Neocortical layer-5 tLTD relies on non-ionotropic presynaptic NMDA receptor signaling
- Centre for Research in Neuroscience, Brain Repair and Integrative Neuroscience Program, Departments of Neurology & Neurosurgery and Medicine, The Research Institute of the McGill University Health Centre, Montreal General Hospital, Canada
- Université Côte d’Azur, CNRS UMR7275, Institute of Molecular and Cellular Pharmacology, France
- Integrated Program in Neuroscience, McGill University, Canada
- Gerald Choa Neuroscience Institute & School of Biomedical Sciences, The Chinese University of Hong Kong, China
Figures
Figure 1
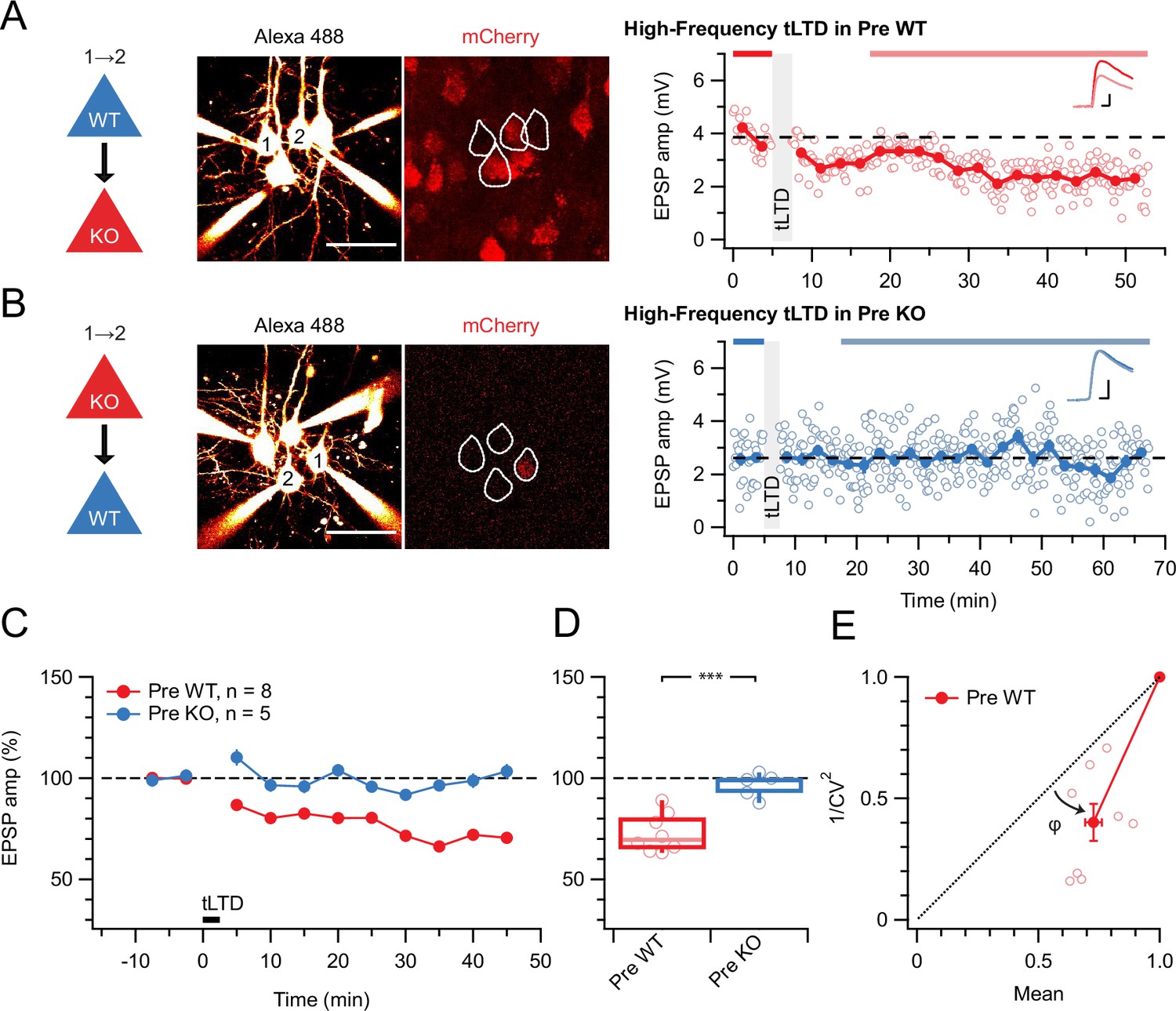
tLTD requires presynaptic NMDARs.
