Loss of olfaction reduces caterpillar performance and increases susceptibility to a natural enemy
- Laboratory of Entomology, Wageningen University and Research, Netherlands
- Biosystematics Group, Wageningen University & Research, Netherlands
- Business Unit Bioscience, Wageningen University & Research, Netherlands
eLife Assessment
This important manuscript investigates the role of olfactory cues in Pieris brassicae larvae, focusing on their interactions with the host plant Brassica oleracea and the parasitoid wasp Cotesia glomerata. The authors' demonstration that impaired olfactory perception reduces caterpillar performance and increases susceptibility to parasitism is convincing. These findings highlight the ecological significance of olfaction in mediating feeding behavior and predator avoidance in herbivorous insects.
https://doi.org/10.7554/eLife.105585.3.sa0Significance of the findings:
Important: Findings that have theoretical or practical implications beyond a single subfield
- Landmark
- Fundamental
- Important
- Valuable
- Useful
Strength of evidence:
Convincing: Appropriate and validated methodology in line with current state-of-the-art
- Exceptional
- Compelling
- Convincing
- Solid
- Incomplete
- Inadequate
During the peer-review process the editor and reviewers write an eLife Assessment that summarises the significance of the findings reported in the article (on a scale ranging from landmark to useful) and the strength of the evidence (on a scale ranging from exceptional to inadequate). Learn more about eLife Assessments
Abstract
Insect herbivores such as caterpillars are under strong selection pressure from natural enemies, especially parasitoid wasps. Although the role of olfaction in host-plant seeking has been investigated in great detail in parasitoids and adult lepidopterans, the caterpillar olfactory system and its significance in tri-trophic interactions remain poorly understood. In this study, we investigated the olfactory system of Pieris brassicae caterpillars and the importance of olfactory information in the interactions among this herbivore, its host-plant Brassica oleracea, and its primary natural enemy Cotesia glomerata. To examine the role of olfaction, we utilized CRISPR/Cas9 to knockout (KO) the odorant receptor co-receptor (Orco). This KO impaired olfactory detection and primary processing in the brain. Orco KO caterpillars exhibited reduced weight and lost preference for their optimal food plants. Interestingly, the KO caterpillars also experienced reduced weight when challenged by the parasitoid C. glomerata whose ovipositor had been removed, and the mortality of the KO caterpillars under the attack of unmanipulated parasitoids increased. We then investigated the behavior of P. brassicae caterpillars in response to volatiles from plants attacked by conspecific caterpillars and volatiles from plants on which the caterpillars were themselves attacked by C. glomerata. After analyzing the volatile compounds involved in these interactions, we concluded that olfactory information enables caterpillars to locate suitable food sources more efficiently as well as to select enemy-free spaces. Our results reveal the crucial role of olfaction in caterpillar feeding and natural-enemy avoidance, highlighting the significance of chemoreceptor genes in shaping ecological interactions.
eLife digest
Many caterpillars are major pests in agriculture, feeding on a variety of crops. They constantly face threats from both predators and the toxic defenses of the plants they eat. While scientists have long studied how predators find their prey, much less is known about how prey – like caterpillars – manage to survive and defend themselves.
Plants have evolved various strategies to fend off insects. When attacked by herbivores, some plants release specific chemical signals known as herbivore-induced plant volatiles. These scents act as a distress call, attracting natural enemies of the herbivores, such as parasitoid wasps, which use the caterpillars for reproduction, killing the herbivores in the process.
In turn, caterpillars have evolved different defense strategies. They have a sophisticated sense of smell, which may help them detect not only the scent of host plants but also the presence of predators or other caterpillars nearby.
Despite how common and important caterpillars are in farming systems, it is still not fully understood how they choose where to feed or how they avoid being eaten. To explore this, Wang et al. studied how much caterpillars rely on their sense of smell to survive and find food.
In insects, the ability to smell relies on a group of sensory proteins, most of which need a key gene called Orco to function. Without this gene, an insect’s ability to smell is severely impaired. Wang et al. used a gene editing tool known as CRISPR/Cas9 to ‘turn off’ the Orco gene in caterpillars of the large cabbage white butterfly (Pieris brassicae), effectively disabling their sense of smell. The researchers then compared the development, survival performance, and behavioral preferences of these mutant caterpillars with normal ones.
This revealed that caterpillars without a working sense of smell gained less weight and were less successful at finding suitable food sources. They were also more likely to be killed by parasitoid wasps. Behavioral experiments showed that caterpillars with an intact Orco gene avoided plants where other caterpillars were under attack by detecting the warning signals in the air. In contrast, mutant caterpillars could not recognize these danger cues and were unable to find suitable host plants or select enemy-free spaces.
The study of Wang et al. shows that caterpillars use smell not only to find food, but also to avoid danger. These insights could help farmers develop new, environmentally friendly ways to manage pests. For example, by planting companion plants that produce specific scents, or by breeding crops that naturally repel pests, we might be able to steer caterpillars away from valuable crops without relying on chemical pesticides.
Introduction
Chemoreception plays a critical role in the insect life cycle. For instance, insects exploit chemical cues to locate food plants, to find and select mates, to evaluate oviposition sites, and to avoid natural enemies (Dicke and Grostal, 2001). Olfaction, a pivotal component of chemoreception, involves multiple processes in insects and has been extensively investigated in many species (Haverkamp et al., 2018), including its role in the interactions with conspecifics and natural enemies (Kannan et al., 2022). Thus far, the study of olfaction in insects has largely centered upon imagos, the final life stage of insects. However, chemical information likely also plays an essential role in host-plant choice and natural-enemy avoidance during the immature life stage, which is especially pronounced in holometabolous insects. Nonetheless, the olfactory system in larval herbivores remains largely unexplored.
Tri-trophic interactions among plants, herbivores, and natural enemies have been widely investigated in the last few decades, and chemoreception is considered to play an important role in these systems (Vet and Dicke, 1992; Turlings and Erb, 2018). In particular, abundant research has demonstrated that olfactory information, such as plant volatiles, is crucial in the coevolution of herbivores and natural enemies (Turlings and Erb, 2018; Dicke and Baldwin, 2010; Haverkamp and Smid, 2020). In addition to plant volatiles, chemicals emitted by the insects themselves also play an important role (Venugopal et al., 2020; Ebrahim et al., 2015). Under strong selection pressure from natural enemies, herbivores have developed several integrated avoidance strategies based on the detection of chemical cues. Drosophila melanogaster fruit flies, for example, prefer to lay eggs on citrus fruits because the volatiles emitted by citrus are repellent to the endoparasitoid Leptopilina boulardi, a natural enemy of fruit fly larvae (Dweck et al., 2013). Similarly, the brown planthopper Nilaparvata lugens deposits eggs on rice plants infested by the striped stem borer Chilo suppressalis, which is repellent to the egg parasitoid Anagrus nilaparvatae (Hu et al., 2020).
Insect olfaction is dominated by two classes of olfactory receptor genes, odorant receptors (ORs) and ionotropic receptors (IRs). ORs detect the majority of volatiles in the insect habitat. In all ORs, the odorant receptor co-receptor (Orco) is required to form a functional ion channel in the membrane of the odorant receptor neurons (ORNs) (Sato et al., 2008; Wicher et al., 2008; Jones et al., 2005). Previous research indicates that silencing the Orco gene can lead to a severe olfaction deficiency, causing a loss of social behavior, impaired mating, and significantly reduced foraging behavior (Trible et al., 2017; Yan et al., 2017; Fandino et al., 2019). Previous reports have also clearly shown that adult insect behavior and neuronal development were greatly affected when Orco is knocked out (Trible et al., 2017; Yan et al., 2017). Yet, potential changes in olfaction-deficient larvae remain largely unexplored, and little is known about how olfaction contributes to the co-adaptation of larvae and their natural enemies, especially in the larvae of non-model insects such as lepidopteran caterpillars.
Pieris brassicae is a very common butterfly species that causes substantial agricultural production loss across Europe and the Indian subcontinent (Feltwell, 1982; Hasan and Ansari, 2011; Hasan and Shafiq Ansari, 2010). The butterflies utilize chemoreception to evaluate oviposition sites and host plants (van Loon et al., 1992a; van Loon et al., 1992b; Wang et al., 2023). The caterpillars also have a well-developed olfactory system (Wang et al., 2024) and chemical communication has been demonstrated to be involved in the interaction between herbivores and their natural enemies (Stam et al., 2014; Dicke and Baldwin, 2010). Natural enemies such as parasitoid wasps are highly attracted to volatiles from cabbage plants infested by P. brassicae caterpillars (Mattiacci et al., 1994). P. brassicae caterpillars are thus under strong selection pressure from natural enemies, though caterpillars can defend themselves by spitting at natural enemies while being attacked (Müller et al., 2003).
To gain comprehensive insight into the role of olfaction in P. brassicae, and to understand the role of olfaction in their coevolution with natural enemies, we employed a well-established research system utilizing Brassica oleracea as host plants, P. brassicae caterpillars as herbivores and Cotesia glomerata parasitoid wasps as natural enemies. In this study, we knocked out Orco by CRISPR/Cas9 in P. brassicae to investigate how olfaction guides the host-plant choice and natural-enemy avoidance of this insect herbivore. The knockout (KO) was verified in caterpillars by staining Orco and glomeruli in larval antennae and the larval antennal center (LAC), respectively, and was further confirmed in butterflies by electrophysiological investigations. We also examined the egg-laying behavior of KO butterflies, which is the basis of establishing a homozygous KO insect colony for our subsequent studies. The development of caterpillars on cabbage plants was evaluated, and the performance of caterpillars under the threat of natural enemies was studied to investigate the ecological significance of larval olfaction in interacting with their natural enemies. The larval host-plant seeking behaviors were further evaluated to determine the importance of olfaction in locating food sources. Volatiles emitted by caterpillars and parasitoids were analyzed to identify candidate chemicals that might play a role in this interaction. Our study provides further knowledge on the olfactory system in insect herbivores and might help to clarify the ecological significance of olfaction in the interaction with their natural enemies.
