The wiring diagram of a glomerular olfactory system
- Harvard University, United States
- Centre for Genomic Regulation, The Barcelona Institute of Science and Technology, Spain
- Universitat Pompeu Fabra, Spain
- Janelia Research Campus, Howard Hughes Medical Institute, United States
Figures
Figure 1 with 3 supplements
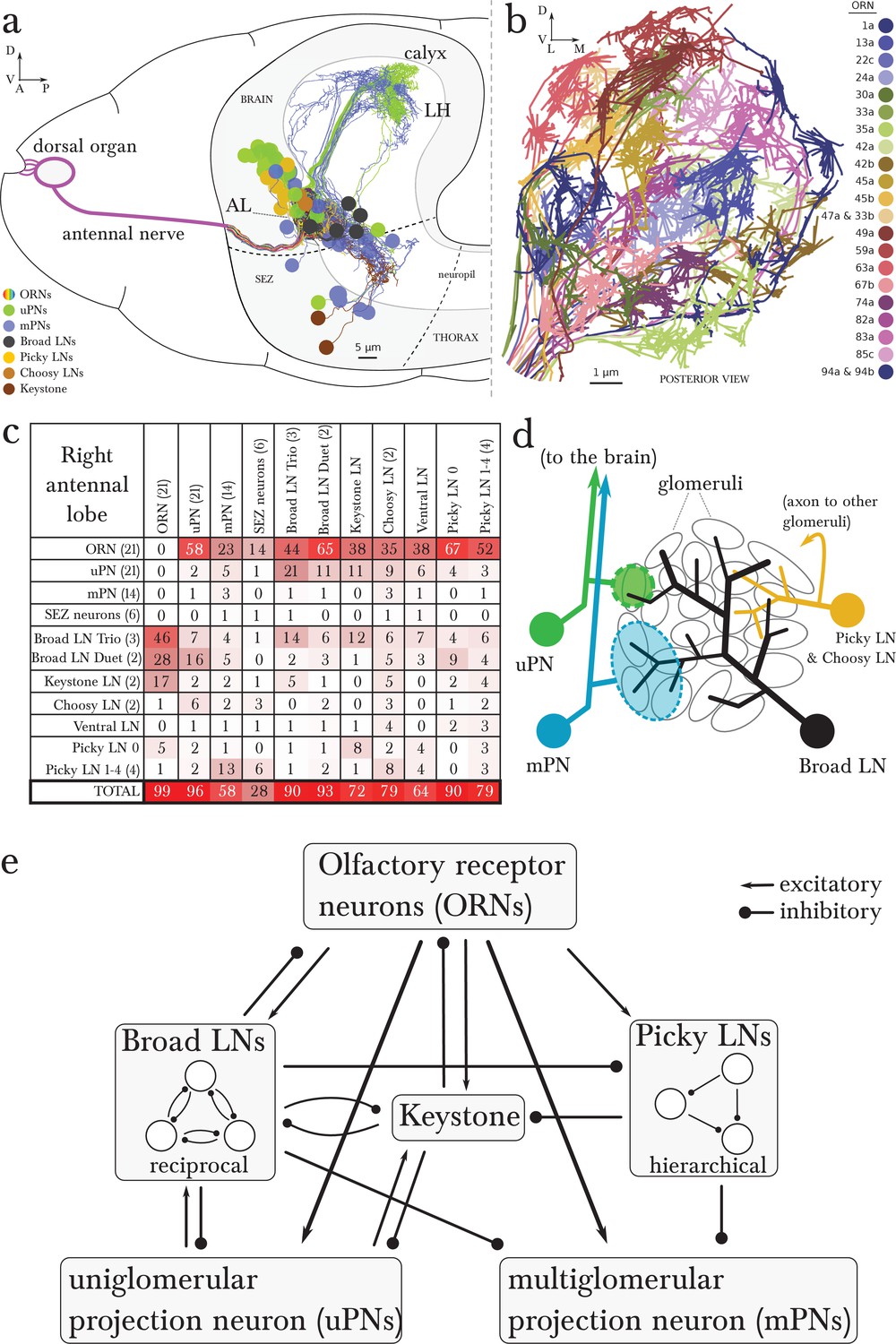
Overview of the wiring diagram of the glomerular olfactory system of the larval Drosophila.
(a) Schematic of the olfactory system of the larval Drosophila with EM-reconstructed skeletons overlaid. The ORN cell bodies are housed in the dorsal organ ganglion, extend dendrites into the dome of the dorsal organ, and emit axons to the brain via the antennal nerve. Like in all insects, neuron cell bodies (circles) reside in the outer layer of the nervous system (grey), and project their arbors into the neuropil (white) where they form synapses. Also shown are the major classes of local neurons (Broad LNs, Picky LNs and Keystone) and the 2 classes of projection neurons (uPNs and mPNs). The arbors of the Broad LNs (black) specifically innervate the AL. LNs and mPN dendrites can extend into the subesophageal zone (SEZ), innervated by sensory neurons of other modalities. uPNs project to specific brain areas (mushroom body calyx and lateral horn; LH), and mPNs mostly project to other nearby brain areas. (b) The larva presents 21 unique olfactory glomeruli, each defined by a single ORN expressing a single or a unique pair of olfactory receptors. We reconstructed each ORN with a skeleton and annotated its synapses, here colored like the skeleton to better illustrate each glomerulus. See Figure 1—figure supplement 1 for individual renderings that aided in the identification of each unique ORN. (c) Summary connectivity table for the right antennal lobe with all major neuron classes (4 neuromodulatory neurons and the descending neuron from the brain were omitted), indicating the percent of postsynaptic sites of a column neuron contributed by a row neuron. For most neurons, the vast majority of their inputs originates in other neurons within the antennal lobe. In parentheses, the number of neurons that belong to each cell type. We show only connections with at least two synapses, consistently found among homologous identified neurons in both the left and right antennal lobes. Percentages between 0 and 0.5 are rounded down to 0. (d) Schematic of the innervation patterns of the main classes of LNs and PNs in the antennal lobe. White ovals represent the glomeruli. Solid circles are cell bodies. Shaded areas with dotted outlines represent the extent of the PN dendritic arbors, with each uPN (green) innervating one glomerulus and each mPN (blue) innervating multiple glomeruli. Their axons (arrows) project to the brain. Broad LNs (black) are axonless and present panglomerular arbors. Picky LN (orange) dendrites span multiple glomeruli and their axons (arrow; not shown) target a different yet overlapping set of glomeruli as well as regions outside the olfactory system. Choosy LNs are similar to the Picky LNs but their axons remain within the antennal lobe. (e) A simplified wiring diagram of the larval olfactory system with only the main connections. ORNs are excitatory. All shown LNs are inhibitory. Broad LNs reciprocally connect to all glomeruli and each other and thus engage in presynaptic inhibition (on ORNs) and postsynaptic inhibition (on uPNs). Picky LNs form a hierarchical circuit and selectively synapse onto mPNs. Another LN, Keystone, receives inputs from ORNs, one Picky LN and non-ORN sensory neurons, and can potentially alter the operational mode of the entire olfactory system by altering the pattern of inhibition (see text).
Figure 1—figure supplement 1

Electron microscopy view of the antennal lobe of Drosophila larva.
(A) Cross-section of the antennal lobe, with dorsal up and lateral to the left. Notice most of the antennal lobe is wrapped by glial cells (darker profiles), but these do not fully enclose it (not shown), opening towards the SEZ (bottom right). Individual glomeruli are not wrapped in glia like in other insects (Oland et al., 2008), but glia do separate the antennal lobe from the neuronal somas and from nearby neuropils. ALT, antennal lobe tract. Corresponds to serial section 648 in the online volume, at bottom left (right antennal lobe from anterior view). (B) Magnification of the box in (A), showing an axon terminal of the 24a ORN synapsing onto the 24a uPN and multiple LNs. The dendrite of the 24a PN contains mitochondria, vesicles and presynaptic sites (magenta arrows), synapsing in this section onto e.g. one of the two Keystone LNs among others. The ORN axon bouton hosts multiple synapses (red arrows) with prominent ribbons or T-bars. The ORN boutons are packed with vesicles, giving them a darker appearance than surrounding PN and LN neurites; also contain numerous mitochondria (not shown in this image). Only some LNs are labeled for clarity; all neurites in this image were reconstructed. Blue arrows point to synapses of labeled LNs.
