Tau and spectraplakins promote synapse formation and maintenance through Jun kinase and neuronal trafficking
- The University of Manchester, United Kingdom
- Institute of Translational Medicine, University of Liverpool, United Kingdom
- Universidad de Valladolid-CSIC, Spain
Figures
Figure 1 with 2 supplements
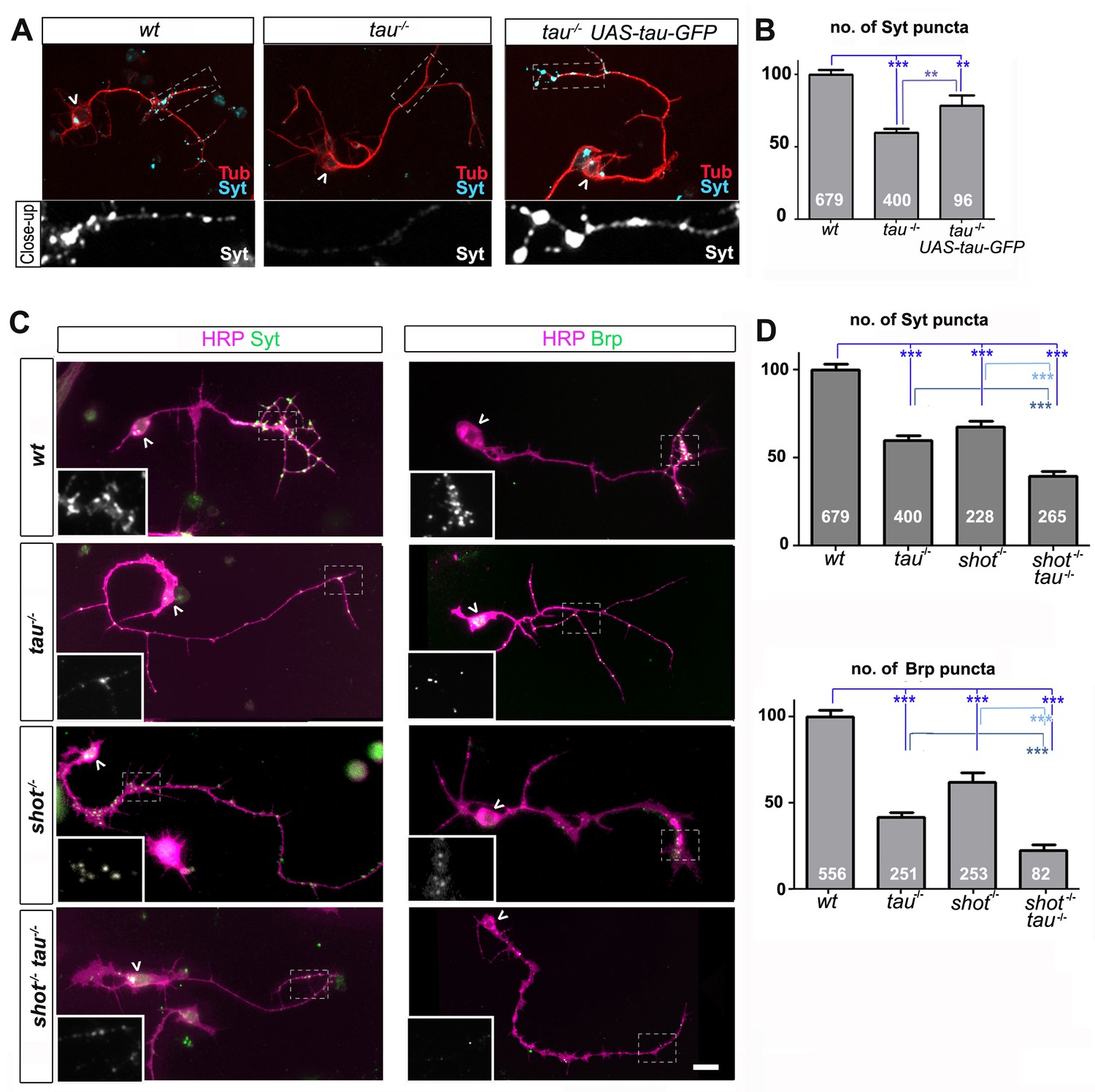
Tau and Shot are required for the formation of synaptic specialisations in axons.
(A) Primary neurons at 2 DIV obtained from embryos that were wildtype (wt), tau-/-, and tau-/- with elav-Gal4 driven expression of UAS-tau-GFP; neurons were stained for tubulin (Tub, red) and the synaptic protein Synaptotagmin (Syt, light blue). (B) Quantification of the experiment shown in A, shown as the number of Syt puncta per neuron, normalised to wildtype (the assessed numbers of neurons are indicated in each bar, ***PMW<0.001, **PMW<0.01). (C) Primary Drosophila neurons at 2DIV, obtained from embryos that were wildtype (wt), tauMR22(tau-/-), shot3(shot-/-), and shot3 tauMR22(shot-/- tau-/-), co-stained with antibodies against HRP (magenta) and the synaptic proteins (green) Syt and Bruchpilot (Brp); areas emboxed with dashed lines are displayed as magnified insets showing the synaptic staining only. (D) Quantification of the experiments in C, displayed as number (no.) of Syt and Brp puncta per neuron, normalised to wildtype (the assessed numbers of neurons are indicated in each bar, ***PMW<0.001; **PMW<0.01; *PMW<0.05,). Scale bar: 10 µm. A statistics summary of the data shown here is available in Figure 1—source data 1.
-
Figure 1—source data 1
Summary of the statistics from Figure 1B and C.
- https://doi.org/10.7554/eLife.14694.004
Figure 1—figure supplement 1
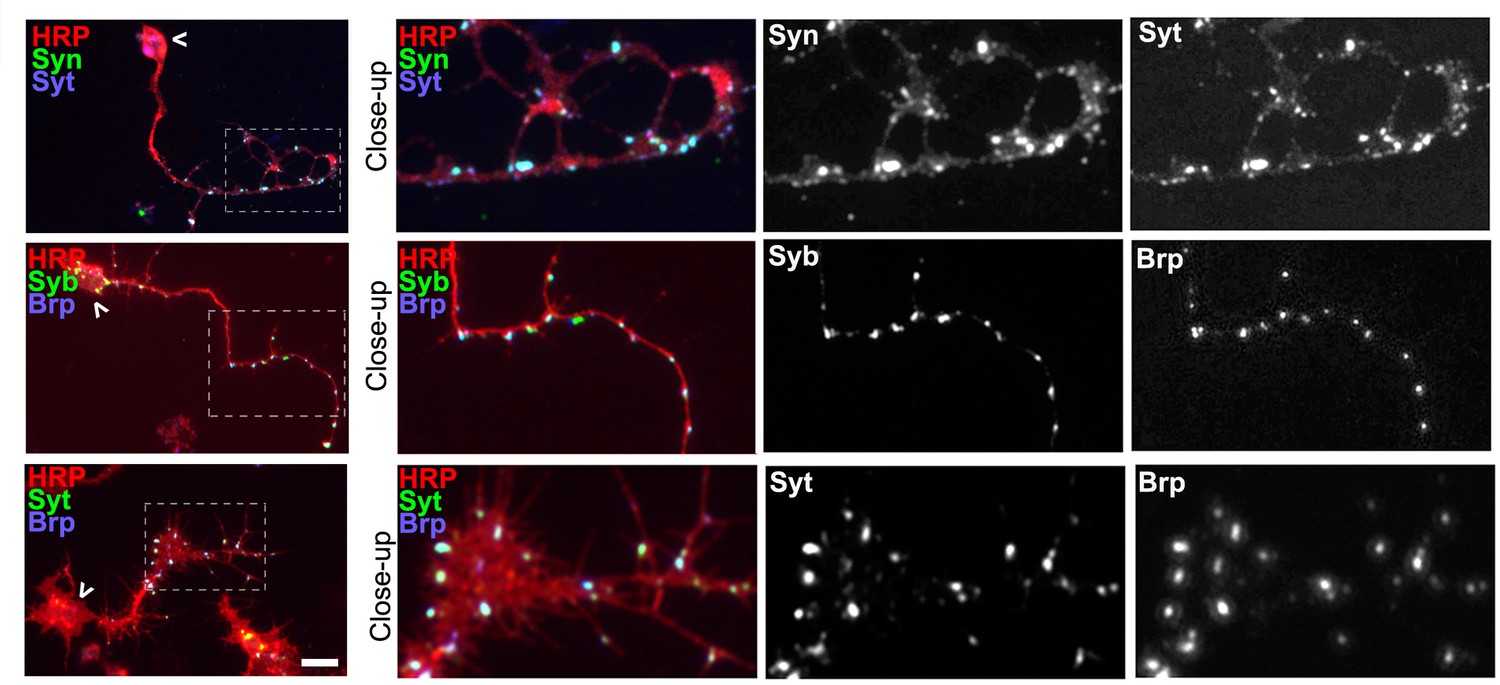
Co-localisation of presynaptic markers reveals presynaptic specialisations.
Primary Drosophila neurons at 2 DIV co-stained with antibodies against the neuronal marker HRP (red) and different presynaptic proteins (green and blue) including Synaptotagmin (Syt), Synapsin (Syn), neuronal Synaptobrevin (nSyb) or Bruchpilot (Brp); whole neurons are shown, with chevrons indicating somata and the emboxed areas indicating the distal axons shown as threefold manified close-ups; presynaptic proteins show a high degree of co-localisation in the axons, 91% co-localisation of Synapsin (Syn) and Synaptotagmin (Syt) labelled spots (sample size = 24 neurons), 84.7% of Syt and Bruchpilot (Brp) (sample size = 8 neurons), and 81% of neuronal Synaptobrevin (nSyb) with Brp (sample size = 8 neurons), suggesting that the majority of dots represent presynaptic specialisations. Scale bar: 10 µm
Figure 1—figure supplement 2
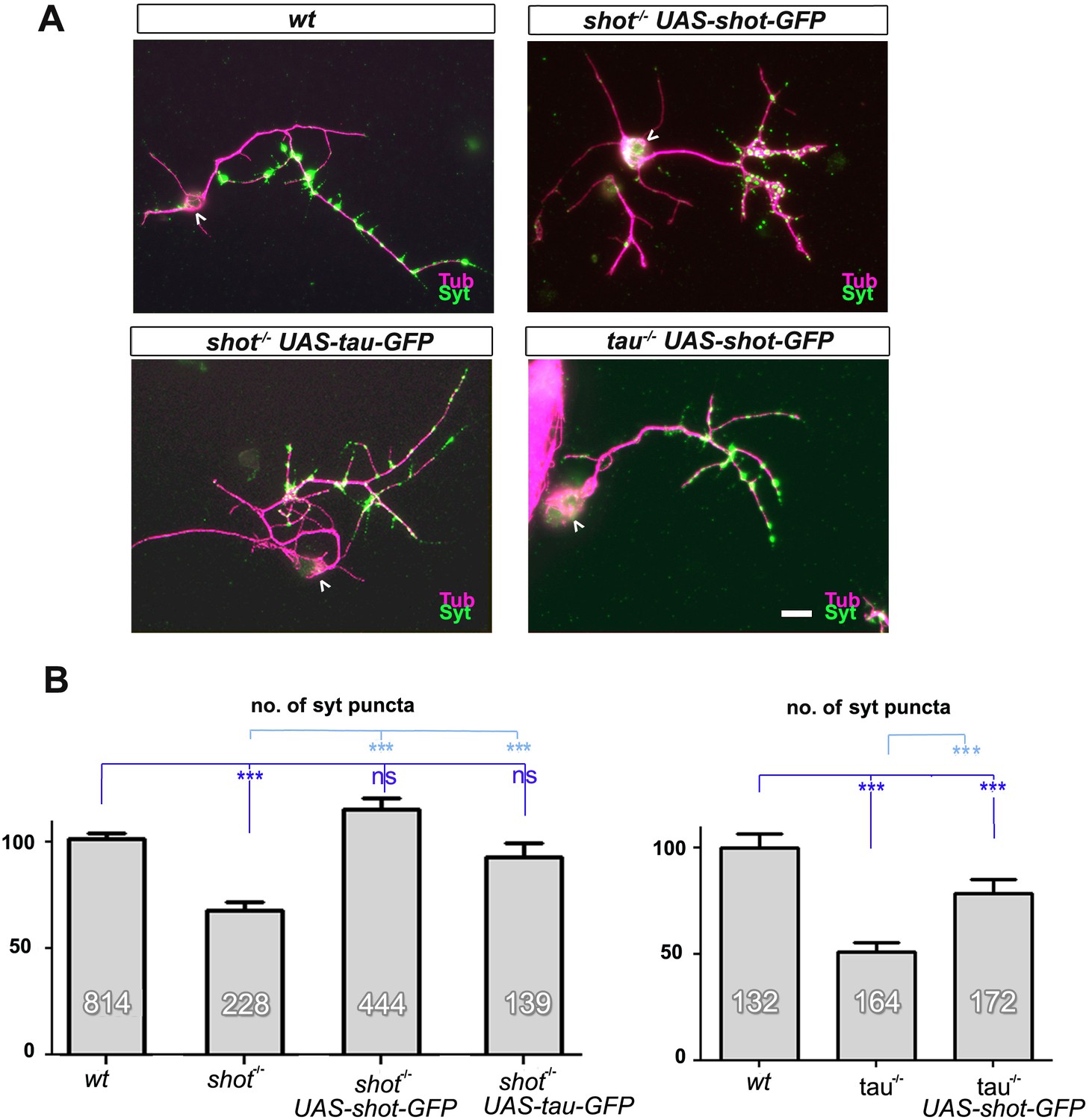
Rescue experiments with Shot and Tau demonstrate redundant roles in synapses.