Results
Orco KO and neuronal deficiency
To test for potential off-target regions of our sgRNAs, we searched these sequences against the P. brassicae genome to identify any such regions. Neither did we find any off-target sites with a less-than-5-nt alignment mismatch from Exonerate searching results, nor did we find any off-target sites with a less-than-4-nt alignment mismatch from CHOPCHOP searching results. Furthermore, we did not find any developmental differences between wildtype (WT) animals and KO when caterpillars were raised on leaf discs in Petri dishes (Figure 1—figure supplement 1) or in the hatching rate of fertilized WT and KO eggs (Figure 1—figure supplement 2). Taken together, these results indicate that no off-target sites were available for our sgRNA. Through screening all butterflies that were reared from injected eggs using PCR and sequencing, we identified an Orco mutant line in P. brassicae. This mutant line exhibited an 11-bp insertion and a 4-bp deletion in the second exon, resulting in a shifted reading frame (Figure 1A). The Orco amino acid sequence showed a significant change mediated by an early stop codon (Figure 1B). These results indicate that the KO of the Orco gene was successful.
Figure 1 with 4 supplements see all
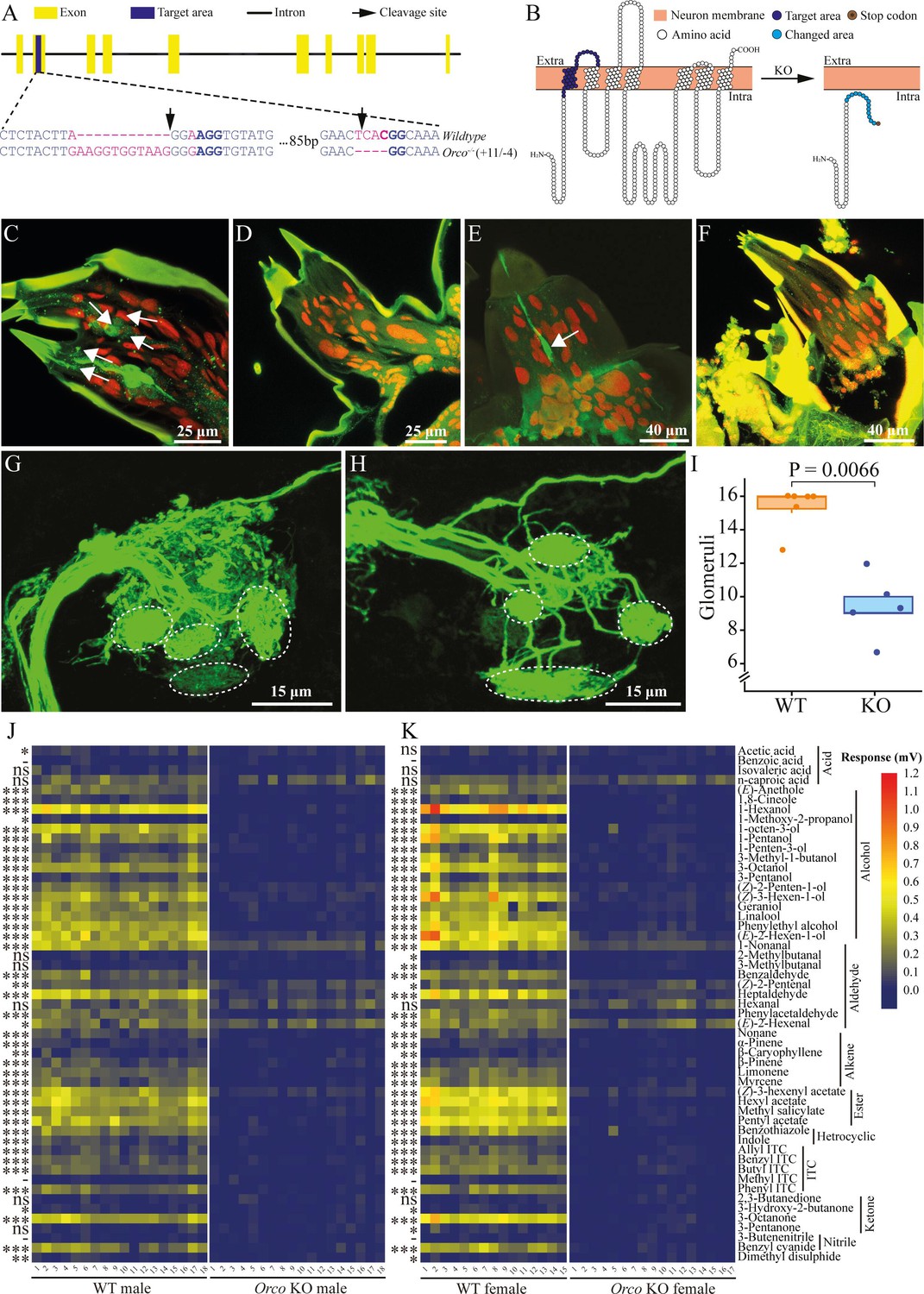
Orco knockout (KO) by CRISPR/Cas9 and verification in Pieris brassicae.
(A) Gene structure of Orco. Yellow blocks indicate exons, and blue blocks indicate target areas in the second exon. Black solid line indicates intron. Black arrows indicate the designated cleavage site of the Cas9 protein. The target KO area is magnified to show the sequence. Blue letters indicate base pairs in the second exon segment, purple letters indicate mutation sites. Protospacer adjacent motif (PAM) sequences are in blue bold. (B) Predicted transmembrane structure of Orco. The left and right panels represent wildtype (WT) Orco and mutated Orco transmembrane domains, respectively. Orange blocks indicate odorant receptor neuron (ORN) membrane, extracellular (Extra), and intracellular (Intra) are shown. White circles indicate amino acids of Orco, blue circles on the left panel indicate the target mutation area. Cyan circles and the brown circle on the right panel indicate the mutated area and the early stop codon, respectively. Orco-positive ORNs and larval antennal center (LAC). P. brassicae larval antennae and LAC staining. (C) ORNs were stained (green cells, several are indicated by white arrows) in WT larval antennae. (D) ORNs were not stained in Orco KO larval antennae. (E) An ORN was stained (green cell, indicated by white arrow) in WT larval palps. (F) ORNs were not stained in KO larval palps. (G) Glomeruli in the LAC of a WT brain. (H) Glomeruli in the LAC of a KO brain. Glomeruli are indicated by white dashed circles in (G) and (H) (not directly corresponding). (I) Number of glomeruli counted in WT caterpillar brain (orange) and Orco KO caterpillar (blue) brain (n = 6 for WT and n = 5 for KO). A significant difference was detected by the Wilcoxon rank-sum test. (J) Electroantennogram (EAG) response of male butterflies (n = 18 for both WT and KO). (K) EAG response of female butterflies (n = 15 for WT and n = 17 for KO). Left panels represent WT butterfly EAG responses, and right panels represent KO butterfly EAG responses. Significant differences between WT and KO butterflies were identified using Student’s t-test when the data were normally distributed and the variances were equal or using a Kruskal–Wallis rank-sum test when these criteria did not apply. Significance levels are indicated by asterisks, ns: p > 0.05; *0.01 < p < 0.05; **0.001 < p < 0.01; ***p < 0.001; -: no response recorded from WT butterfly antennae. Butterfly antennal responses (mV) to the tested chemicals are indicated by a color scale from navy (0 mV) via yellow to red (1.2 mV).
To further validate the KO in P. brassicae and investigate the neuronal mechanism underlying olfactory deficiency in KO caterpillars, we performed staining of Orco in third-instar larvae (L3) using a lepidopteran-specific Orco antibody, which has been validated previously (Fandino et al., 2019; Wang et al., 2024; Nolte et al., 2016). In L3 WT caterpillar antennae, a total of 34 ORNs were successfully identified (Wang et al., 2024; Figure 1C). However, no Orco-positive ORNs were found in the antennae of KO caterpillars (Figure 1D). Similarly, we detected a low number of Orco-positive neurons in the maxillary palps of L3 WT caterpillars (Figure 1E), but no Orco-positive signal was detected in the maxillary palps of KO caterpillars (Figure 1F). By tracing the ORN axons to their glomeruli in the brain, we observed some differences between WT and Orco KO caterpillars. In the LAC, glomeruli displayed well-defined shapes with clear boundaries in the WT LAC (Figure 1G). However, in the KO LAC, the glomeruli were less clearly separated (Figure 1H). Only 7–12 glomeruli were identified in KO LAC, which was a significant reduction (p = 0.0066, Wilcoxon rank-sum test) compared to WT LAC (Figure 1I).
Butterfly electrophysiological response to chemical compounds and oviposition
We then conducted additional tests comparing the antennal electrophysiological responses of male and female butterflies from both genotypes using electroantennography (EAG), to further confirm the KO of Orco in butterflies. Orco KO butterflies of both sexes displayed a complete loss of responses to the tested chemicals with the exception of isovaleric acid, n-caproic acid, hexanal, and 2,3-butanedione, where no significant difference was detected between KO and WT butterflies. Reduced responses were recorded in KO butterflies to 1-nonanal, (Z)-2-pentenal, (E)-2-hexenal, and (Z)-3-hexenyl acetate. Besides, WT male (n = 18) and female (n = 15) butterflies responded similarly to most of the volatile compounds with a few exceptions where WT females showed significantly different responses to 2-methylbutanal (p = 0.0183, Kruskal–Wallis test), 3-methylbutanal (p = 0.0030, Kruskal–Wallis test), and 3-pentanone (p = 0.0263, Student’s t-test) compared to KO females (n = 17), and WT males (n = 18) responded significantly stronger to acetic acid than KO males (p = 0.0217, Student’s t-test). Moreover, both WT males and females showed no response to benzoic acid, methyl isothiocyanate (ITC), and 3-butenenitrile (p > 0.05, Student’s t-test) (Figure 1J, K).
