Transforming a fragile protein helix into an ultrastable scaffold via a hierarchical AI and chemistry framework
- State Key Laboratory of Coordination Chemistry, Nanjing Drum Tower Hospital, Affiliated Hospital of Medical School, School of Chemistry and Chemical Engineering, Chemistry and Biomedicine Innovation Center (ChemBIC), Frontier Interdisciplinary Science Research Center, Nanjing University, China
Figures
Figure 1 with 13 supplements

A hierarchical chemical blueprint for multi-axis stabilization of α-helical protein.
(a) The erythrocyte membrane skeleton, which relies on spectrin repeats to withstand shear stress. Inset: Structure of a single, naturally fragile spectrin repeat (PDB 3F57), with hydrophobic core residues shown. (b) The mechanical unfolding pathway of a spectrin repeat under tensile force, probed computationally by steered molecular dynamics (SMD) to generate force–extension curves. (c) Overview of the two-stage design strategy. Stage I (architectural stabilization): AI-guided backbone construction and computational screening generate four-helix designs with optimized hydrophobic cores. Stage II (precision functionalization): Rational installation of inter-helical salt bridges and metal-coordination motifs reinforces specific mechanical interfaces. (d) Stage I, backbone construction. RFdiffusion appends a fourth helix to the native three-helix template, generating 100 initial backbones. Five optimal four-helix scaffolds are selected, and ProteinMPNN is used to generate 100,000 sequences per scaffold. (e) Stage I, computational screening. A multi-step funnel prioritizes candidates through successive filters: developability (GRAVY score ≤−0.3), foldability (ESMFold triage followed by AlphaFold2 refinement with root mean square deviation [RMSD] ≤2.0 Å and pLDDT ≥90), and stability assessed via molecular dynamics. The process efficiently narrows ~106 initial designs to an experimentally tractable shortlist.
Figure 1—figure supplement 1

2D structure and sequence of the designed protein.
Figure 1—figure supplement 2

The steered molecular dynamics (SMD) simulation results of natural spectrin, SpecAI88, SpecAI41, and SpecAI89.
Figure 1—figure supplement 3

Thermal stability of natural spectrin and SpecAI designs via annealing molecular dynamics (MD).
Figure 1—video 1
Steered molecular dynamics (SMD) video of Spectrin unfolding under mechanical pulling.
Figure 1—video 2
Steered molecular dynamics (SMD) video of SpecAI88 unfolding under mechanical pulling.
Figure 1—video 3
Steered molecular dynamics (SMD) video of SpecAI41 unfolding under mechanical pulling.
Figure 1—video 4
Steered molecular dynamics (SMD) video of SpecAI89 unfolding under mechanical pulling.
Figure 1—video 5
Molecular dynamics (MD) video of Spectrin and SpecAI series unfolding under mechanical pulling.
Figure 1—video 6
Annealing molecular dynamics (AMD) video of Spectrin upon thermal denaturation.
Figure 1—video 7
Annealing molecular dynamics (AMD) video of SpecAI88 upon thermal denaturation.
Figure 1—video 8
Annealing molecular dynamics (AMD) video of SpecAI41 upon thermal denaturation.
Figure 1—video 9
Annealing molecular dynamics (AMD) video of SpecAI89 upon thermal denaturation.
Figure 1—video 10
Annealing molecular dynamics (AMD) video of Spectrin and SpecAI series upon thermal denaturation.
Figure 2 with 1 supplement

Thermal and chemical robustness of AI-designed spectrin variants.
(a) MALDI-TOF mass spectrometry confirms molecular weights for SpecAI88, SpecAI41, and SpecAI89. (b) Far-UV circular dichroism (CD; 190–260 nm) shows α-helical signatures with minima at 208 and 222 nm. Temperature-dependent CD (20–100°C, blue to red) indicates substantial retention of ellipticity at 195 nm, with melting temperatures exceeding 100°C. (c) CD at 222 nm recorded in GdnHCl demonstrates persistence of α-helical signal at high denaturant concentrations (~3 M), indicating high chemical resistance.
Figure 2—figure supplement 1