(A) Primary neurons at 2 DIV obtained from embryos that were wildtype (wt), shot-/- with elav-Gal4 driven expression of UAS-shot-GFP, shot-/- with sca-Gal4 driven expression of UAS-tau-GFP, and tau-/- with elav-Gal4 driven expression of UAS-shot-GFP; neurons were stained for tubulin (Tub, magenta) and Syt (green). (B) Quantification of the experiment in A, shown as the number of Syt puncta per neuron, normalised to wildtype (the assessed numbers of neurons are indicated in each bar, ***PMW<0.001; *PMW<0.05; ns, not significant PMW>0.05). Scale bar: 10 µm. A statistics summary of the data shown here is available in Figure 1—figure supplement 2—source data 1.
-
Figure 1—figure supplement 2—source data 1
Summary of the statistics from Figure 1—figure supplement 2B.
- https://doi.org/10.7554/eLife.14694.007
Figure 2 with 1 supplement
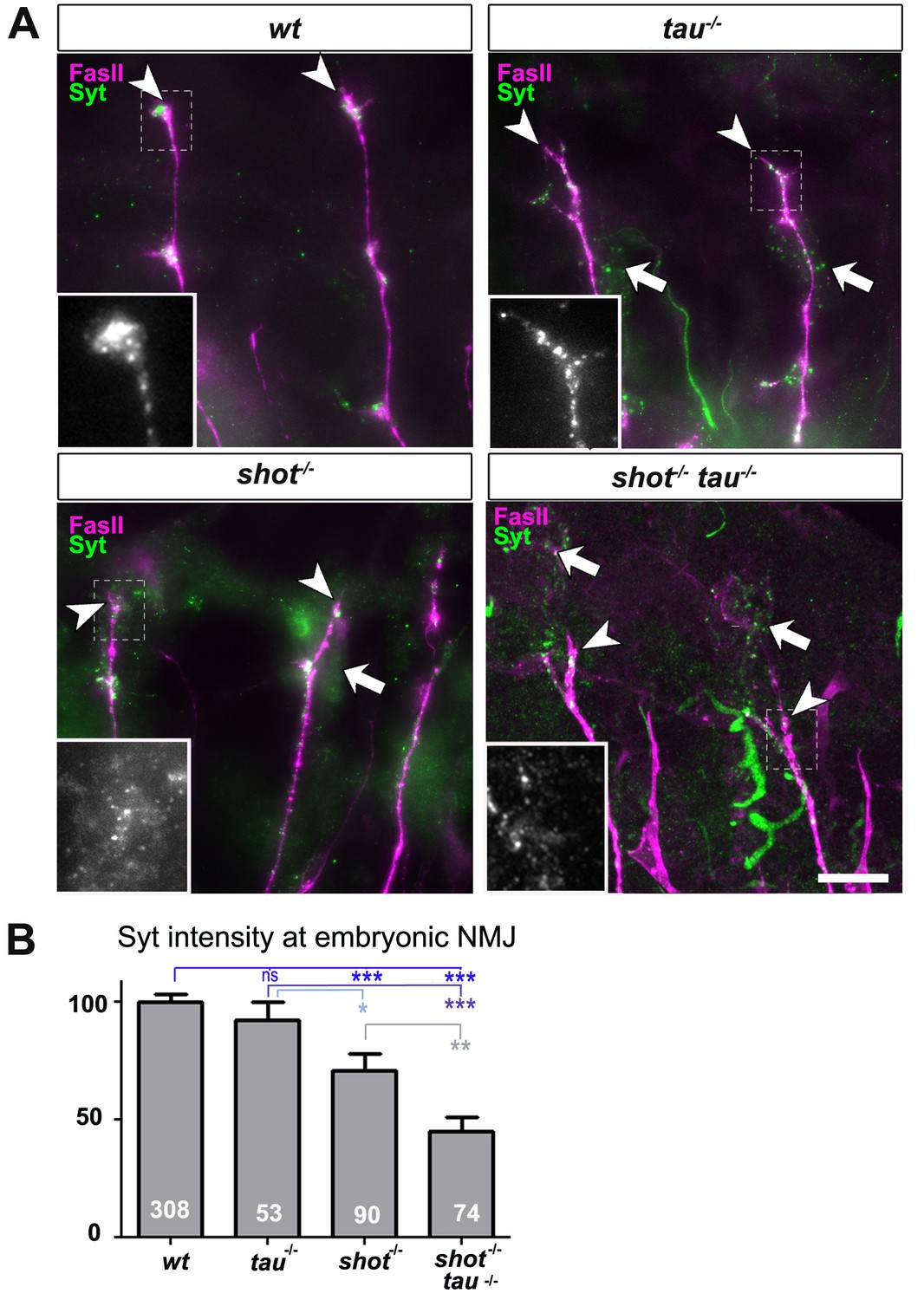
Tau and Shot regulate the localisation of presynaptic proteins at the embryonic NMJ in vivo.
(A) Images show the dorsal segment of inter-segmental motornerves (Landgraf et al., 2003) in stage 16 embryos that were wildtype (wt), tauMR22 (tau-/-), shot3 (shot-/-), and shot3 tauMR22 (shot-/- tau-/-), stained with antibodies against Syt (green) and the motorneuron-specific cell membrane protein Fasciclin II (FasII, magenta). Arrowheads depict the distal end of the motoraxons where the nascent NMJs are forming; boxed areas are displayed as enlarged insets showing anti-Syt staining only. Note that cell bodies of sensory neurons contain visible levels of Syt in the mutant (white arrows) but not in wildtype neurons (open arrow). (B) Quantification of the experiments in A, shown as the average intensity of Syt at the nerve ending normalised to wildtype (the sample number of NMJs is indicated in each bar, ***PMW<0.001; **PMW<0.01; *PMW<0.05; ns, not significant PMW>0.05). Scale bars: 10 µm. A statistics summary of the data shown here is available in Figure 2—source data 1.
-
Figure 2—source data 1
Summary of the statistics from Figure 2B.
- https://doi.org/10.7554/eLife.14694.009
Figure 2—figure supplement 1
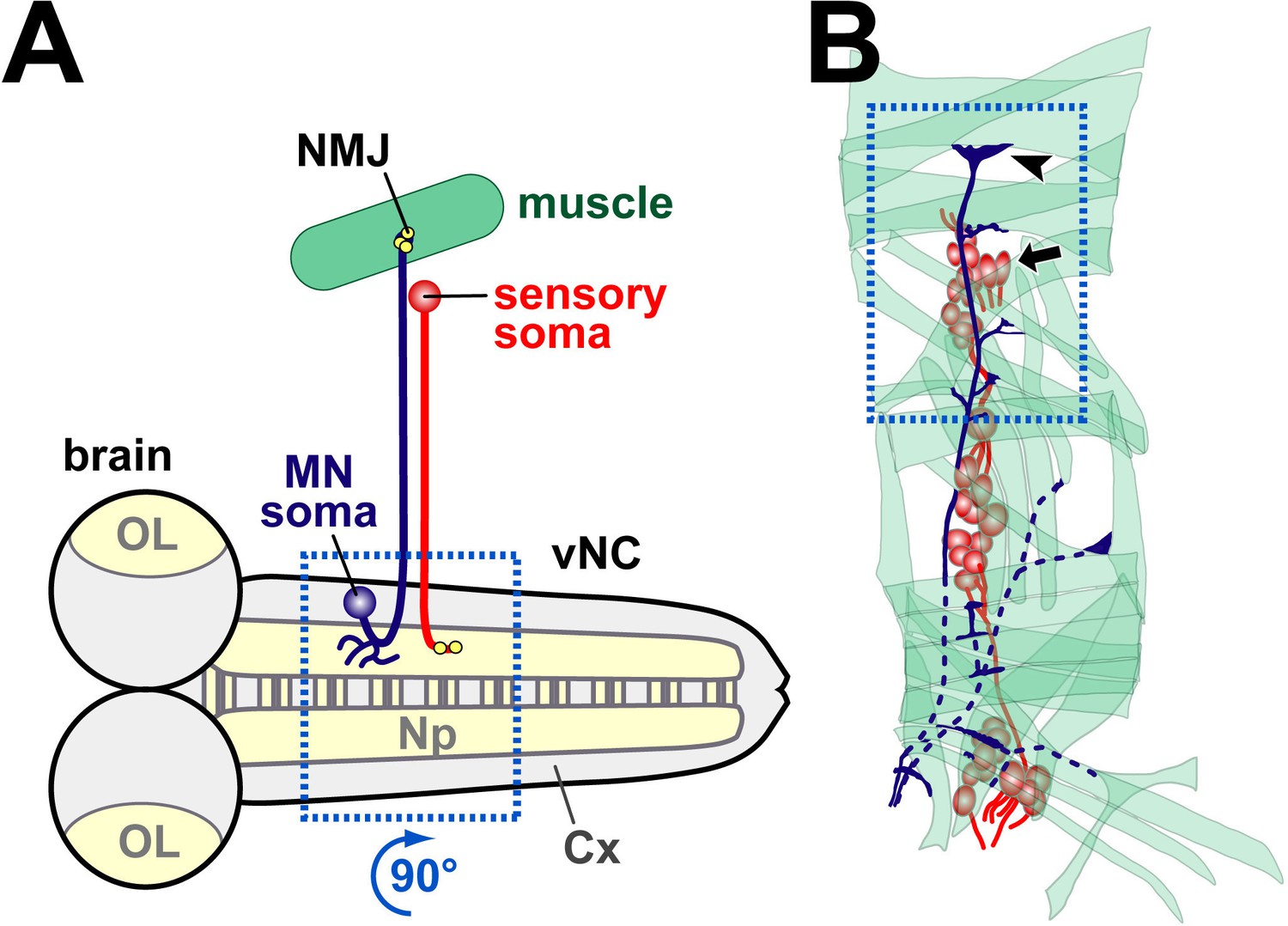
Schematic drawings of embryonic tissues analysed in this study.
(A) Schematic horizontal view of the embryonic nervous system (brain; OL, optic lobe; vNC, ventral nerve cord) showing an exemplary motorneuron (dark blue), muscle (green) and sensory neuron (red). Note that cell bodies in the nervous system lie in the cortex (Cx), i.e. outside the synaptic area (Np, neuropile), and that sensory neurons are positioned in the periphery, often adjacent to synaptic endings of motorneurons (NMJs, neuromuscular junctions). Synapses are shown as yellow dots. The stippled blue frame shows the area (rotated 90 degrees clockwise) shown in Figures 5C,D, 8F. (B) More detailed representation of the periphery with muscles, motorneuronal projections and sensory neurons (same colours as in A). The stippled blue frame emboxes the area shown in Figures 2, 4E, 5C,E, 6D,F, 7 and 8C, including an arrow head pointing at the most dorsal NMJ and arrow indicating somata of sensory neurons.