To investigate the role of olfaction in butterfly mating and oviposition and evaluate the feasibility of establishing a homozygous Orco KO colony for subsequent larval studies, pairs of newly emerged butterflies were collected and reared in a cage and allowed to mate freely. Upon dissection, it was found that KO butterflies (n = 14) mated fewer times than WT butterflies (p = 0.0199, n = 11; Wilcoxon rank-sum test), since the majority of the KO butterflies (9 out of 14) did not mate during the experiment (Figure 1—figure supplement 3). Out of the five KO butterflies that did mate, all but one mated singly. Of the WT butterflies, 8 out of 11 mated, with most mating twice and one individual even thrice. Female KO butterflies also deposited fewer eggs than WT butterflies during the experiment (p < 0.0001, generalized linear model [GLM] negative binomial) (Figure 1—figure supplement 4). To determine the hatching rate, the number of newly hatched caterpillars of both genotypes was counted. It was found that virgin butterflies of both genotypes did not deposit any fertilized eggs, while mated butterflies exhibited similar fecundity with comparable hatching rates between the two genotypes without any significant difference (Figure 1—figure supplement 2).
Caterpillar performance on cabbage plants
To assess the ecological significance of knocking out Orco in P. brassicae caterpillars, we first conducted a comparison of caterpillar growth between the two genotypes on their optimal host plant. The results show that in the absence of parasitoids, WT caterpillars exhibited better performance, gaining more weight compared to KO caterpillars (p = 0.0268, n = 17 for both genotypes; Student’s t-test) (Figure 2A). However, we did not find significant survival differences between the two genotypes while collecting caterpillars after the experiment (WT mortality = 8.8 %, n = 17; KO mortality = 2. 9%; p = 0.0880, n = 17; Wilcoxon test). Nor was there a developmental difference when caterpillars were reared in a Petri dish supplied with leaf discs (p = 0.9698, n = 10 for both genotypes; Wilcoxon test) (Figure 1—figure supplement 1). Subsequently, to determine if the differences in feeding and survival are dependent on food-plant location, a two-choice assay was conducted with cabbage leaf discs, tomato leaf discs, and green paper discs in a glass Petri dish to investigate the role of olfaction in host-plant selection (Figure 2B). Both genotypes of caterpillars chose the cabbage leaf disc significantly more often than the green paper discs (p = 0.0016 for KO, χ2 = 9.92; p < 0.0001 for WT, χ2 = 21.41; Chi-square test), and no difference was detected between the two genotypes (p = 0.1460, χ2 = 2.11; Chi-square test). Similarly, when the cabbage leaf disc was replaced with a tomato leaf disc, both genotypes preferred the tomato leaf disc over the paper disc (p = 0.0143 for KO, χ2 = 6.00; p = 0.0009 for WT, χ2 = 10.97; Chi-square test) and no difference was found between the two genotypes (p = 0.5477, χ2 = 0.36; Chi-square test). However, when caterpillars were provided with both a cabbage leaf disc and a tomato leaf disc, a contrasting preference was observed between the two genotypes (p = 0.0004, χ2=12.68; Chi-square test). KO caterpillars significantly preferred the tomato leaf disc (p = 0.0466, χ2 = 3.96; Chi-square test), while WT caterpillars preferred the cabbage leaf disc (p = 0.0028, χ2=8.97; Chi-square test).
Figure 2
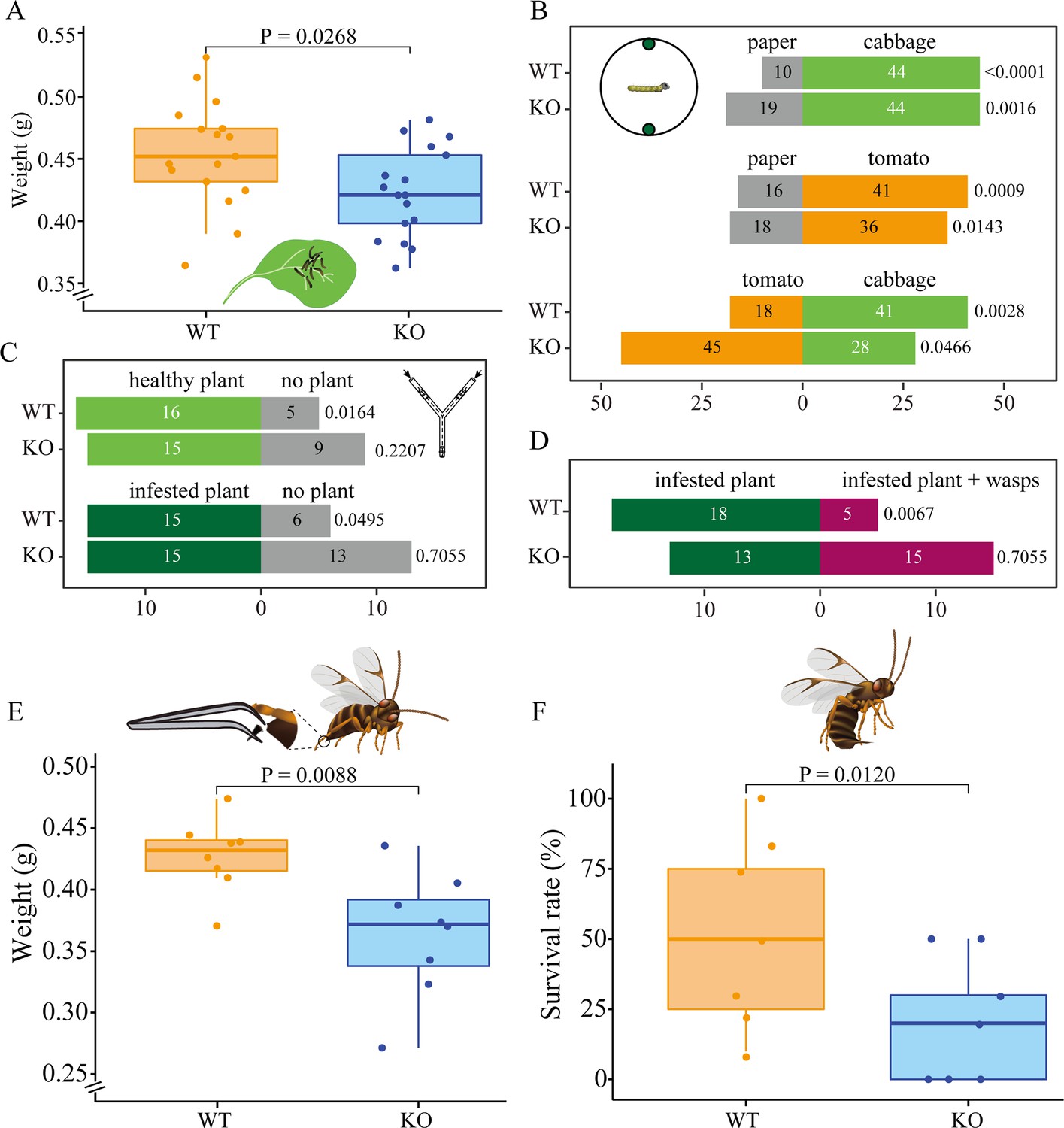
Pieris brassicae caterpillar growth and foraging behavior.
(A) P. brassicae caterpillar growth on cabbage plants, y-axis shows weight (g) of wildtype (WT) and knockout (KO) caterpillars after 10 days of feeding, n = 17. (B) Caterpillar behavioral choices in a Petri dish. The numbers of caterpillars that chose one of the two discs are shown in the respective bars (n = 54–73). Schematic drawing shows behavioral setup. Petri dish diameter was 188 mm, disc diameters were 13 mm. Discs in the Petri dish represent cabbage, paper, and tomato leaf discs. Gray bars indicate paper disc choices, light green indicates cabbage leaf disc choices, and orange indicates choices for tomato leaf discs. (C) Caterpillar behavioral choices in Y-tube olfactometer without parasitoid wasps. A schematic drawing shows the Y-tube olfactometer. The dashed line indicates a black metal Y wire in the center of glass Y-tube olfactometer. The main arm of the Y-olfactometer is 200 mm length, the lateral arms are 275 mm length, and the angle between the lateral arms is 80°. Light green, dark green, and gray bars represent choices for healthy plant, infested plant, and no plant, respectively. Healthy plant, plants were not treated; no plant, an empty jar without any insect or plant; infested plant, plants were infested by early L3 caterpillars. (D) Caterpillar behavioral choices in Y-tube olfactometer with parasitoid wasps. Different treatments are in different colors. Dark green, infested plant − wasps; magenta, infested plant + wasps. Significant differences were tested between WT and KO caterpillars or between two discs by Chi-square test in panels B–D, p-values are shown on the right side of each bar. (E) P. brassicae caterpillar growth when exposed to disarmed C. glomerata female parasitoids, y-axis shows weight (g) of caterpillars after 10 days of feeding, n = 8 for both genotypes. Schematic drawing shows disarmed female C. glomerata (ovipositor removed). (F) P. brassicae caterpillar survival rate when exposed to healthy C. glomerata female wasps, n = 7 for both genotypes. Significant differences in development and survival rate were assessed using a one-tailed Student’s t-test, p-values are indicated above boxplots. A schematic drawing shows a healthy female C. glomerata (unmanipulated). In both panels, orange boxplots indicate WT caterpillars, and blue boxplots indicate Orco KO caterpillars.