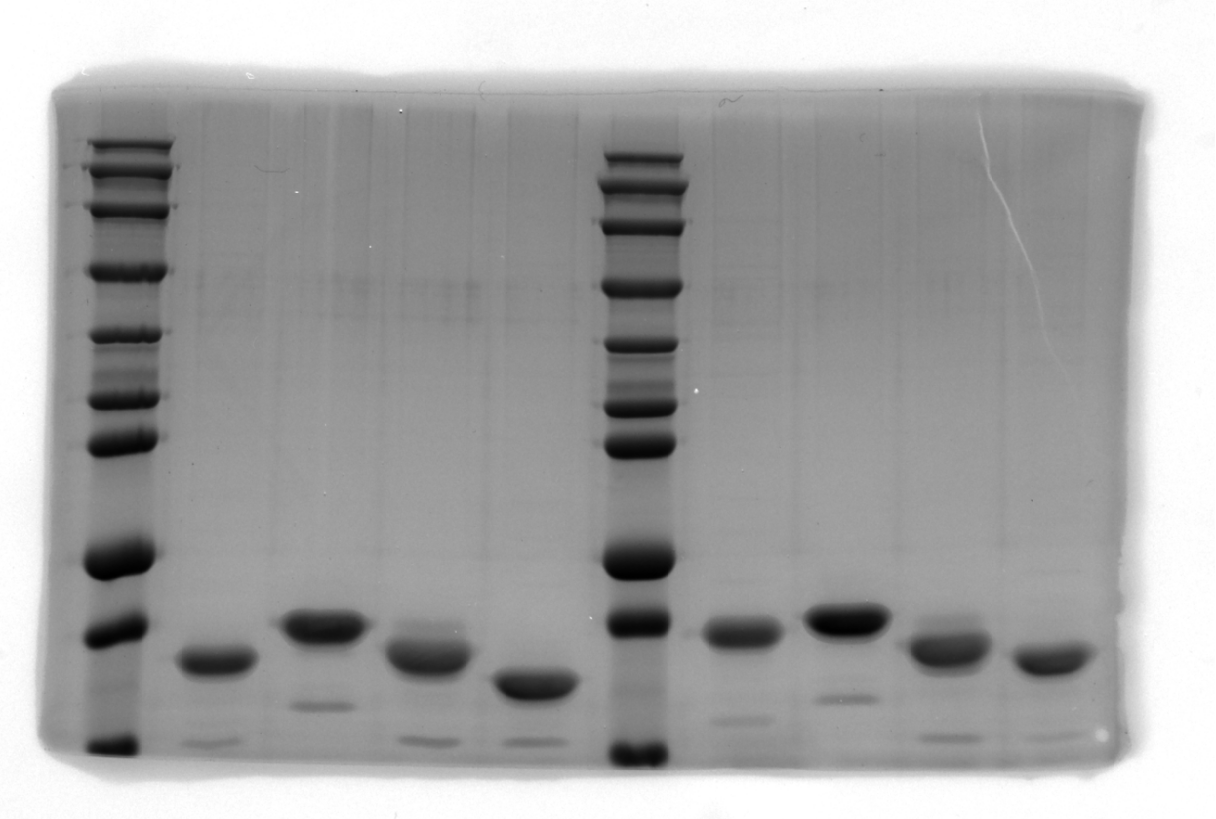
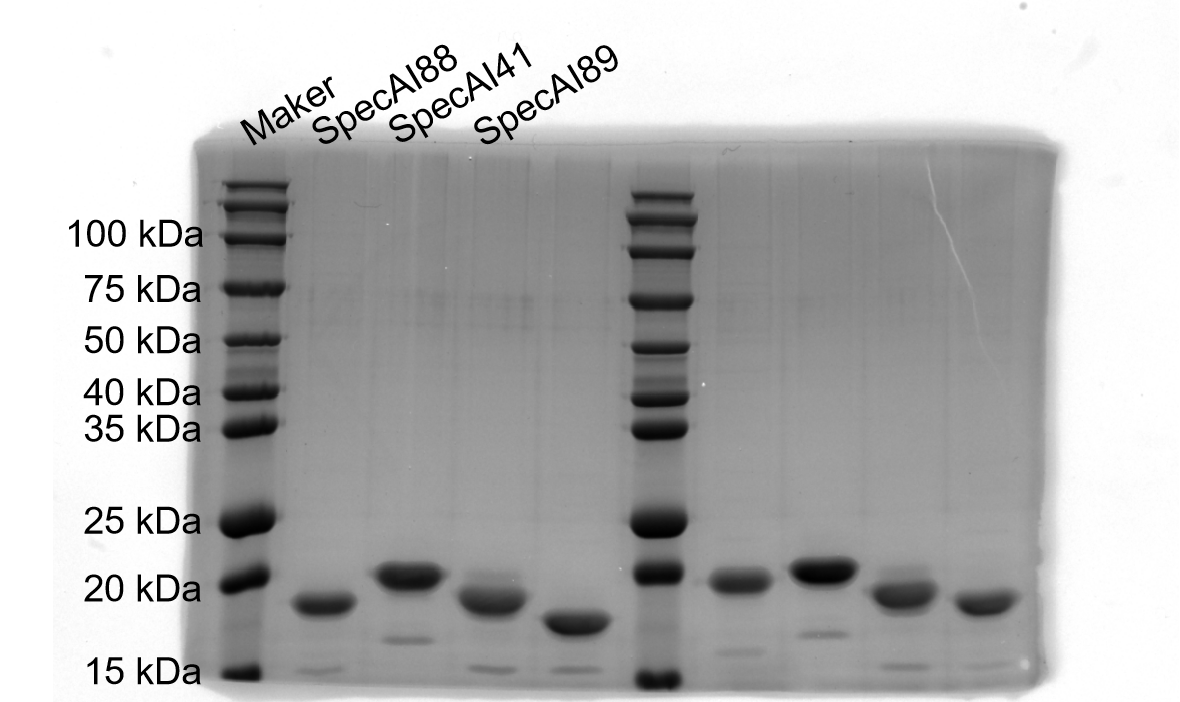
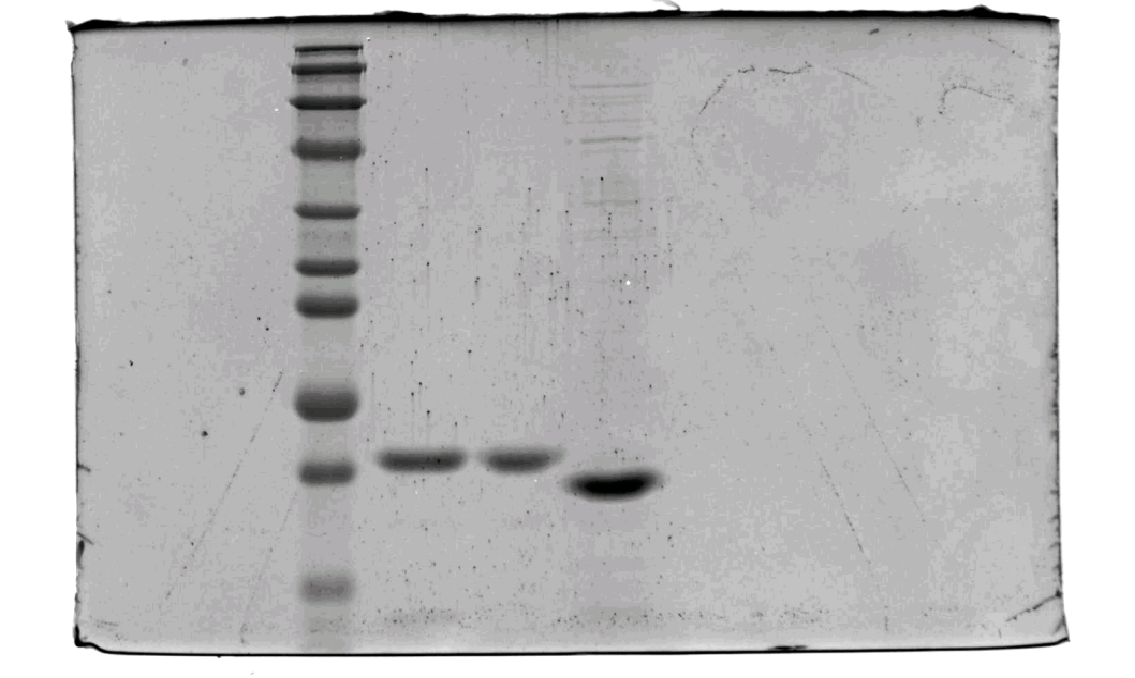
SDS–PAGE results of SpecAI variants, show their successful expression with expected MW of ~19 kDa.
-
Figure 2—figure supplement 1—source data 1
Original SDS-PAGE gel.
