Imaging a memory trace over half a life-time in the medial temporal lobe reveals a time-limited role of CA3 neurons in retrieval
- Ruhr University Bochum, Germany
- Otto von Guericke University, Germany
- Leibniz-Institute for Neurobiology, Germany
- Osaka University, Japan
Figures
Figure 1

Contextual fear-conditioning task and memory performance.
(A) Schematic description of the task. Mice were exposed to a conditioning chamber they explored freely for 2.5 min after which they received a single 1 mA footshock and were returned to their home-cage after 3.5 additional min. Memory for the association of the footshock with the conditioning context was tested after either 1 day, 1 week, 1 month, 6 months or a year by re-exposing the conditioned mice to the conditioning chamber and measuring freezing levels (n=4 mice per delay; n=20 total). Upon completion of the task, mice were sacrificed and their brain processed for imaging. ‘No-shock’ age-matched control groups were exposed to the exact same experimental conditions but did not receive any footshock (n=4 mice per delay; n=20 total). Of note, additional age-matched home-cage control groups were generated to control for Arc baseline expression (n=4 mice per delay; n=20 total; not shown here). (B) Differences in freezing indices between ‘shock’ and ‘no-shock’ groups (e.g. normalized freezing indices) in function of the age of the memory trace. All shocked groups showed stronger freezing than their no-shock controls at test as shown by differences in freezing levels significantly higher than 0 (a 55.9 ± 3.4% increase in average). Importantly, freezing levels were similar across delays, suggesting that memory strength did not significantly differ as memory aged. (C) Normalized freezing index induced by