We then aimed to determine whether caterpillars would orient toward healthy plants, plants infested with conspecifics, and plants on which conspecifics are attacked by parasitoid wasps, using a Y-tube olfactometer. We first compared the behaviors of both WT and Orco KO caterpillars in response to clean air, volatiles from a healthy plant, and volatiles from a caterpillar-infested plant (Figure 2C). Both WT and Orco KO caterpillars tended to prefer a healthy cabbage plant to a clean air control; however, WT caterpillars were significantly attracted by volatiles from a healthy plant (p = 0.0164, χ2 = 5.76; Chi-square test) while the preference for Orco KO caterpillars was not significant (p = 0.2207, χ2 = 1.50; Chi-square test). Subsequently, we challenged the caterpillars with volatiles from an infested plant and a clean air control and found that WT caterpillars prefer volatiles from the infested plant to the control (p = 0.0495, χ2 = 3.86; Chi-square test), while Orco KO caterpillars did not show any preference between the two provided options (p = 0.7055, χ2 = 0.14; Chi-square test). We further compared larval host-plant seeking behaviors with the presence of natural enemies on caterpillar-infested plants (Figure 2D). WT caterpillars showed a significant preference for infested plants without parasitoid wasps C. glomerata (p = 0.0067, χ2 = 7.35; Chi-square test), while Orco KO caterpillars did not show any preference for either odor source (p = 0.7055, χ2 = 0.14; Chi-square test).
To understand the ecological significance of the odor-guided behavior for the natural-enemy avoidance in P. brassicae caterpillars, we then evaluated caterpillar performance under the potential threat of natural enemies by exposing the caterpillar to parasitoids after removing the ovipositor of the female wasps (Figure 2E). The results show a significantly higher weight in WT caterpillars than in KO caterpillars (p = 0.0088, n = 8 for both genotypes; Student’s t-test). To further explore the effect of knocking out Orco on the interaction of the caterpillars with their natural enemies, we compared the survival rates of caterpillars when exposed to two mated unmanipulated female C. glomerata wasps. The findings show that KO caterpillars had significantly lower survival rates compared to WT caterpillars (p = 0.0120, n = 7 for both genotypes; GLM beta-binomial) (Figure 2F).
Volatile headspace composition
Furthermore, we investigated if caterpillar- and parasitoid-derived chemicals are involved in the behavioral responses. We identified a total of 45 chemical compounds among the five treatments belonging to seven classes, including alcohols, aldehydes, aromatics, ketones, nitrogen and/or sulfur-containing chemicals, terpenoids, and others (Supplementary file 1A). The contribution of identified chemicals toward separating different treatment groups was analyzed using principal component analysis (PCA). This analysis showed a significant separation of most groups with PC 1 carrying 65.4% of the variance (p = 0.001, PERMANOVA). P. brassicae caterpillars (Pb, n = 10), P. brassicae caterpillar spit (Pb-S, n = 12), C. glomerata female wasps (Cg, n = 12), and caterpillar frass (Pb-Fr, n = 10) treatment groups were clearly separated (Pb vs. Cg, Pb vs. Pb-S, Pb vs. Pb-Fr, Pb-S vs. Cg, and Pb-Fr vs. Cg, p = 0.0013; Pb-S vs. Pb-Fr, p = 0.0022 pairwise PERMANOVA, Figure 3A), whereas the identified and measured volatile blend from Pb was similar to that of the P. brassicae caterpillars–C. glomerata parasitoids combination (Pb–Cg, n = 14) (p = 0.0330, pairwise PERMANOVA). The amount of volatiles among the five treatments was also compared. Most of the identified compounds were found in significantly higher amounts in Pb-S and/or Pb-Fr compared to the other treatments (Figure 3B, Supplementary file 1A). We also found that some chemicals are strongly correlated. The identified chemicals are classified into several subgroups in the hierarchical clustering heatmap according to the abundance among treatments. For instance, the seven chemicals (Z)-3-hexen-1-ol, 1-penten-3-ol, dimethyl disulfide, methyl (methylthio) methyl sulfide, dimethyl trisulfide, 2-methylbutanal, and 3-methylbutanal formed a clade. Similarly, the six chemicals, 3-methylbutanoic acid, 1-pentanol, benzyl cyanide, 3-(methylthio) propanal, (Z)-2-hexen-1-ol, and phenylacetaldehyde formed another clade. The clustering analysis demonstrated that chemicals within the same clade are highly correlated. The multivariate data analysis (MVDA) also showed similar clustering when comparing the different treatments (Figure 3—figure supplements 1–5).
Figure 3 with 7 supplements see all
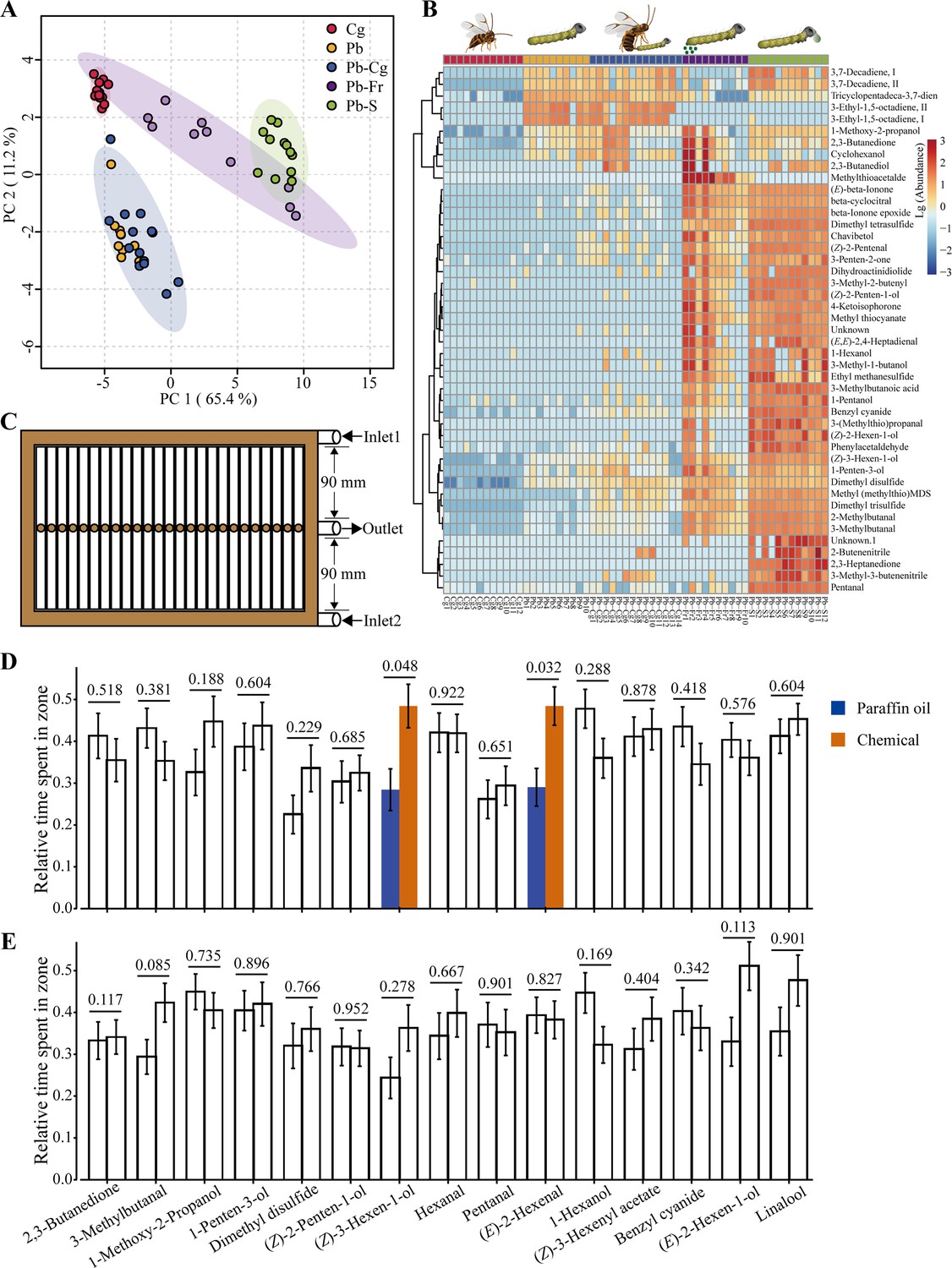
Overview of caterpillar- (Pieris brassicae) and parasitoid wasp- (Cotesia glomerata) associated volatile compounds.
(A) PCA (principal component analysis) two-dimensional score plot of five treatment groups: Cg, Cotesia glomerata female parasitoid wasps (n = 12); Pb, Pieris brassicae caterpillars (n = 10); Pb–Cg, P. brassicae caterpillars in the presence of C. glomerata female parasitoid wasps (n = 14); Pb-Fr, P. brassicae caterpillar frass (n = 10); and Pb-S, P. brassicae spit (n = 12), based on their volatile blend composition. (B) Hierarchical clustering heatmap showing the abundance of each identified volatile compound of each treatment. Clustering chemicals in the heatmap indicates higher correlation. (C) Schematic drawing of the custom-designed multichannel arena. (D) Wildtype (WT) caterpillar behavioral preference to the tested 15 odorants. (E) Orco knockout (KO) caterpillar behavioral preference to the tested 15 chemicals. Unfilled bars represent tests that exhibit no difference between paraffin oil and the odorant of interest. Blue bars indicate the cumulative duration ratio that caterpillars stayed in the paraffin oil zone, dark orange bars indicate the cumulative duration ratio that caterpillars stayed in the odorant zone. Relative time spent in odorant zone = Cumulative duration in a specific zone/Cumulative duration in the arena. Error bars indicate standard errors. In (D) and (E), differences were tested between the two zones using a Wilcoxon rank-sum test, p-values are presented above bars (n = 31–50). For each comparison, the left bar represents paraffin oil and the right bar represents chemical compound.