- https://cdn.elifesciences.org/articles/109753/elife-109753-fig2-figsupp1-data1-v1.png
-
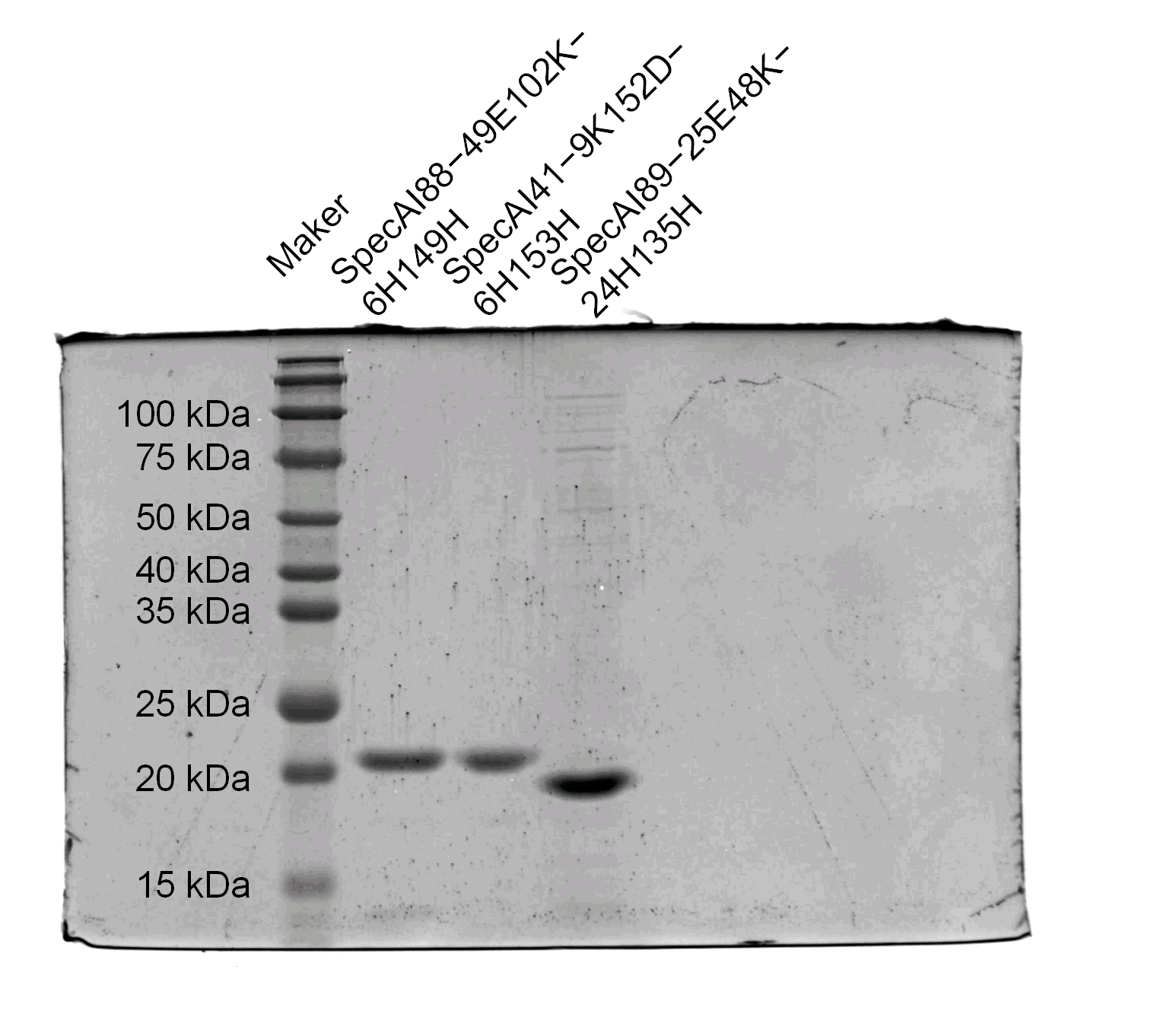
Figure 2—figure supplement 1—source data 2
Original SDS-PAGE gel, indicating the relevant bands.
- https://cdn.elifesciences.org/articles/109753/elife-109753-fig2-figsupp1-data2-v1.png
Figure 3