Figure 3 with 4 supplements
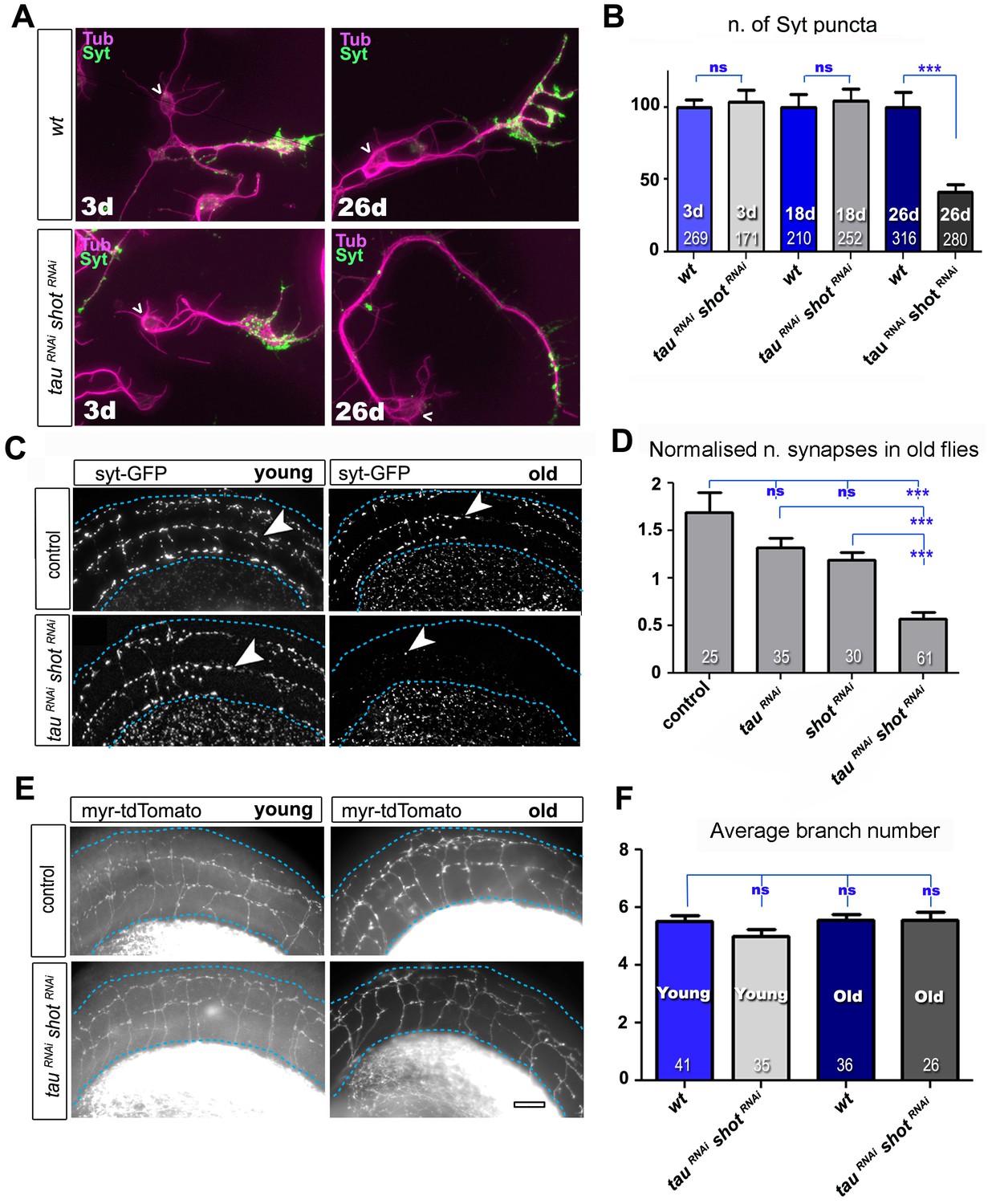
Tau and Shot are required for the maintenance of synaptic markers in cultured neurons and the ageing adult fly brain.
(A) Primary neurons at 3 DIV and 26 DIV cultured from embryos that were wildtype or jointly expressing UAS-tauRNAi and UAS-shotRNAi in all neurons driven by the pan-neuronal driver elav-Gal4 (tauRNAi shotRNAi). Neurons are stained with anti-tubulin and anti-Syt; at 26 DIV, tauRNAi shotRNAi neurons display a reduction in the number of Syt puncta when compared to wildtype. (B) Quantification of the experiments in A, shown as the number of Syt puncta per neuron at 3 DIV, 18 DIV and 26 DIV, normalised to wildtype controls (the number of assessed neurons is indicated in each bar; ***PMW<0.001; ns, not significant PMW>0.05). (C) A region of Drosophila adult brains including the medulla (delimited by dashed lines) where Syt-GFP is expressed in dorsal cluster neurons using atonal-Gal4, in the absence (control) or together with tauRNAi and shotRNAi (tauRNAi shotRNAi). Brains are stained with anti-GFP at 2–5 days (young) and 24–29 days (old) after eclosion. Note that GFP-labelled synapses (arrowheads) are decreased in old brains upon shot and tau knock-down. (D) Quantification of the experiments in C, showing the normalised number of Syt-GFP-labelled puncta in old specimen per mean number of puncta in young specimens for the following phenotypes: ato-Gal4 UAS-syt-GFP alone (control), co-expressing UAS-tauRNAi (tauRNAi), UAS-shotRNAi(shotRNAi), or both knock-down constructs (tauRNAi shotRNAi; the number of analysed brains is indicated in each bar, ***PMW<0.001; ns, not significant PMW>0.05). (E) Brain regions as in C, of animals expressing the membrane marker myr-tdTomato driven by ato-Gal4 revealing the morphology of the projections of dorsal cluster neurons within the medulla ; brains were from adults at 2–5 days (young) and 24–29 days (old) after eclosure, expressing myr-tdTomato either alone (control) or together with tauRNAi and shotRNAi(tauRNAi shotRNAi). (F) Quantification of the experiments in E, displayed as number of branches per axon projecting into the medulla (the number of axons analysed is indicated in each bar; ns, not significant PMW>0.05). Scale bar: 10 µm in A and 40 µm in C and E. A statistics summary of the data shown here is available in Figure 3—source data 1.
-
Figure 3—source data 1
Summary of the statistics from Figure 3B,D,F.
- https://doi.org/10.7554/eLife.14694.012
Figure 3—figure supplement 1
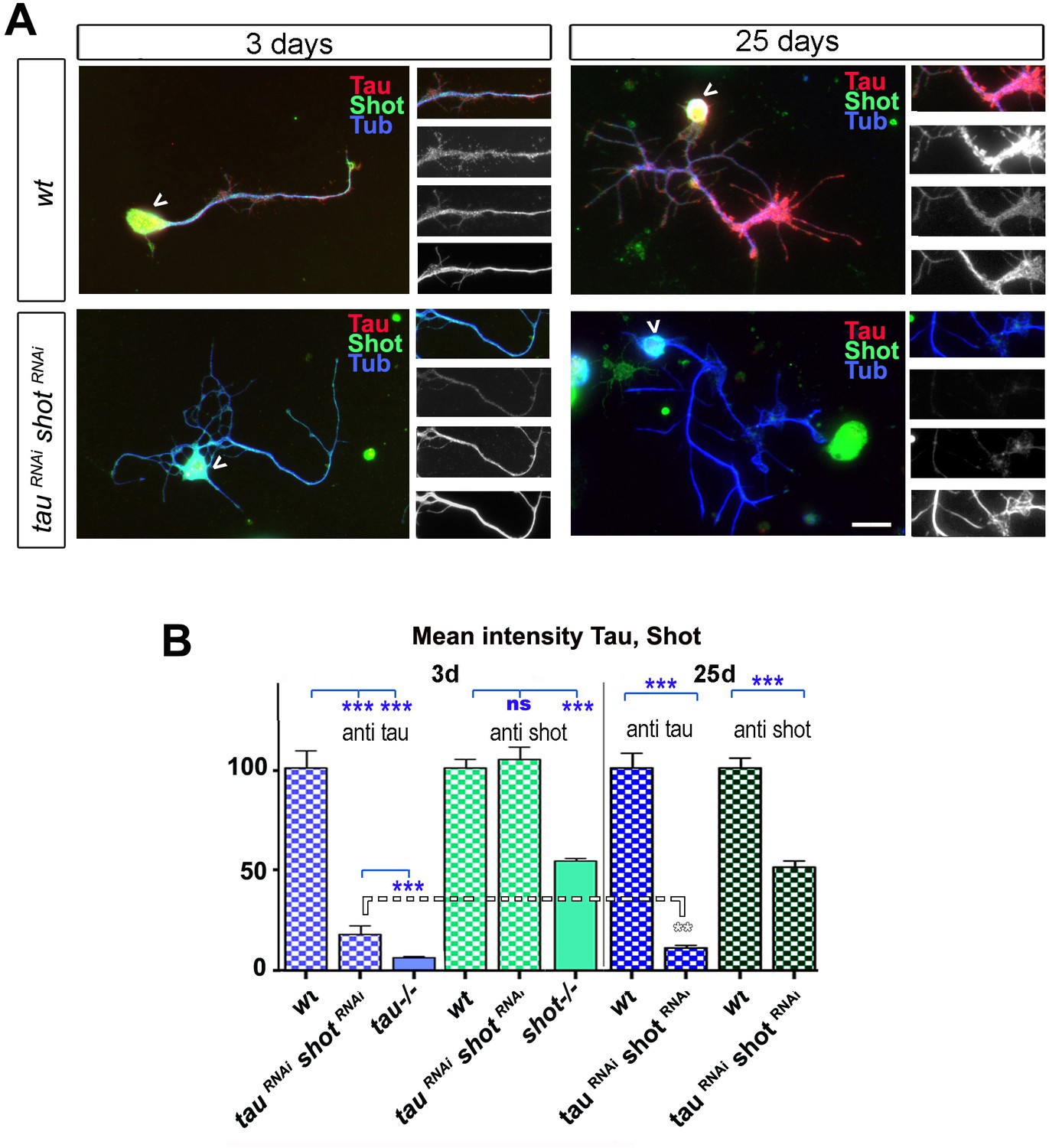
Delayed effect of RNAi mediated knock-down of Shot and Tau.
(A) Primary neurons at 3 DIV and 25 DIV cultured from embryos that were wildtype or jointly expressing UAS-tauRNAi and UAS-shotRNAiin all neurons driven by the pan-neuronal driver elav-Gal4 (tauRNAi shotRNAi). Neurons are stained with antibodies against Tau, Shot and Tubulin (red, green and blue respectively); images on the right show: a selected axon segment taken from the main image (top) followed by grey scale images of the separated channels for Tau (2nd from top), Shot (3rd from top) and Tubulin (bottom). At 25 DIV, tauRNAi shotRNAi neurons display a reduction in both Tau and Shot when compared to wildtype. (B) Quantification of the experiments in A, shown as mean intensity of Tau or Shot signal per neuron at 3 DIV and 25 DIV, normalised to wildtype controls (30–39 neurons were assessed per genotype; ***PMW<0.001; **PMW<0.01; ns, not significant PMW>0.05). Comparative data for shot3 and tauMR22 homozygous mutant neurons are given as control, indicating low Tau background staining and incomplete knock-down of Tau at 3 DIV, but high Shot background suggesting strong or complete Shot knock-down at 25 DIV. A statistics summary of the data shown here is available in Figure 3—figure supplement 1—source data 1.
-
Figure 3—figure supplement 1—source data 1
Summary of the statistics from Figure 3—figure supplement 1B.
- https://doi.org/10.7554/eLife.14694.014
Figure 3—figure supplement 2
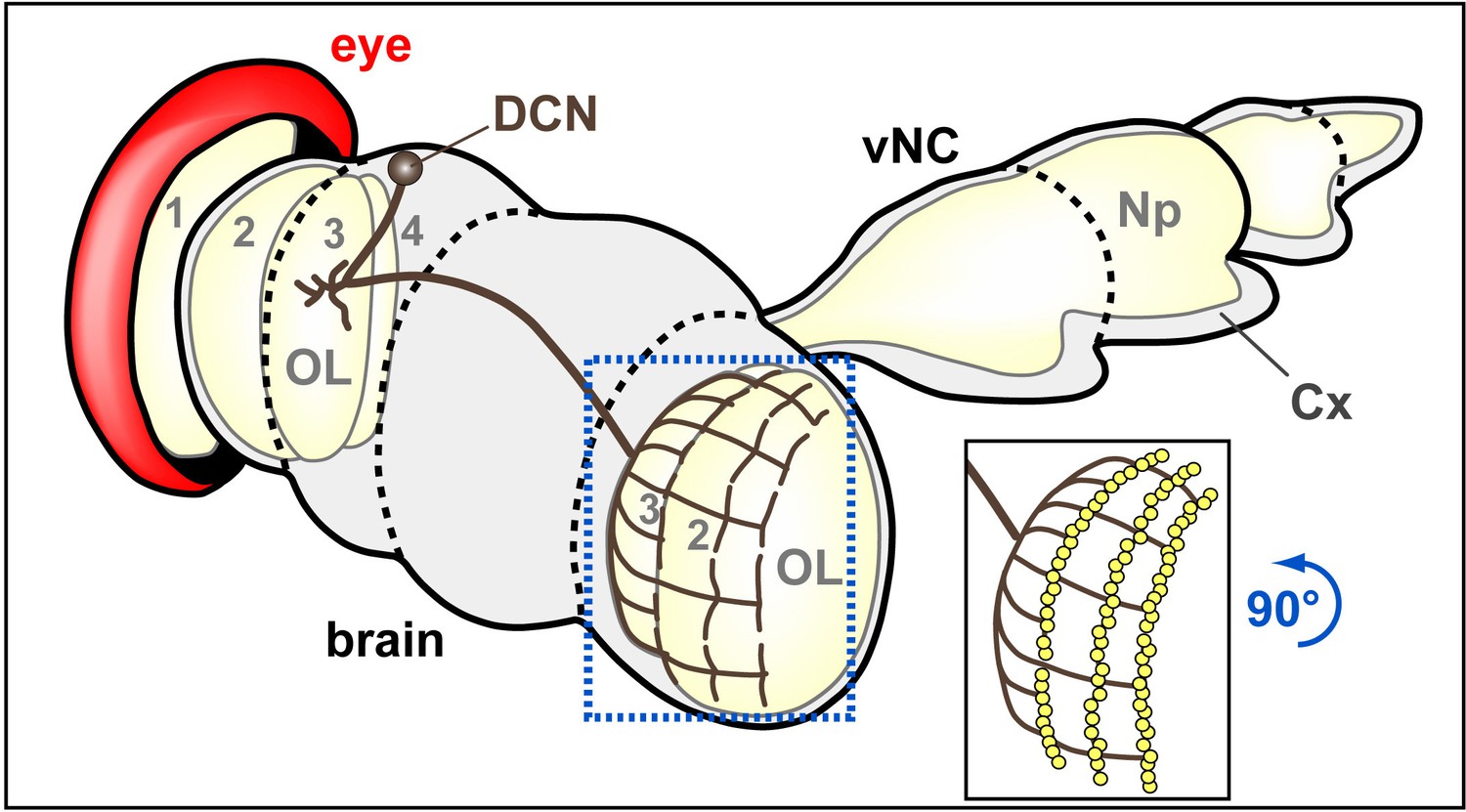
Schematic drawings of brain areas analysed in this study.