(A) Sample quadruple recording with L5 PCs visualized with Alexa 488 (left, scale bar 50 µm) and NMDAR deletion indicated by mCherry (right). In this wild-type (WT) → KO sample, HF tLTD at 20 Hz (see Methods) was intact (after/before = 68%, p<0.001), suggesting postNMDARs were not required. Traces (right) were averaged over periods indicated by blue lines. Scale bars: 5 ms, 1 mV. (B) For this KO→WT sample, however, the same induction elicited no plasticity (after/before = 100%, p=0.99), suggesting tLTD needed preNMDARs. Scale bars as in A. (C, D) HF tLTD was robustly evoked for presynaptic WT (Pre WT, blue) but not presynaptic KO pairs (Pre KO, red), verifying the need for preNMDARs in tLTD that we previously demonstrated (Sjöström et al., 2003; Sjöström et al., 2007). Pre WT: WT→WT and WT→KO pooled. Pre KO: KO→WT and KO→KO pooled. (E) CV analysis indicated that tLTD was presynaptically expressed (φ=19°±4°, n=8, one-sample t-test vs. diagonal, p0.001), in agreement with our prior studies (Sjöström et al., 2003; Sjöström et al., 2007).
Figure 2 with 1 supplement
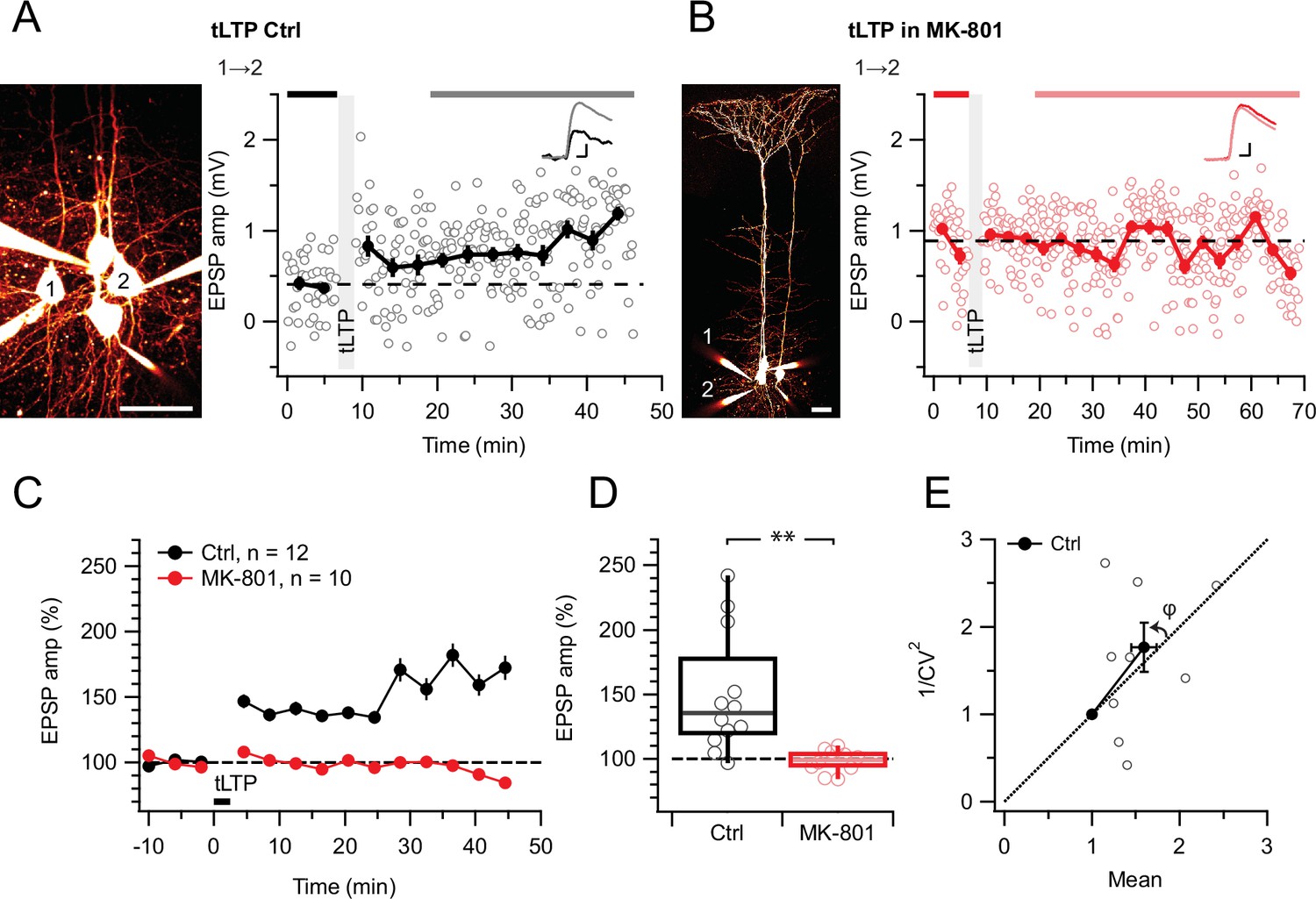
tLTP requires ionotropic NMDAR signaling.