Stage I four-helix bundle designs exhibit enhanced mechanical stability by single-molecule force spectroscopy.
(a) Schematic of the AFM–SMFS experimental setup. A dockerin (Doc)-functionalized AFM tip engages a cohesin (Coh)-tagged SpecAI construct immobilized on the surface, enabling single-molecule pulling. The construct includes three GB1 domains as fingerprint markers. (b) Representative force–extension curves. Curve 1: Unfolding of the natural spectrin repeat (ΔLc ≈ 32 nm, red), followed by three GB1 (ΔLc ≈ 18 nm per domain, black). Curves 2–4: Unfolding of SpecAI variants (ΔLc ≈ 53 nm). Sometimes, the Doc unfolds, showing additional peak (Curve 3). Dashed lines are worm-like chain model fit. (c) Unfolding force histograms of SpecAI (bin size = 30 pN) with Gaussian fits demonstrate a significant increase in mechanical stability compared to the native spectrin repeat (56 ± 3 pN, n = 212, bin size = 13.75 pN): SpecAI88, 116 ± 2 pN (n = 224); SpecAI41, 156 ± 4 pN (n = 218); and SpecAI89, 121 ± 4 pN (n = 174). The corresponding ΔLc distributions are centered near 52 nm: SpecAI88, 52 ± 2 pN (n = 224); SpecAI41, 53 ± 1 pN (n = 218); and SpecAI89, 52 ± 2 pN (n = 174), consistent with the full unfolding of the designed domain.
Figure 4 with 5 supplements

