Structural mechanisms of PIP2 activation and SEA0400 inhibition in human cardiac sodium-calcium exchanger NCX1
- Department of Physiology, The University of Texas Southwestern Medical Center, United States
- Department of Biophysics, The University of Texas Southwestern Medical Center, United States
- Howard Hughes Medical Institute, United States
- Department of Medicine (Cardiology), UCLA, United States
- Department of Anesthesiology and Perioperative Medicine, Division of Molecular Medicine, David Geffen School of Medicine, University of California Los Angeles, United States
Figures
Figure 1
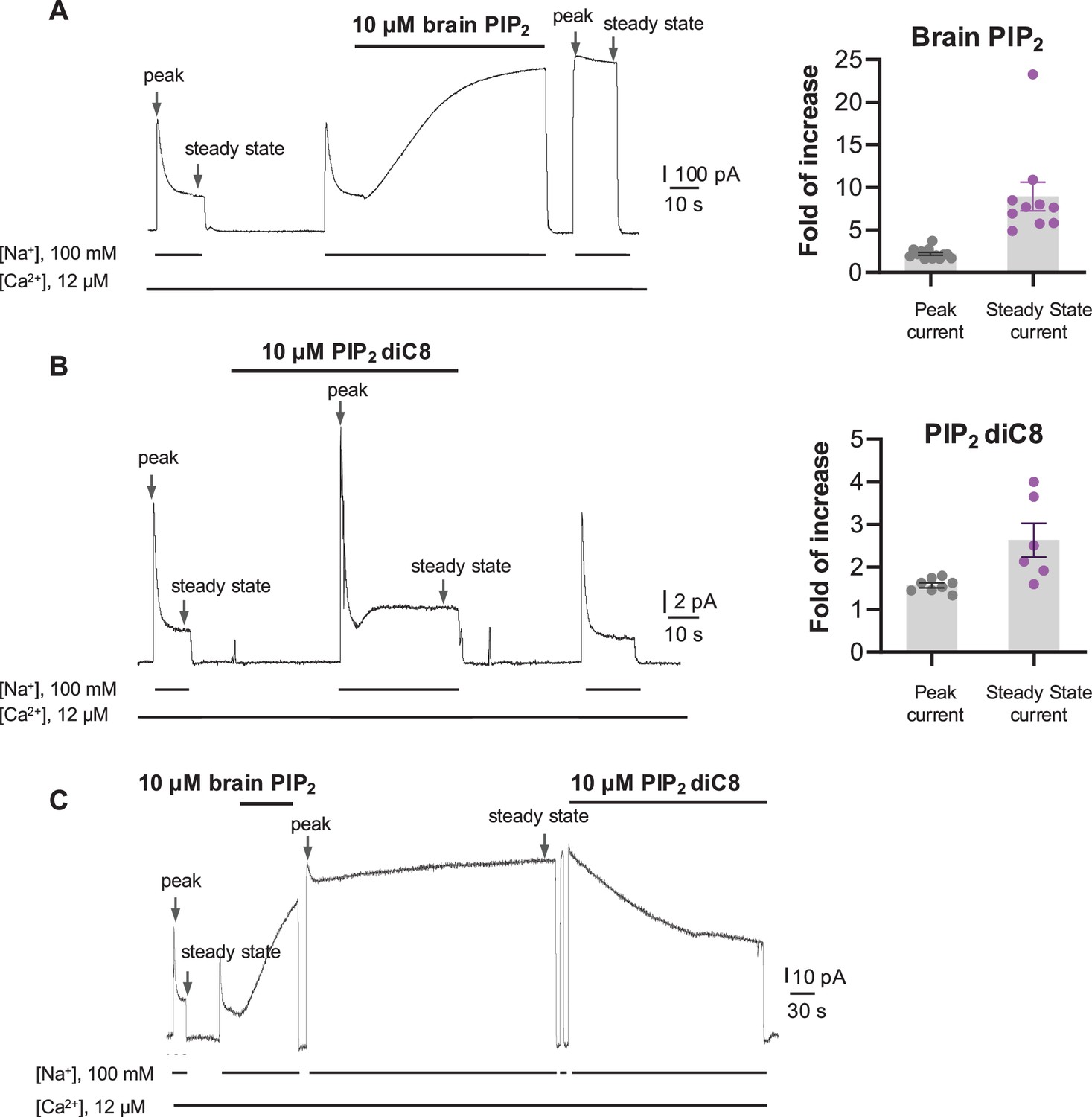
Phosphatidylinositol 4,5-bisphosphate (PIP2) enhances HsNCX1 activity.
(A) Representative outward currents recorded from oocytes expressing the human NCX1 before and after application of long-chain brain PIP2. Currents were activated by replacing cytosolic Cs+ with Na+. Application of 10 µM brain PIP2 enhanced HsNCX1 current and abolished the Na+-dependent inactivation irreversibly. Perfusion time of PIP2 is indicated above traces, while lines below traces indicate solution exchange. Arrows mark the peak and steady currents used to measure the fold of increase upon PIP2 application. The fold of current increase was calculated by comparing the peak or steady-state current before and after PIP2 application (fold of increase in peak current = 2.2 ± 0.2, n=14; steady-state current = 8.9 ± 1.7, n=10; data points are mean ± s.e.m.). (B) Representative outward currents recorded before and after application of short-chain PIP2 diC8 (10 µM). PIP2 diC8 was perfused from the cytosolic side before HsNCX1 activation (in the presence of Cs+ for 30 s) and during transport (in the presence of Na+). Both peak and steady-state currents of HsNCX1 are enhanced by PIP2 diC8, and the effect is reversible (fold of current increase in peak current = 1.6 ± 0.1, n=8; steady-state current = 2.6 ± 0.4, n=6; data points are mean ± s.e.m.). The Na+-dependent inactivation remains in the presence of PIP2 diC8. (C) Representative outward currents recorded with the application of brain PIP2 and PIP2 diC8. The NCX1 current was first potentiated by applying 10 µM brain PIP2 at the steady state. The PIP2 effect was not reversible over the 5 min washout with a solution containing 100 mM Na+ and 12 µM Ca2+. The same patch was then perfused with the same solution in the presence of 10 µM PIP2 diC8. Application of the short-chain PIP2 diC8 facilitates the decrease of brain PIP2-potentiated current, suggesting that both lipids compete for the same binding site.
-
Figure 1—source data 1
The fold of current increase in the peak or steady-state current before and after PIP2 application.