Dorso-fronto-lateral view onto a schematic adult CNS composed of the brain and ventral nerve cord (vNC). Beige areas indicate some synaptic areas, in particular the ventral nerve cord neuropile (Np) and the optic lobes (OL) composed of lamina (1), medulla (2), lobula (3) and lobula plate (4). DCN neurons project to the optic lobe of the contralateral brain half where they branch out in a layered fashion. The blue stippled frame and image inset embox the area (rotated 90 degrees counterclockwise) shown in Figures 3E and 8E without and with synaptic markers (yellow dots), respectively.
Figure 3—figure supplement 3
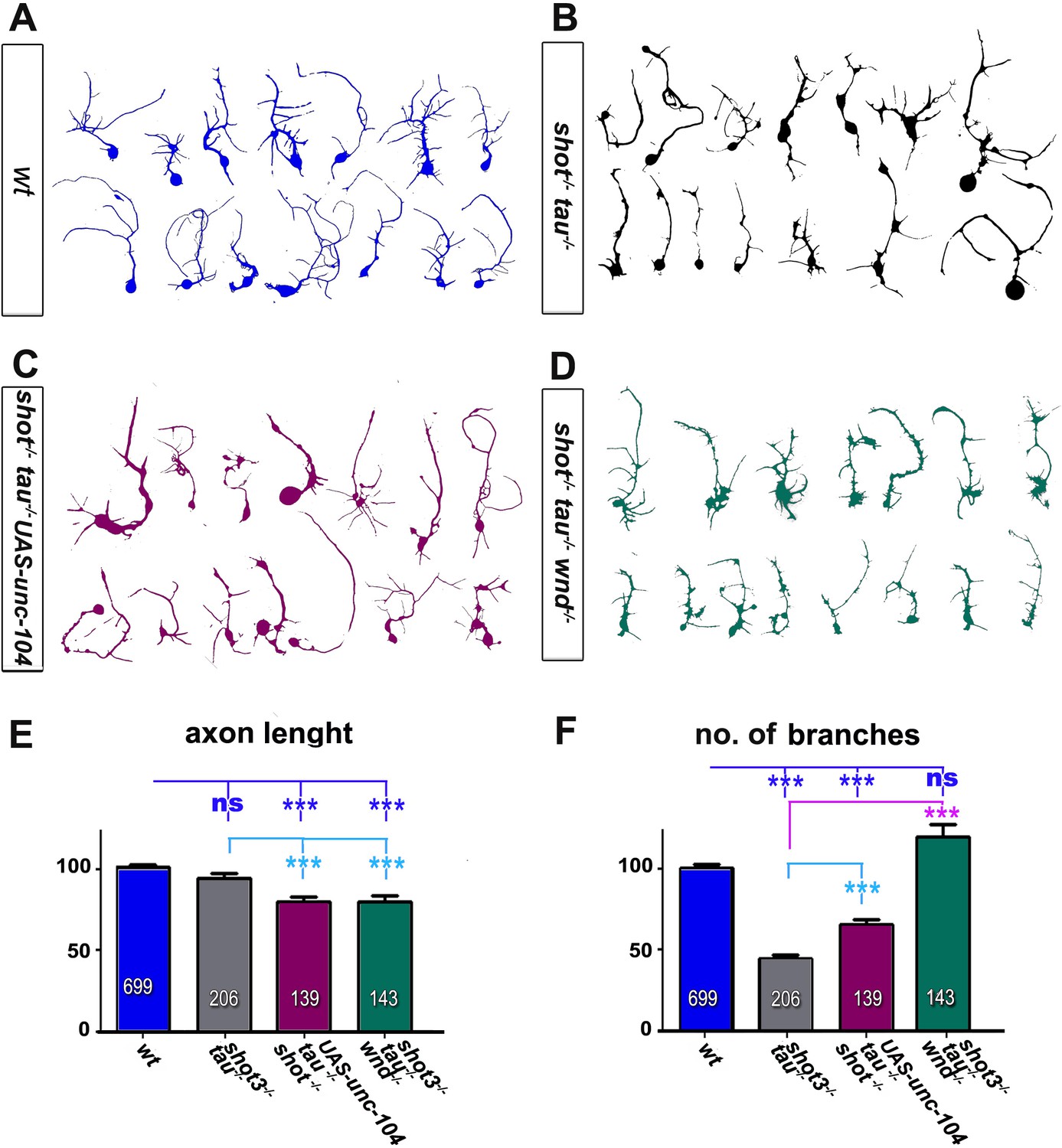
Loss of function mutations in shot and tau induce morphological changes.
(A–D) Representative examples of the shape of primary neurons at 2 DIV obtained from embryos that were wildtype (wt, A), shot3 tauMR22 (shot-/- tau-/- B), shot-/- tau-/- with elav-Gal4 driven expression of UAS-unc-104 (shot-/- tau-/- UAS-unc-104, C) and shot3 tauMR22 wnd2 (shot-/- tau-/- wnd-/- D). (E–F) Quantification of morphological parameters of experiment in (A–D), including the length of axons (E) and the number of branches (F), (the assessed numbers of neurons are indicated in each bar, ***PMW<0.001; ns, not significant PMW>0.05). Note that wnd2 (wnd-/-) restored the number of branches in shot-tau mutant neurons, suggesting that JNK not only mediates synapse regulation but also morphogenetic processes downstream of the Shot-Tau deficiency. A statistics summary of the data shown here is available in Figure 3—figure supplement 3—source data 1.
Figure 3—figure supplement 4
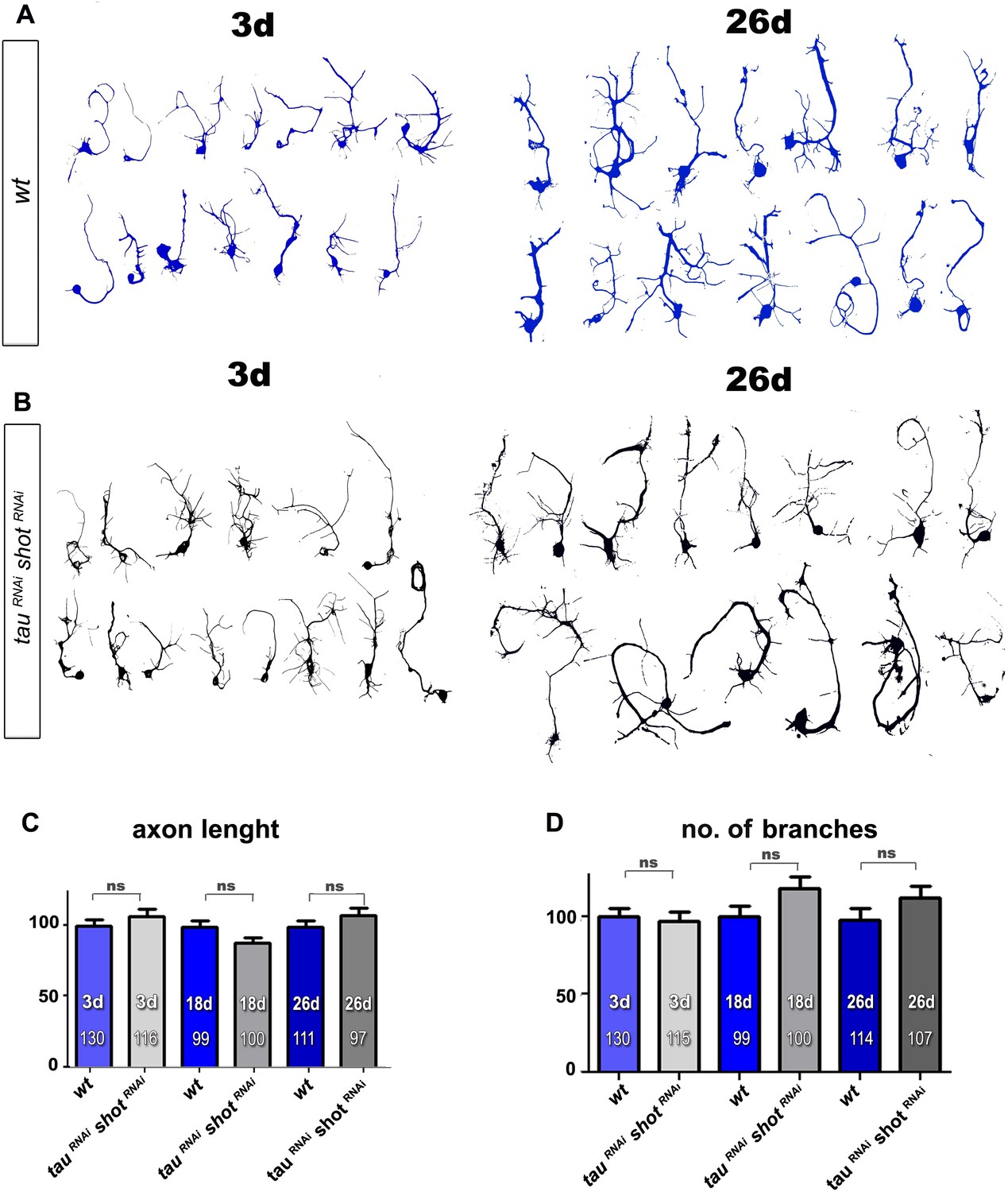
RNAi-mediated knock-down of Shot and Tau has no effect on axonal length and branch number.
(A–B) Representative examples of the shape of primary neurons at 3, 18 and 26 DIV obtained from embryos that were wildtype (wt, A) or jointly expressing UAS-tauRNAi and UAS-shotRNAi in all neurons driven by the pan-neuronal driver elav-Gal4 (tauRNAi shotRNAi, B). (C–D) Quantification of morphological parameters of experiment in (A–B), including the length of axons (C) and the number of branches (D), (the assessed numbers of neurons are indicated in each bar, ns, not significant PMW>0.05). A statistics summary of the data shown here is available in Figure 3—figure supplement 4—source data 1.
-
Figure 3—figure supplement 4—source data 1
Summary of the statistics from Figure 1—figure supplement 3C and D.
- https://doi.org/10.7554/eLife.14694.018
Figure 4
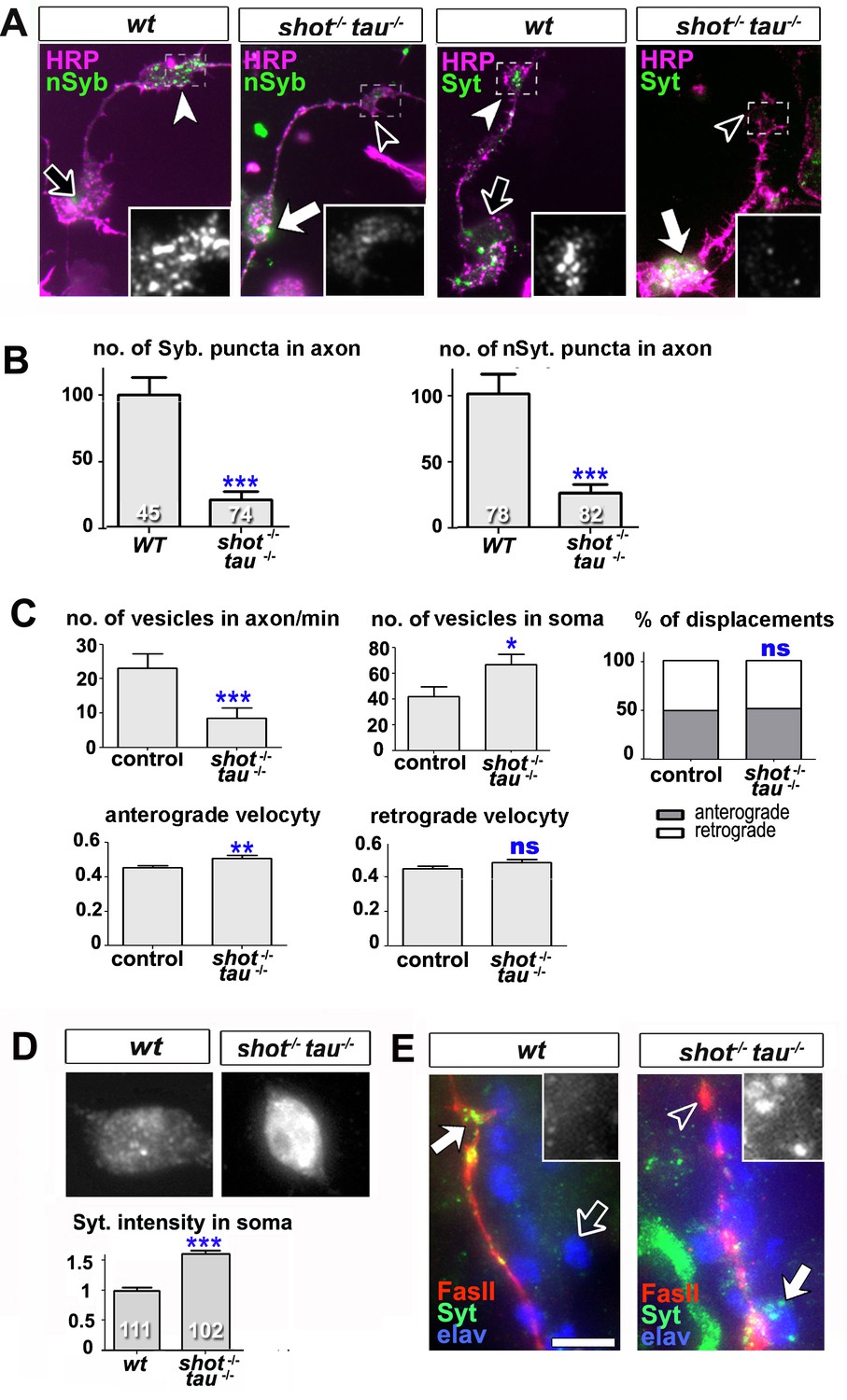
Intracellular transport of synaptic proteins is defective in shot-tau mutant neurons.