exposure either to the conditioning chamber or to a new context. Two additional age-matched groups (‘new context’ groups) were conditioned and normalized freezing levels were evaluated as mice were exposed at test to a new context instead of the conditioning chamber one day or one year after conditioning (n=4 mice per delay; n=8 total). Conditioned mice reexposed to the conditioning chamber froze in average 36 ± 4.7% more than the conditioned mice tested in the new context, demonstrating that the freezing levels were specific to the conditioning chamber. Error bars are mean ± SEM. ‘o’ indicate a significant comparison to 0 at p<0.05; asterisks a significant effect of the context at p<0.05 for ‘*’ and p<0.01 for ‘***’.
Figure 2

Imaging brain activity in the medial temporal lobe (MTL).
(A) Location of counting frames for CA1, CA3, the perirhinal (PER), the lateral entorhinal (LEC), the postrhinal (POR) and the medial entorhinal cortices (MEC). Task-induced Arc nuclear RNAs’ expression was detected on three nonconsecutive brain slices for each target area covering approx. 800 microns. (B-G) Cells activated during the retrieval of one day–old and one year-old memory traces in CA1, CA3 and the LEC: Arc positive cells (red arrowheads) and an exemplar of non-activated cells Arc negative cells (white arrowheads). Cell nuclei are labeled in blue with DAPI. (B,C) CA1 is engaged independently of the age of the memory trace while (D,E) activity levels in CA3 are negligible for very remote memories. (F-G) The parahippocampal areas, for example the LEC, are maximally engaged during the retrieval of very remote memories.
Figure 3
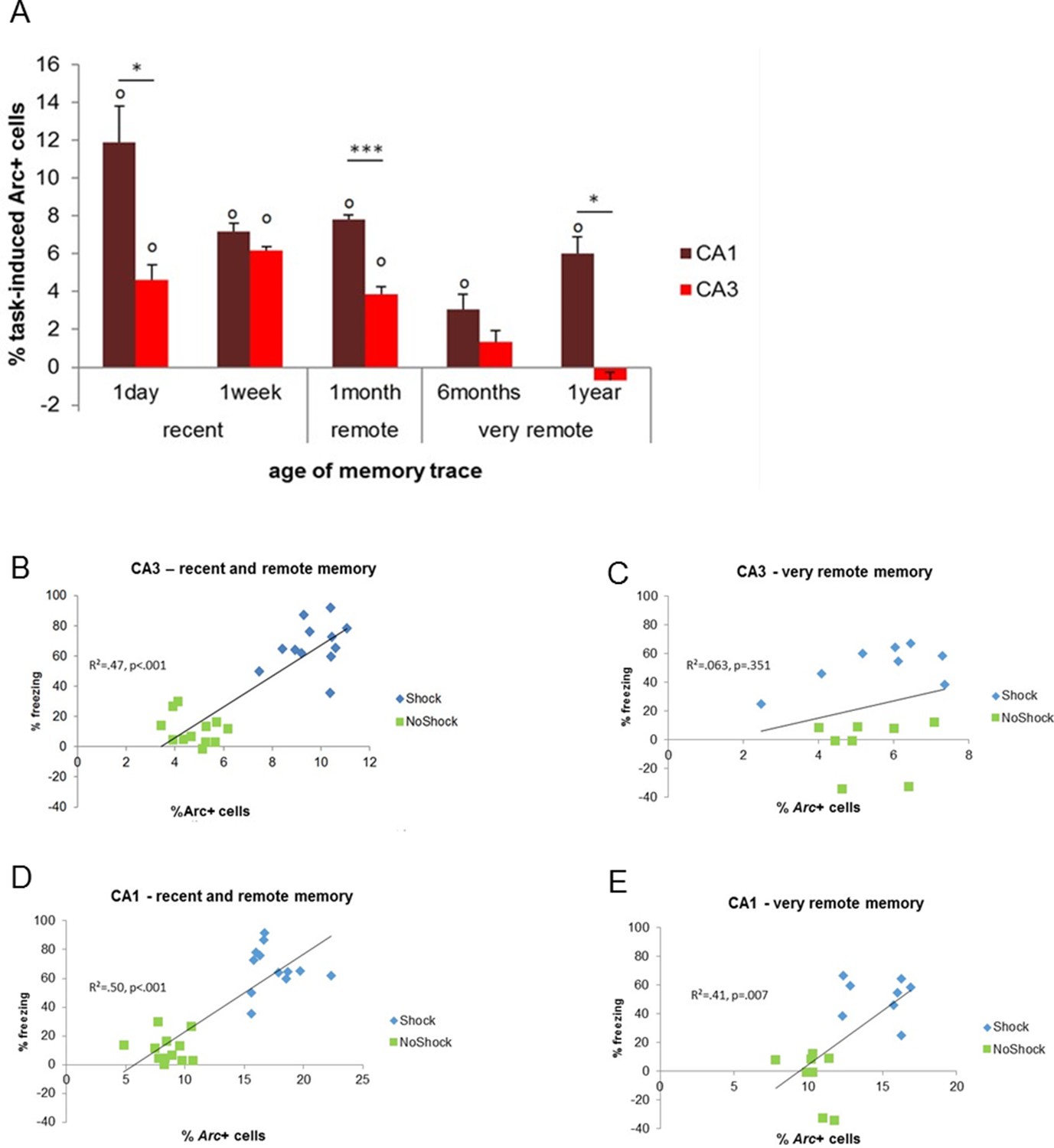
Activity patterns in CA1 and CA3 over time and correlations with memory performance.