- https://cdn.elifesciences.org/articles/105396/elife-105396-fig1-data1-v1.xlsx
Figure 2 with 4 supplements
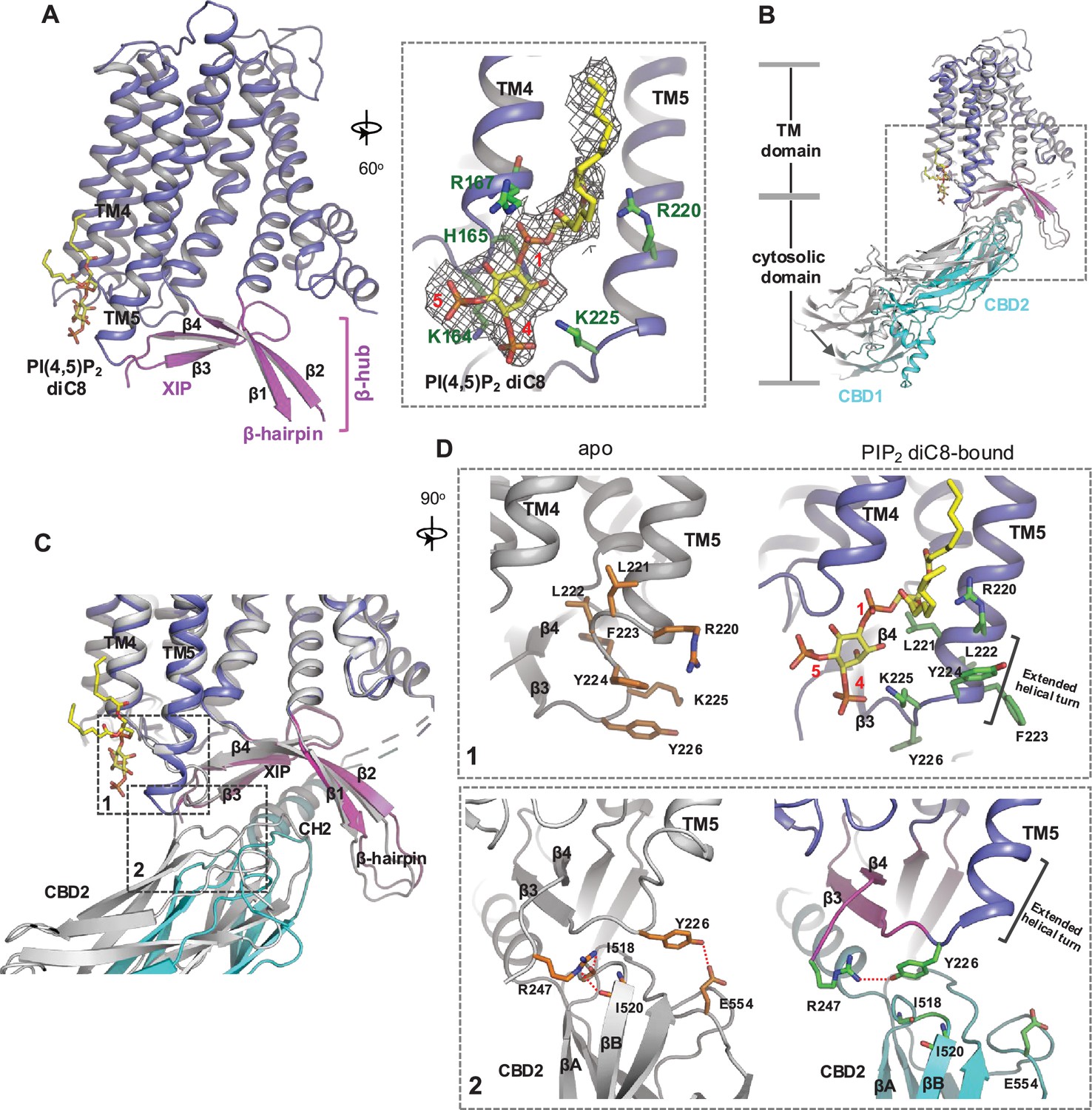
Phosphatidylinositol 4,5-bisphosphate (PIP2) binding in NCX1.
(A) Structure of the transmembrane (TM) and β-hub regions of PIP2 diC8-bound NCX1 with a zoomed-in view of the lipid-binding site. The density of PIP2 diC8 is shown as a gray mesh contoured at 5.5σ. (B) Structural comparison between apo (gray) and PIP2 diC8-bound (color) NCX1. Upon PIP2 diC8 binding, there is a rigid-body downward swing movement (marked by an arrow) at CBDs caused by the partial detachment of the CBD2 domain from XIP. The conformational change at the TM domain is subtle and mainly occurs at the C-terminus of TM5 as illustrated in (C) and (D). (C) Zoomed-in view of the structural comparison (boxed area in (B)). The two major conformational changes occur in the boxed regions. (D) Zoomed-in views of the two conformational changes between apo (left in gray) and PIP2 diC8-bound (right in color) state. Top: conformational change 1 at the C-terminus of TM5. Bottom: conformational change 2 at the interface between XIP and CBD2.