Precision stabilization of designed proteins via electrostatic interactions and metal coordination.
(a) Schematic of Stage II design: introducing inter-helical ion pairs and metal-coordination sites into AI-designed backbones to stabilize specific interfaces. (b) AlphaFold3-predicted structure of variant SpecAI41-9K152D, showing an engineered salt bridge (Lys9–Asp152, 3 Å) designed for electrostatic stabilization without perturbing the core. The color of protein is based on the pLDDT value, showing a high-confidence structure prediction (>80). (c) A representative force–extension curve for the salt-bridge variant shows an unfolding event with a ΔLc of 53 nm, consistent with the parent scaffold. (d) Unfolding force histograms reveal a ~25 pN increase in mechanical stability for salt-bridge variants compared to their Stage I parents, with forces reaching 180 ± 4 pN (n = 197). (e) Introduction of a metal-binding site in variant SpecAI41-9K152D-6H153H, with two histidines positioned 2 Å apart, compatible with Ni2+ coordination. (f) A representative force–extension curve recorded in 200 µM Ni2+ shows the unfolding event (ΔLc ≈ 53 nm). (g) Unfolding force histograms confirm enhanced mechanical stability, with forces reaching 208 ± 3 pN (n = 222) , a significant gain over the salt-bridge parent.
Figure 4—figure supplement 1