(A) CA3’s contribution to memory retrieval depends on the age of the memory trace, not CA1’s, as CA3’s was no longer significantly recruited for the retrieval of very remote memories. (B,C) CA3’s activity levels were predictive of memory performance only for the retrieval of recent and early remote memories while D,E) CA1’s activity was independently of the age of the memory trace, Error bars are mean ± SEM. ‘o’ indicate a significant comparison to 0 at p<0.05; asterisks a t-test at p<0.05 for ‘*’ and at p<0.01 for ‘***’.
Figure 4
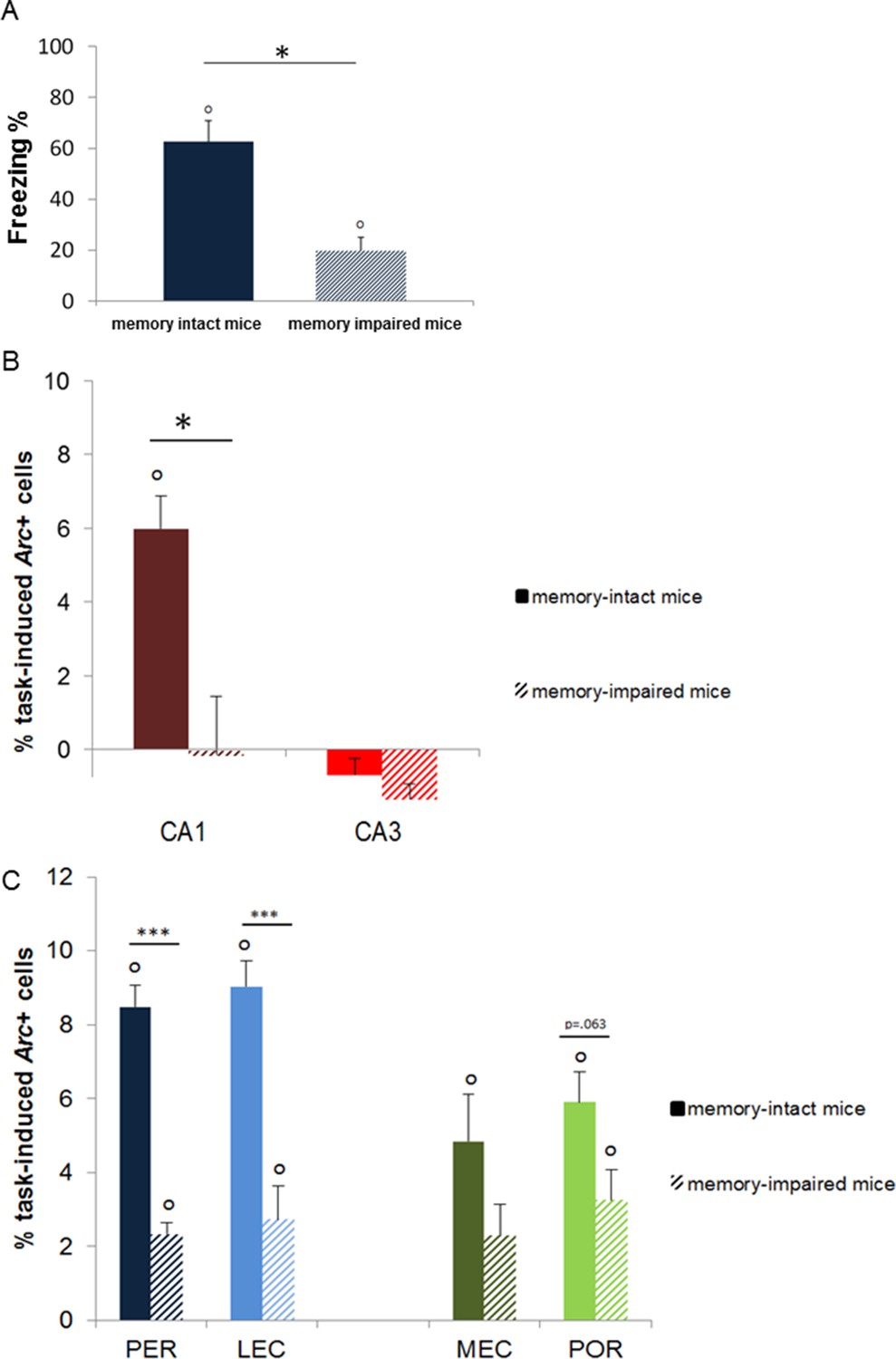
Memory performance of ‘memory-intact’ and ‘memory- impaired’ mice tested one year after conditioning and corresponding MTL patterns of activity.
(A) Behavioral performance: ‘memory-impaired’ mice froze significantly less than ‘memory-intact’ mice at test, reflecting impaired memory retrieval in this group. (B) Activity patterns in CA1 and CA3: in contrast to mice that successfully retrieved the footshock-context association one year after conditioning, CA1 was not recruited in those that had impaired memory. In addition, activity levels in CA3 were comparable between the two groups, underlining the critical role of CA1 in the retrieval of very remote memories. (C) Activity patterns in parahippocampal areas: parahippocampal areas of the ‘memory-impaired’ mice were recruited to much lesser extent than those of ‘memory-intact’ animals. Since this reduced activation did not lead to successful memory retrieval, this result suggests that CA1 might play a role as important as the cortical areas in the retrieval of very remote memories in memory-intact mice. Error bars are mean ± SEM. ‘o’ indicate a significant comparison to 0 at p<0.05; asterisks a t-test at p<0.05 for ‘*’ and at p<0.01 for ‘***’.