Figure 2—figure supplement 1
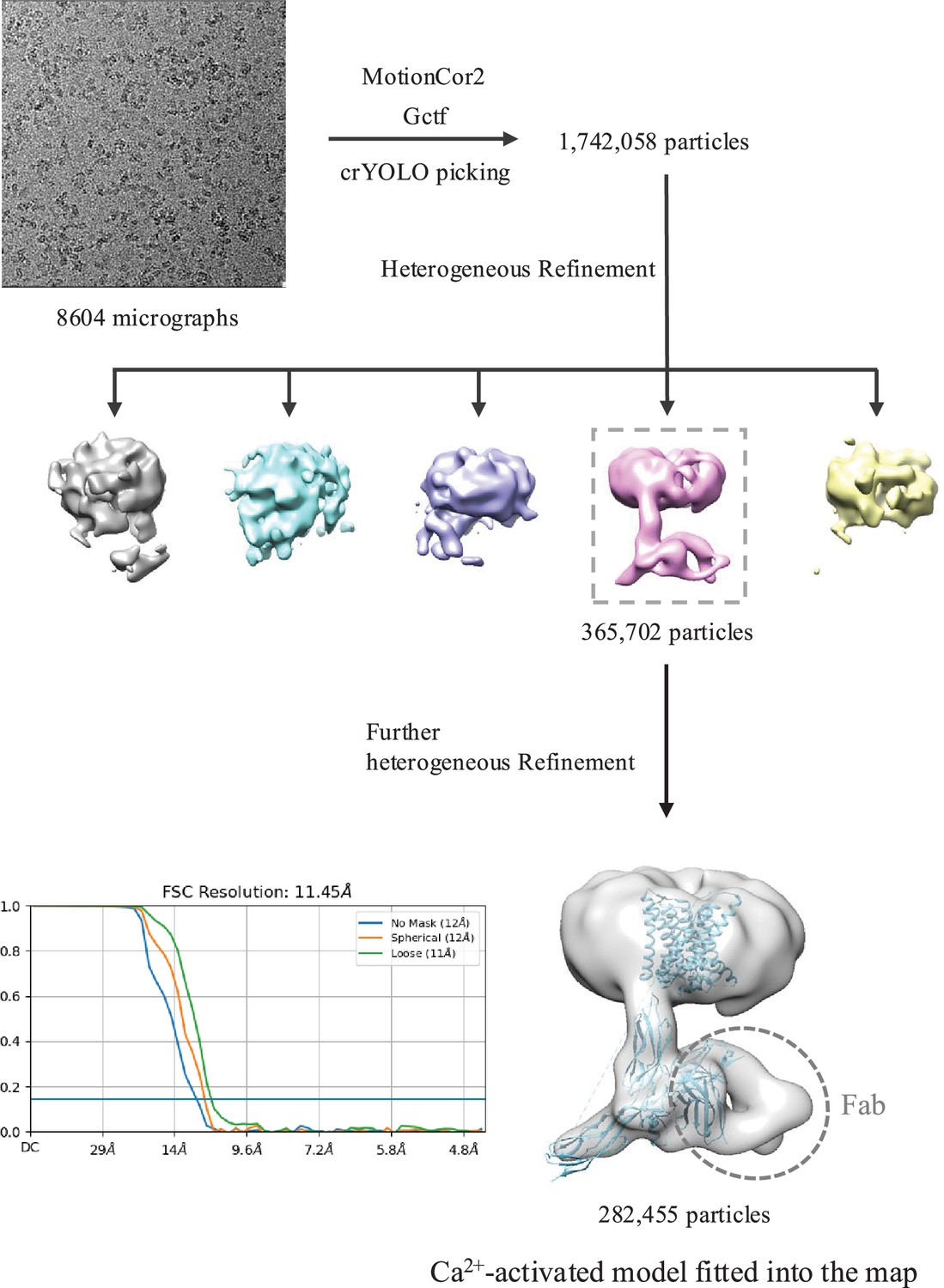
Cryo-EM data processing of HsNCX1 in the presence of long-chain porcine brain phosphatidylinositol 4,5-bisphosphate (PIP2).
The structural model of Ca2+-activated HsNCX1 from a previous study (PDB 8SGT) was directly fitted into the low-resolution EM map (~11.5 Å). The Fab fragment from a monoclonal antibody against NCX1 was used as a fiducial marker to facilitate the single-particle alignment.
Figure 2—figure supplement 2
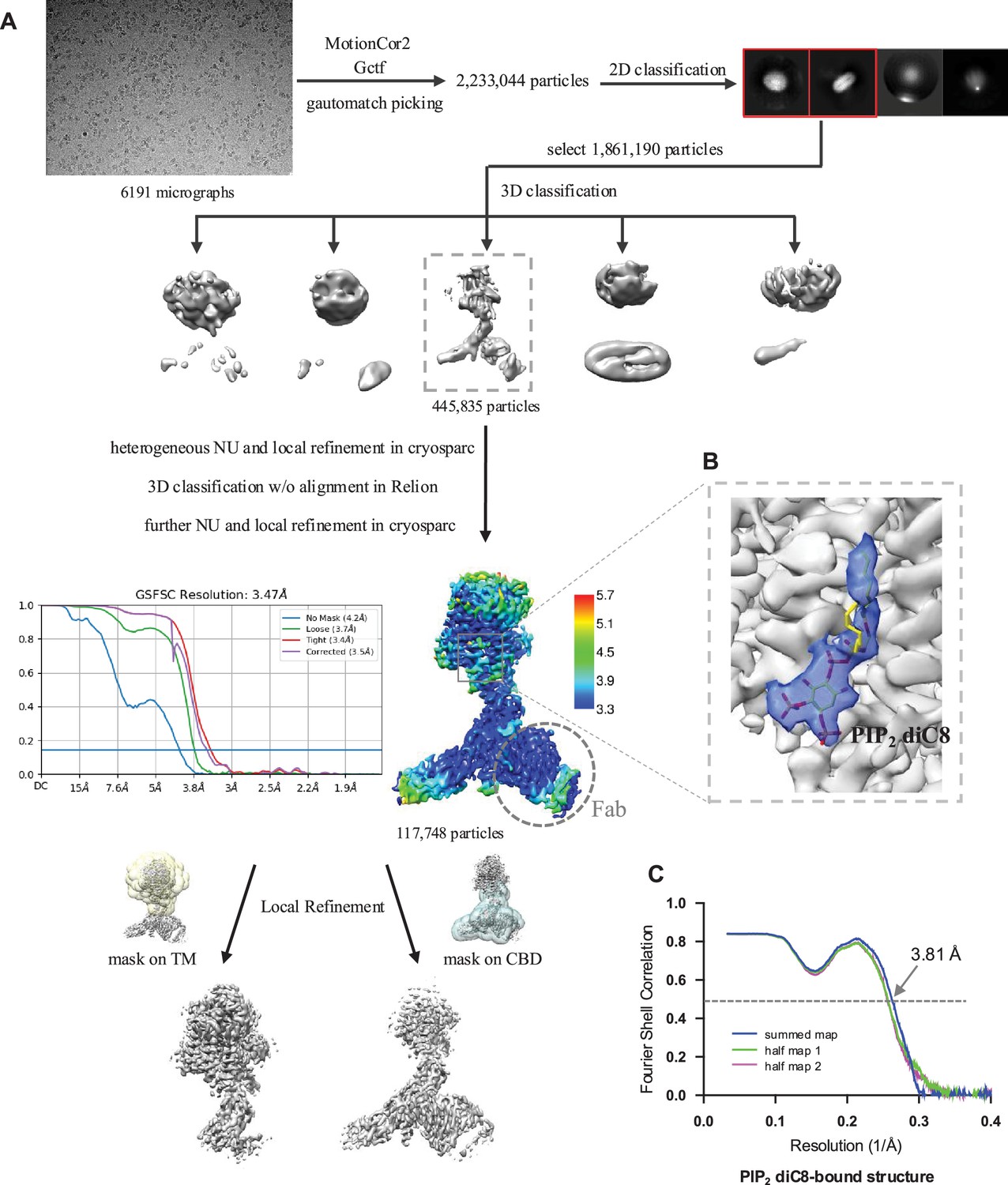
Structure determination of HsNCX1-PIP2 diC8 complex.
(A) Cryo-EM data processing of HsNCX1 in complex with short-chain PIP2 diC8. The Fab fragment from a monoclonal antibody against NCX1 was used as a fiducial marker to facilitate the single-particle alignment. (B) Zoomed-in view of the density map of the bound PIP2 diC8 contoured at the threshold level of 0.35 using ChimeraX. (C) The Fourier shell correlation (FSC) curves for cross-validation between the maps and the models.