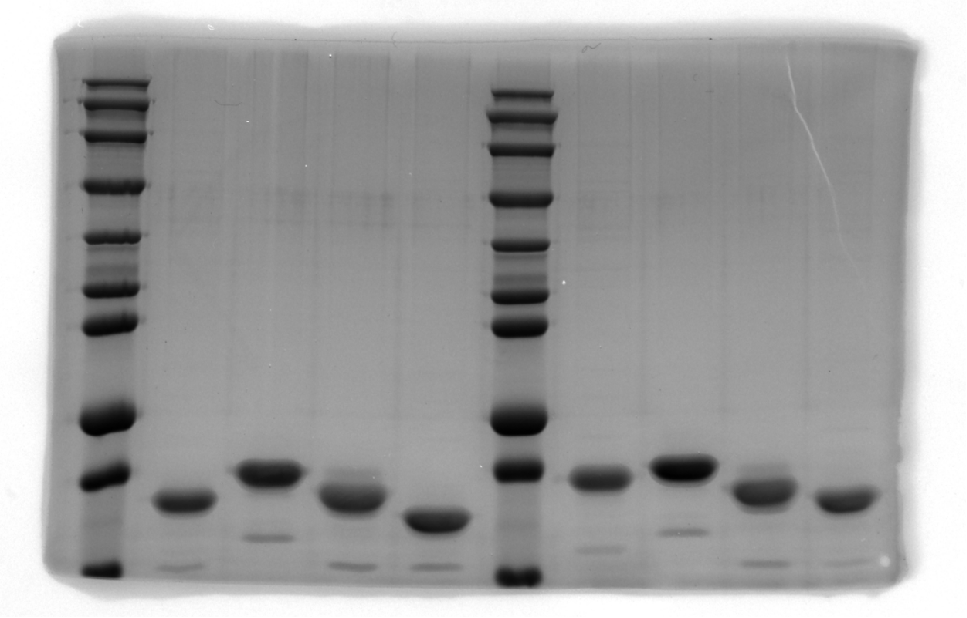
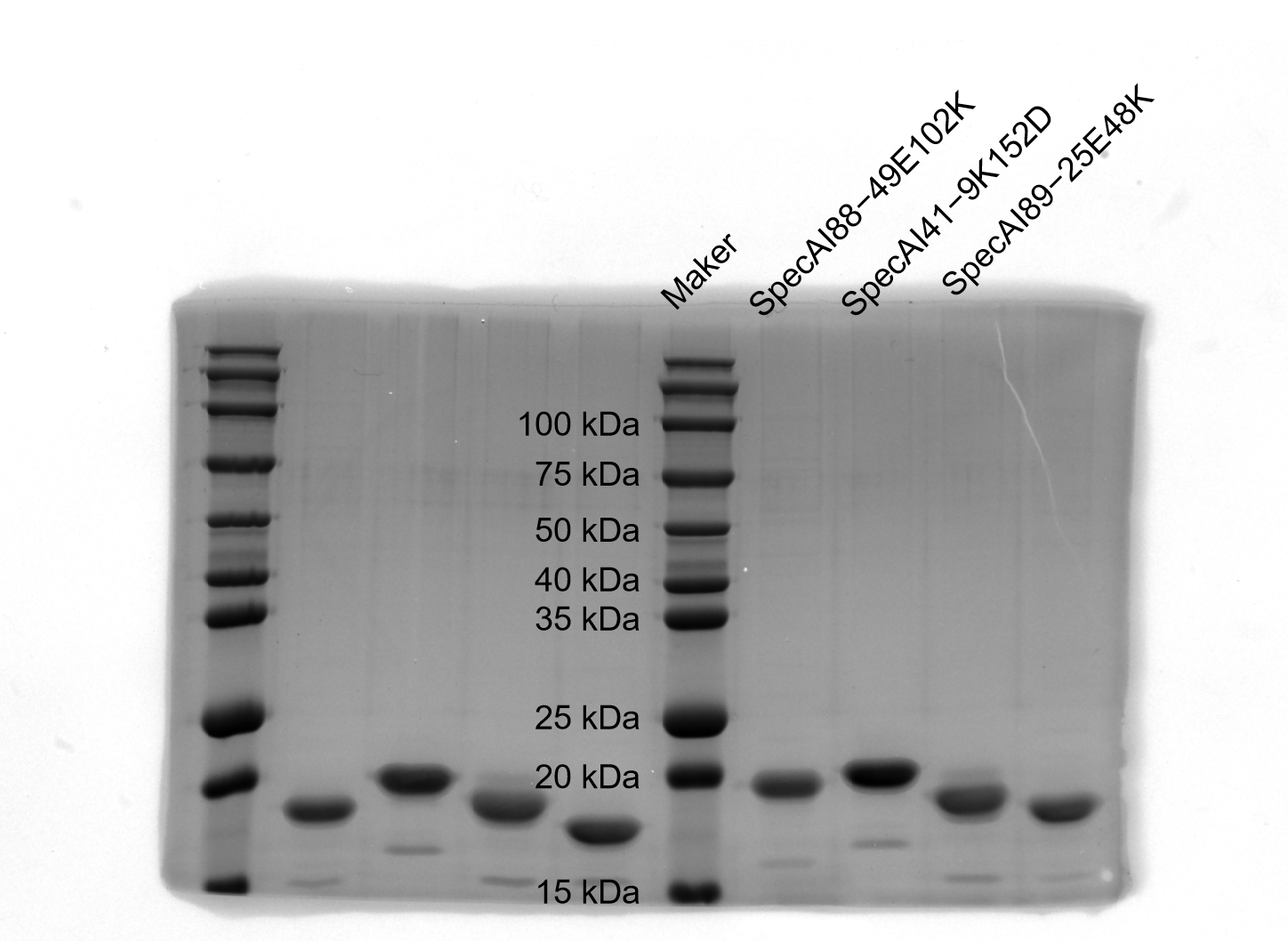
SDS–PAGE results of SpecAI variants with salt bridge, show their successful expression with expected MW of ~19 kDa.
-
Figure 4—figure supplement 1—source data 1
Original SDS-PAGE gel.
- https://cdn.elifesciences.org/articles/109753/elife-109753-fig4-figsupp1-data1-v1.png
-
Figure 4—figure supplement 1—source data 2
Original SDS-PAGE gel indicating the relevant bands.
- https://cdn.elifesciences.org/articles/109753/elife-109753-fig4-figsupp1-data2-v1.png
Figure 4—figure supplement 2