Figure 1—figure supplement 2
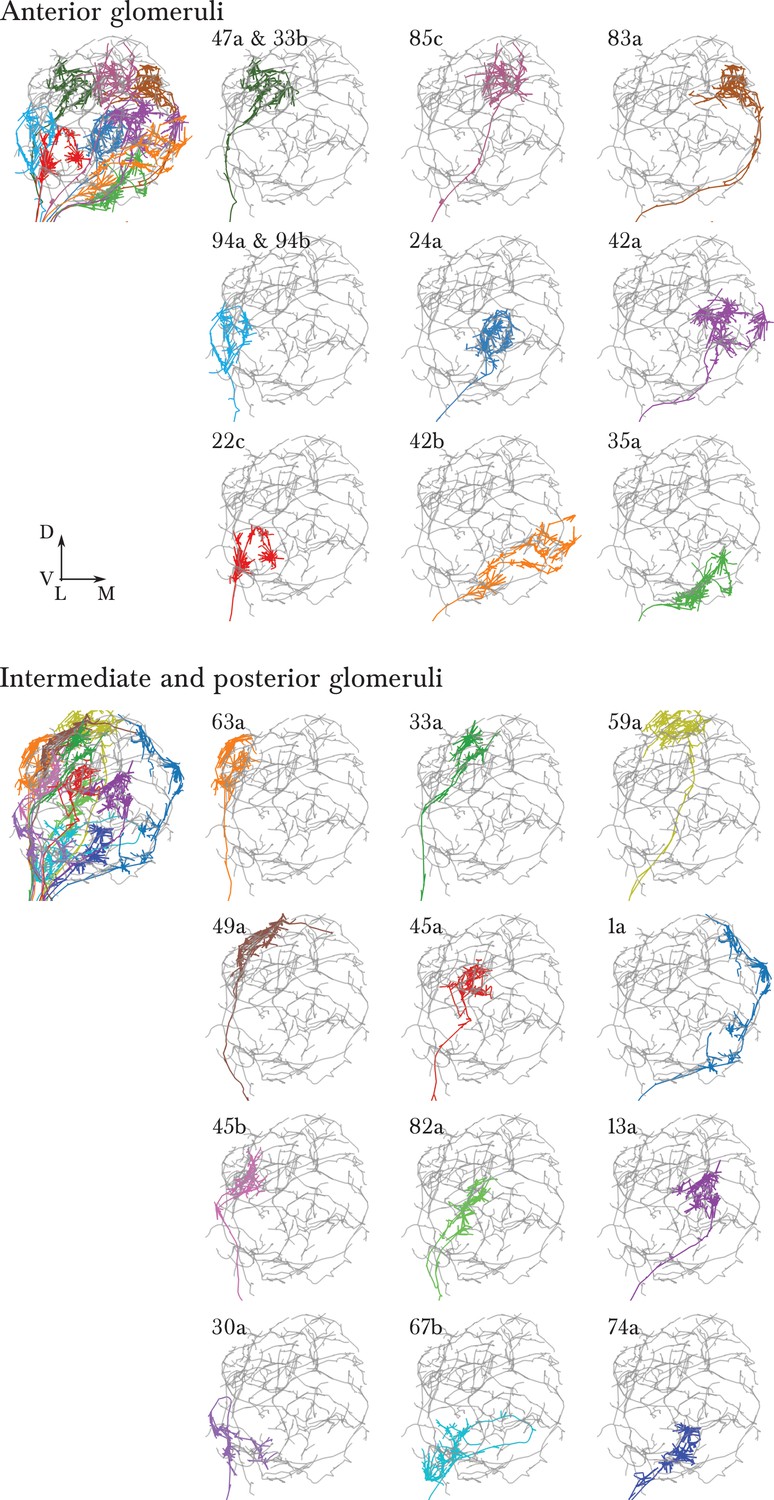
A single, identified ORN for each glomerulus in the antennal lobe of the first instar larva.
Each panel shows an EM-reconstructed arbor of an ORN (colored) over the background of a Broad LN Duet (grey). ORN synapses are rendered in the same color as the skeleton. To the left, all ORNs of each half of the antennal lobe are rendered together. The orientation (lateral to the left, dorsal up) and relative position of each ORN has been chosen to exactly match the arrangement in the supplementary Figure 1 of Masuda-Nakagawa et al. (2009), where each individual ORN was identified and labeled with GFP using genetic driver lines.
Figure 1—figure supplement 3
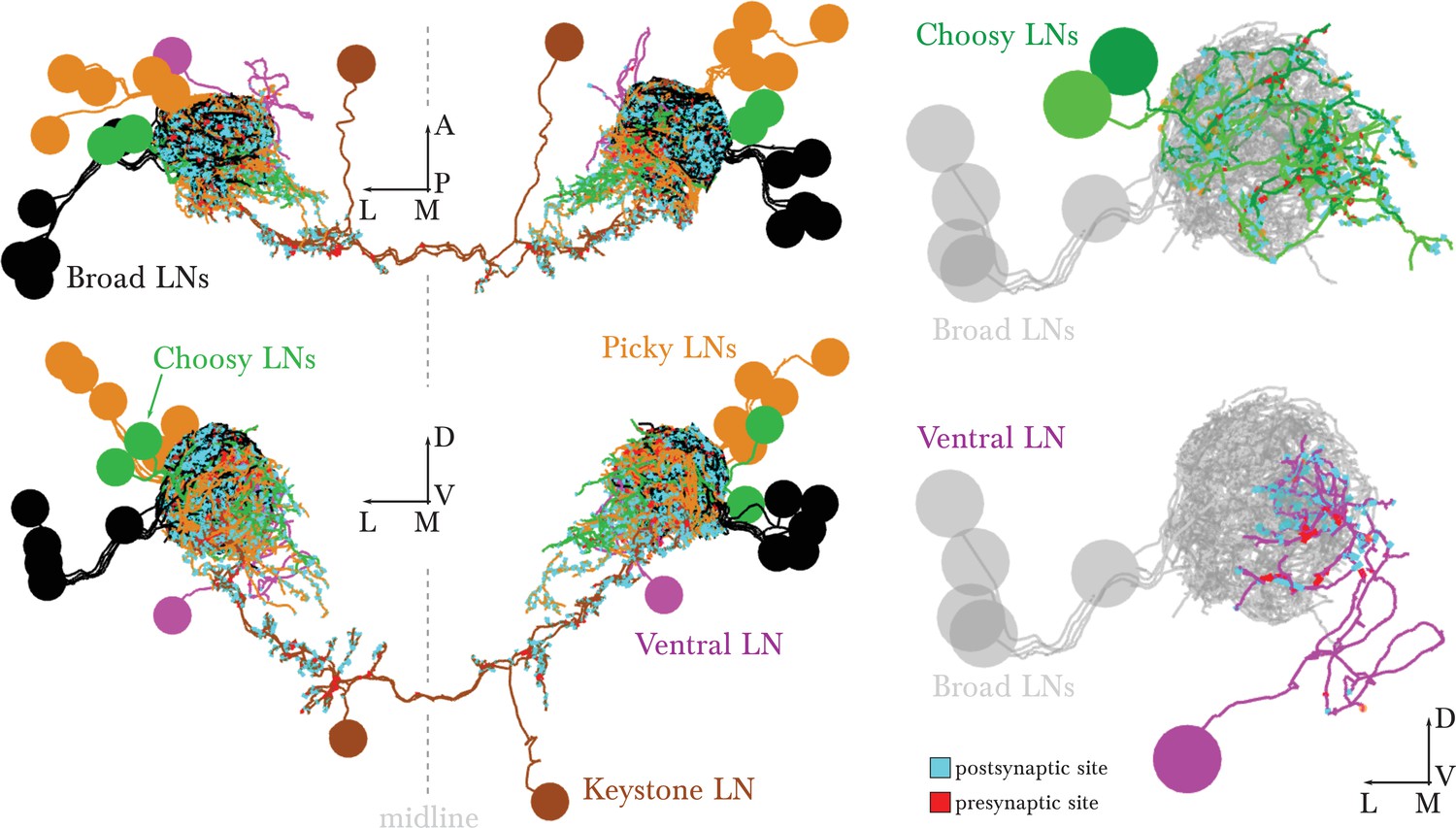
EM-reconstructed arbors of all LNs.
Left dorsal and posterior views of all LNs. The bundling of their primary axon tracts suggests that all LNs derive from 5 neuronal lineages (5 on the left and 5 on the right), shown in 5 different colors. Right, renderings of the left antennal lobe, posterior view. These identified neurons present similar morphology and connectivity in the right antennal lobe. Broad LNs are shown for reference. The morphology, cell body position and number of Choosy LNs matches that of the pair of GABAergic LNs described in Figure 2 L–O of Thum et al., 2011. Scale bar: a cell body measures about 4 micrometers in diameter.
Figure 2 with 1 supplement

Percentage of synapses of LNs from/onto specific cell types.