(A) At this sample connection (PC 1→2, see Alexa 594 fills at left, scale bar: 50 µm), tLTP induction was successful (after/before = 210%, p<0.001). Traces (right) were averaged over time periods indicated in black/gray. To avoid MK-801 affecting short-term depression (Abrahamsson et al., 2017; Sjöström et al., 2003), baseline spiking was 0.1 Hz. Scale bars: 5ms, 0.2 mV. (B) In this sample, MK-801 abolished tLTP with the same induction protocol (after/before = 95.44%, p=0.55). Baseline spiking and scale bars as in (A). (C, D) Ensemble data revealed that tLTP was robustly expressed in controls (black) but abolished in MK-801 (red). (E) CV analysis indicated that tLTP was expressed both pre- and postsynaptically to varying degrees across different pairs (φ=11°±15°, n=10, p=0.44, two data points without potentiation were excluded, see Methods), in agreement with our prior work (Sjöström et al., 2007).
Figure 2—figure supplement 1
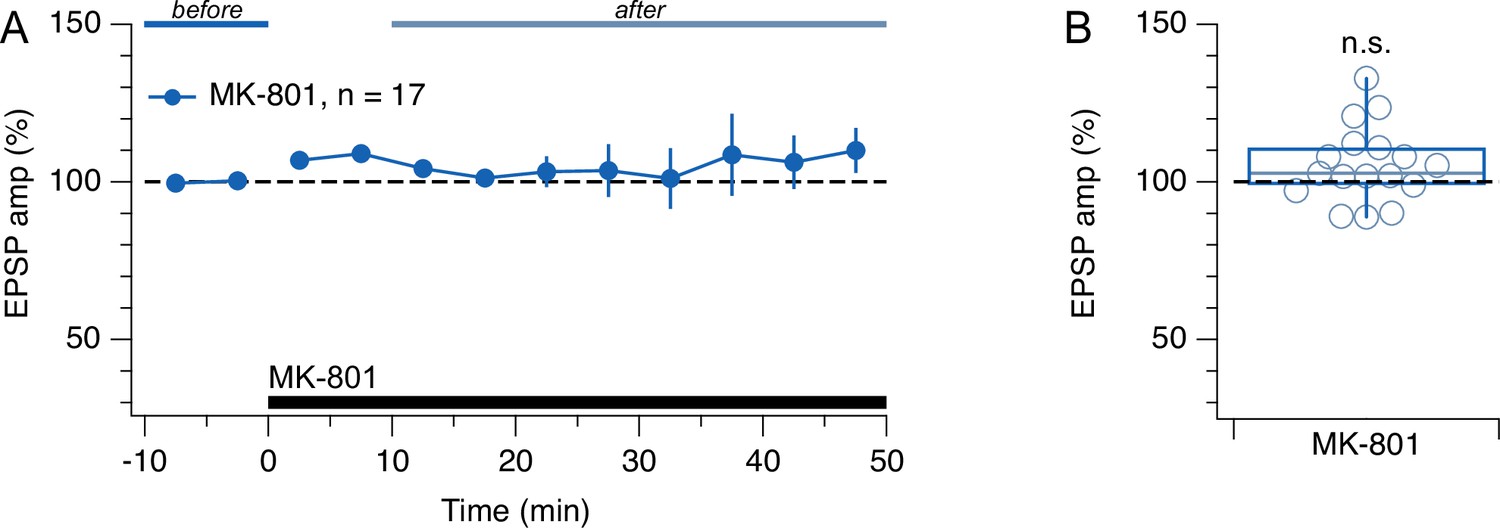
Low-frequency evoked release did not respond to MK-801.
(A, B) Washing in MK-801 had no appreciable effect on EPSP amplitude (one-sample t-test for normalized EPSP vs. 100%, p=0.076). Presynaptic cells spiked once every 10 s. This outcome is consistent with the finding that preNMDAR activation requires firing above a critical frequency (Abrahamsson et al., 2017; Buchanan et al., 2012; Sjöström et al., 2003; Wong et al., 2024). Before (blue) and after lines (light blue) indicate periods used for EPSP normalization.