Figure 5
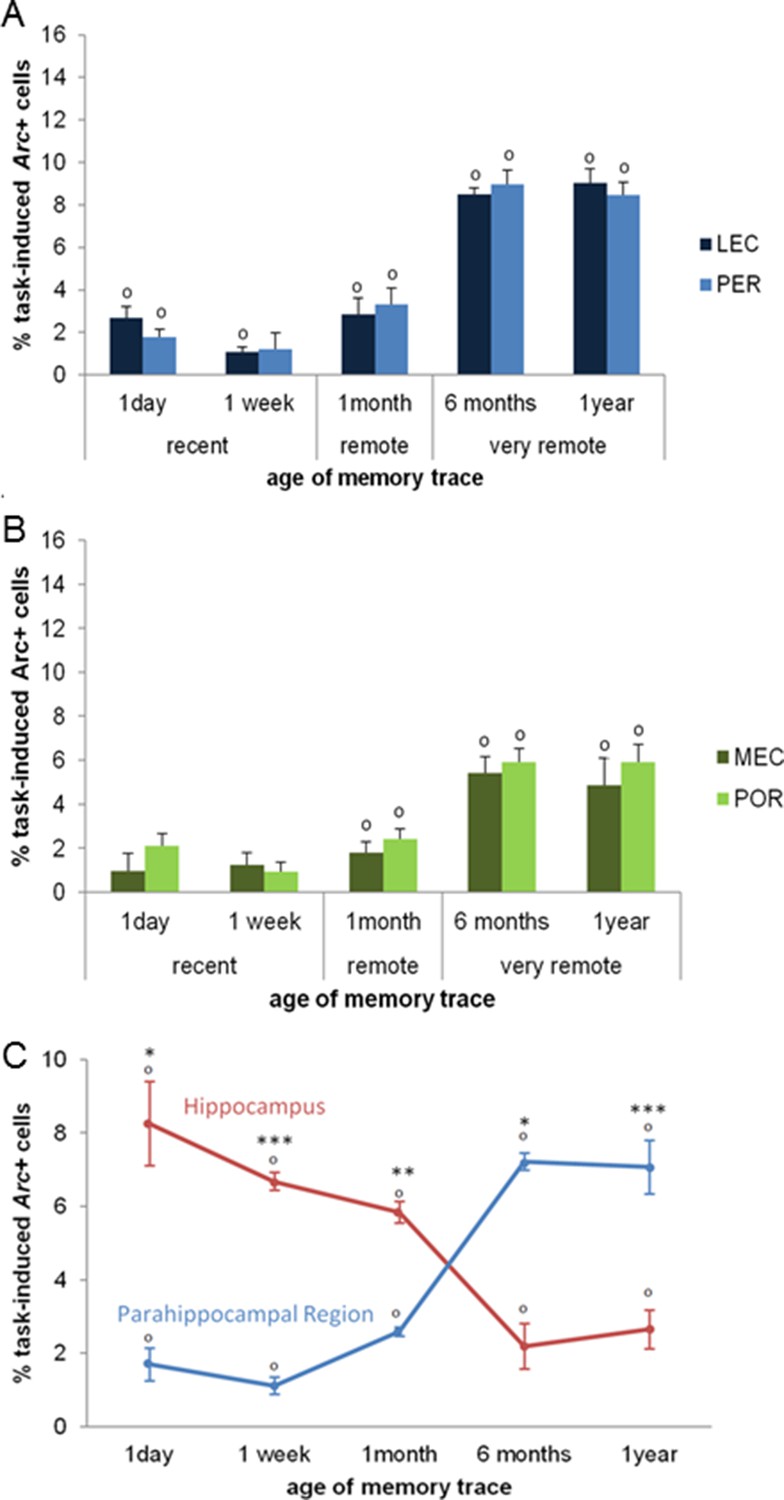
Activity patterns in the parahippocampal areas over time and apparent (and possibly misleading) over-time shift from the involvement of the hippocampus to the involvement of the parahippocampal region in memory retrieval.
All parahippocampal areas were maximally engaged for the most remote memories and patterns of activity were comparable between the (A) LEC and PER and (B) the MEC and POR. Furthermore, in line with the existence of stronger projections from the amygdala to the PER and LEC than to the POR and MEC, and a more important role of the cortical areas for the most remote memories than for more recent ones, the levels of activity in the PER and the LEC were higher than those in the POR and MEC during the retrieval of very remote memories, providing further support to an emerging theory according to which the parahippocampal areas might be segregated in terms of memory types/processes rather than in terms of information content (spatial versus spatial information; Eichenbaum et al., 2007; of note, for the sake of clarity, significant area differences between graphs A and B are not shown). (C) Contribution of the hippocampal CAs and parahippocampal region to memory retrieval over time: when CA1 and CA3 activity levels are not dissociated, the overall contribution of the hippocampus to the retrieval of very remote memories is largely underestimated when compared to that of CA1 shown in Figure 3A. Even in this case though, the hippocampus is still significantly recruited at all times. In contrast, overall activity of the parahippocampal region is comparable to that of any of the parahippocampal areas, with a maximal activation during the retrieval of very remote memories (see Figure 5A and B). These results underline the need of dissociating CA1 and CA3 activity patterns to better understand the contribution of the hippocampus to the retrieval of memory over time. Importantly, when these contributions are not dissociated, the apparent over-time shift remains relative at most (and not absolute) since both areas are significantly activated at all delays. Error bars are mean ± SEM. ‘o’ indicate a significant comparison to 0 at p<0.05; asterisks a t-test at p<0.05 for ‘*’ and at p<0.01 for ‘***’.
Author response image 1
we have now integrated these points in the results section of revised manuscript b adding following sentence.
https://doi.org/10.7554/eLife.11862.008Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Imaging a memory trace over half a life-time in the medial temporal lobe reveals a time-limited role of CA3 neurons in retrieval
eLife 5:e11862.
https://doi.org/10.7554/eLife.11862




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}