(A) Primary Drosophila neurons at 8HIV, obtained from embryos that were wildtype (wt) and shot-tau (shot-/- tau-/-) stained with antibodies against pan-neuronal HRP (magenta), Syt (green) or nSyb (green); nSyb and Syt are reduced in the growth cones (open versus white arrow heads) but enriched in cell bodies (open versus white arrows) of shot-tau mutant neurons. (B) Quantification of the experiments from A, given as the number of nSyb or Syt puncta in axons and growth cones; the number of analysed neurons is given in the bars (***PMW<0.001). (C) Quantification of various transport parameters generated from live movies of axons of wildtype or shot-tau mutant neurons (shot-/- tau-/-) at 8 HIV with elav-Gal4 driven expression of UAS-Syt-GFP. Axonal anterograde and retrograde velocities show only subtle or no alteration in the axons of shot-tau neurons. On the contrary, the numbers of vesicles in axons of shot-tau neurons are sharply decreased and increased in the somata (**PMW<0.01; *PMW<0.05; ns, not significant PMW>0.05). (D) Magnified views of the somata from primary Drosophila neurons at 2 DIV, obtained from wildtype (wt) and shot-tau mutant embryos (shot-/- tau-/-), co-stained with antibodies against Syt. To document the protein content within cell bodies, several z stacks per neuron were obtained and fused as maximal projection; the cell bodies show higher levels of Syt in shot-tau mutant neurons as compared to wildtype (number of assessed cells is indicated in the bars, average staining intensity normalised to wildtype; ***PMW<0.001). (E) The dorsal peripheral nervous system (PNS) of wildtype andshot-tau embryos at late stage 16 (stages according to) (Campos-Ortega and Hartenstein, 1997) stained for Syt (green), FasII (red) and the pan-neuronal nuclear marker Elav (blue). The nascent NMJ at the tip of the inter-segmental motornerve (red) in wildtype contains high levels of Syt (white arrow) whereas the somata of sensory neurons (blue and grey in insets) contain low levels (open arrow); in shot-tau homozygous embryos the somata of sensory neurons have high levels of Syt (arrow and inset), whereas there is only little staining at the nerve tip (open arrowhead). Scale bars: 10 μm in A, 5 μm in D and 5 μm in E. A statistics summary of the data shown here is available in Figure 4—source data 1.
-
Figure 4—source data 1
Summary of the statistics from Figure 4B–D.
- https://doi.org/10.7554/eLife.14694.020
Figure 5 with 1 supplement
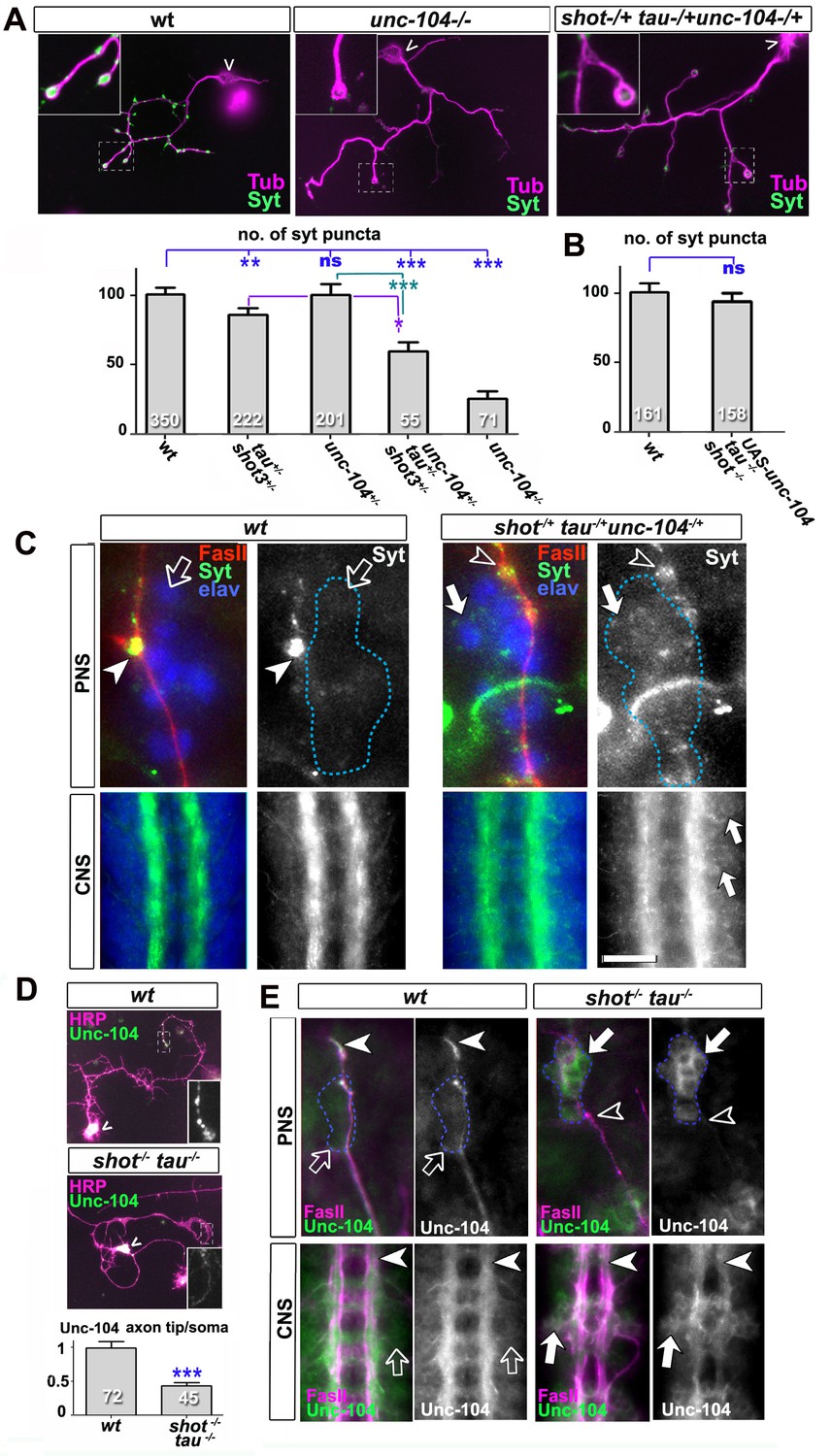
Defects in kinesin-3 function mediate synaptic deficits in shot-tau mutant neurons.
Shot and Tau interact with Unc-104 and regulate its subcellular distribution. (A) Primary Drosophila neurons at 2 DIV, obtained from embryos which were wildtype, homozygous for unc104imac170 (unc104-/-), or triple-heterozygous for shot3 tauMR22 unc104imac170 mutations (shot+/- tau+/-unc104+/-), co-stained with antibodies against tubulin (Tub, magenta) and Syt (green). The graph shows the quantification of the data including also unc104-/+ and shot-/- tau-/- controls. (B) Quantification of Syt puncta in two day old neurons, obtained from embryos that were wildtype or shot-/- tau-/- with elav-Gal4 driven expression of UAS-unc-104 (compare Figure 1D). (C) The dorsal peripheral nervous system (PNS) and the central nervous system (CNS) of wildtype and shot3 tauMR22 unc104imac170 triple heterozygous embryos at late stage 16 (stages according to Campos-Ortega and Hartenstein, 1997) stained for Syt (green), FasII (red in upper panel) and the pan-neuronal nuclear marker Elav (blue); for illustration of the imaged tissue see Figure 2-figure supplement 1. The nascent NMJ at the tip of the inter-segmental motornerve (red in upper panels) in wildtype contains high levels of Syt (arrowheads) whereas the somata of sensory neurons (blue; demarcated by dashed lines) contain low levels (open arrows); in shot3 tauMR22 unc104imac170 triple heterozygous embryos the somata of sensory neurons have high levels of Syt (arrows), whereas there is only little staining at the nerve tip (open arrowhead). In the ventral nerve cord of wildtype (lower panels), Syt is confined to the neuropile (synapse containing CNS compartment; arrowheads) and excluded from the cortex (compartment with the cell bodies of inter- and motorneurons); in the ventral nerve cord of shot3 tauMR22 unc104imac170 triple heterozygous embryos, there are segmental groups of cell bodies displaying higher Syt levels (arrows). (D) Primary Drosophila neurons at 2 DIV, obtained from wildtype (wt) and tau-shot mutant embryos, stained with antibodies against pan-neuronal HRP (magenta) and Unc-104 (green); Unc-104 in distal axon segments (emboxed and magnified in insets) is enriched in wildtype but much weaker in shot-tau mutant neurons (chevrons indicate neuronal somata). Data were quantified as average intensity of Unc-104 at the distal end of the axon divided by the average intensity at the soma. (E) Upper and lower panels show the same locations of late stage 16 embryos as shown in C, but taken from wildtype and shot-tau mutant embryos, stained for FasII (magenta) and Unc-104 (green). Note the stark decrease of Unc-104 at the end of motor nerves (open versus white arrow heads) and the unusual accumulations of Unc-104 in the cell bodies of sensory neurons as well as in the CNS cortex in shot-tau embryos (open versus white arrows). In all graphs, the number of assessed neurons is indicated in each bar; ***PMW<0.001; *PMW<0.05; ns, not significant PMW>0.05; scale bars: 18 μm in A, 5 μm in C/PNS, 35 μm in C/CNS, 15 μm in D and E/PNS, 35 μm in E/CNS. A statistics summary of the data shown here is available in Figure 5—source data 1.
-
Figure 5—source data 1
Summary of the statistics from Figure 5.
- https://doi.org/10.7554/eLife.14694.022
Figure 5—figure supplement 1
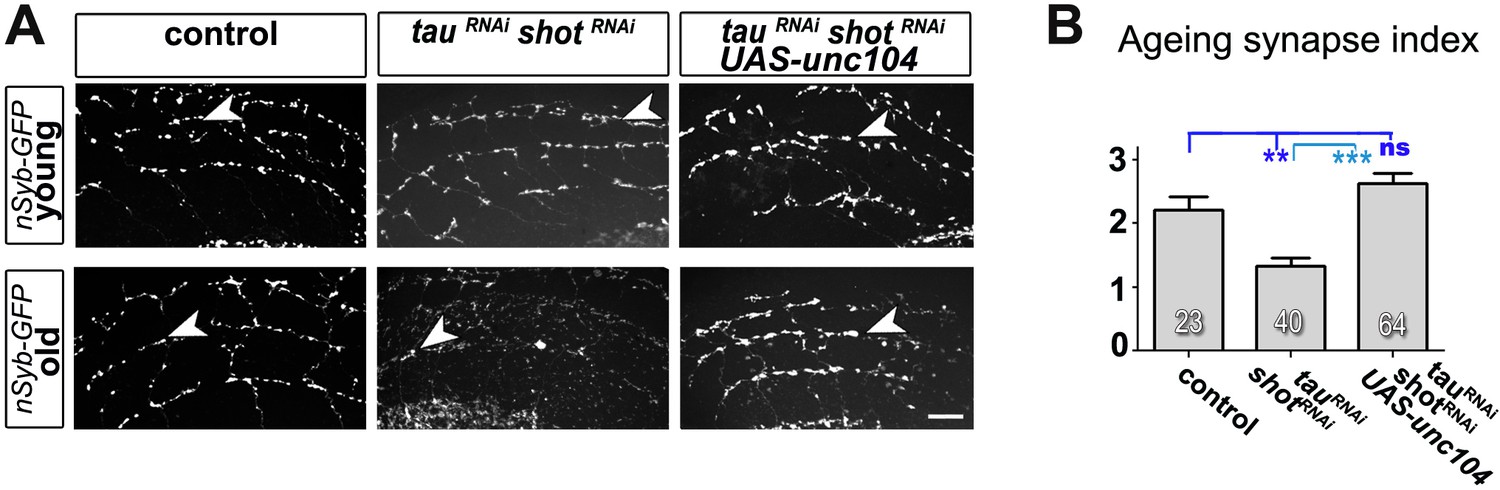
Expression of Unc-104 rescues synaptic defects in aged adult brains.