Figure 2—figure supplement 3

Side-by-side comparison of the densities at the phosphatidylinositol 4,5-bisphosphate (PIP2)-binding site between the PIP2-bound structure (EMD-60921) and the apo structure (EMD-40457).
The local-refined maps focused on the transmembrane (TM) domain are used in the comparison. The density map of the bound PIP2 is contoured at a threshold level of 0.35 using ChimeraX.
Figure 2—figure supplement 4
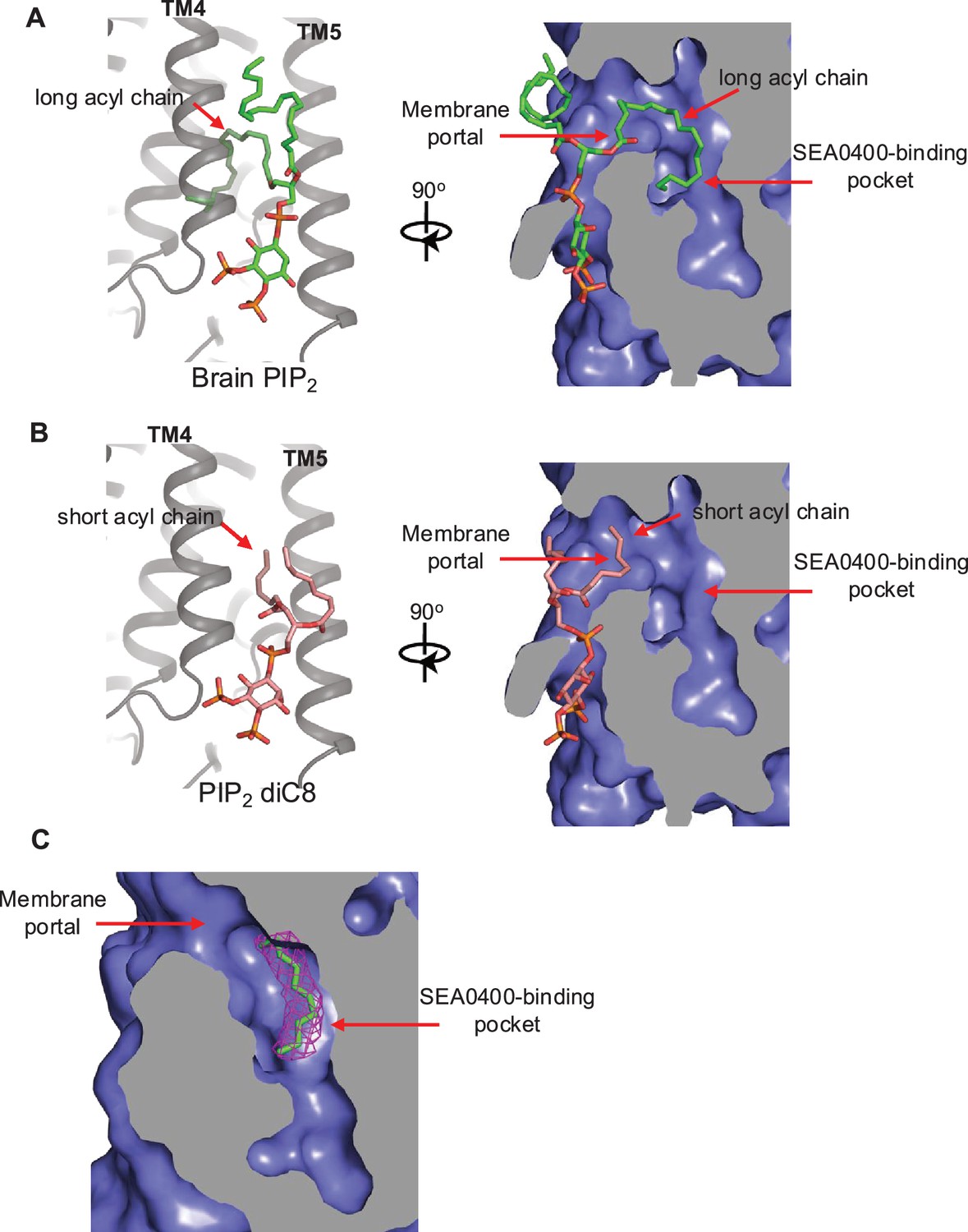
Proposed structural basis underlying the different binding affinity between long- and short-chain phosphatidylinositol 4,5-bisphosphate (PIP2).
(A) A docking model for native brain PIP2 binding in NCX1 showing the insertion of its long acyl chain into the SEA0400-binding pocket. (B) A docking model for short-chain PIP2 diC8 binding in NCX1 showing that the SEA0400-binding pocket is not accessible to shorter acyl chain. (C) The density (red mesh, contoured at 6σ) of an acyl chain likely from a native lipid is observed in the SEA0400-binding pocket of the apo NCX1 structure (EMD-40457, local-refined map at the transmembrane [TM] domain).