The entry for each neuron presents two bars, for the left and right homologs. Top row, Broad LNs and Keystone. T: Trio, D: Duet. Left, differences between the Trio and Duet subtypes are evident in the fraction of inputs that originates in ORNs, uPNs and Keystone. The Duet subtype presents a far larger fraction of its inputs from ORNs, and barely receives any inputs from Keystone. By its pattern of inputs, Keystone resembles a Broad LN Trio neuron, except for the large fraction of non-ORN inputs and the inputs from Picky LNs (specifically from Picky LN 0). Right, note how the Trio subtype devote about 25% of their synapses to each other, whereas the Duet subtype preferrentially targets uPNs, providing postsynaptic inhibition to the glomeruli (both lateral and feedforward inhibition). Keystone differs from the Broad LNs in that it targets uPNs much more weakly, preferring instead the Broad LN Trio and a variety of other neurons. Bottom row, Picky LNs, Choosy LNs and Ventral LN. Left, the fraction of inputs from ORNs stands out as a large difference among Picky LNs, with Picky LN 3 and 4 receiving substantially fewer, similarly to Choosy LNs. The fraction of inputs received from other Picky LNs (green) is among the most distinguishing feature of Picky LN 0, which receives close to none. Right, in contrast to the similar patterns of inputs onto all Picky LNs, Picky LN 0 stands out as very different from other Picky LNs in its choice of downstream synaptic partners, spreading approximately evenly between ORNs, uPNs, mPNs, other Picky LNs and Keystone. Choosy LNs strongly prefer uPNs, being therefore strong providers of postsynaptic inhibition to glomeruli. Notice that Picky LNs, Choosy LNs and Ventral LN have a larger fraction of synapses to/from 'others', with their arbors spreading towards adjacent sensory neuropils in the SEZ.
Figure 2—figure supplement 1

Neurotransmitters of Keystone LN and Picky LNs.
Genetic driver lines specific for Keystone LN (GAL4 line GMR27F08) and Picky LNs (split-GAL4 lines JRC_SS04499, JRC_SS04500, JRC_SS04260) driving GFP expression specifically in these neurons were labeled with anti-GABA and anti-vGlut (A–U), and also anti-Chat (all negative; not shown). Keystone presents immunoreactivity to anti-GABA (textbf A–C), and at least 4 of the 5 Picky LNs are positive to anti-vGlut and negative to anti-GABA (D–U). These neurons derive from the BAla2 lineage (Das et al., 2013). JRC_SS04260 drives expression specifically and uniquely a Picky LN, likely Picky LN 4, which presents anti-vGLut immunoreactivity (P–R). Left unlettered panels show the homologous identified EM-reconstructed neurons, with Broad LNs in grey for reference. Asterisks mark the location of cell bodies when there is not labeling, such as in panels I, O and U. Broad LNs and Choosy LNs are GABAergic (see Thum et al., 2011 at Figure 2 D–G for Broad LNs and L–O for Choosy LNs).
Figure 3 with 2 supplements
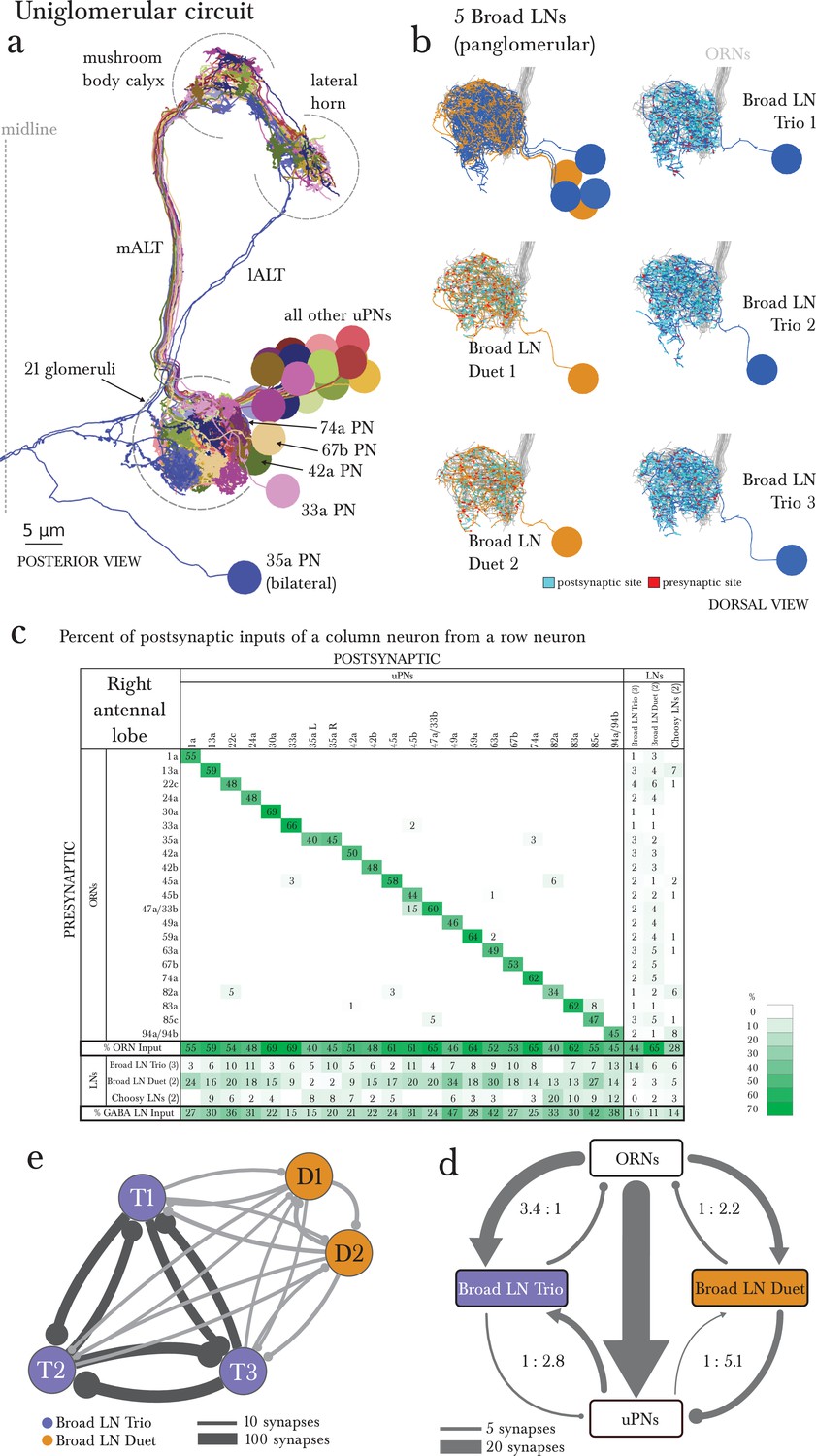
The uniglomerular circuit consists of 21 glomerular-specific projection neurons, which interact primarily with their corresponding ORN and with the 5 panglomerular LNs (Broad LNs), each an identified neuron.
(a) Posterior view of the EM-reconstructed uPNs of the right antennal lobe. The dendrites of each uPN delineate the glomerular boundaries, and the axons project to both the mushroom body (required for learning and memory; [Michels et al., 2011]) and the lateral horn (implicated in innate behaviors; [de Belle and Heisenberg, 1994]). 19 uPNs are likely generated by the same neuroblast lineage BAmv3 (Das et al., 2013) (although the uPNs for 42a, 74a, and 67b are slightly separated from the rest), and the other two (the uPNs for 33a and 35a) clearly derive from two other neuroblasts. Notice that the 35a uPN is bilateral, ascends through a different tract, and receives additional inputs outside of the antennal lobe. The 33a uPN does not synapse within the calyx and the 82a uPN does not continue to the lateral horn. The left antennal lobe (not shown) is a mirror image of the right one. (b) Dorsal view of the EM-reconstructed, axonless Broad LNs (Duet in orange; Trio in blue) shown together and individually. All neurons are on the same lineage: BAlc (Das et al., 2013). The pre-(red) and post-(cyan) synaptic sites on these panglomerular neurons are fairly uniformly distributed. ORNs in grey for reference. These neurons extend posteriorly out of the olfactory glomeruli to receive synapses from 2 non-ORN sensory neurons that enter the brain via the antennal nerve. (c) Percentage of the total number of postsynaptic sites on the dendrite of an uPN, Broad LN or Choosy LN (columns) that originate in a given ORN or LN (rows) for the right antennal lobe. Since the 35a uPN is bilateral, we include inputs to it from both antennal lobes. We show only connections with at least two synapses, consistently found among homologous identified neurons in both the left and right antennal lobes. Percentages between 0 and 1 are rounded to 1, but totals are computed from raw numbers. The uniglomerular nature of uPNs (notice the green diagonal) and panglomerular nature of Broad LNs is evident. The Broad LN Duet generally contributes more synapses onto uPNs than the Broad LN Trio does. While the number of synapses that an ORN makes onto its uPN varies widely (24–120 synapses; see Supplementary file 1 and 2), this number is tailored to the size of the target uPN dendrite given that percentage of inputs the ORN contributes to the uPN is much less varied (mostly 45–65%). For an extended version of this table that includes all LNs, see Figure 3—figure supplement 1. (d) Both Broad LN types (Trio and Duet) mediate presynaptic inhibition (synapses onto ORN axons) similarly, but the Duet shows far stronger postsynaptic inhibition (synapses onto uPN dendrites) while the Trio receives far more dendro-dendritic synapses from uPNs. Connections among Broad LNs are not shown for simplicity. Each arrow is weighted linearly by the number of synapses for an average single Broad LN of each type. (e) The 5 Broad LNs that govern this circuit synapse reciprocally, with the Trio type synapsing more strongly onto each other. Shown here for the right antennal lobe with arrow thickness weighted by the square root of the number of synapses.