Based on the results of our chemical analysis, we selected 11 candidate chemicals (1-hexanol, 1-methoxy-2-propanol, 1-penten-3-ol, 2,3-butanedione, 3-methylbutanal, benzyl cyanide, dimethyl disulfide, pentanal, (E)-2-hexen-1-ol, (Z)-2-penten-1-ol, and (Z)-3-hexen-1-ol) with high variable importance in the projection scores (>1.0) (Supplementary file 1A–F), and another four common plant volatiles (linalool, (Z)-3-hexenyl acetate, hexanal, and (E)-2-hexenal). These odorants were chosen to test the behavioral response of L3 caterpillars. Using a custom-made multiple channel setup (Figure 3C), we conducted behavioral experiments with the caterpillars. The results showed that WT caterpillars displayed a preference for two plant chemicals, namely (Z)-3-hexen-1-ol and (E)-2-hexenal over paraffin oil (p = 0.0482 and p = 0.0321) (Figure 3D). In contrast, KO caterpillars did not exhibit any significant attraction or avoidance behavior toward the tested chemicals (Figure 3E). Additionally, we found that WT caterpillars were more active in the arena, moving around (Figure 3—figure supplement 6), while KO caterpillars were less active in the arena, often remaining at the same position within the channels (Figure 3—figure supplement 7).
Discussion
Chemical communication plays a significant role in the interaction between plants, herbivores, and their natural enemies. Infested host plants emit herbivore-induced plant volatiles (HIPVs) and thereby attract natural enemies such as parasitoid wasps (Mattiacci et al., 1994). While the role of adult herbivores’ olfaction in mediating these tri-trophic interactions has been investigated, the significance of caterpillar olfaction in tri-trophic interactions of plants, herbivores, and their natural enemies has remained largely underexplored. In this study, we severely impaired the olfaction of P. brassicae by knocking out Orco using CRISPR/Cas9 to investigate the ecological importance of olfaction in host-plant choice and enemy avoidance by P. brassicae larvae. The KO of olfaction was successful, as we did not find Orco proteins in the antennae of Orco KO caterpillars and showed an altered brain structure in the Orco KO caterpillars in comparison to WT caterpillars. Furthermore, the near-complete lack of response to odors by Orco KO butterfly antennae compared to WT confirms that olfaction was largely lost. Intriguingly, although mating and oviposition were significantly reduced, KO butterflies still produce some homozygous offspring. After this characterization of our mutant line, we aimed to test the role of larval olfaction in a multitrophic framework. We found that the loss of olfaction had a significant effect on the foraging behavior of the caterpillars, which exhibited a reduced growth rate and a greater vulnerability to the parasitoid wasp C. glomerata.
The perception of chemical cues is mediated by the chemosensory signal transduction in insect antennae and further processing of the information in the brain. In P. brassicae KO caterpillar antennae, no Orco protein was detected, and chemical signals can therefore not be converted into electrical signals (Sato et al., 2008; Wicher et al., 2008). Interestingly, the non-functional Orco decreased the number of glomeruli in the LAC, indicating that Orco is likely to play a role in the glomerular organization of the LAC (Figure 1C–I). Similar changes have been reported for adult insects of a few hymenopteran species: the ants Harpegnathos saltator and Ooceraea biroi and the honeybee Apis mellifera, where the number and total size of glomeruli decreased while the volume of individual glomeruli increased in Orco KO mutants (Trible et al., 2017; Yan et al., 2017; Chen et al., 2021). In contrast, the antennal lobe morphology of D. melanogaster was not significantly changed when Orco was knocked out (Larsson et al., 2004). In Lepidoptera, the size and number of glomeruli in Helicoverpa armigera were found to be comparable between WT and KO adult moths without any significant difference (Fan et al., 2022). However, silencing the pheromone receptor, SlitOR5, led to a decreased size of a glomerulus in the adult moth Spodoptera littoralis, and knocking out Orco also led to a reduced size of OR-related glomeruli in adult Manduca sexta (Koutroumpa et al., 2022; Fandino et al., 2019). The loss of olfactory glomeruli in the caterpillar might, therefore, either indicate that the formation of glomeruli in the embryo is mediated by a different mechanism than during pupation (Oland and Tolbert, 2011), or that the loss of neuronal activity reduces axonal innervations and thereby the size of the glomeruli to an extent that they could no longer be visualized by our method. However, the mechanism of glomerular formation and the roles that olfactory genes play in this process need to be further investigated in both immature and adult insects (Williams et al., 2022).
The role of olfaction in the behavioral ecology of adult insects has already been studied in quite some detail, and silencing Orco has been an important tool for unraveling the importance of chemical communication in adults of various insect species. For instance, in Orco KO adults of several insect species, there was a significant reduction in the electrophysiological responses to volatile chemicals such as alcohols and esters (Fandino et al., 2019; Sun et al., 2020). Consequently, Orco mutants exhibited notable changes in host-plant location and severely reduced fecundity due to impaired pheromone perception and mating, which also resulted in significantly reduced egg-laying behavior (Fan et al., 2022; Fandino et al., 2019; Liu et al., 2023). Similar to other species, Orco KO P. brassicae butterflies exhibited a loss of electrophysiological responses to most plant volatiles we tested (Fan et al., 2022; Sun et al., 2020; Figure 1J, K), and the mating behavior of P. brassicae Orco KO butterflies was also disrupted (Figure 1—figure supplement 3). However, we observed some differences: P. brassicae Orco KO butterflies were still able to mate to a limited degree (Figure 1—figure supplement 2). Whereas Orco KO adults in some other insect species did not exhibit any mating behavior (Fandino et al., 2019; Fan et al., 2022). This finding suggests that additional sensory systems beyond olfaction, such as vision, may be involved in the mating processes of this day-active butterfly (Carpenter and Sparks, 1982; Obara et al., 2008). Interestingly, while many P. brassicae butterflies with impaired olfaction failed to mate, several KO butterflies still mated and deposited fertilized eggs when the oviposition substrate was supplied in our laboratory setting. The mated Orco KO butterflies laid a comparable number of fertilized eggs on cabbage plants to WT butterflies, which is in line with the results from different moth species (Fandino et al., 2019; Fan et al., 2022; Sun et al., 2023), suggesting oviposition decisions are mostly guided by non-volatile compounds when no host-plant location is required (van Loon et al., 1992a). Furthermore, the hatching rate of eggs laid by mated Orco KO butterflies was similar to that of WT butterflies (Figure 1—figure supplement 2), indicating that the reduced fecundity in these insects is primarily due to impaired mating behavior and reduced mating frequency, which may result from a lack of pheromonal communication.
When directly placed on their host plant, we found that WT caterpillars gained more weight than Orco KO caterpillars (Figure 2A). We speculate that olfaction is needed to evaluate plant tissues and to select different leaves or leaf parts in order to feed efficiently, even when the caterpillar is already on the plant. Secondary metabolites are often unevenly distributed even within a single leaf, and these concentration differences have been shown to influence the performance of different caterpillars (Kester et al., 2002; Yuan et al., 2022). It is conceivable that these localized differences in plant secondary metabolites are also detectable by the herbivore through its olfactory system (Hanson and Dethier, 1973). The difference between WT and KO caterpillars was amplified in the presence of disarmed parasitoids, which were able to attack but not injure the caterpillars, indicating that KO caterpillars were more susceptible to harassment by their natural enemies (Figure 2E). We hypothesize that this further reduction in caterpillar weight was due to the less efficient foraging behavior of the KO caterpillars, which resulted in an extended time period in which the caterpillars were vulnerable to the harassment by the disarmed parasitoids. This harassment would then further reduce the foraging efficiency of the KO caterpillars and thereby exacerbate the effect seen in Figure 2A.
P. brassicae caterpillars are most vulnerable to C. glomerata during the first two larval instars. Due to their reduced growth rate, KO caterpillars spent a longer time in these vulnerable stages and, therefore, in a natural setting would likely have suffered more strongly from parasitoid attack than WT caterpillars (Brodeur et al., 1996; Haverkamp and Smid, 2020). In addition, caterpillars might have been avoiding plant parts with natural enemies similar to choosing infested plants without parasitoids over infested plants with parasitoids present (Figure 2D). This suggests that caterpillars might be able to use their olfactory system to find enemy-free spaces based on volatiles emitted by the plant or the frass and spit of conspecifics. Similarly, when the caterpillars were exposed to unmanipulated parasitoids, the KO caterpillars were more susceptible and exhibited a higher mortality rate, demonstrating again that olfaction is crucial in larvae to survive under the pressure of natural enemies (Figure 2F). Subsequently, we inferred that the differences in weight gain and survival rate between WT and KO caterpillars are due to different sensitivity to plant volatiles. WT caterpillars can sooner reach a body size in which they are no longer vulnerable to attack by C. glomerata by locating an available food source efficiently (Brodeur et al., 1996). The higher mortality rate of KO caterpillars might, therefore, be partly due to a reduced foraging efficiency caused by the lack of olfactory information, which in turn led to an extended period in which the caterpillars were vulnerable to attack by their natural enemies. In addition, the WT caterpillars might have been better able to use olfactory cues to avoid plant parts where conspecifics are under attack by parasitoids, again reducing their mortality in comparison to KO caterpillars (Figure 2D).
Searching for host plants suitable for feeding and free of natural enemies is largely accomplished via the olfactory system of herbivorous insects (Carrasco et al., 2015; Hu et al., 2020). Without functional ORs, P. brassicae caterpillars could still discriminate plant tissues from paper discs; however, Orco KO caterpillars preferred the non-host tomato plant over their natural food plant (Figure 2B). We surmise that OR-mediated olfaction is required in caterpillars to locate and identify their host plants, and losing this part of their olfactory receptor repertoire would make it challenging to distinguish host from non-host plants (Liu et al., 2023). Nonetheless, Orco KO caterpillars still retain ORNs expressing ionotropic receptors, which commonly detect acids and amines that have been found to elicit aversion in insect herbivores (Zhang et al., 2019). Fatty acids are present at the edge of both artificially damaged and caterpillar-infested cabbage leaves (Horikoshi et al., 1997), which, in the absence of ORs detecting attractive plant compounds, may have caused avoidance behavior in the KO caterpillar to their natural host plant. Interestingly, the parasitoid C. glomerata also uses the fatty acids produced by the plant to locate Pieris caterpillars, and it has been argued that herbivores might avoid these compounds to escape from their natural enemies (Horikoshi et al., 1997; Zhang et al., 2019).