Figure 3
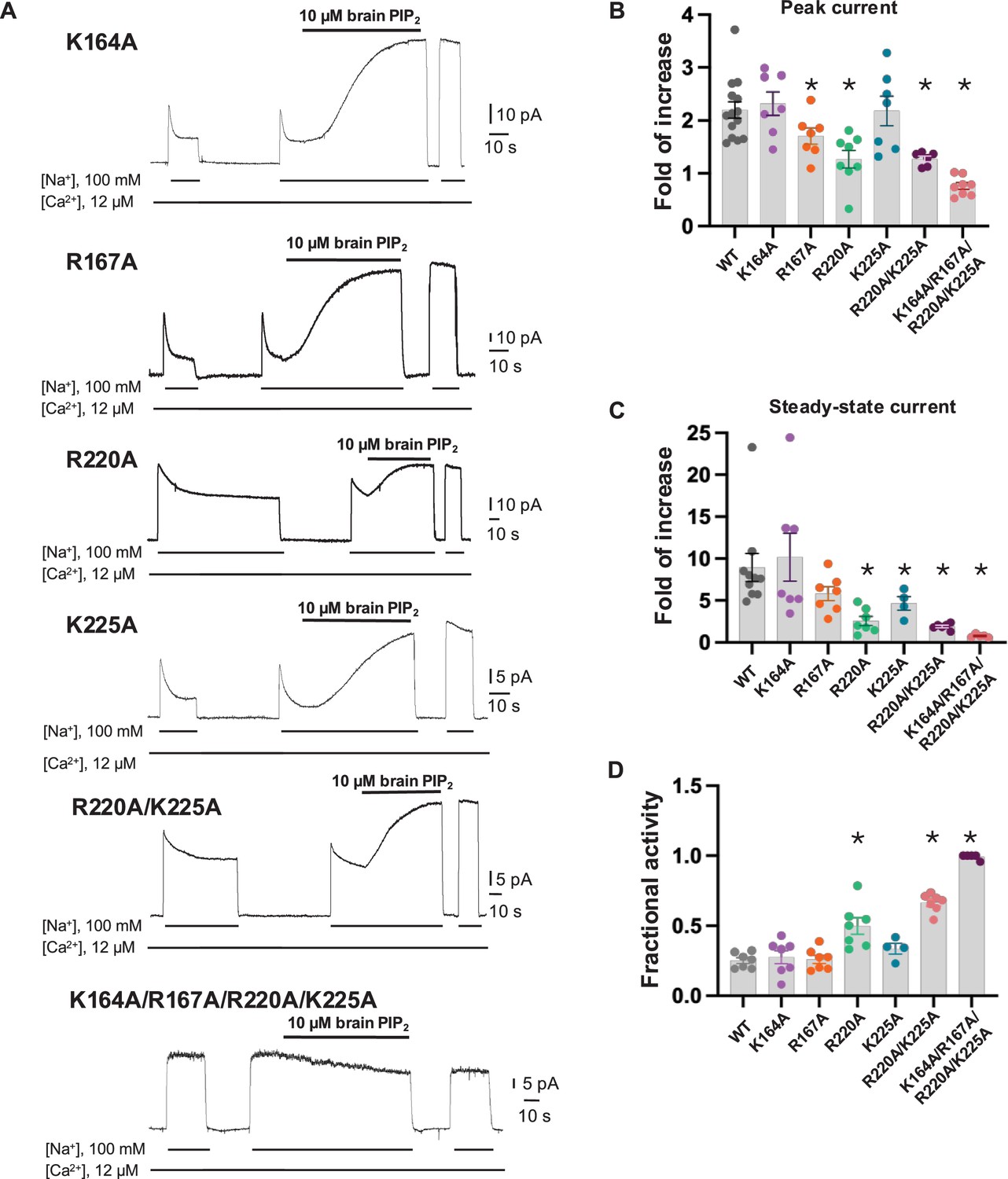
Mutagenesis at the phosphatidylinositol 4,5-bisphosphate (PIP2) binding.
(A) Representative NCX1 currents of PIP2 site mutants before and after perfusion of 10 µM brain PIP2 to the cytosolic side of the patch. (B, C) Summary graphs demonstrating the effects of PIP2 on the enhancement of peak (B) and steady-state currents (C). Potentiation (fold of increase) was measured by comparing the current magnitude before and after PIP2 application. Mutants R167A, R220A, and K225A showed some decreased response to PIP2, whereas the R220A/K225A mutant shows a more profound decrease in PIP2 response. Compared to WT, the PIP2 potentiation of R220A/K225A at the steady state is decreased by ~70–90% (fold of increase WT = 8.9 ± 1.6, n=10 vs R220A/K225A=1.9 ± 0.1, n=6). PIP2 has no potentiation effect on the quadruple K164A/R167A/R220A/K225A mutant. Data points are mean ± s.e.m. (*p<0.1). (D) The extent of Na+-dependent inactivation was measured as the ratio between steady-state and peak currents (fractional activity), and values for WT and the indicated mutants are shown. Mutants R220A and R220A/K225A displayed significantly higher fractional activity values when compared to WT, indicating that the Na+-dependent inactivation was less pronounced in these mutant exchangers. K164A/R167A/R220A/K225A shows no Na+-dependent inactivation.
-
Figure 3—source data 1
Fold of increase in peak and steady-state current, and fractional activity values for WT and indicated mutants.
- https://cdn.elifesciences.org/articles/105396/elife-105396-fig3-data1-v1.xlsx
Figure 4 with 2 supplements
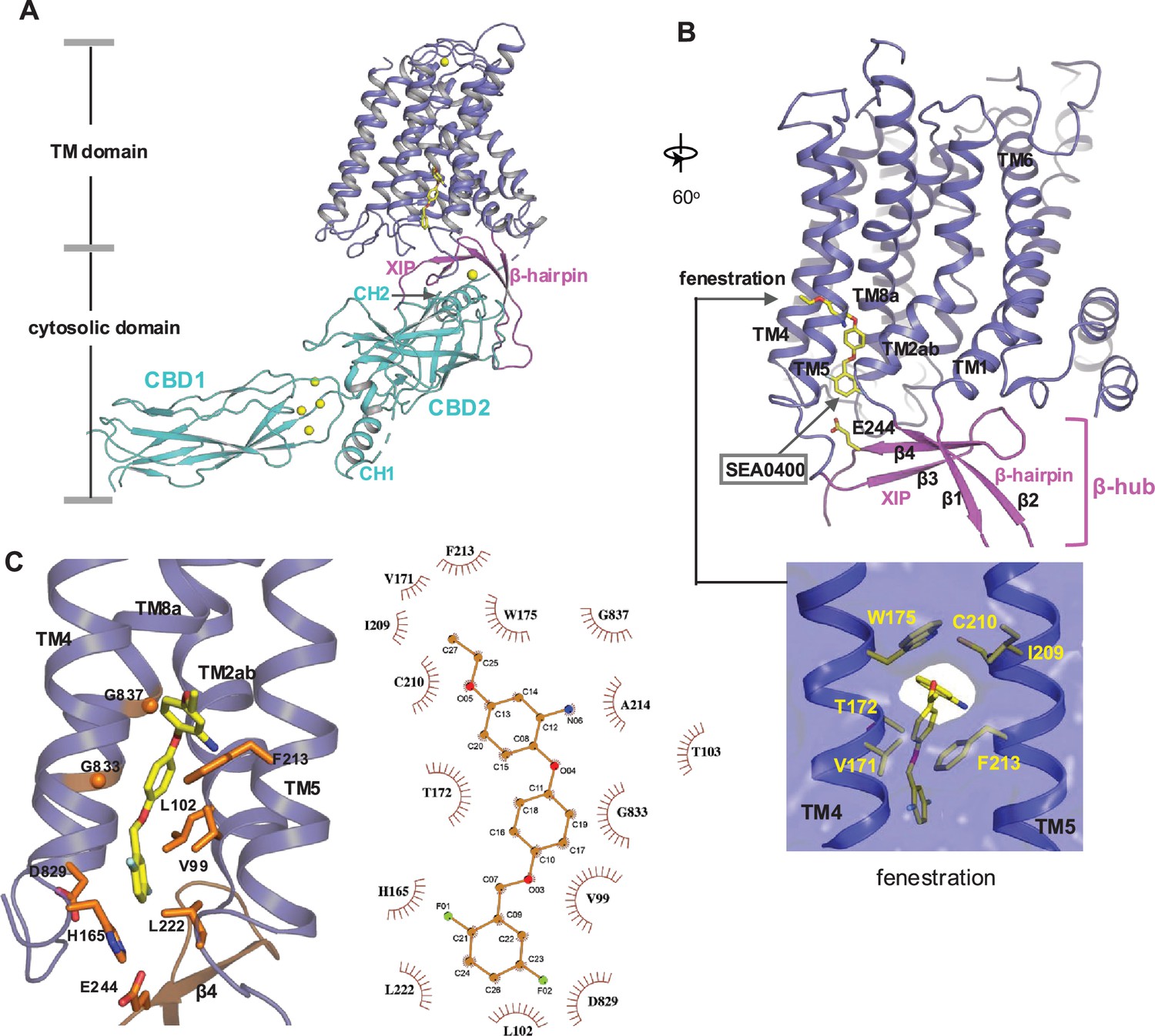
SEA0400 binding in NCX1.