Figure 3—figure supplement 1
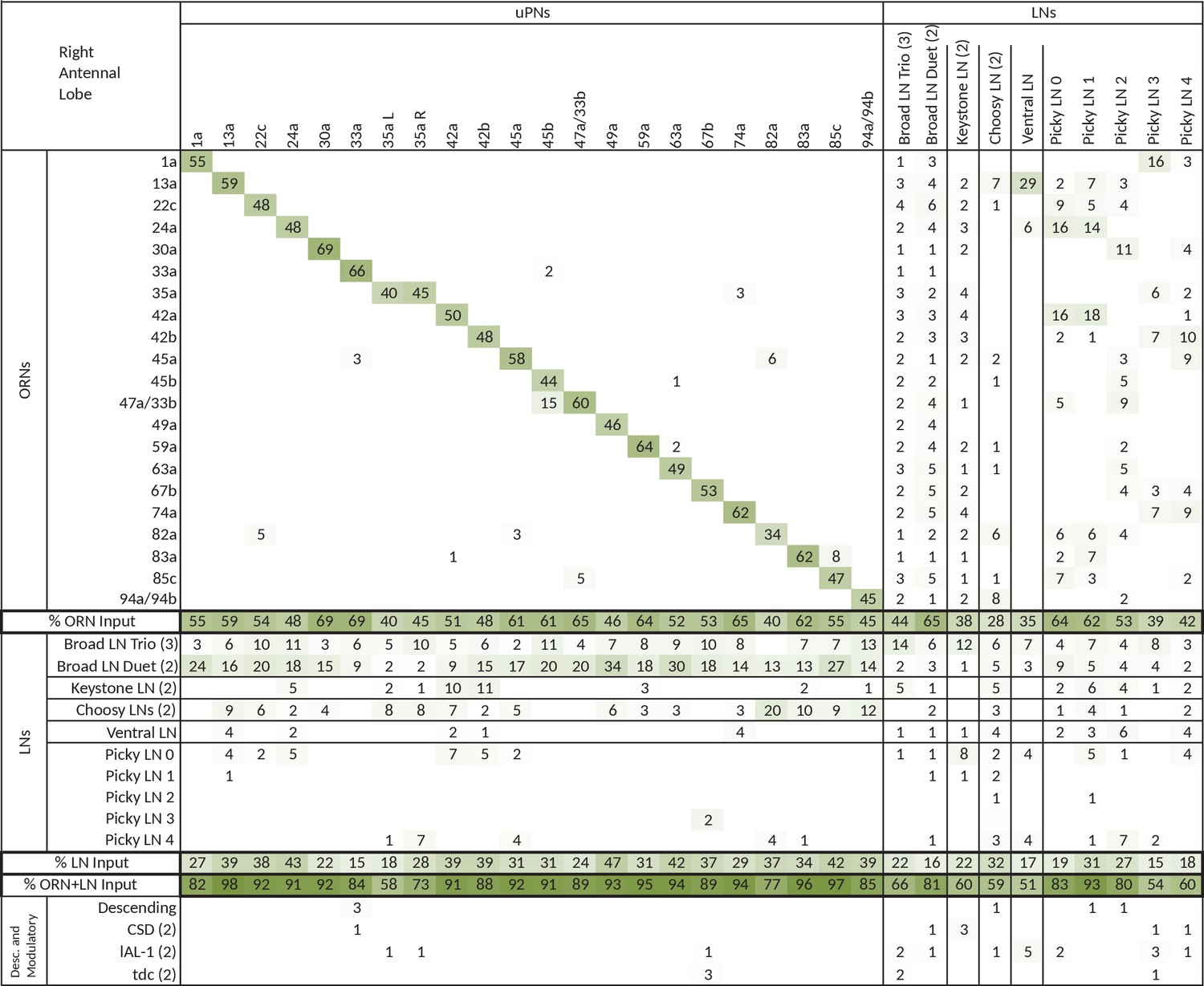
Extended version of table in Figure 3c, including all other olfactory-related neurons.
Tables show percent of postsynaptic sites of a column neuron contributed by a row neuron. We show only connections with at least two synapses, consistently found among homologous identified neurons in both the left and right antennal lobes. Percentages between 0 and 0.5 are removed. For bilateral neurons, inputs from both sides are included.
Figure 3—figure supplement 2
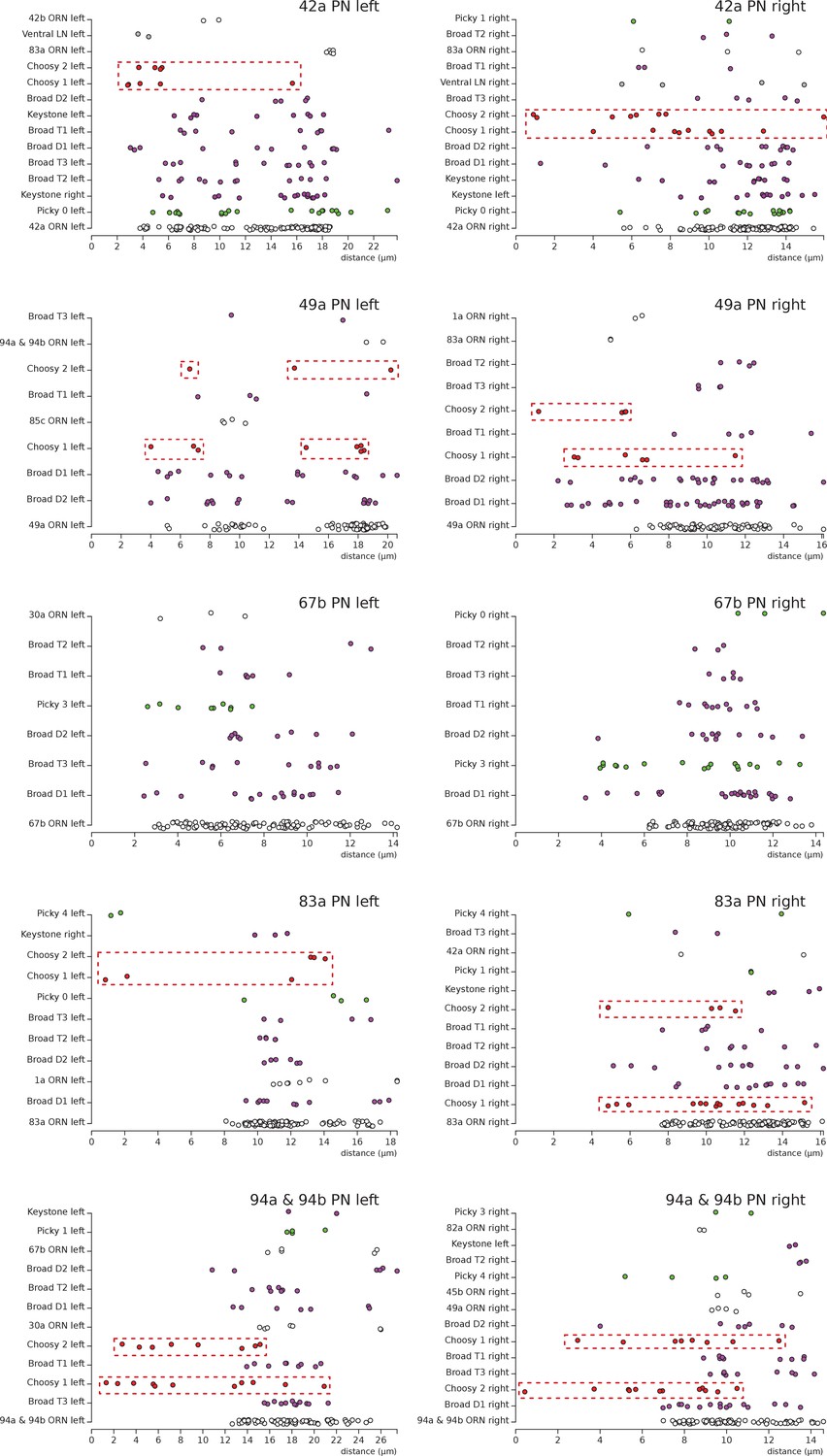
Distribution of postsynaptic sites on the uPN dendrites.
