ UTR of nrg1 into the upstream region of renilla luciferase on the psiCheck2 plasmid. Following construction, 25 pg of the LCR plasmid was co-injected with either 100 pg of tcf3b mRNA or 1 ng of tcf3b MO into one-cell stage zebrafish embryos. At 48 hpf, 20 embryos were gathered into one group and fully lysed. Subsequently, firefly and renilla luciferase activities were sequentially measured using a microplate reader with the dual luciferase reporter gene assay kit (Yeasen, 11402ES60), according to the manufacturer’s instructions. The experiment was independently replicated three times. The relative renilla luciferase activity, normalized by firefly luciferase activity, served as an indicator of nrg1 expression level under the influence of tcf3b overexpression or reduction.
Image processing and statistical analysis
Request a detailed protocolWhole-mount in situ hybridization images were captured using a Leica M165FC stereomicroscope. Live imaging of zebrafish embryos involved mounting anesthetized embryos in 1% low melting agarose (Sangon Biotech, A600015) and manually orienting them for optimal visual access to the heart. Confocal images were obtained with a Nikon Ti2 confocal microscope. Fluorescence intensity and cell number counting were processed using Nikon NIS-Elements AR analysis and ImageJ software. Statistical analysis was performed using GraphPad Prism 8 software. No statistical methods were used to predetermine sample size. Unpaired two-tailed Student’s t-tests were used to determine statistical significance. Data are presented as mean ± s.e.m., *p<0.05 was considered to be statistically significant.
Data availability
The authors declare that all data supporting the findings in the paper are available in the article and the supplementary files. RNA-seq data have been deposited in GEO under accession number GSE295737 and GSE295738.
-
NCBI Gene Expression OmnibusID GSE295737. Effect of depletion of id2b on gene expression during zebrafish heart development.
-
NCBI Gene Expression OmnibusID GSE295738. Transcriptome analysis identifies id2b as a blood flow sensitive gene.
References
-
Neuregulin-1 is essential for nerve plexus formation during cardiac maturationJournal of Cellular and Molecular Medicine 22:2007–2017.https://doi.org/10.1111/jcmm.13408
-
Purification of hearts from zebrafish embryosBioTechniques 40:112135.https://doi.org/10.2144/000112135
-
Foxn4 directly regulates tbx2b expression and atrioventricular canal formationGenes & Development 22:734–739.https://doi.org/10.1101/gad.1629408
-
Dynamic smad-mediated BMP signaling revealed through transgenic zebrafishDevelopmental Dynamics 240:712–722.https://doi.org/10.1002/dvdy.22567
-
Blood flow and Bmp signaling control endocardial chamber morphogenesisDevelopmental Cell 30:367–377.https://doi.org/10.1016/j.devcel.2014.06.020
-
STAR: ultrafast universal RNA-seq alignerBioinformatics 29:15–21.https://doi.org/10.1093/bioinformatics/bts635
-
Two tcf3 genes cooperate to pattern the zebrafish brainDevelopment 130:1937–1947.https://doi.org/10.1242/dev.00402
-
Mechanotransduction in cardiovascular morphogenesis and tissue engineeringCurrent Opinion in Genetics & Development 57:106–116.https://doi.org/10.1016/j.gde.2019.08.002
-
Cardiomyocyte maturation: new phase in developmentCirculation Research 126:1086–1106.https://doi.org/10.1161/CIRCRESAHA.119.315862
-
Characterization of the zebrafish cell landscape at single-cell resolutionFrontiers in Cell and Developmental Biology 9:743421.https://doi.org/10.3389/fcell.2021.743421
-
Expression of Id2 in the second heart field and cardiac defects in Id2 knock-out miceDevelopmental Dynamics 240:2561–2577.https://doi.org/10.1002/dvdy.22762
-
A heterozygous mutation in cardiac troponin T promotes Ca2+ dysregulation and adult cardiomyopathy in zebrafishJournal of Cardiovascular Development and Disease 8:46.https://doi.org/10.3390/jcdd8040046
-
Genetically encoded tension heterogeneity sculpts cardiac trabeculationScience Advances 11:eads2998.https://doi.org/10.1126/sciadv.ads2998
-
A dual role for ErbB2 signaling in cardiac trabeculationDevelopment 137:3867–3875.https://doi.org/10.1242/dev.053736
-
Dependence of cardiac trabeculation on neuregulin signaling and blood flow in zebrafishDevelopmental Dynamics 240:446–456.https://doi.org/10.1002/dvdy.22526
-
Fluid forces shape the embryonic heart: Insights from zebrafishCurrent Topics in Developmental Biology 132:395–416.https://doi.org/10.1016/bs.ctdb.2018.12.009
-
E2A proteins: regulators of cell phenotype in normal physiology and diseaseThe International Journal of Biochemistry & Cell Biology 40:1431–1436.https://doi.org/10.1016/j.biocel.2007.05.014
-
SIX2 and BMP4 mutations associate with anomalous kidney developmentJournal of the American Society of Nephrology 19:891–903.https://doi.org/10.1681/ASN.2006111282
-
Embryonic alcohol exposure in zebrafish predisposes adults to cardiomyopathy and diastolic dysfunctionCardiovascular Research 120:1607–1621.https://doi.org/10.1093/cvr/cvae139
-
Functional heterogeneity within the developing zebrafish epicardiumDevelopmental Cell 52:574–590.https://doi.org/10.1016/j.devcel.2020.01.023
Article and author information
Author details
Funding
National Natural Science Foundation of China (32170823)
- Peidong Han
National Key Research and Development Program of China (2023YFA1800600)
- Peidong Han
National Natural Science Foundation of China (92468104)
- Peidong Han
National Natural Science Foundation of China (31871462)
- Peidong Han
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
We thank Dr. Pengfei Xu for providing morpholinos. We also thank Dr. Jia Li for the support in generating the id2b:eGFP line. This work was supported by the National Natural Science Foundation of China (32170823, 92468104, 31871462), and the National Key R&D Program of China (2023YFA1800600).
Ethics
All animal procedures were approved by the Animal Care and Use Committee of the Zhejiang University School of Medicine (application no.29296).
Version history
- Sent for peer review:
- Preprint posted:
- Reviewed Preprint version 1:
- Reviewed Preprint version 2:
- Version of Record published:
Cite all versions
You can cite all versions using the DOI https://doi.org/10.7554/eLife.101151. This DOI represents all versions, and will always resolve to the latest one.
Copyright
© 2024, Chen, Liang et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 1,445
- views
-
- 84
- downloads
-
- 1
- citation
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 1
- citation for umbrella DOI https://doi.org/10.7554/eLife.101151
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Contraction-induced endocardial id2b plays a dual role in regulating myocardial contractility and valve formation
eLife 13:RP101151.
https://doi.org/10.7554/eLife.101151.3




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}