(A) Overall structure of human NCX1 in complex with SEA0400 obtained in high Na+ and low Ca2+ conditions. Yellow spheres represent the bound Ca2+ in CBD1 and XIP. (B) Cartoon representation of the transmembrane (TM) domain and β-hub of the complex with surface-rendered view of the fenestration in the middle of the membrane. The β-hub is assembled by β-hairpin (β1 and β2) and XIP (β3 and β4). (C) Zoomed-in view of the SEA0400-binding site, and the schematic diagram detailing the interactions between NCX1 residues and SEA0400.
Figure 4—figure supplement 1
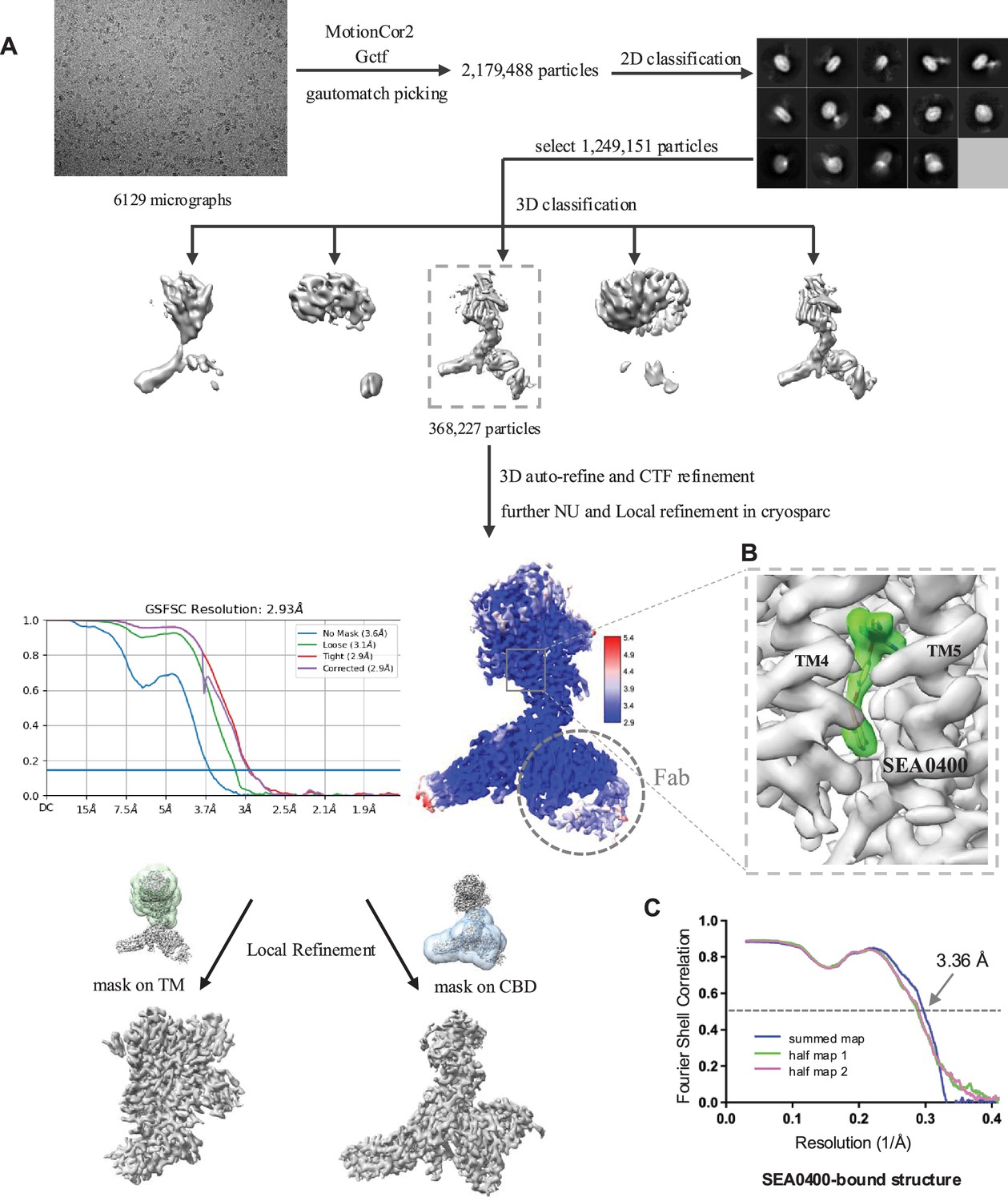
Structure determination of HsNCX1-SEA0400 complex.
(A) Cryo-EM data processing scheme of HsNCX1 in complex with SEA0400 inhibitor. The Fab fragment from a monoclonal antibody against NCX1 was used as a fiducial marker to facilitate the single-particle alignment. (B) Zoomed-in view of the density map of the bound SEA0400 inhibitor contoured at the threshold level of 0.52 using ChimeraX. (C) The Fourier shell correlation (FSC) curves for cross-validation between the maps and the models.
Figure 4—figure supplement 2

Structural comparison between the apo (gray) and SEA0400-bound (color) HsNCX1.