We found that WT caterpillars prefer caterpillar-infested plants free of wasps over caterpillar-infested plants with wasps, which suggests that caterpillars are able to detect certain volatiles that are derived from the interaction with caterpillars and wasps. By analyzing headspace samples of caterpillars, parasitoids, as well as the interaction of caterpillars and parasitoids, we successfully identified some chemical compounds which were already known plant volatiles, such as alcohols and aldehydes. However, we did not find any compounds that were specifically abundant in Pb-Cg or Cg. This suggests that a volatile signal serving as a ‘danger signal’ directly emitted by the wasps or caterpillars might not exist, or it might not be detectable with our methods (Figure 3B). Alternatively, indirect cues such as the volatiles emitted from the caterpillar spit during their defensive behavior might have triggered the avoidance behavior of the WT caterpillars to infested plants with natural enemies. When we tested the most abundant compounds in a two-choice behavioral assay against a solvent control, we found that P. brassicae WT larvae showed a preference to the plant volatiles (Z)-3-hexen-1-ol and (E)-2-hexenal (Figure 3D), which are known HIPVs. Previous studies on other species have demonstrated that caterpillars are capable of detecting different HIPVs (Di et al., 2017; de Fouchier et al., 2018). For caterpillars, HIPVs can serve as indicators for the location of potential food plants and the presence of conspecifics (Di et al., 2017; Zhang et al., 2019), but they also attract natural enemies and enhance the risk of being attacked (Bernays, 1997; Ngumbi and Fadamiro, 2012; Yang et al., 2016). Most HIPVs identified in this study were found abundantly in caterpillar saliva that caterpillars spit onto parasitoids to defend against attack. In our study, caterpillars were attracted by HIPVs emitted by plants and by individual HIPVs (Figure 3D). We speculate that caterpillars use low to intermediate concentrations of HIPVs to find suitable host plants and high concentrations to avoid sites where other caterpillars are under attack by their natural enemies. Therefore, perceiving plant leaf volatiles and approaching the food plants accordingly will increase the herbivores’ feeding efficiency and enhance survival chances. Interestingly, ITCs, which are hydrolyzed from glucosinolates after leaf damage in crucifer plants, did not elicit strong responses from the butterfly antenna (Figure 1J, K). Because all receptors expressed in the caterpillar are also present in the butterfly and because of the limited number of ORs and ORNs in larvae and low expression levels of ORs (Wang et al., 2024), we found it less likely that ITCs are used by P. brassicae caterpillars as foraging cues, even though these compounds are recognized by Plutella xylostella ORs (Liu et al., 2020). In our study, volatile cues of wasps were only found in very low quantities, and these compounds hardly elicited any behavioral response in the caterpillars; however, these chemicals might still play a role in the interaction between the caterpillars and wasps at a close range (Ebrahim et al., 2015).
The way that larval insects react to the potential threat from their natural enemies is still largely unknown for most insects. Our study that exploited CRISPR/Cas9 shows that olfaction is of high significance to caterpillars for locating food sources and likely affects their survival when under the selection pressure from natural enemies. We provided novel insights into the tri-trophic interactions of plants, herbivores, and their natural enemies from the perspective of a caterpillar and highlighted the role of olfaction for both foraging and escaping natural enemies.
Materials and methods
Plant and insect rearing
Request a detailed protocolCabbage plants (B. oleracea var. gemmifera cv. Cobelius; Brussels sprouts) and tomato plants (Solanum lycopersicum cv. moneymaker) were planted in individual pots and grown for 4 weeks after germination before being used in the experiments or to feed the caterpillars. The cabbage plants were reared under glasshouse conditions at 22 ± 3°C, RH 50–80% and 16 hr light:8 hr dark cycles. P. brassicae caterpillars and butterflies, as well as C. glomerata wasps, were collected from laboratory colonies maintained at the Laboratory of Entomology, Wageningen University & Research, The Netherlands. The caterpillars were reared on Brussels sprouts plants at 22 ± 3°C, RH 25–35% with 14 hr light:10 hr dark cycles. The adults were supplied with 10% sugar water as nutrient under the same environmental conditions. After successful gene editing, KO insects were transferred and reared in a glass house compartment under conditions at 25 ± 3°C, RH 25–35%. Several KO caterpillars and butterflies from each generation were randomly collected for genotype screening to ensure that homozygosity remains. Cotesia glomerata adults were fed with organic honey, P. brassicae second-instar larvae (L2) were supplied to the parasitoid wasps for parasitization to maintain the colony. The wasp cocoons were collected in Petri dishes and incubated at 22°C, RH 50–70% with the same photoperiod as that of caterpillars. All the subsequent comparisons between WT and KO in this study were conducted in the glass house compartment under the rearing condition of KO insects unless specified.
Orco KO
Request a detailed protocolThe second exon of Orco was targeted to knockout the gene. The sgRNA was designed with Geneious Prime 2021 (Geneious, New Zealand) by searching N20 target + NGG PAM sequences in the second exon, sgRNA primers were selected according to activity scorings (Doench et al., 2016). The designated sgRNA off-target regions were evaluated by searching against the genome by est2genome function with a loose 50-score cut-off by using Exonerate 2.0 (Slater and Birney, 2005) and CHOPCHOP v3 (Labun et al., 2019). The sgRNAs that had no off-target sites in coding areas were selected. sgRNA templates were prepared by mixtures of 50 µl Q5 high-fidelity 2× master mix (NEB, USA), 5 µl of the following forward primers: sgRNA1: ATTTAGGTGACACTATACATGTCAACTCTACTTAGGAGTTTTAGAGCTAGAAATAGCAAG; sgRNA2: ATTTAGGTGACACTATACGATGAAGTAAACGAACTCAGTTTTAGAGCTAGAAATAGCAAG; 5 µl constant reverse primer: AAAAGCACCGACTCGGTGCCACTTTTTCAAGTTGATAACGGACTAGCCTTATTTTAACTTGCTATTTCTAGCTCTAAAAC and 40 µl H2O. The mixture was incubated at 98°C for 2 min, followed by 38 cycles of 98°C for 20 s, 65°C for 10 s, and 72°C for 10 s, finalized at 72°C for 5 min. The PCR products were checked by 1.0% gel electrophoresis and purified by QIAGEN PCR purification kit (QIAGEN, Germany) according to the manufacturer’s instructions. sgRNA was synthesized by a reaction system of 2 µl 10× SP6 enzyme mix (Invitrogen, USA), 2 µl 10× SP6 reaction buffer, 2 µl each of ATP, UTP, GTP, and CTP, 200 µg sgRNA template and add RNase-free H2O up to 20 µl. The mixture was incubated at 37°C for 4 hr, followed by adding 1 µl Turbo DNase and incubating at 37°C for 15 min. The reaction products were purified by Monarch RNA cleanup kit (NEB, USA). The concentration of sgRNA was determined by DeNovix (DeNovix, USA). sgRNA was mixed with Cas9 protein (NEB, USA) and incubated at 25°C for 10 min.
Newly laid eggs, not older than half an hour, were collected from our laboratory colony and were fixed on glass slides. The sgRNA/Cas9 mixture was colored with 1 µl food dye to stain the eggs as an indicator that injection with a Femto-jet (Eppendorf, Germany) was successful. The injected eggs were incubated at 25°C for 4–5 days until hatching. The caterpillars were reared until the emergence of the butterflies, a leg of each butterfly was cut for genotyping. gDNA of the leg samples was extracted with MyTaq Extract-PCR Kit (Bioline, Germany). Mutant screening was performed by PCR with 15 µl 2× MyTaq HS Red mix, 2 µl forward primer (TCTGGCTTCGGTATTACATTTC), 2 µl reverse primer (CTTTTATGGCGTGTTTTATTTG), 2 µl gDNA template, and 9 µl H2O. The PCR products were checked by gel electrophoresis and sequenced with the same primers by Eurofins (Eurofins, The Netherlands).
ORN staining and axon tracing
Request a detailed protocolL3 caterpillars (n = 6 for WT and n = 5 for KO) were anesthetized on ice and decapitated. Heads were cut along the midline with micro-scissors to facilitate penetration of the fixative and antibodies. The collected heads were immersed in freshly prepared 4% formaldehyde in 0.1 M phosphate buffer at pH 7.3 and fixed overnight at 4°C. The fixed samples were rinsed in 70% ethanol, followed by incubation in a mixture of 96% ethanol: 30% hydrogen peroxide at 1:1 ratio for 7 days at 4°C. The incubated samples were then dehydrated in a graded series of ethanol, degreased in xylene for 2 min, rehydrated, and incubated in 0.1 M phosphate buffered saline (Oxoid, Dulbecco A) with 0.5% Triton X-100 (PBS-T) followed by four washes in PBS-T for 15 min each. Preincubation of head samples was performed in 10% normal goat serum (Bio-connect services, The Netherlands) in PBS-T (PBS-T-NGS) for 1 hr, and then incubated in rabbit anti-Orco antibody, the specificity of which was tested previously (Nolte et al., 2016), diluted in 1: 500 in PBS-T-NGS for 3 days at 4°C, followed by 6 washes in PBS-T for 30 min each. Samples were subsequently incubated in goat anti-rabbit conjugated to Alexa fluor 488 (Thermo Fisher Scientific, USA) diluted 1:100 and TO-PRO-3 iodide (Thermo Fisher Scientific, USA) diluted 1:1000 in PBS-T-NGS for 2 days at 4°C. Heads were washed two times in PBS-T for 30 min and once overnight at 4°C, following with two 1 hr PBS washes at room temperature. The samples were then dehydrated with a graded series of ethanol, cleared in xylene, and mounted in DPX. Confocal microscopy was performed with a Leica Stellaris 5- DIM 8 confocal microscope (Leica, Germany), using a 63× oil-immersion plan APO objective NA1.4 and a spectrally flexible white light laser for excitation of the two fluorophores using the pre-sets for Alexa fluor 488 and To-Pro-3 iodide.