(A) A region of Drosophila adult brains including the medulla (see illustration in Figure 3—figure supplement 2); UAS-nSyb-GFP is expressed in dorsal cluster neurons using atonal-Gal4, either alone (control), together with tauRNAi and shotRNAi(tauRNAi shotRNAi) or together with tauRNAi, shotRNAi and UAS-unc-104 (tauRNAi shotRNAi UAS-unc-104). Brains are shown at 2–6 days (young) and 26–30 days after eclosion (old); GFP-labelled synapses are decreased in old brains with shot-tau knock-down when compared to controls, and this effect is rescued by the expression of Unc-104. (B) Quantification of experiments in A, shown as number of GFP-labelled synapses in old specimen per mean number of GFP-labelled synapses in young specimens of the respective genotype (number of analysed brains is indicated in the bars; ***PMW<0.001; **PMW<0.01; ns, not significant PMW>0.05). Scale bars: 5 μm in A. A statistics summary of the data shown here is available in Figure 5—figure supplement 1—source data 1.
-
Figure 5—figure supplement 1—source data 1
Summary of the statistics from Figure 5—figure supplement 1B.
- https://doi.org/10.7554/eLife.14694.024
Figure 6 with 1 supplement
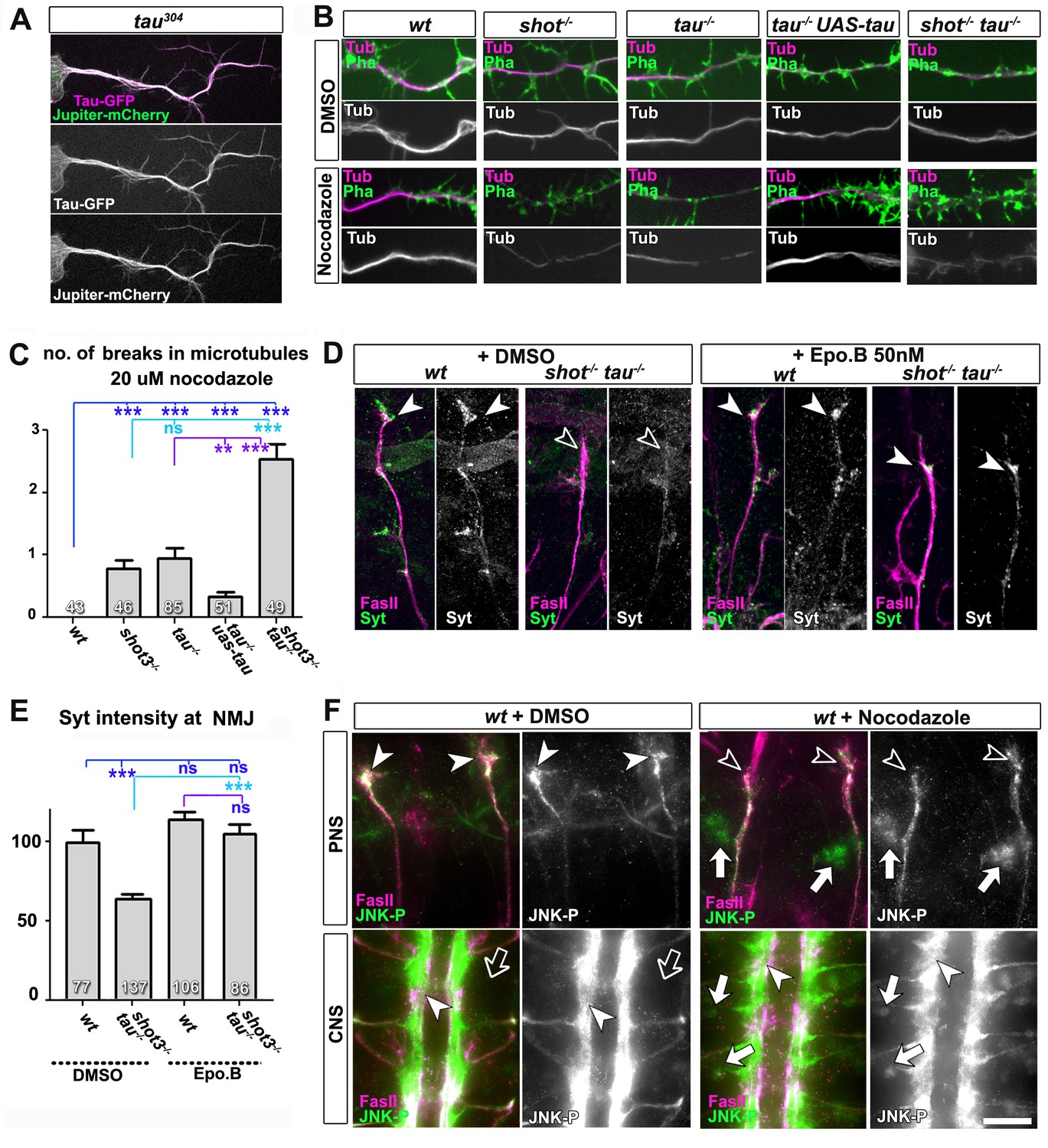
Microtubule instability mediates aberrant JNK signalling and synaptic defects.
(A) Live imaging of Drosophila neurons at 2 DIV, obtained from embryos carrying tau304 (a protein trap line where the endogenous tau gene is genomically tagged with GFP) and the microtubule binding protein Jupiter-Cherry. Endogenous Tau (in magenta) is observed in a pattern reminiscent of microtubules, and colocalises with Jupiter (shown in green). (B) Axons of Drosophila neurons at 6 HIV with the following genotypes: wildtype (wt), shot3 (shot-/-), tauMR22 (tau-/-), tau rescue (tau-/- UAS-tau) and shot-tau (shot-/- tau-/-). Neurons were treated for 2.5 hrs with vehicle (DMSO) or 20 µM nocodazole, fixed and stained with anti-Tubulin (Tub, magenta and white) and phalloidin (Pha, green). Only shot3, tauMR22, and shot-tau double mutant displayed gaps in their axonal microtubule bundles upon nocodazole treatment, but not wildtype and tau mutant embryos with Tau rescue. (C) Quantification of the experiments in B, indicated as the number of breaks in the microtubule staining per axon (number of analysed neurons is indicated in bars; ***PMW<0.001; **PMW<0.01; ns, not significant PMW>0.05). (D) Embryonic motoraxons of wildtype and shot-tau embryos at late stage 16 treated with vehicle (DMSO) or 50 nM of the microtubule stabilising drug epothilone B for 3 hr and stained with FasII (magenta) and Syt (green); in wildtype, the nascent NMJ at the nerve tip contains high levels of Syt (arrowheads); in shot-tau embryos there is only little Syt staining at the nerve tip (open arrowhead). Treatment of shot-tau embryos with 50nM epothilone B increases the levels of Syt at the tip of motornerves (arrowheads). (E) Quantification of the experiments shown in D, measured as the average intensity of Syt at nascent NMJs and normalised to wildtype (number or assessed NMJ is indicated in bars; ***PMW<0.001; ns, not significant PMW >0.05). (F) Upper (PNS) and lower (CNS) panels show the same locations of late stage 16 wildtype embryos as shown in Figure 5C, stained for FasII (magenta) and activated phospho-JNK (JNK-P); treatment with 100 µm nocodazole for 2 hrs induced a relocation of JNK-P from nascent NMJs (open versus white arrow heads) to cell bodies of sensory neurons and in the CNS cortex (white versus open arrows). Scale bar: 5 μm in A, 4 μm in B, 10 μm in E, 15 μm in D/PNS and 35 μm in D/CNS. A statistics summary of the data shown here is available in Figure 6—source data 1.
-
Figure 6—source data 1
Summary of the statistics from Figure 6C and E.
- https://doi.org/10.7554/eLife.14694.026
Figure 6—figure supplement 1
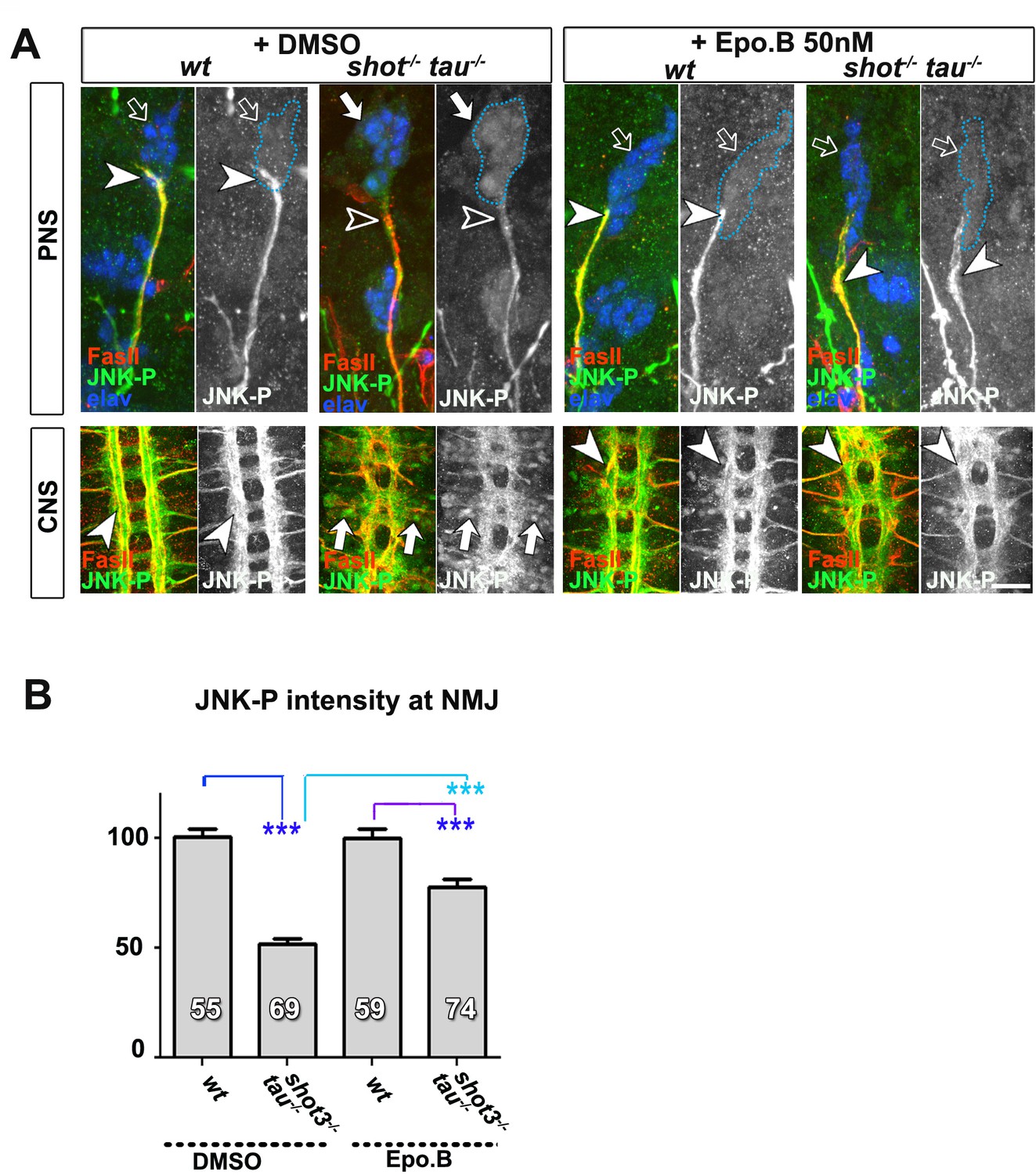
Treatment of shot-tau mutant neurons with epothilone B, increases the localisation of JNK-P at axonal tips.
(A) Embryonic motoraxons of wildtype and shot-tau embryos at late stage 16 treated with vehicle (DMSO) or 50 nM of the microtubule stabilising drug epothilone B for 3 hr and stained with FasII (magenta) and JNK-P (green); In wildtype, JNK-P is high at nerve endings (white arrowheads) and below detection levels in cell bodies of sensory neurons and in the CNS cortex (open arrows). This pattern is inverted in shot-tau embryos where JNK-P levels are low at nerve tips (open arrowhead) and high in cell bodies of sensory neurons and in the CNS cortex (white arrows). Treatment of shot-tau embryos with 50 nM epothilone B increases the levels of JNK-P at the tip of motornerves (white arrowheads). (B) Quantification of the experiments shown in A, measured as the average intensity of JNK-P at nascent NMJs and normalised to wildtype (number of assessed NMJ is indicated in bars; ***PMW<0.001). Scale bars: 15 μm in PNS panels and 35 μm in CNS panels. A statistics summary of the data shown here is available in Figure 6 —figure supplement 1—source data 1.