Figure 3 with 1 supplement
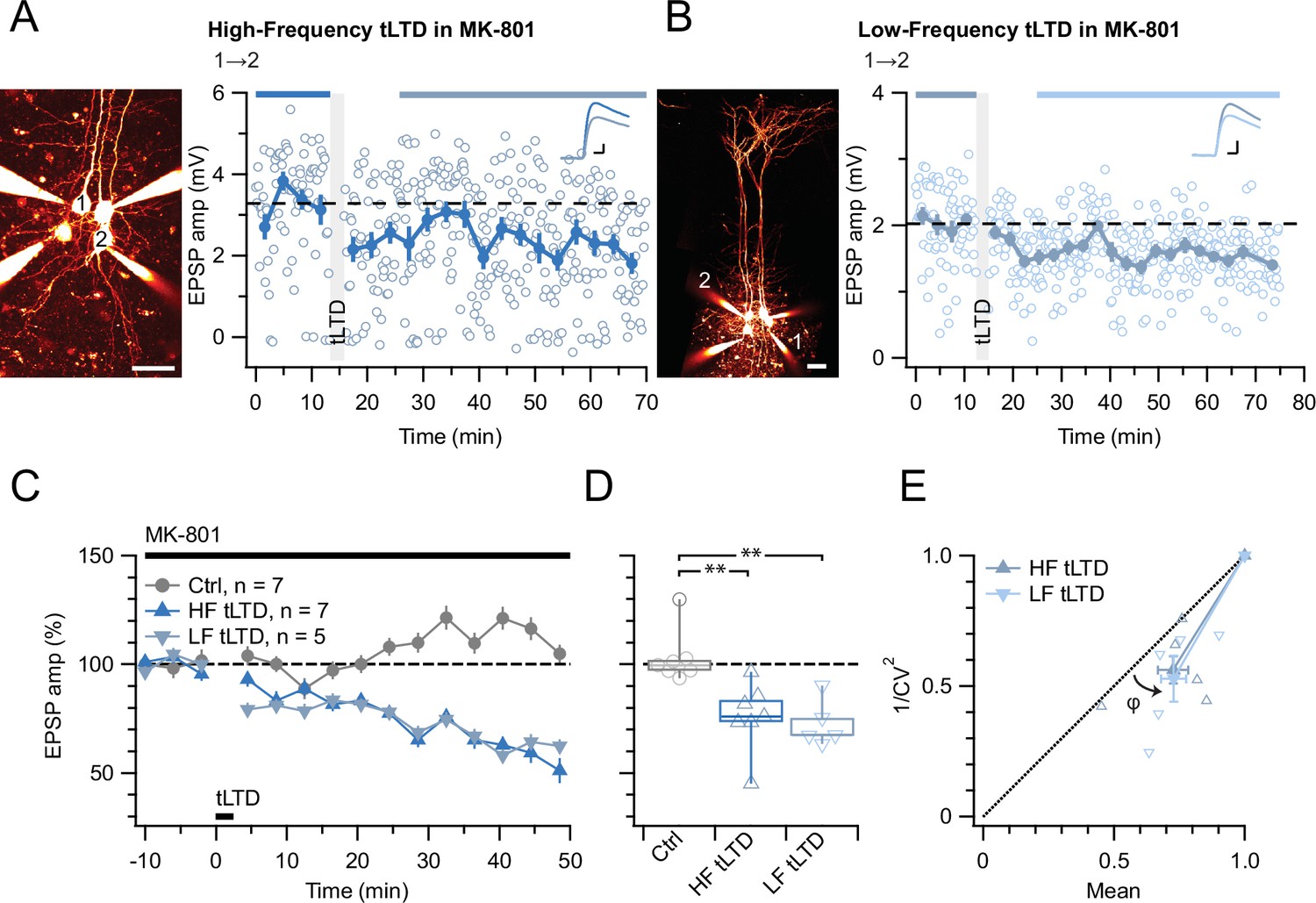
tLTD does not require ionotropic NMDAR signaling.
(A) At this sample connection (PC 1→2, see Alexa 594 fills left, scale bar: 50 µm), HF tLTD was successfully induced in the presence of MK-801 (after/before = 74%, p<0.001). Traces were averaged over time periods indicated in blue/light blue. Baseline spiking was 0.1 Hz. Scale bars: 5 ms, 0.5 mV. (B) In this sample, LF tLTD at 1 Hz (see Methods) was successful in MK-801 (after/before = 78%, p<0.001). Baseline spiking and scale bars as in A. (C, D) Ensembles revealed robust tLTD at both low and high frequencies in MK-801 (blue triangles) compared to no-induction MK-801 controls (gray, Welch ANOVA p<0.01), suggesting that tLTD does not rely on ionotropic NMDAR signaling. (E) CV analysis indicated that tLTD at both induction frequencies was presynaptically expressed (HF LTD: φ=13°±5°, n=6, p<0.05; LF LTD: 15°±4°, n=5, p<0.05), in agreement with our prior findings (Figure 1; Sjöström et al., 2003; Sjöström et al., 2007). One experiment with <5% plasticity was excluded, see Methods.