We show 5 examples, plotting the distance (along the cable) of individual postsynaptic sites (colored dots) to the axon initial segment of each uPN. The same type of presynaptic neuron presents the same color across all plots. Notice how Choosy LN inputs (red, framed in a red box) onto uPNs are generally more proximal to the axon initial segment than other inhibitory inputs such as from Broad LNs; particularly noticeable for 42a PN (top row) and 94 & 94b PN (bottom row). No noticeable difference exists between Broad LN Duet and Trio. Notice that the left 49a PN presents an arbor with two main dendrites, with one being further than the other from the axon initial segment, explaining the split in the distribution of distances of postsynaptic sites. While 67b PN (third row) does not receive inputs from Choosy LNs, the Picky LN 3 (light green), which specifically targets 67b PN and no other uPN, provides proximal inputs. Presynaptic neurons are ordered with the largest contributor at the bottom of each plot.
Figure 4 with 4 supplements
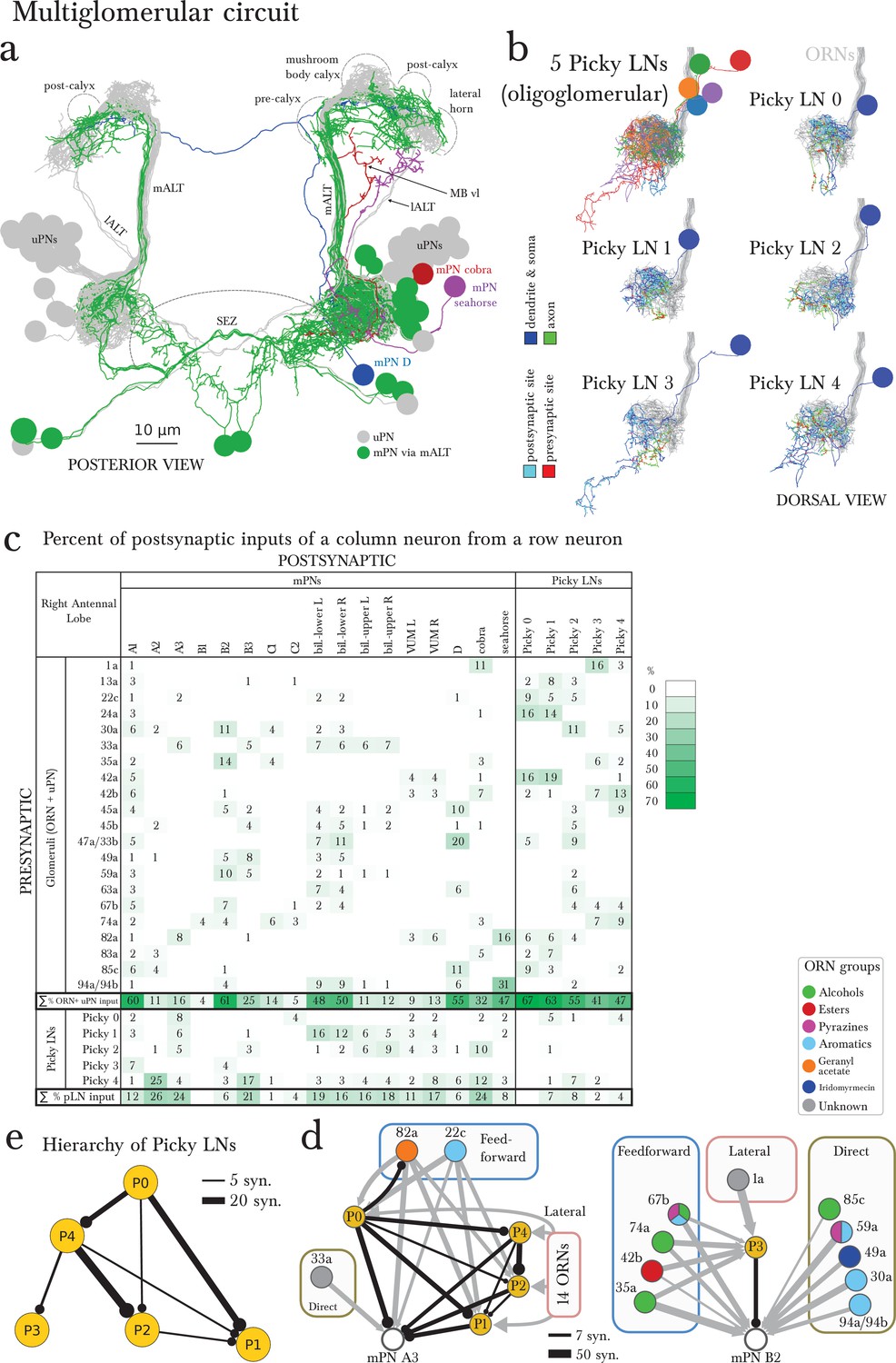
The multiglomerular circuit consists of 14 mPNs that project to the brain and 5 Picky LNs, each an identified neuron.
(a) Posterior view of EM-reconstructed mPNs that innervate the right antennal lobe (in color; uPNs in grey for reference), each receiving inputs from a subset of olfactory glomeruli but many also from non-ORN sensory neurons in the subesophageal zone (SEZ). Most mPNs (green) project via the same tract as the uPNs (mALT). They can project via other tracts (other colors), but never via the mlALT used by the iPNs of the adult Drosophila. The mPNs project to many regions including a pre-calyx area, a post-calyx area, the lateral horn (LH) and the mushroom body vertical lobe (MB vl). mPNs are generated by diverse neuroblast lineages including BAlp4, BAla1, and others (Das et al., 2013). (b) Dorsal view of the EM-reconstructed Picky LNs shown together and individually. When shown individually, the Picky LNs are in 2 colors: blue for the dendrites and soma, and green for the axon. Zoom in to observe that presynaptic sites (red) are predominantly on the axon, whereas postsynaptic sites (cyan) are mostly on dendrites. Collectively, the dendritic arbors of the 5 Picky LNs tile the olfactory glomeruli. The dendrites of the Picky LN 3 and 4 extend significantly into the SEZ. They all originate from the same neuroblast lineage: BAla2 (Das et al., 2013). (c) Percentage of the total number of postsynaptic sites on the dendrite of a mPN or Picky LN (column neuron) that originate from a given glomerulus or Picky LN (row neurons). Here we define the glomerulus as connections from the ORN or via dendro-dendritic synapses from a given ORN’s uPN. This is most relevant for mPN A1, which can receive more synapses from an ORN’s uPN than the ORN itself (see suppl. Adjacency Matrix). We show the inputs to the mPNs and Picky LNs for the right antennal lobe, but for all bilateral mPNs (bil.-lower, bil.-upper, and VUM) we include inputs from both sides. We show only connections with at least two synapses, consistently found among homologous identified neurons in both the left and right antennal lobes. Percentages between 0 and 1 are rounded to 1, but totals are computed from raw numbers. Connections in this table are stereotyped (when comparing the left and right antennal lobes) and selective. Note that mPNs that receive many inputs from non-ORN sensory neurons in the SEZ have a low total of ORN+uPN input. For an extended version of this table that includes all LNs see Figure 4—figure supplement 2. (d) The direct upstream connectivity for two mPNs, with ORNs colored by the groups emerging from the PCA analysis of odor tuning. Connections from ORNs and Picky LNs to mPNs create 3 different types of motifs: direct excitatory connections from ORNs, lateral inhibitory connections from ORNs only via Picky LNs, and feedforward loops where an ORN connects both directly to the mPN and laterally through a Picky LN. Note that the activity of Picky LN 0 could alter the integration function for mPN A3 and indirectly for B2, as well as many other mPNs (not shown). Arrow thicknesses are weighted by the square root of the number of synapses between neurons. (e) The Picky LN hierarchy, dominated by Picky LN 0, here showing connections with 2 or more consistent synapses between bilaterally homologous neurons. Some of these connections are axo-axonic (see Figure 4—figure supplement 3).