-
Figure 6—figure supplement 1—source data 1
Summary of the statistics from Figure 6—figure supplement 1B.
- https://doi.org/10.7554/eLife.14694.028
Figure 7
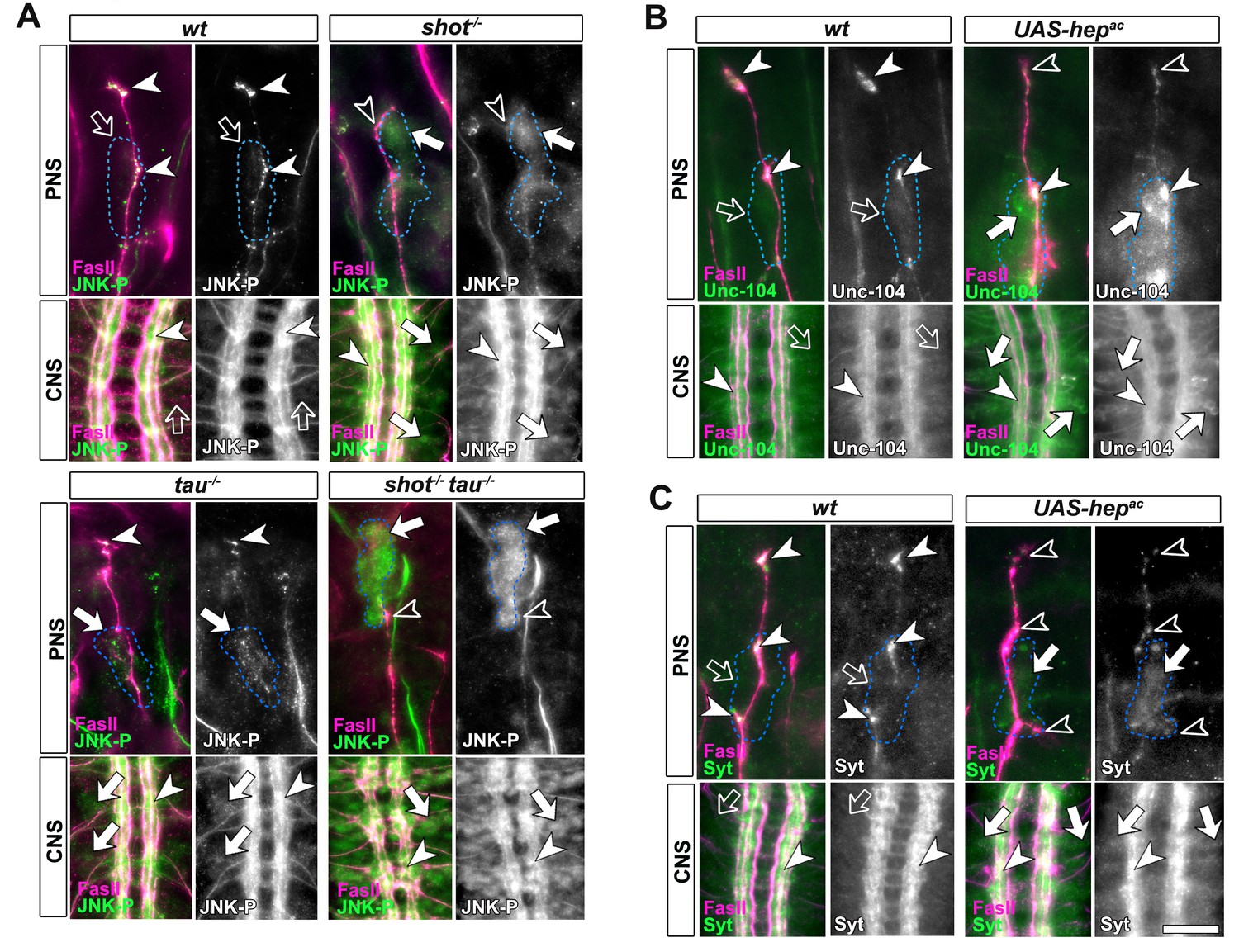
Activated JNK correlates with the subcellular localisation of Unc-104 and Syt.
Upper (PNS) and lower (CNS) panels in A-C show the same locations of late stage 16 embryos as shown in Figure 5E, but embryos are of different genotypes and stained with different antibodies, as indicated; genotypes: wildtype (wt), shot3 (shot-/-), tauMR22 (tau-/-), shot-tau (shot-/- tau-/-), elav-Gal4 driven expression of UAS-hepac (UAS-hepac); used antibodies detect FasII (magenta), Syt (green), Unc-104 (green), activated phospho-JNK (JNK-P). (A) In wildtype, JNK-P is high at nerve endings (white arrow heads) and below detection levels in cell bodies of sensory neurons and in the CNS cortex (open arrows); this pattern is inverted in tauMR22 and shot3 mutant embryos and even stronger in shot-tau embryos, i.e. Syt is reduced at nerve endings (open arrowheads) and upregulated in cell bodies (white arrows). (B, C) Artificial activation of JNK with neuronal expression of Hepac suppresses high levels of Unc-104 and Syt at nascent NMJs (open versus white arrow heads) and increases their levels in cell bodies (white versus open arrows). Scale bars: 15 μm in PNS panels and 35 μm in CNS panels.
Figure 8 with 2 supplements
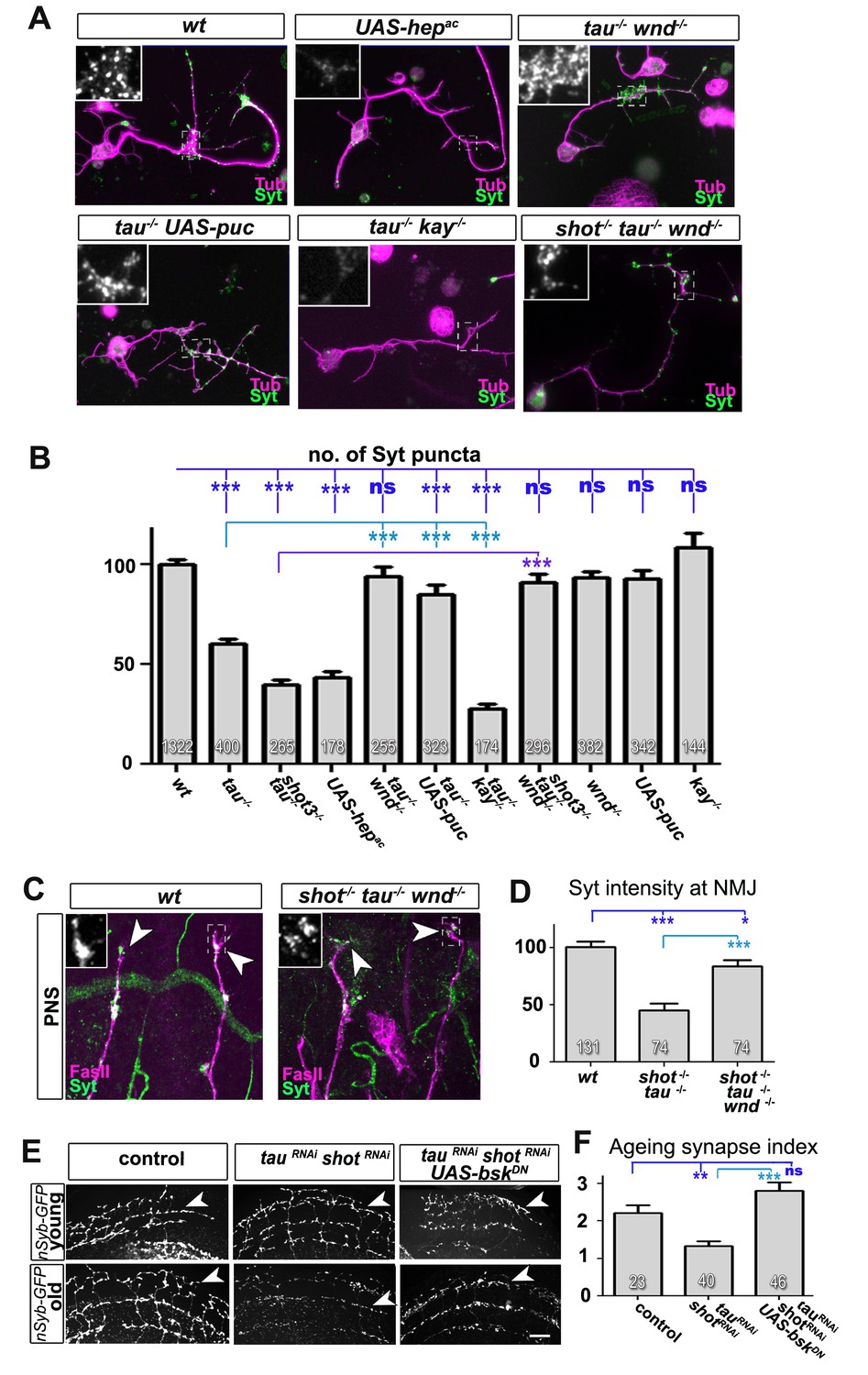
Inhibition of the JNK pathway rescues synaptic defects in shot-tau mutant neurons.
(A) Primary Drosophila neurons at 2 DIV, obtained from embryos of the following genotypes: wildtype (wt), elav-Gal4 driven expression of UAS-hepac (UAS-hepac), tauMR22 (tau-/-), wnd2 (wnd-/-), tau-/- with elav-Gal4 driven expression of UAS-puc (tau-/- UAS-puc), tauMR22 kay2 (tau-/- kay-/-) and shot3 tauMR22 wnd2 (shot-/- tau-/- wnd-/-), all stained with antibodies against Tubulin (tub, magenta) and Syt (green). Insets correspond to emboxed areas and show a magnified view of the Syt staining. (B) Quantification of experiments in A, shown as the number of Syt puncta normalised to wildtype (number of assessed neurons is shown in the bars, ***PMW<0.001; **PMW<0.01; *PMW<0.05; ns, not significant PMW>0.05). (C) Inter-segmental motornerves in the dorsal area of wildtype and shot3 mutant embryos at late stage 16, stained against FasII (magenta) and Syt (green); insets correspond to emboxed areas and show a magnified view of the most dorsal nascent NMJs stained for Syt; note the rescue of Syt localisation if Wnd is absent in tau-shot mutant background. (D) Quantification of the experiments in C, measured as the average intensity of Syt normalised to wt (number of assessed NMJs is shown in the bars; ***PMW<0.001; *PMW<0.01). (E) A region of Drosophila adult brains including the medulla; UAS-nSyb-GFP is expressed in dorsal cluster neurons using atonal-Gal4, either alone (control), together with tauRNAi and shotRNAi (tauRNAi shotRNAi) or together with tauRNAi, shotRNAi and UAS-bskDN. Brains are shown at 2–6 days (young) and 26–30 days at 29°C after eclosion (old); GFP-labelled synapses are decreased in old brains with shot-tau knock-down when compared to controls, and this effect is rescued by the expression of BskDN. (F) Quantification of experiments in E, shown as number of GFP-labelled synapses in old specimen per mean number of GFP-labelled synapses in young specimens of the respective genotype (number of analysed brains is indicated in the bars; ***PMW<0.001; **PMW<0.01). Scale bars: 5 μm in A, 10 μm in C and 40 μm in E. A statistics summary of the data shown here is available in Figure 8—source data 1.
-
Figure 8—source data 1
Summary of the statistics from Figure 8B,D and F.