SDS–PAGE gel results of SpecAI variants with bis-histidine mutation, show their successful expression with expected MW of ~19 kDa.
-
Figure 4—figure supplement 2—source data 1
Original SDS-PAGE gel.
- https://cdn.elifesciences.org/articles/109753/elife-109753-fig4-figsupp2-data1-v1.png
-
Figure 4—figure supplement 2—source data 2
Original SDS-PAGE gel indicating the relevant bands.
- https://cdn.elifesciences.org/articles/109753/elife-109753-fig4-figsupp2-data2-v1.png
Figure 4—figure supplement 3

Engineered salt bridges and metal coordination enhance mechanical stability in SpecAI88 and SpecAI89.
(a-c) Introduction of a Glu49-Lys102 salt bridge (3 Å) in SpecAI88 increases its mean unfolding force to 141 ± 3 pN (n = 202) . (d-f) Adding a His6-His149 metal-binding site (4 Å) to the salt-bridge variant further raises the unfolding force to 190 ± 3 pN (n = 232). (g-l) Similar stabilization in the SpecAI89 series: a Glu25-Lys48 salt bridge (4 Å) confers 153 ± 4 pN (n = 264) stability, increased to 173 ± 3 pN (n = 204) with an added His24-His135 metal-coordination motif (2 Å). All variants show a consistent unfolding ΔLc of 53 nm. Data from AFM-SMFS experiments; structures predicted by AlphaFold3.
Figure 4—figure supplement 4

Dynamic force spectroscopy reveals the kinetic stabilization of the SpecAI41 series.
For SpecAI41, the unfolding forces at 400 nm/s, 1000 nm/s, 2000 nm/s, and 4000 nm/s were F = 134 ± 48 pN (n = 143), F = 156 ± 59 pN (n = 218), F = 174 ± 56 pN (n = 197), and F = 200 ± 60 pN (n = 145), respectively. For SpecAI41-9K152D, the unfolding forces at the corresponding pulling speeds were F = 169 ± 37 pN (n = 154), F = 180 ± 56 pN (n = 197), F = 201 ± 62 pN (n = 156), and F = 220 ± 36 pN (n = 142), respectively. For SpecAI41-9K152D-6H153H, the unfolding forces were F = 182 ± 35 pN (n = 136), F = 208 ± 45 pN (n = 222), F = 222 ± 60 pN (n = 144), and F = 237 ± 60 pN (n = 147), respectively. Data here are shown as mean ± SD (normal distribution assumed).
Figure 4—figure supplement 5