L4 caterpillars of similar head size were anesthetized on ice and immobilized in clay. The tip of each antenna was peeled, and a layer of petroleum jelly (Vaseline) was applied around the antenna. A 0.5-µl drop of 2.5% biotin–dextran solution was added to the cut and covered with petroleum jelly, then incubated at 4°C overnight. Brains were dissected in PBS, fixed in 4% formaldehyde in PBS at 4°C overnight, dehydrated in xylene, and rehydrated in PBS-T, followed by three washes in PBS-T for 2 hr each. Brains were then incubated in 10% NGS in PBS-T for 1 hr, followed by incubation in 1:200 streptavidin 488 and 1:1000 To-Pro iodide in PBS-T at 4°C for 3 days. After further rinsing in PBS-T overnight, brains were mounted in DPX. Whole-mount samples were scanned using confocal microscopy with settings similar to those used for ORN staining.
Electroantennography
Request a detailed protocolA panel of 54 chemicals (Supplementary file 1G) including esters, alcohols, aldehydes, alkenes, ITCs, heterocyclics, and nitriles was selected from the literature (van Loon et al., 1992b; Liu et al., 2020; Zhu et al., 2015; Bourne et al., 2023) and our current volatile identification data, to investigate the EAG responses of both KO and WT butterfly antennae (n = 15–18) of both sexes. Filter papers loaded with 10 µl of selected chemicals, diluted in paraffin oil (10–2 vol/vol), were inserted into Pasteur pipettes, and clean filter paper loaded with 10 µl paraffin oil was employed as negative control. The left antennae of butterflies were excised at the basal end, and the distal tip was also removed for better conductivity. Antennae were placed into glass capillaries filled with EAG Ringer solution. Glass capillaries were placed on Ag-AgCl wires connected to a ground electrode and a 10× high-impedance DC amplifier (Ockenfels Syntech, Germany). The electrical signals were transformed using an IDAC-232 (Ockenfels Syntech, Germany) analog-digital converter connected to a personal computer. Signals were finally recorded using the software EAG Pro (Ockenfels Syntech, Germany). The first puff of each chemical was abandoned before the first round of testing. Antennae were exposed to the chemicals in random order and were given intervals of at least 20 s for recovering until the baseline stabilized.
Female oviposition behavior
Request a detailed protocolA mating pair of newly emerged butterflies (n = 11 for WT and n = 14 for KO) were put in a cage and supplied with a 4-week-old Brussels sprouts cabbage plant as oviposition substrate and sugar water as nutrients. The numbers of eggs were counted on a daily basis for 7 days. The female butterflies were then collected and dissected to compare mating frequency between WT and KO females. Mating frequency was determined by the number of spermatophores dissected from the females’ abdomens. The eggs remained on the plants for another 4–5 days to allow hatching. The number of caterpillars hatched enabled us to compare the number of fertilized eggs and hatching rate between the two butterfly genotypes.
Caterpillar performance on plants
Request a detailed protocolCaterpillar performance on cabbage plants was evaluated by three different treatments:
For each genotype, a group of ten L1 caterpillars (n = 17) was placed on one leaf of a 4-week-old cabbage plant. Caterpillars were reared for 10 days, and more plants were provided when needed. The caterpillars were then collected for weighing to compare the growth between the two genotypes.
The growth of both genotypes of caterpillars (n = 8) was further compared by rearing in the presence of C. glomerata parasitoid wasps from which the ovipositor had been removed (hereafter: ‘disarmed’). Groups of ten L1 caterpillars were placed on individual 4-week-old cabbage plants as described above. The parasitoid wasps were anesthetized on a CO2 plate, and the ovipositor was removed with fine tweezers to ensure that the parasitoid wasps can interact with the caterpillars but cannot parasitize them. The activity of disarmed parasitoids was observed for several minutes to ensure that they would behave similarly to unmanipulated parasitoids. Two disarmed parasitoid wasps were released in each rearing cage 24 hr after the caterpillars had been placed on the plants. The disarmed parasitoid wasps were replaced by newly disarmed parasitoids every day, the caterpillars were reared on the plants for 10 days and then collected for weighing.
To compare the survival rate of caterpillars (n = 7 groups of ten caterpillars) on the plants under the threat of C. glomerata parasitoid wasps, a group of ten L1 caterpillars was placed on a leaf of a 4-week-old cabbage plant 2 days prior to placing two female parasitoid wasps. The parasitoid wasps were replaced every day to ensure that caterpillars were always exposed to natural enemies. The caterpillars were collected after 10 days to count the number that survived; the collected caterpillars were further reared until they pupated or C. glomerata larvae emerged.
Larval host-plant seeking behavior
Request a detailed protocolCaterpillar (n = 54–73) food-plant seeking behavior was evaluated in a greenhouse under the same conditions in which the caterpillars were reared. Plant leaf discs or green paper discs (Trophée 1224 ‘Forest Green’, Clairefontaine, France), with 12 mm diameter were placed at opposite positions along the edge of a 188-mm-diameter Petri dish. A single L3 caterpillar was then placed at the center of the arena. Each caterpillar was observed for 10 min, the first choice that the caterpillars made and the time spent were recorded. The caterpillars were provided with a choice between either (1) a cabbage leaf disc and a paper disc, (2) a tomato leaf disc and a paper disc, or (3) a cabbage leaf disc and a tomato leaf disc. A choice was recorded when caterpillars began feeding on the cabbage leaf disc or contacted the tomato leaf/green paper discs. New discs were used for every caterpillar, and the positions of discs were exchanged after every ten tests.
To further test the host-plant locating behavior of L3 caterpillars, we investigated the behaviors of caterpillars in a Y-tube olfactometer (diameter 35 mm) with a built-in black Y-shaped wire, allowing caterpillars to crawl along the wire and make a choice smoothly in the laboratory. Each arm was provided with a 1.1 l/min inlet airflow which had been purified by charcoal and humidified by water before entering the treatment jars. Healthy plants were untreated, infested plants were infested with 30 early L3 caterpillars for 24 hr and with caterpillars on plants while testing. Differently treated plants were placed in jars to test caterpillar behaviors. The caterpillars had been starved for approximately 5 hr before testing. Each caterpillar was tested and observed for 5 min, and caterpillar behavior was monitored by a camera. The position of inlet airflow was changed every five tests to avoid position bias.
Volatile analysis
Request a detailed protocolVolatiles were collected from L3 P. brassicae caterpillars, 3-day-old female C. glomerata wasps, P. brassicae caterpillars and C. glomerata wasps together, caterpillar frass collected from L3 caterpillars and caterpillar saliva also collected from L3 caterpillars using glass capillaries following Mattiacci et al., 1994. Fresh frass and saliva samples were stored in a freezer at –20°C and thawed overnight at room temperature before volatile collection. Headspace collection of volatiles was performed from five treatments: 20 female wasps (Cg) in a 10-ml glass vial (n = 12); 20 L3 caterpillars (Pb) in a 10-ml glass vial (n = 10); 20 L3 caterpillars and 2 female wasps (Pb–Cg) in a 10-ml glass vial (n = 14); 100 mg caterpillar frass (Pb-Fr) in a 1.5-ml glass vial (n = 10); and 50-µl caterpillar saliva (Pb-S) in a 1.5-ml glass vial (n = 12). Clean empty glass vials (10 and 1.5 ml) were used as a negative control. Volatiles were collected through dynamic headspace sampling using Tenax TA adsorbent material (20/35 mesh; Camsco, USA). Synthetic air (Air Synthetic 4.0 Monitoring from Linde Gas, The Netherlands) at 110 ml min–1 to the samples in 10 ml vials and 55 ml min–1 to the samples in 1.5 ml vials were constantly supplied as a carrier for the volatiles, while simultaneously the volatiles were trapped by drawing air at 100 and 50 ml min–1, respectively, through a stainless-steel tube filled with 200 mg Tenax TA for 2 hr.
The collected volatiles were thermally released from the Tenax TA adsorbent using an Ultra 50:50 thermal desorption unit (Markes, UK) at 250°C for 10 min under a helium flow of 20 ml min–1, while simultaneously re-collecting the volatiles in a thermally cooled universal solvent trap: Unity (Markes, UK) at 0°C. Once the desorption process was completed, the volatile compounds were released from the cold trap by ballistic heating at 40°C s–1 to 280°C, which was then kept for 10 min, while all the volatiles were transferred to a ZB-5MS analytical column 30 ml × 0.25 mm ID × 1 mm F.T. with a 10 m built-in guard column (Phenomenex, USA), placed inside the oven of a Thermo Trace GC Ultra (Thermo Fisher Scientific, USA), for separation of volatiles. The GC oven temperature was initially held at 40°C for 2 min and was immediately raised at 6°C min–1 to a final temperature of 280°C, where it was kept for 4 min under a constant helium flow of 1 ml min–1. For the detection of volatiles, a Thermo Trace DSQ quadrupole mass spectrometer (Thermo Fisher Scientific, USA) coupled to the GC was operated in an electron impact ionization mode at 70 eV in a fullscan mode with a mass range of 35–400 amu at 4.70 scans s–1. The MS transfer line and ion source were set at 275 and 250°C, respectively.
Automated baseline correction, peak selection (S/N >3), and alignments of all extracted mass signals of the raw data were processed following an untargeted metabolomic workflow using MetAlign software, producing detailed information on the relative abundance of mass signals representing the available metabolites (Lommen, 2009). This was followed by the reconstruction of the extracted mass features into potential compounds using the MSClust software through data reduction by means of unsupervised clustering and extraction of putative metabolite mass spectra (Tikunov et al., 2012). Tentative identification of volatile metabolites was based on a comparison of the reconstructed mass spectra with those in the NIST 2008 and Wageningen Mass Spectral Database of Natural Products MS libraries, as well as experimentally obtained linear retention indices.