Figure 5
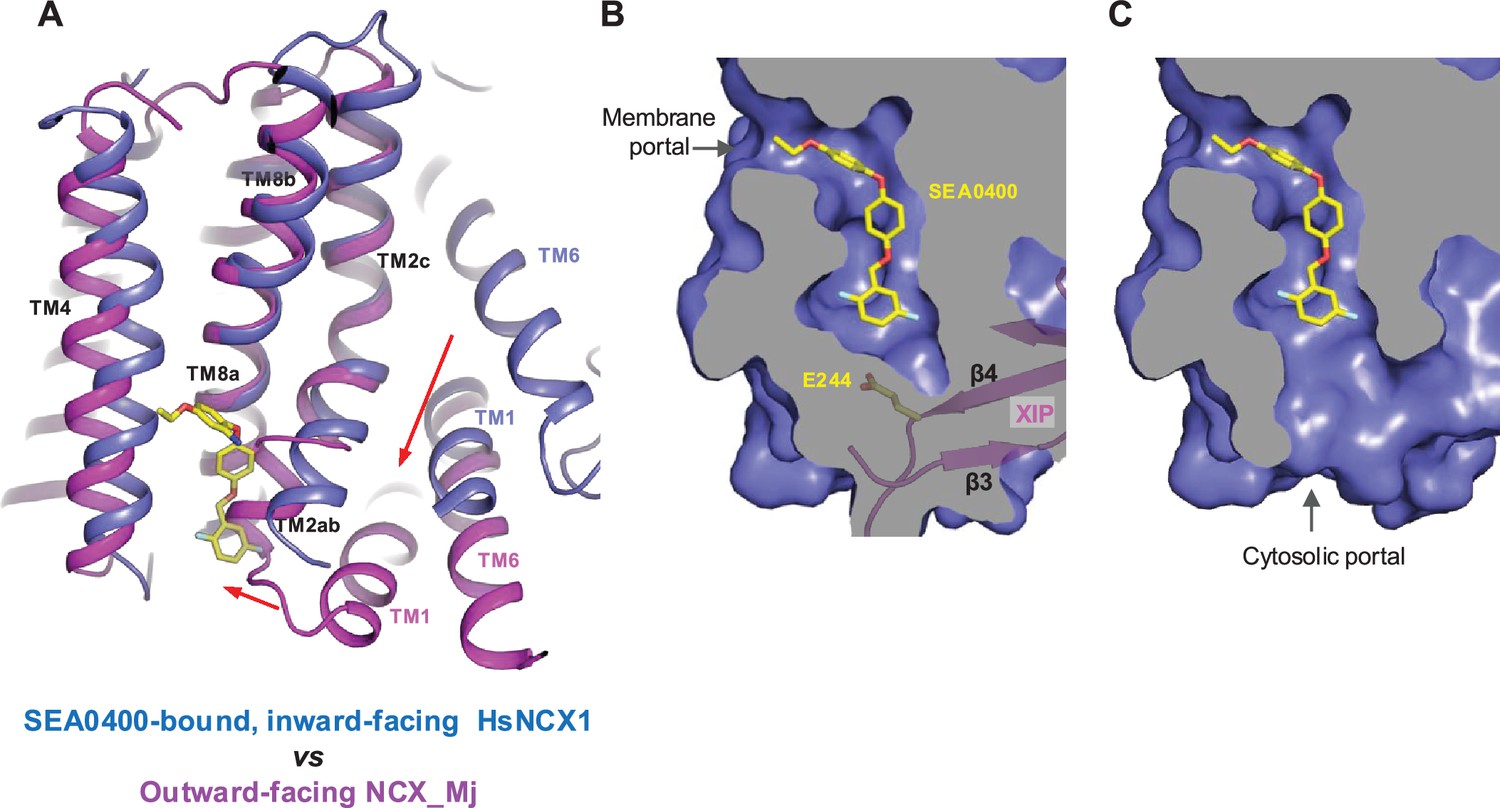
Structural mechanism of SEA0400 inhibition.
(A) Structural comparison at the core part of the transmembrane (TM) domain between the SEA0400-bound, inward-facing HsNCX1 and the outward-facing NCX_Mj (PDB 3V5U). Red arrows mark the sliding movement of TMs 1 and 6 and the bending of TM2ab from inward to outward conformation. (B) Surface-rendered views of the SEA0400-binding pocket sealed off from the cytosolic side by E244 from XIP in the inactivated state. (C) Removal of XIP would generate a cytosolic portal that facilitates the release of SEA0400.
Author response image 1
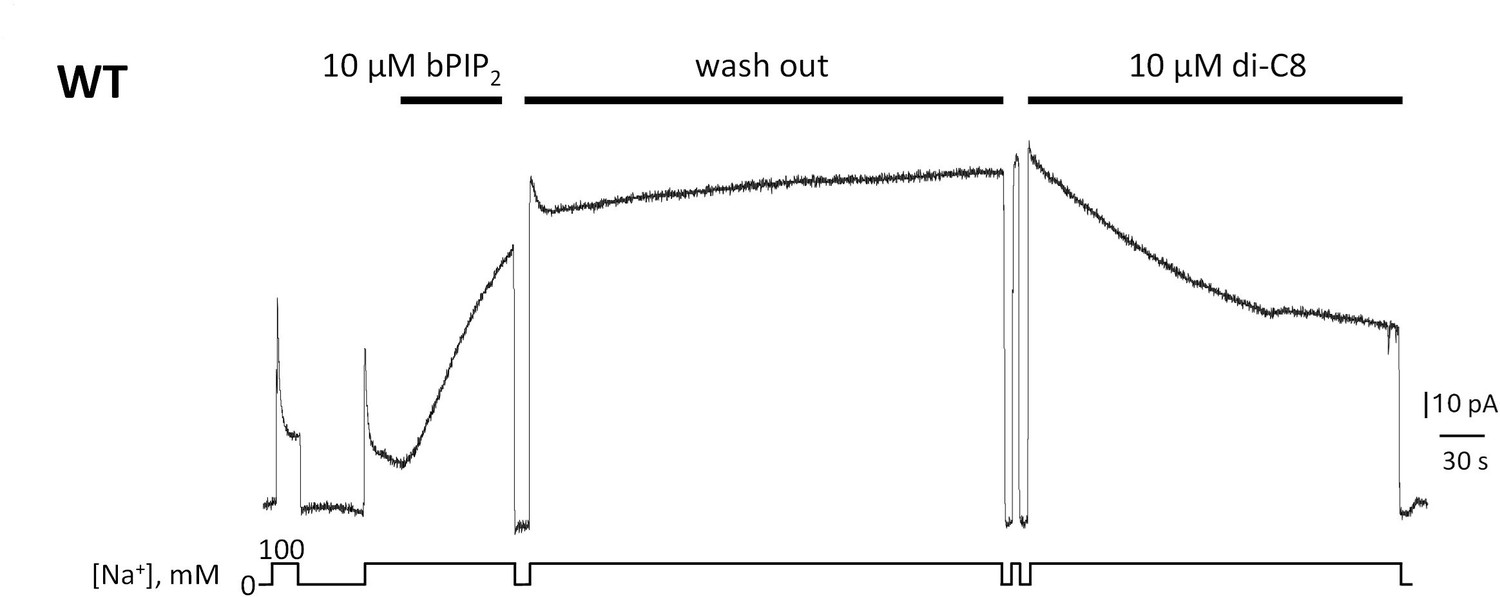
Giant patch recording of the human WT exchanger.