Thermal and chemical stability profiles of Stage II constructs.
Figure 5

Hierarchical computational design of ultrastable proteins through multi-scale stabilization.
Schematic summarizing the two-stage design strategy for additive mechanical reinforcement. Stage I establishes a stable architectural framework through molecular dynamics (MD) screening of AI-designed hydrophobically optimized cores. Stage II introduces precision functionalization via inter-helical salt bridges and bi-histidine metal-coordination motifs, guided by AlphaFold3 structural models. The integration of global architectural stability with local chemical cross-linking produces additive mechanical reinforcement, enabling the creation of ultrastable protein domains.
Author response image 1

Analysis of the minimum atomic distance between the protein and its periodic images under periodic boundary conditions.
Left: SpecAI89 during SMD simulations, showing that the minimum protein–image distance remains above 1.0 nm for the majority of the simulation time. Right: WT during AMD simulations, where transient proximity to the periodic boundary is observed at very late stages due to large conformational fluctuations.
Author response image 2

The backbone RMSD of SpecAI89 over time during simulation.
Author response image 3

The bimodal fit for unfolding force of SpecAI88-49E102K-6H149H show the same 190 pN unfolding for the first peak as previous fit.
Author response image 4

Spearman correlation between SMD and AFM unfolding forces for natural spectrin and SpecAI designs.
SMD force (x-axis) versus experimental AFM force (y-axis); each point represents one protein.
Tables
Table 1
Stability of AI-designed spectrin with four α-helices.
| Protein | SMD (pN)1 nm/ns | Tm (°C) | Yield (mg/l) | AFM (pN) | ΔLc (nm) | ΔF (pN) |
|---|---|---|---|---|---|---|
| Spectrin | 599 ± 62 | ~50 | ~50 | 56 ± 3 | 32 ± 1 | N/A |
| SpecAI88 | 735 ± 80 | >100 | 580 ± 36 | 116 ± 2 | 52 ± 2 | 60 |
| SpecAI41 | 689 ± 84 | >100 | 625 ± 28 | 156 ± 4 | 53 ± 1 | 100 |
| SpecAI89 | 670 ± 84 | >100 | 450 ± 16 | 121 ± 4 | 52 ± 2 | 61 |
Table 2
SpecAI stability further enhanced by site-specific functionalization.
| Protein | Force (pN) | ΔLc (nm) | ΔF (pN) | Total ΔF |
|---|---|---|---|---|
| SpecAI88-49E102K | 141 ± 3 | 54 ± 1 | 25 | 74 |
| +6H149H | 190 ± 3 | 53 ± 1 | 49 | |
| SpecAI41-9K152D | 180 ± 4 | 53 ± 2 | 24 | 52 |
| +6H153H | 208 ± 3 | 53 ± 2 | 28 | |
| SpecAI89-25E48K | 153 ± 4 | 53 ± 1 | 32 | 52 |
| +24H135H | 173 ± 3 | 53 ± 1 | 20 |
-
Total ΔF means the unfolding force increase over SpecAI.
Additional files
-
MDAR checklist
- https://cdn.elifesciences.org/articles/109753/elife-109753-mdarchecklist1-v1.docx
-
Source data 1
Candidate proteins identified through ESMFold screening, followed by AlphaFold2 structural prediction and subsequent molecular dynamics simulations.
- https://cdn.elifesciences.org/articles/109753/elife-109753-data1-v1.xlsx
-
Supplementary file 1
Protein sequences for AFM-SMFS measurement.
- https://cdn.elifesciences.org/articles/109753/elife-109753-supp1-v1.docx
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Transforming a fragile protein helix into an ultrastable scaffold via a hierarchical AI and chemistry framework
eLife 15:RP109753.
https://doi.org/10.7554/eLife.109753.3




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}