Figure 3—figure supplement 1
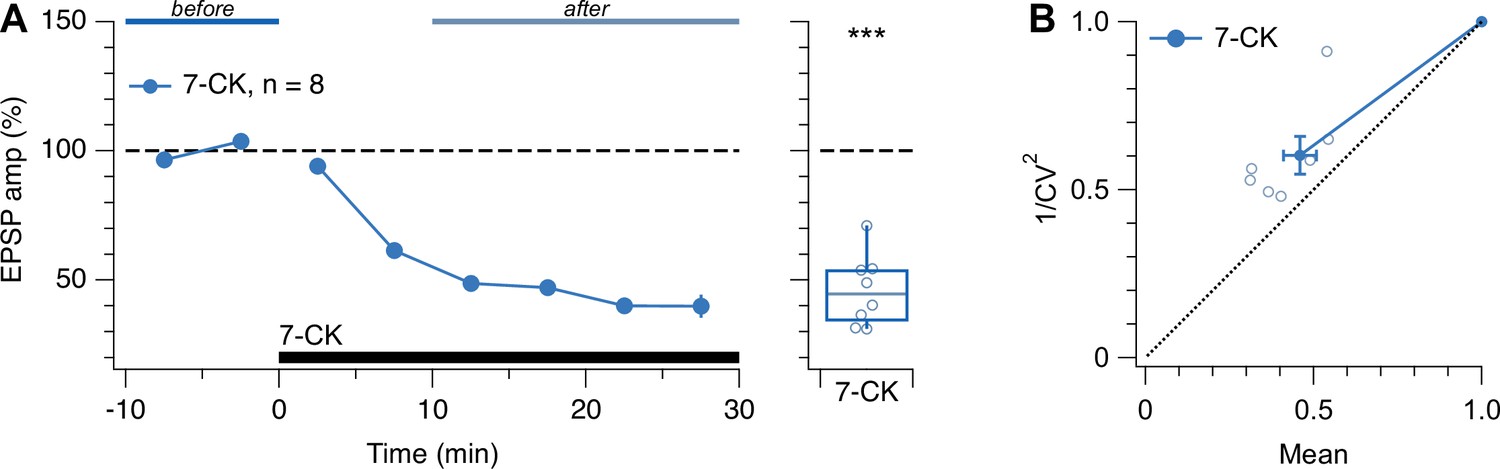
Low-frequency evoked release was suppressed by 7-CK.
(A) Washing in 7-CK drastically reduced EPSP amplitude (one-sample t-test for normalized EPSP vs. 100%, p<0.001). Presynaptic cells spiked once every 10 s. Experiments with 7-CK (Figure 4), therefore, required waiting for responses to asymptotically stabilize. Blue and light-blue lines indicate periods used for the quantification of EPSP amplitude. (B) CV analysis of the 7-CK-mediated suppression gave rise to data points above the diagonal (φ = –11°±4°, n=7, p<0.05), suggesting reduced quantal amplitude (Brock et al., 2020). This outcome is consistent with 7-CK also acting as a competitive inhibitor of L-glutamate transport into synaptic vesicles (Bartlett et al., 1998), so that after 7-CK, vesicles may contain less neurotransmitter. However, 1/CV2 was also decreased (after/before = 0.6 ± 0.06, n=7, one-sample t-test vs. 1, p<0.001), suggesting that prelease was simultaneously reduced (Brock et al., 2020).