Figure 4—figure supplement 1
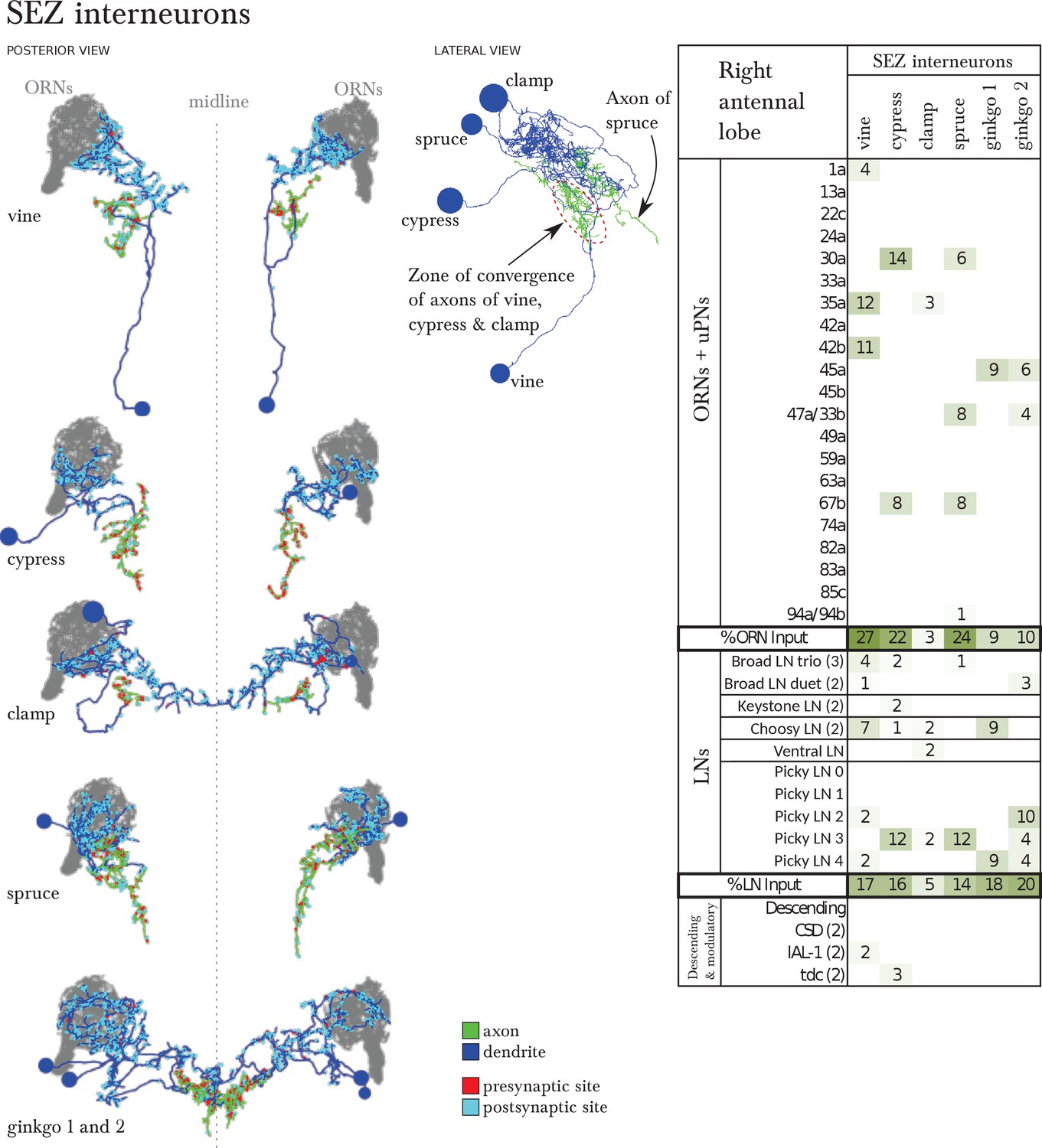
Six SEZ neurons receive specific inputs from some ORNs and from some antennal lobe LNs.
Left, EM-reconstruction of the 6 SEZ neurons (vine, cypress, clamp, spruce and ginkgo 1 and 2), with their axons labeled green and their dendrites blue. Presynaptic sites in red and postsynaptic sites in cyan. Middle, 3 of these SEZ neurons project to the same unidentified region of the SEZ. Spruce projects to a more posterior area. Lateral view, anterior to the left. Right, table of percent of postsynaptic sites of a column neuron contributed by a row neuron, illustrating how some ORNs and LNs specifically target these SEZ neurons. We show only connections with at least two synapses, consistently found among homologous identified neurons in both the left and right antennal lobes. Percentages between 0 and 0.5 are removed. Notice how Picky LNs 2, 3 and 4 synapse strongly onto SEZ neurons.
Figure 4—figure supplement 2
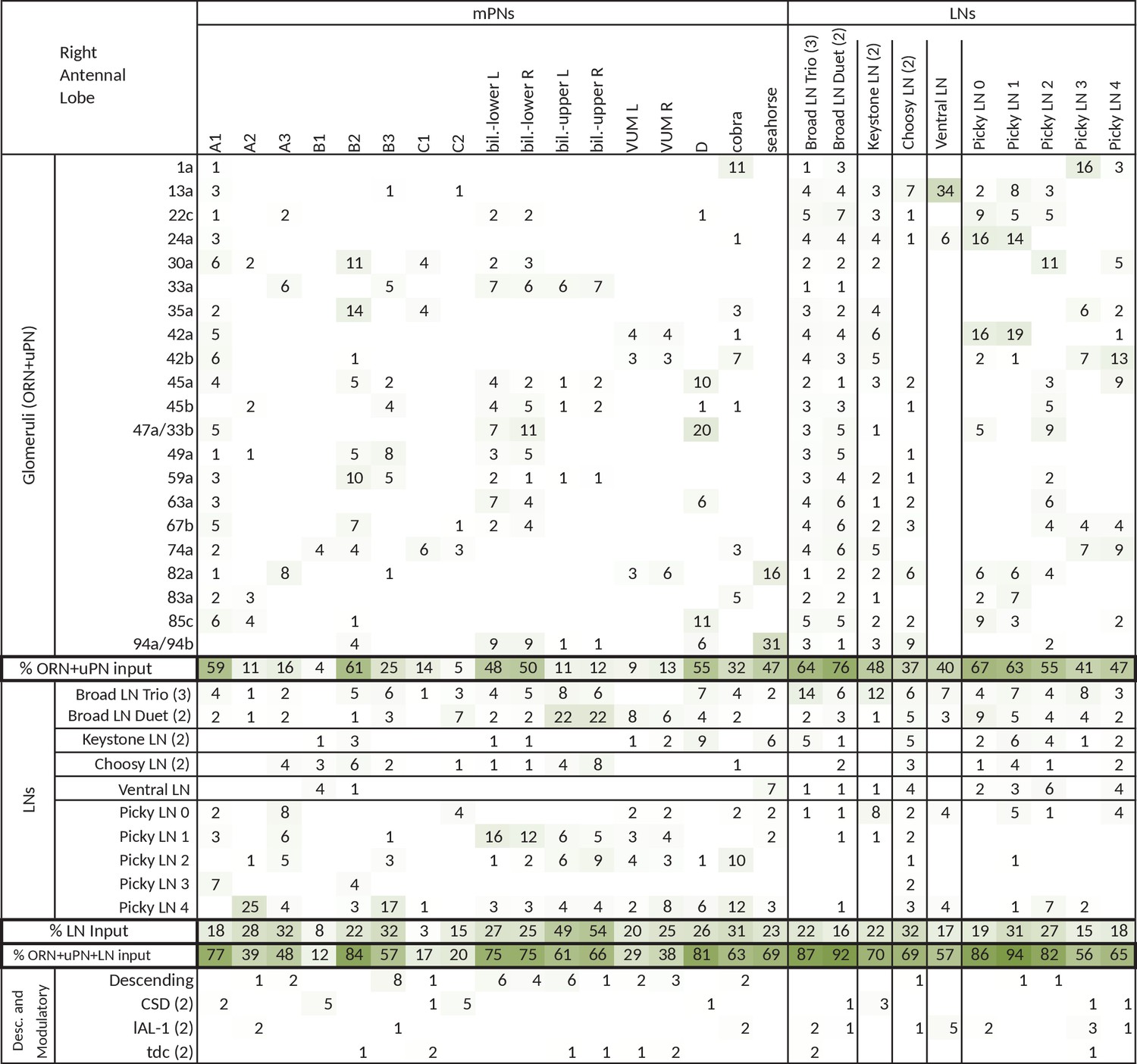
Extended version of table in Figure 4c, including all other olfactory-related neurons.