Caterpillar response to individual chemical compounds
Request a detailed protocolL3 caterpillars (n = 31–50 for each compound, each genotype) were tested in a custom-made set-up consisting of 25 separate arenas in the laboratory (Figure 3C). Each arena allowed caterpillars to move freely along two channels of 90 mm located in opposite directions of a central opening, which permitted the caterpillar to crawl into the arena. At the end of the arenas, airflows are introduced to deliver volatile chemicals. The airflow was sucked out through the channel below the central opening simultaneously. Single caterpillars were placed in the central well of the individual arenas before the test. Caterpillars were given a choice between a selected volatile chemical of interest (Supplementary file 1H) and paraffin oil. Input odors were delivered at 1.1 l min–1 for both chemicals and solvent controls, and the odors were sucked out at 2.2 l min–1 at the same time from the center. The arena was cleaned by ethanol and dried by high-pressure air. The position of chemicals was exchanged after each test. The behavior of caterpillars was recorded for 10 min at 100 frames per minute using a Canon TV lens JF16mm (Canon, Japan). Behavioral data were extracted by EthoVision XT 11.5 software (Noldus, The Netherlands). Data were removed from the dataset when the caterpillar stayed in the arena for less than 10 s. All tests were performed at room temperature.
Statistical analysis
Request a detailed protocolUnless stated otherwise, statistical tests were performed in R (version 4.4.0; R Core Team, 2016) in combination with RStudio (Posit, USA). Datasets were tested for normal distribution using a Shapiro–Wilk test and for equal variances using Levene’s test. The difference of glomerular number between WT and Orco KO caterpillars was tested with a Wilcoxon rank-sum test. Electroantennographic responses between the two genotypes were tested statistically by using the Student’s t-test when the values were normally distributed and had homogenous variance or tested by using the Kruskal–Wallis rank-sum test when the requirements for the t-test were not met. The difference in butterfly mating frequency was analyzed by using the Wilcoxon rank-sum test; differences in oviposition dynamics were tested with a GLM with a negative binomial distribution. The development differences between WT and KO caterpillars on full plants and under the thread of disarmed parasitoids were tested using a Student’s t-test, and survival rate difference between the two genotypes was tested using a GLM with a beta-binomial distribution. Caterpillar host-plant selection behaviors in Petri dishes and in the Y-tube were analyzed with a Chi-square test. Differences in the relative time spent by the caterpillars between the control side and the odor side of the multichannel arenas were compared by a non-parametric Wilcoxon rank-sum test. PCA of volatile blends was achieved by MetaboAnalyst (Pang et al., 2024). Data were normalized by logarithmic transformation and scaled in auto mode. Ward’s method and Euclidean distance were employed in the hierarchical clustering heatmap by default settings without clustering samples. In addition, the abundance of volatiles as peak heights was imported into SIMCA-P 17 statistical software (Umetrics, Sweden), and the data were analyzed by MVDA. Supervised Orthogonal Projection to Latent Structures Discriminant Analysis (OPLS-DA) was employed to compare and correlate treatment groups.
Data availability
All data generated during this study and the corresponding scripts for the statistical analysis are available in the supplementary files and under DOI: https://doi.org/10.4121/610e707d-b10a-467e-8afe-ead477827914.v1.
-
4TU.ResearchDataData underlying the publication "Loss of olfaction reduces caterpillar performance and increases susceptibility to a natural enemy".https://doi.org/10.4121/610e707d-b10a-467e-8afe-ead477827914.v1
References
-
Feeding by lepidopteran larvae is dangerousEcological Entomology 22:121–123.https://doi.org/10.1046/j.1365-2311.1997.00042.x
-
Effects of vision on mating behavior of the male corn earwormJournal of Economic Entomology 75:248–250.https://doi.org/10.1093/jee/75.2.248
-
Insect host plant selection in complex environmentsCurrent Opinion in Insect Science 8:1–7.https://doi.org/10.1016/j.cois.2015.01.014
-
Behavioral effect of plant volatiles binding to Spodoptera littoralis larval odorant receptorsFrontiers in Behavioral Neuroscience 12:264.https://doi.org/10.3389/fnbeh.2018.00264
-
Design of larval chemical attractants based on odorant response spectra of odorant receptors in the cotton bollwormInsect Biochemistry and Molecular Biology 84:48–62.https://doi.org/10.1016/j.ibmb.2017.03.007
-
Chemical detection of natural enemies by arthropods: an ecological perspectiveAnnual Review of Ecology and Systematics 32:1–23.https://doi.org/10.1146/annurev.ecolsys.32.081501.113951
-
The evolutionary context for herbivore-induced plant volatiles: beyond the “cry for help”Trends in Plant Science 15:167–175.https://doi.org/10.1016/j.tplants.2009.12.002
-
Optimized sgRNA design to maximize activity and minimize off-target effects of CRISPR-Cas9Nature Biotechnology 34:184–191.https://doi.org/10.1038/nbt.3437
-
Olfactory preference for egg laying on citrus substrates in DrosophilaCurrent Biology 23:2472–2480.https://doi.org/10.1016/j.cub.2013.10.047
-
BookLarge White Butterfly: The Biology, Biochemistry and Physiology of Pieris BrassicaeDr W. Junk Publishers, The Hague.https://doi.org/10.1007/978-94-009-8638-1_10
-
Rôle of gustation and olfaction in food plant discrimination in the tobacco hornworm, Manduca sextaJournal of Insect Physiology 19:1019–1034.https://doi.org/10.1016/0022-1910(73)90028-0
-
Effect of different cole crops on the biological parameters of Pieris brassicae (L.) (Lepidoptera: Pieridae) under laboratory conditionsJournal of Crop Science and Biotechnology 13:195–202.https://doi.org/10.1007/s12892-010-0025-2
-
A neuronal arms race: the role of learning in parasitoid-host interactionsCurrent Opinion in Insect Science 42:47–54.https://doi.org/10.1016/j.cois.2020.09.003
-
Cotesia glomerata female wasps use fatty acids from plant–herbivore complex in host searchingJournal of Chemical Ecology 23:1505–1515.https://doi.org/10.1023/B:JOEC.0000006418.49537.5d
-
CHOPCHOP v3: expanding the CRISPR web toolbox beyond genome editingNucleic Acids Research 47:W171–W174.https://doi.org/10.1093/nar/gkz365
-
The molecular basis of host selection in a crucifer-specialized mothCurrent Biology 30:4476–4482.https://doi.org/10.1016/j.cub.2020.08.047
-
Species and sexual differences in behavioural responses of a specialist and generalist parasitoid species to host-related volatilesBulletin of Entomological Research 102:710–718.https://doi.org/10.1017/S0007485312000326
-
Better mate in the shade: enhancement of male mating behaviour in the cabbage butterfly, Pieris rapae crucivora, in a UV-rich environmentThe Journal of Experimental Biology 211:3698–3702.https://doi.org/10.1242/jeb.021980
-
Plant interactions with multiple insect herbivores: from community to genesAnnual Review of Plant Biology 65:689–713.https://doi.org/10.1146/annurev-arplant-050213-035937
-
Mutagenesis of the orco odorant receptor co-receptor impairs olfactory function in the malaria vector Anopheles coluzziiInsect Biochemistry and Molecular Biology 127:103497.https://doi.org/10.1016/j.ibmb.2020.103497
-
Knockout of the odorant receptor co-receptor, orco, impairs feeding, mating and egg-laying behavior in the fall armyworm Spodoptera frugiperdaInsect Biochemistry and Molecular Biology 152:103889.https://doi.org/10.1016/j.ibmb.2022.103889
-
Electroantennogram responses to plant volatiles in two species of Pieris butterfliesEntomologia Experimentalis et Applicata 62:253–260.https://doi.org/10.1111/j.1570-7458.1992.tb00665.x
-
Ecology of infochemical use by natural enemies in a tritrophic contextAnnual Review of Entomology 37:141–172.https://doi.org/10.1146/annurev.en.37.010192.001041
-
Sympatric Pieris butterfly species exhibit a high conservation of chemoreceptorsFrontiers in Cellular Neuroscience 17:1155405.https://doi.org/10.3389/fncel.2023.1155405
-
A unique sense of smell: development and evolution of a sexually dimorphic antennal lobe – a reviewEntomologia Experimentalis et Applicata 170:303–318.https://doi.org/10.1111/eea.13145
Article and author information
Author details
Berhane T Weldegergis

Alexander Haverkamp
Funding
China Scholarship Council (China Scholarship Council grant)
- Qi Wang
Nederlandse Organisatie voor Wetenschappelijk Onderzoek (016.Veni.192.116)
- Alexander Haverkamp
The funders had no role in study design, data collection, and interpretation, or the decision to submit the work for publication.
Acknowledgements
We appreciate Janneke Bloem for helping with genome editing, and the rearing team at the Laboratory of Entomology for providing insects for our experiments in this study. In addition, we would like to thank the Wageningen Light Microscopy Centre for their support in obtaining the confocal images.
Version history
- Sent for peer review:
- Preprint posted:
- Reviewed Preprint version 1:
- Reviewed Preprint version 2:
- Version of Record published:
Cite all versions
You can cite all versions using the DOI https://doi.org/10.7554/eLife.105585. This DOI represents all versions, and will always resolve to the latest one.
Copyright
© 2025, Wang et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 1,968
- views
-
- 219
- downloads
-
- 4
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 1
- citation for Reviewed Preprint v1 https://doi.org/10.7554/eLife.105585.1
-
- 1
- citation for Reviewed Preprint v2 https://doi.org/10.7554/eLife.105585.2
-
- 2
- citations for Version of Record https://doi.org/10.7554/eLife.105585.3
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Loss of olfaction reduces caterpillar performance and increases susceptibility to a natural enemy
eLife 14:RP105585.
https://doi.org/10.7554/eLife.105585.3




{kind=link}
{kind=link}
{kind=link}