Currents were first activated by intracellular application of 10 µM brain PIP2. Afterwards, a solution containing 100 mM Na+ and 12 µM Ca2+ was perfused for about 5 min (washout). The PIP2 effects was not reversible during this time. The same patch was then perfused internally with the same solution in presence of 10 µM di-C8. Application of the shorted-chained di-C8, partially decreased the current suggesting that that PIP2 and diC8 compete for the binding site.
Tables
Table 1
Cryo-EM data collection and model statistics.
| Sample preparation conditions | 25 mM HEPES pH 7.4,200 mM NaCl0.9 mM SEA0400 | 25 mM HEPES pH 7.4,200 mM NaCl,0.47 mM PI(4,5)P2diC8 |
|---|---|---|
| SEA0400-bound state (EMD-40456, PDB 8SGI) | PI(4,5)P2diC8-bound state (EMD-60921, PDB 9IV8) | |
| Data collection and processing | ||
| Magnification | 105k | 105k |
| Voltage (kV) | 300 | 300 |
| Electron exposure (e–/Å2) | 60 | 60 |
| Defocus range (μm) | –0.9 to –2.2 | –0.9 to –2.2 |
| Pixel size (Å) | 0.83 | 0.84 |
| Symmetry imposed | C1 | C1 |
| Initial particle images (no.) | 1,249,151 | 2,233,044 |
| Final particle images (no.) | 368,227 | 117,748 |
| Map resolution (Å) FSC threshold | 2.93 0.143 | 3.47 0.143 |
| Refinement | ||
| Initial model used (PDB code) | 8SGJ | 8SGJ |
| Model resolution (Å) FSC threshold | 3.36 0.5 | 3.81 0.5 |
| Model composition Non-hydrogen atoms Protein residues Ligands | 7667 982 3: Na 6: Ca 1: H2O 1: SEA0400 | 5947 750 5: Ca 1: PI(4,5)P2diC8 |
| B factors (Å2) Protein Ligands | 66.21 58.55 | 50.47 106.20 |
| R.m.s. deviations Bond lengths (Å) Bond angles (°) | 0.005 0.696 | 0.004 0.679 |
| Validation MolProbity score Clashscore Poor rotamers (%) | 1.36 5.24 0 | 1.28 5.22 0 |
| Ramachandran plot Favored (%) Allowed (%) Disallowed (%) | 97.62 2.38 0 | 98.24 1.76 0 |
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Gene (Homo sapiens) | NCX1 (sodium/calcium exchanger 1) | Uniprot | P32418 | |
| Strain, strain background (Escherichia coli) | TOP10 | Thermo Fisher Scientific | Cat# 18258012 | |
| Strain, strain background (E. coli) | DH10Bac | Thermo Fisher Scientific | Cat# 10361012 | |
| Cell line (Spodoptera frugiperda) | Sf9 cells | Thermo Fisher Scientific | Cat# 11496015; RRID:CVCL_0549 | |
| Cell line (H. sapiens) | Expi293 GnTI- Cells | Thermo Fisher Scientific | Cat# A39240; RRID:CVCL_B0J7 | |
| Transfected construct (H. sapiens) | pEZT-BM-NCX1-Cstrep | This paper | N/A | |
| Recombinant DNA reagent | pEZT-BM | Morales-Perez et al., 2016 | RRID:Addgene_74099 | |
| Sequence-based reagent | NCX1_F_primer | This paper | PCR primers | gtacttaatacgactcactataggctagcgccaccatgtacaacatgcggcgattaagtc |
| Sequence-based reagent | NCX1_R_primer | This paper | PCR primers | gatggctccatgagccaccAgcggccgcgaagccttttatgtggcagtaggc |
| Chemical compound, drug | Sodium Butyrate | Sigma-Aldrich | Cat# 303410 | |
| Chemical compound, drug | Lauryl Maltose Neopentyl Glycol | Anatrace | Cat# NG310 | |
| Chemical compound, drug | Digitonin | Acros Organics | Cat# 11024-24-1 | |
| Chemical compound, drug | Biotin | Sigma-Aldrich | Cat# B4501 | |
| Software, algorithm | MotionCor2 | Zheng et al., 2017 | RRID:SCR_016499 | |
| Software, algorithm | GCTF | Zhang, 2016 | RRID:SCR_016500 | |
| Software, algorithm | RELION | Scheres, 2012 | RRID:SCR_016274 | http://www2.mrc-lmb.cam.ac.uk/relion |
| Software, algorithm | cryoSPARC | Punjani et al., 2017 | RRID:SCR_016501 | https://cryosparc.com/ |
| Software, algorithm | Chimera | Pettersen et al., 2004 | RRID:SCR_004097 | https://www.cgl.ucsf.edu/chimera |
| Software, algorithm | PyMol | Schrödinger | RRID:SCR_000305 | https://pymol.org/2 |
| Software, algorithm | Coot | Emsley et al., 2010 | RRID:SCR_014222 | https://www2.mrc-lmb.cam.ac.uk/personal/pemsley/coot/ |
| Software, algorithm | MolProbity | Chen et al., 2010 | RRID:SCR_014226 | http://molprobity.biochem.duke.edu/ |
| Software, algorithm | PHENIX | Adams et al., 2010 | RRID:SCR_014224 | https://www.phenix-online.org |
| Other | Superdex 200 Increase 10/300 GL | Cytiva | Cat# 28990944 | |
| Other | Strep-Tactin resin | IBA | Cat# 2-5010 | |
| Other | Amicon Ultra-15 Centrifugal Filter Units | MilliporeSigma | Cat# UFC9100 | |
| Other | Quantifoil R 1.2/1.3 grid Au300 | Quantifoil | Cat# Q37572 | |
| Other | Cellfectin | Thermo Fisher Scientific | Cat# 10362100 | |
| Other | Sf-900 II SFM medium | Thermo Fisher Scientific | Cat# 10902088 | |
| Other | FreeStyle 293 Expression Medium | Thermo Fisher Scientific | Cat# 12338018 | |
| Other | Antibiotic Antimycotic Solution | Sigma-Aldrich | Cat# A5955 |
Additional files
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Structural mechanisms of PIP2 activation and SEA0400 inhibition in human cardiac sodium-calcium exchanger NCX1
eLife 14:RP105396.
https://doi.org/10.7554/eLife.105396.3




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}