Figure 4
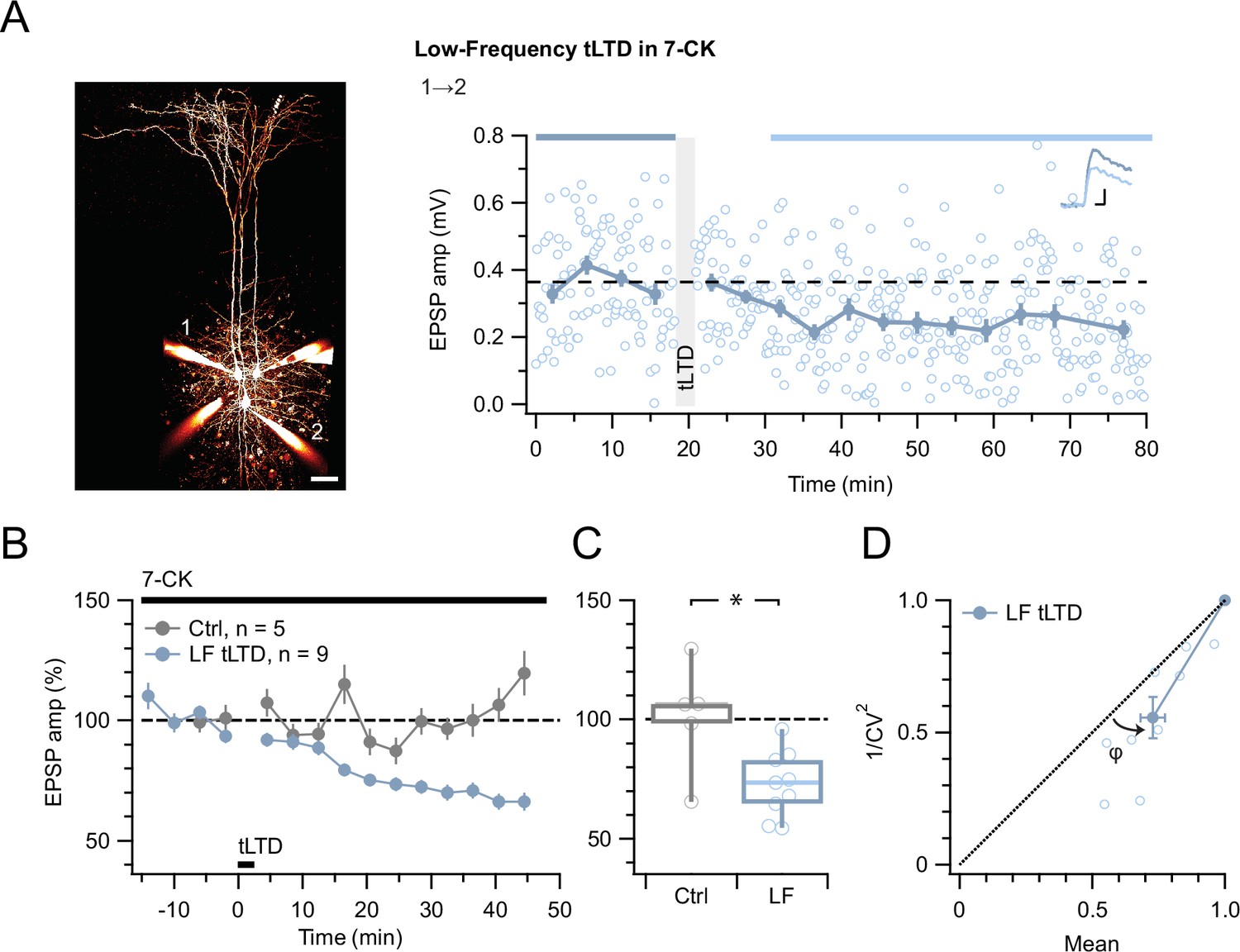
tLTD does not require ligand binding at NMDAR co-agonist site.
(A) At this sample connection (PC 1→2, see Alexa 594 fills at left, scale bar 50 µm), LF tLTD at 1 Hz (see Methods) persisted in the presence of 7-CK (after/before = 68 %, p<0.001). Traces were averaged over time periods indicated in blue/light blue. Baseline spiking was 0.1 Hz. Scale bars: 5 ms, 0.1 mV. (B, C) Ensemble revealed that LF tLTD persisted in 7-CK (blue) compared to no-induction 7-CK controls (gray), verifying that it does not require ionotropic NMDAR signaling. (D) CV analysis verified that LF tLTD was presynaptically expressed (φ=14°±3°, n=9, p<0.01), in agreement with our prior findings (Figures 1 and 3E; Sjöström et al., 2003; Sjöström et al., 2007).
Figure 5
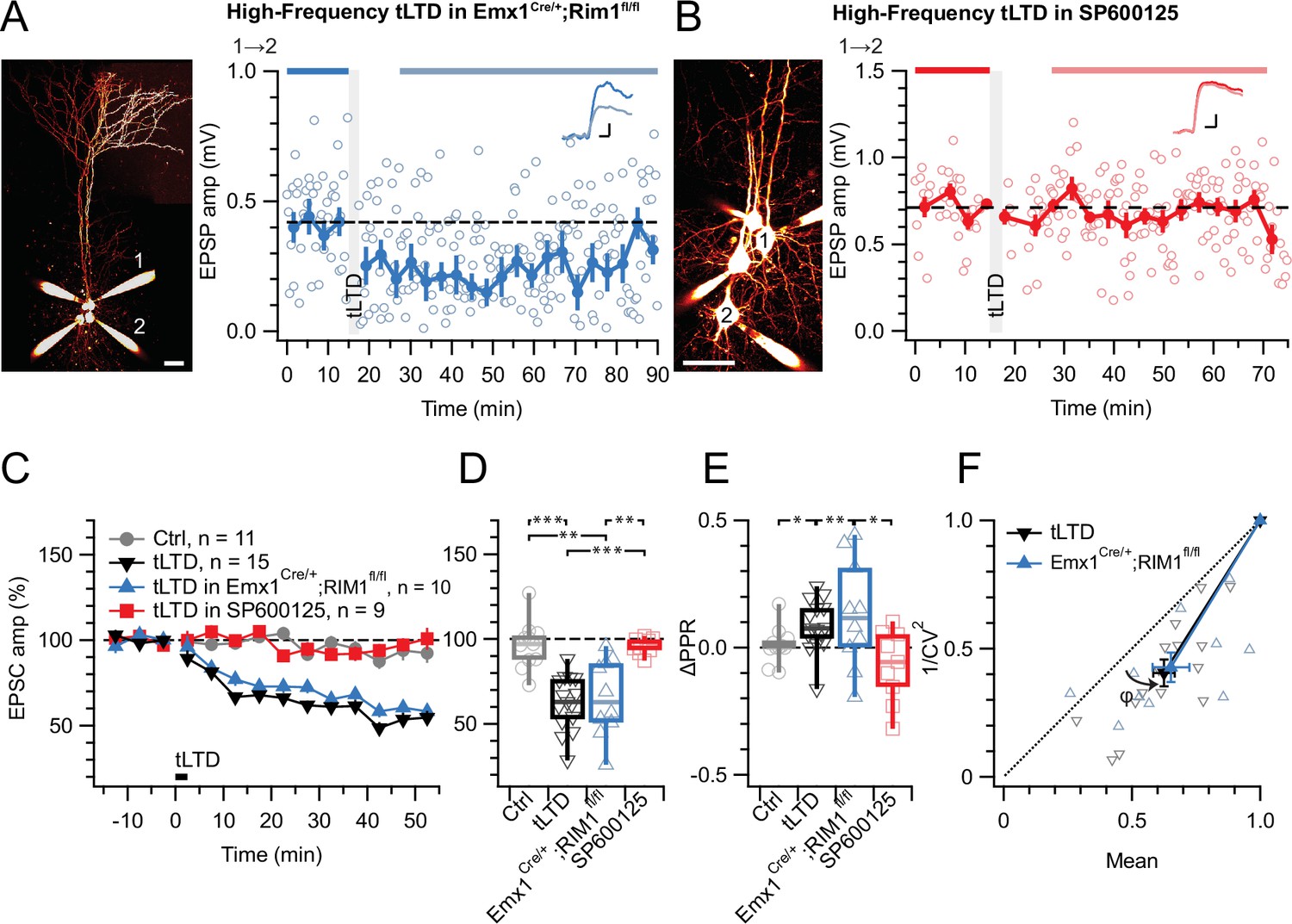
tLTD does not require RIM1αβ but relies on JNK2.
(A) At this sample PC 1→2 connection (left: Alexa 594 fills, scale bar 50 µm) in an acute slice from a homozygous RIM1αβ deletion mouse, tLTD persisted (after/before = 59%, p<0.001). Scale bars: 5ms, 0.1 mV. (B) For this sample connection, the JNK2-blocker SP600125 (Bennett et al., 2001) abolished tLTD (after/before = 99%, p=0.86). Scale bars: 5ms, 0.2 mV. (C, D) Ensemble data revealed that tLTD after homozygous RIM1αβ deletion (blue) was indistinguishable from WT tLTD (black), thereby dissociating tLTD from RIM1αβ. However, SP600125 robustly disrupted tLTD (red), with an outcome indistinguishable from no-induction controls (gray). Welch ANOVA p<0.001. (E) tLTD increased PPR (black and blue) compared to controls (gray), suggesting presynaptic expression. Welch ANOVA p<0.05. (F) CV analysis indicated that tLTD was presynaptically expressed, whether RIM1αβ was deleted (φ=16°±4°, n=10, p<0.01) or not (φ=13°±2°, n=15, p<0.001), in agreement with our prior findings (Figures 1 and 3E, Figure 4D; Sjöström et al., 2003; Sjöström et al., 2007).