- https://doi.org/10.7554/eLife.14694.032
Figure 8—figure supplement 1
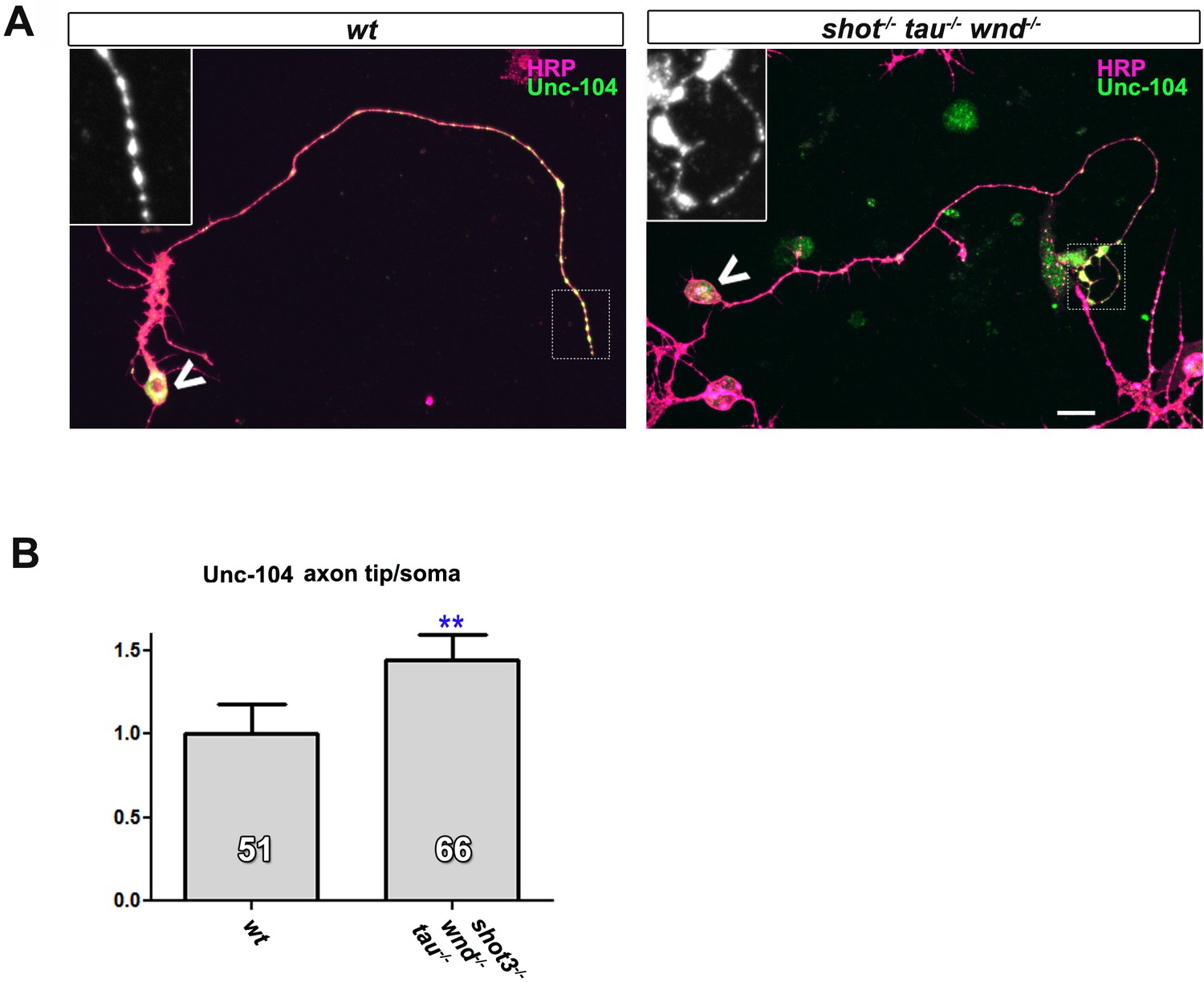
Attenuation of the JNK pathway rescues aberrant Unc-104 localisation in shot-tau mutant neurons in culture.
(A) Primary Drosophila neurons at 2 DIV, obtained from wildtype (wt) and shot3 tauMR22 wnd2 mutant embryos (shot-/- tau-/- wnd-/-), stained with antibodies against pan-neuronal HRP (magenta) and Unc-104 (green); Unc-104 in distal axon segments (emboxed and magnified in insets) is enriched in wildtype and in shot-/- tau-/- wnd-/- mutant neurons (chevrons indicate neuronal somata). This is in contrast to shot-tau mutant neurons, in which Unc-104 in distal axon segments is much weaker (for reference see Figure 5D). (B) Data were quantified as average intensity of Unc-104 at the distal end of the axon divided by the average intensity at the soma (**PMW<0.01). Scale bars: 5 μm in A. A statistics summary of the data shown here is available in Figure 8—figure supplement 1—source data 1.
-
Figure 8—figure supplement 1—source data 1
Summary statistics from Figure 8—figure supplement 1B.
- https://doi.org/10.7554/eLife.14694.034
Figure 8—figure supplement 2
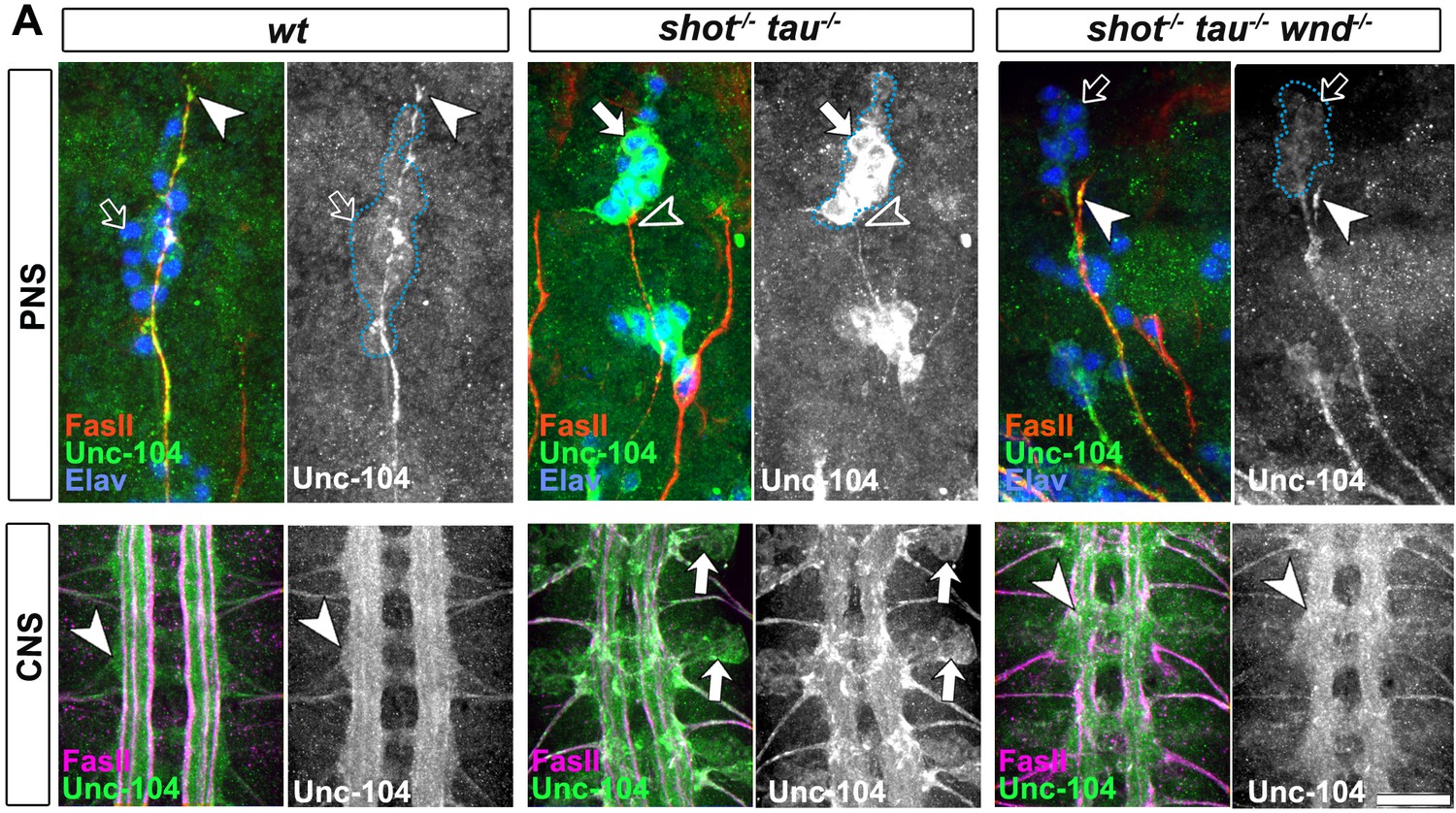
Attenuation of the JNK pathway rescue aberrant unc-104 localisation in shot-tau mutant embryos.
(A) The dorsal peripheral nervous system (PNS) and the central nervous system (CNS) of wildtype wildtype, shot3 tauMR22 and shot3 tauMR22 wnd2 mutant embryos (shot-/- tau-/- wnd-/-) at late stage 16 stained with FasII (red), Unc-104 (green) and elav (blue) for the PNS panels and FasII (magenta) and Unc-104 (green) in the CNS panels. Note the stark decrease of Unc-104 at the end of motor nerves (open versus white arrow heads) and the unusual accumulations of Unc-104 in the cell bodies of sensory neurons as well as in the CNS cortex in shot-tau embryos (open versus white arrows). This is in contrast to shot-/- tau-/- wnd-/- mutant neurons in which Unc-104 is increased at the end of motor nerves and decreased in the cell bodies of sensory neurons as well as in the CNS cortex. Scale bars: 15 μm in PNS panels and 35 μm in CNS panels.
Figure 9
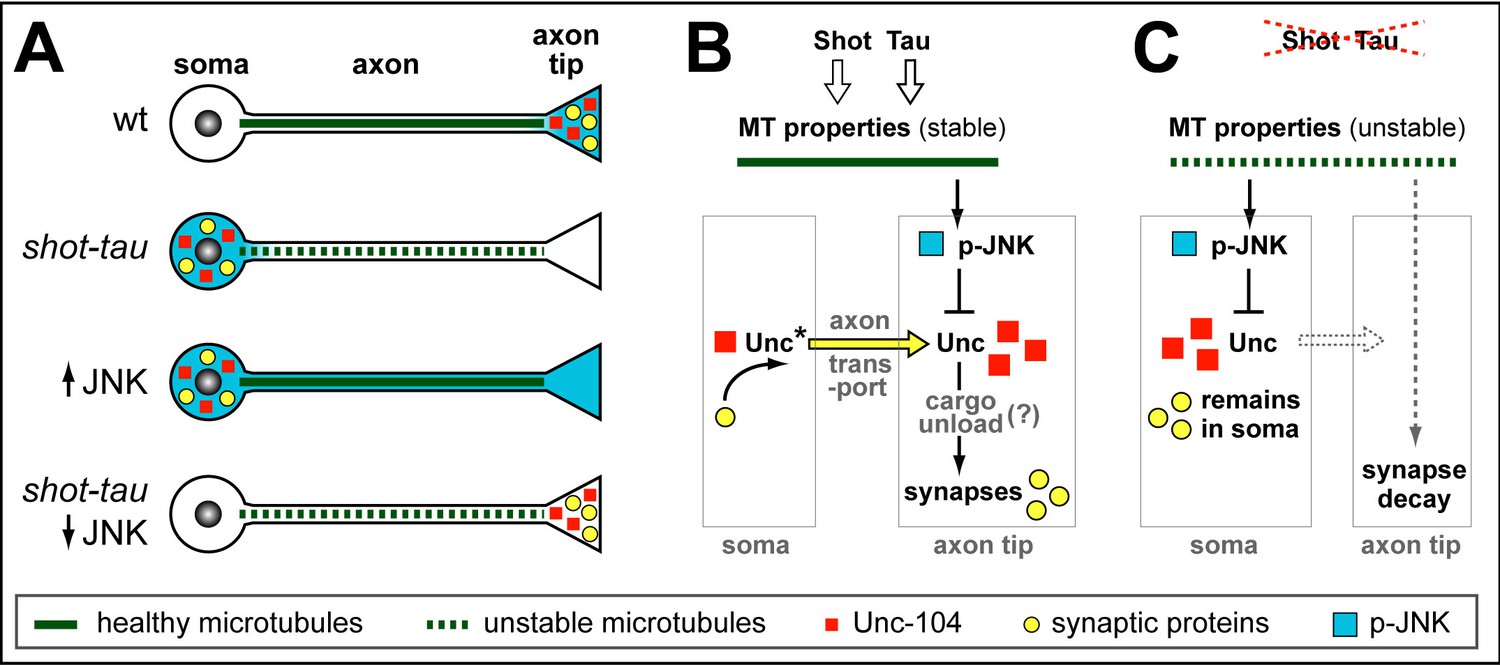
Schematic model of proposed function for Tau and Shot.
(A) Neurons illustrating different phenotypes: in wildtype neurons (wt), microtubules are stable (green line) and levels of Unc-104 (red square), synaptic markers (yellow dots) and p-JNK (turquoise background) are high in axon tips; in shot-tau mutant neurons, microtubules are unstable (stippled green lines), and the above listed proteins accumulate in cell bodies (soma); ubiquitous activation of JNK (↑JNK) causes similar somatic accumulation of Unc-104 and synaptic markers; down-regulation of JNK (↓JNK) rescues the shot-tau mutant phenotypes. (B,C) Schematic representation of the underlying mechanisms: In wildtype neurons (B), Unc-104 is activated (Unc*) and mediates axonal transport of synaptic proteins (yellow arrow) to the axon tip, where we propose (?) that active JNK inhibits Unc-104, thus releasing its cargo for synaptic incorporation. In shot-tau mutant neurons (C), unstable microtubules cause upregulation of JNK in the soma, thus inhibiting Unc-104 and trapping it as well as its cargo proteins in the soma.
Videos
Video 1
Live imaging of Drosophila neurons at 2 DIV, obtained from embryos carrying tau304 (a protein trap line where the endogenous tau gene is genomically tagged with GFP) and the microtubule binding protein Jupiter-Cherry.
Endogenous Tau (in green) is observed in a pattern reminiscent of microtubules, and colocalises with Jupiter (shown in red). The time laps were obtained every 15 s with a 3i Marianas Spinning Disk Microscope.
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Tau and spectraplakins promote synapse formation and maintenance through Jun kinase and neuronal trafficking
eLife 5:e14694.
https://doi.org/10.7554/eLife.14694




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}