Tables show percent of postsynaptic sites of a column neuron contributed by a row neuron. We show only connections with at least two synapses, consistently found among homologous identified neurons in both the left and right antennal lobes. Percentages between 0 and 0.5 are removed. For bilateral neurons, inputs from both sides are included.
Figure 4—figure supplement 3
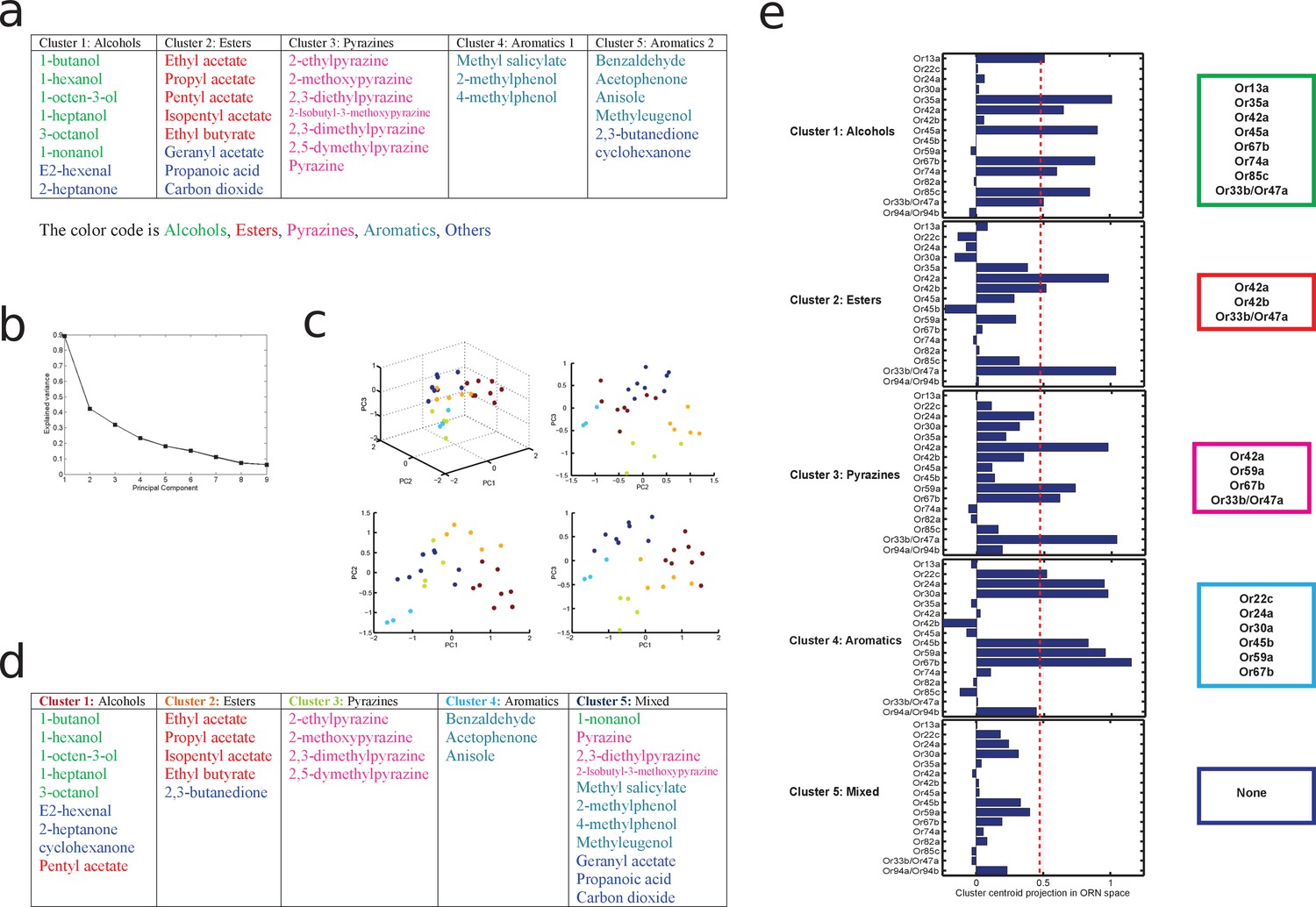
principal component analysis of odors leading to a principled clustering of orns.
(a) Clustering of odors by odorant-descriptor. Results of K-means clustering of odors in the 32 dimensional odor-descriptor space proposed in Haddad et al. (2008). Odors cluster into five groups that are well correlated with odor chemical type (alcohols, aromatics, esters, pyrazines, and others). (b–e) Clustering of odors by ORN response. (b) The variance explained for the odors in ORN response space as a function of the number of principal components (dimensions). The 'elbow' of this curve is composed of the principal components used for the clustering analysis of the odors by ORN-response. (c) How the odors span the space of the first 3 principal components of ORN response space. The odors are individual points colored by which of the five clusters, calculated via an affinity propagation clustering algorithm, they belong to. (d) How each of the odors fit into the clusters in ORN response space. Each cluster tends to group odors of similar chemical type. (e) The ORNs that represent the centroid of each cluster, calculated using a threshold obtained via Otsu’s method. See Materials and methods for further details.
Figure 4—figure supplement 4
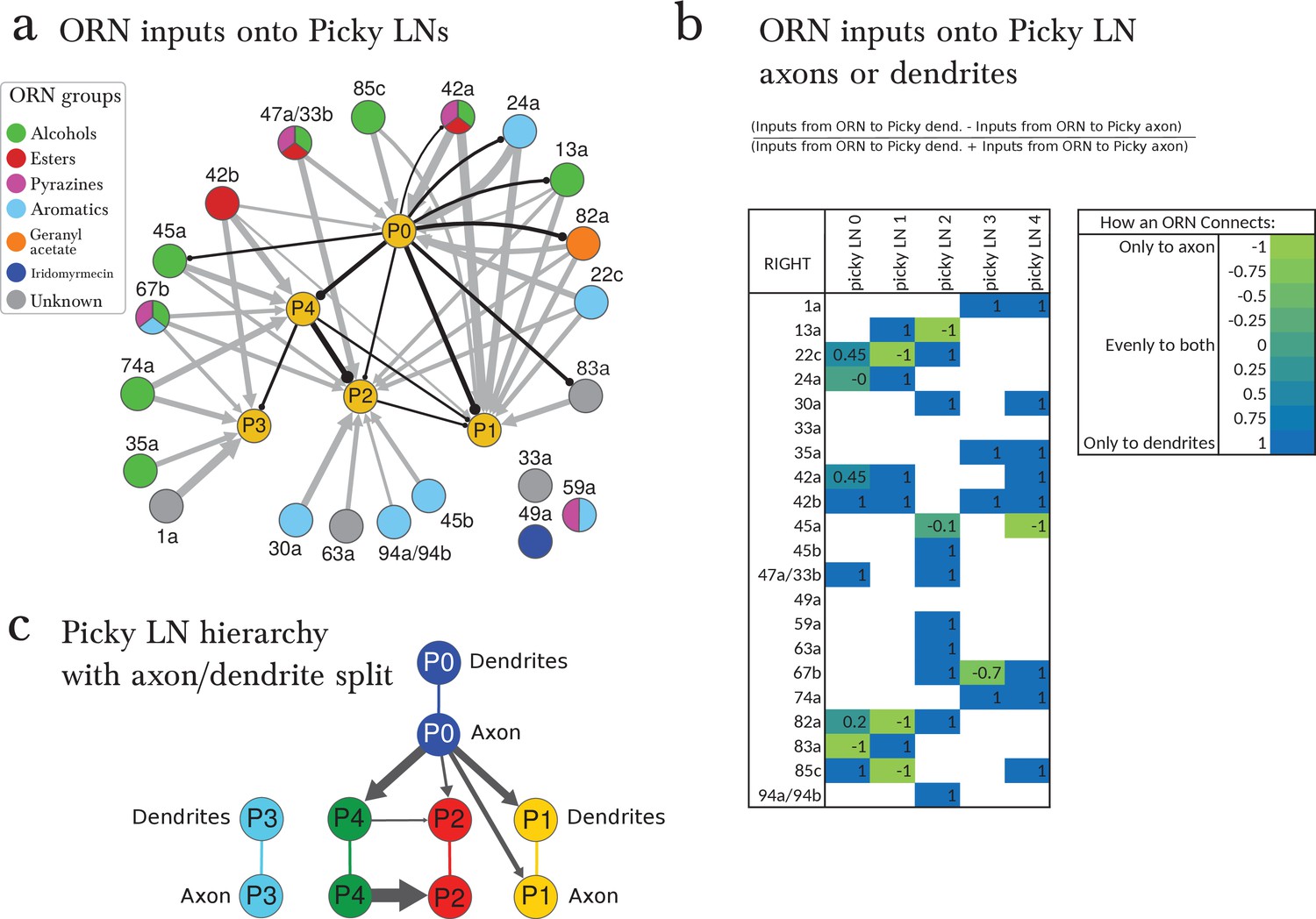
Pattern of ORN inputs onto Picky LNs.