Figure 6
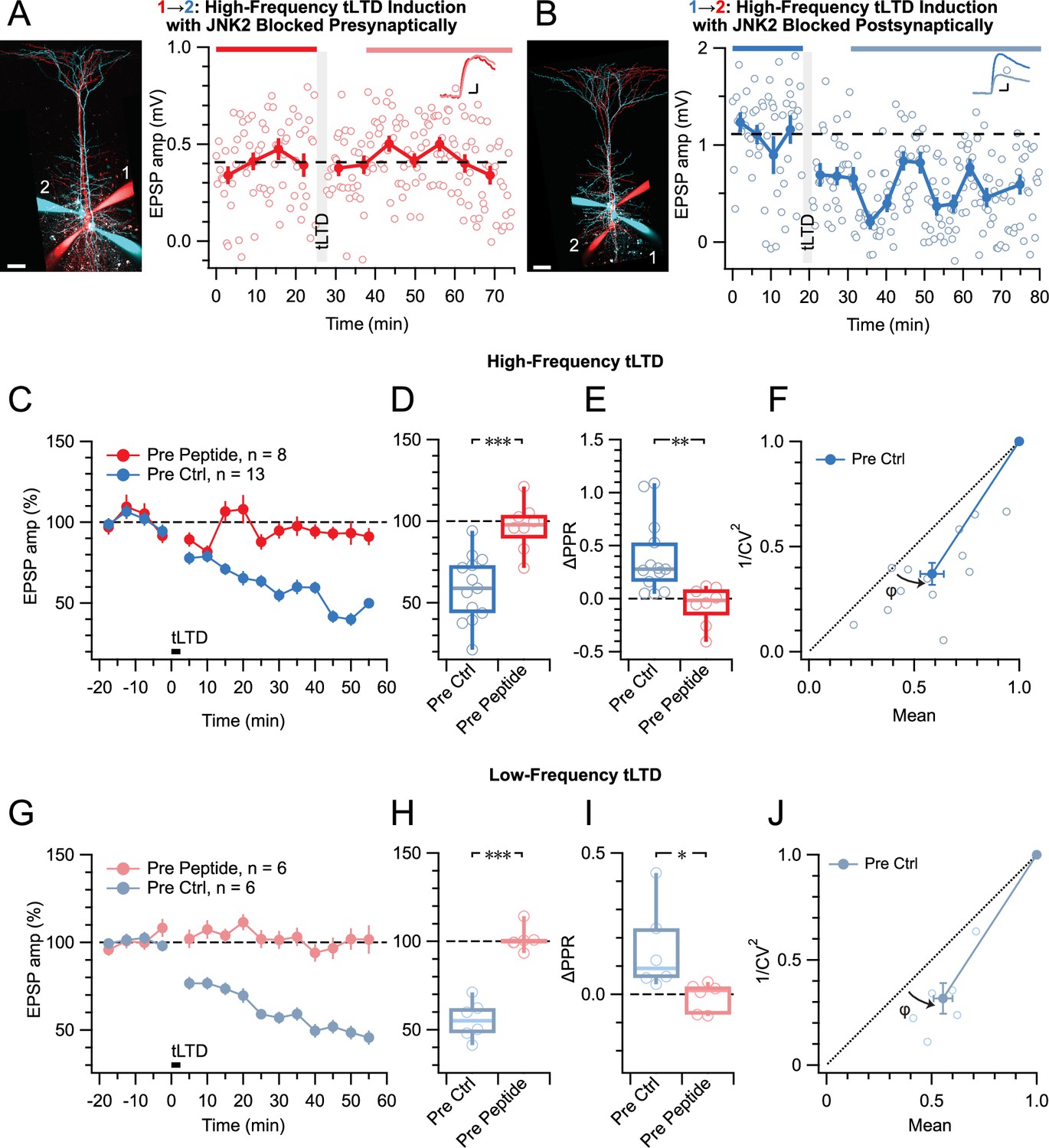
tLTD relies on presynaptic JNK2/STX1 signaling.
(A) Presynaptic loading of peptide disrupting JNK2/STX1 interactions (Marcelli et al., 2019) in this PC 1→2 pair abolished HF tLTD (after/before = 105%, p=0.52). Red cells were loaded with peptide, whereas blue cells were not. Traces were averaged over time periods indicated in red/pink. Scale bar 50 µm, inset scale bars 5 ms, 0.1 mV. (B) Postsynaptic peptide loading in this PC 1→2 pair had no effect on HF tLTD (after/before = 47%, p<0.001). Inset scale bars 5ms, 0.2 mV. (C, D) Presynaptic (red) but not postsynaptic/no peptide loading (blue) abolished HF tLTD, demonstrating a need for presynaptic JNK2/STX1 signaling. (E) Without presynaptic peptide (blue), HF tLTD induction increased PPR as expected from reduced release, whereas with presynaptic peptide (red), PPR expectedly remained unaffected. (F) CV analysis of HF tLTD with no presynaptic peptide agreed with presynaptic expression (φ=13°±3°, n=13, p<0.001). (G, H) Presynaptic (red) but not postsynaptic/no peptide loading (blue) abolished LF tLTD, demonstrating that the need for presynaptic JNK2/STX1 signaling was not frequency dependent like ionotropic preNMDAR signaling is (Abrahamsson et al., 2017). (I) Without presynaptic peptide (blue), LF tLTD induction increased PPR in keeping with presynaptic expression, but with presynaptic peptide (red), PPR was unaffected. (J) CV analysis of LF tLTD with no presynaptic peptide agreed that release was reduced (φ=11°±2°, n=6, p<0.01).
Additional files
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Neocortical layer-5 tLTD relies on non-ionotropic presynaptic NMDA receptor signaling
eLife 14:RP106284.
https://doi.org/10.7554/eLife.106284.3




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}