(a) The connections of ORNs onto the hierarchy of Picky LNs. ORNs are colored by the groups emerging from the PCA analysis of odor tuning. Inhibitory connections from Picky LNs are shown in black (only connections with 2 or more synapses among bilaterally homologous neuron pairs are show). Excitatory connections from ORNs are shown in grey (only connections with 4 or more synapses among bilaterally homologous neuron pairs are shown). See Supplementary file 1 and 2 (containing the adjacency matrices) for the complete set of connections. The thickness of the arrows is proportional to the square root of the number of synapses. Some of these connections are axo-axonic (see c). (c) ORNs can synapse onto the Picky neurons at either their dendrites or their axons. This table shows values from -1 to 1 based on the written formula. Values between -1 and 0 correspond to the ORN synapsing more to the axon of the Picky LN than the dendrite, and values between 0 and 1 correspond to the ORN synapsing more to the dendrite of the Picky LN than the axon. Only consistent connections between ORNs and Picky dendrites or ORNs and Picky axons with a threshold of at least 2 consistent synapses per side are used to calculate these ratios. For values that are not 1 or -1, the value can differ from side to side. Because the threshold is lowered from that of A, more connections appear, but since we only consider connections consistent in how they connect to the Picky LNs (to dendrite or axon), some of the weakest connections also drop out compared to a. (c) The Picky LN hierarchy shown with the Picky LNs split into axon and dendrite, showing that not all connections are from the axon of a Picky LN to the dendrites of another. We are only showing connections that are consistent both in their motif (axo-axonic, dendro-dendritic, etc) and with a consistent threshold of 2 synapses on both sides. Because these criteria are more stringent than those used in a, some connections drop out (such as Picky LN 4 to Picky LN 3).
Figure 5 with 1 supplement
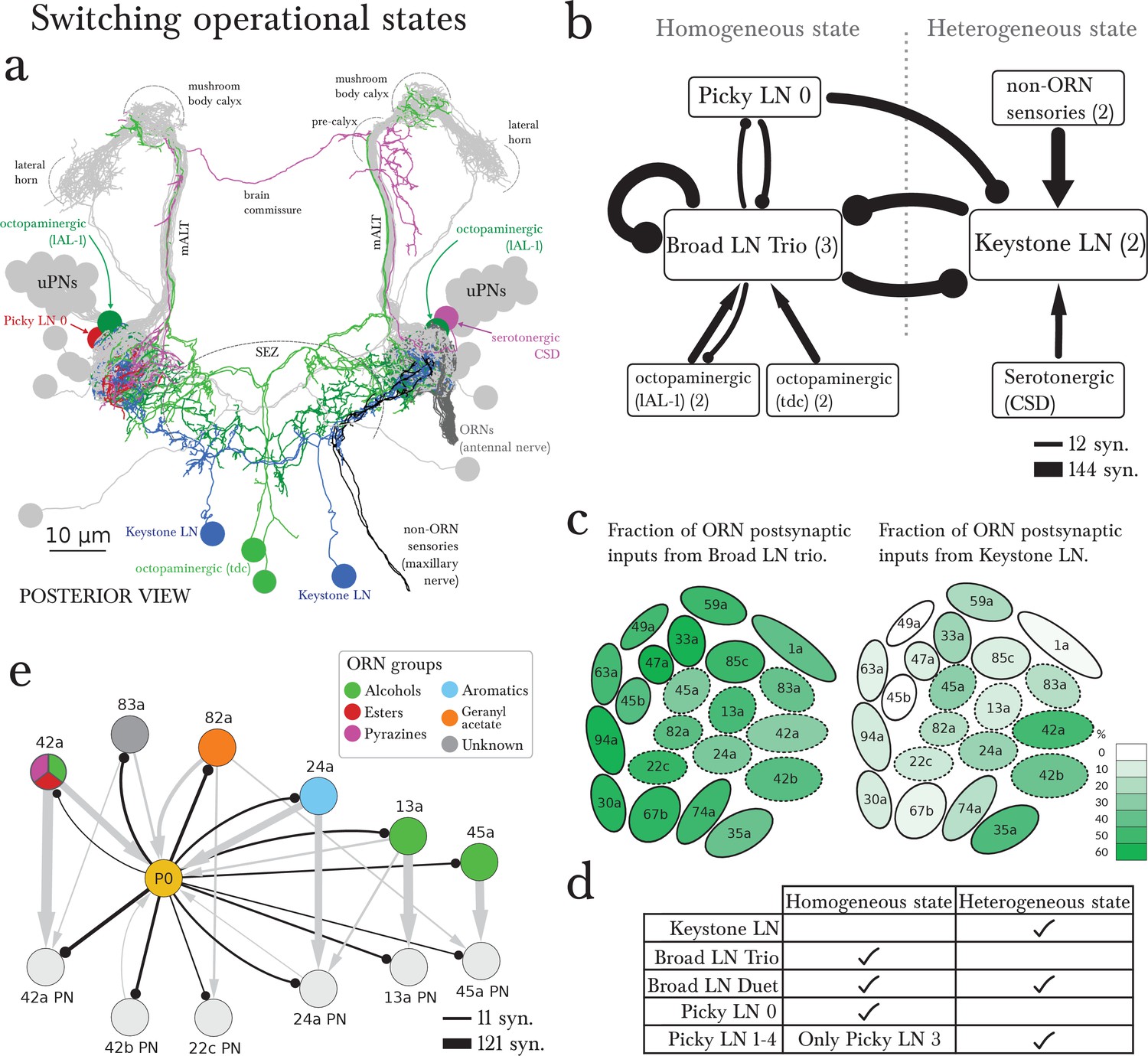
The wiring diagram suggests two operational states: homogeneous or heterogeneous presynaptic inhibition.
(a) Posterior view of the EM-reconstructed neurons innervating the left antennal lobe that could govern the switch (uPNs in grey and right ORNs in dark grey for reference). The Keystone LN (blue) has a symmetric bilateral arbor and additionally innervates the SEZ, receiving inputs from non-ORN sensory neurons (in black). Neuromodulatory neurons that make direct morphological synapses onto LNs are serotonergic (CSD in pink; projects contralaterally after collecting inputs from near the MB calyx) and octopaminergic (lAL-1 and two tdc, in dark and light green), and all arborize well beyond the antennal lobe. Also included is Picky LN 0 (red). (b) A wiring diagram outlining the strong LN-LN connections, showing the core reciprocal inhibition between Broad LN Trio and Keystone that could mediate the switch between homogeneous (panglomerular) presynaptic inhibition and heterogenous (selective) presynaptic inhibition. For simplicity, neurons are grouped together if they belong to the same neuron type, with the number of neurons belonging to each group indicated in parentheses. Connections are weighted by the square root of the number of synapses between groups of neurons. The self-arrow for the Broad LN Trio represents the average number of synapses that one of the Trio neurons receives from the other two. Picky LN 0 inhibits Keystone, thereby disinhibiting the Broad LN Trio and promoting homogeneous presynaptic inhibition. The maxillary nerve sensory neurons are the top input providers of Keystone and may drive the system towards heterogeneous presynaptic inhibition (see C). The effect of direct inputs from neuromodulatory neurons is unknown, but at least it has been suggested that octopaminergic neurons may have an excitatory effect on inhibitory LNs (Linster and Smith, 1997). (c) Cartoon of glomeruli colored by the percentage of inputs onto ORN axon terminals provided by the Broad LN Trio and from Keystone, indicating the amount of presynaptic inhibition (onto ORNs) in either state. The inhibition provided by Broad LN Trio is much more uniform than the inhibition provided by Keystone. Dotted lines indicate glomeruli that receive Picky LN 0 input on either the ORN or uPN. (d) The LNs putatively active in each state. (e) Unlike other Picky LNs, Picky LN 0 makes synapses onto ORN axon terminals and many uPNs. Here connections with 2 or more synapses consistent between bilaterally homologous neuron pairs are shown. Arrow thicknesses are weighted by the square root of the number of synapses between neurons. With the exception of 45a, all shown ORNs and uPNs belong to glomeruli that synapse onto Picky LN 0 as well. Thus Picky LN 0 provides both pre- and postsynaptic inhibition to a small set of glomeruli.
Figure 5—figure supplement 1
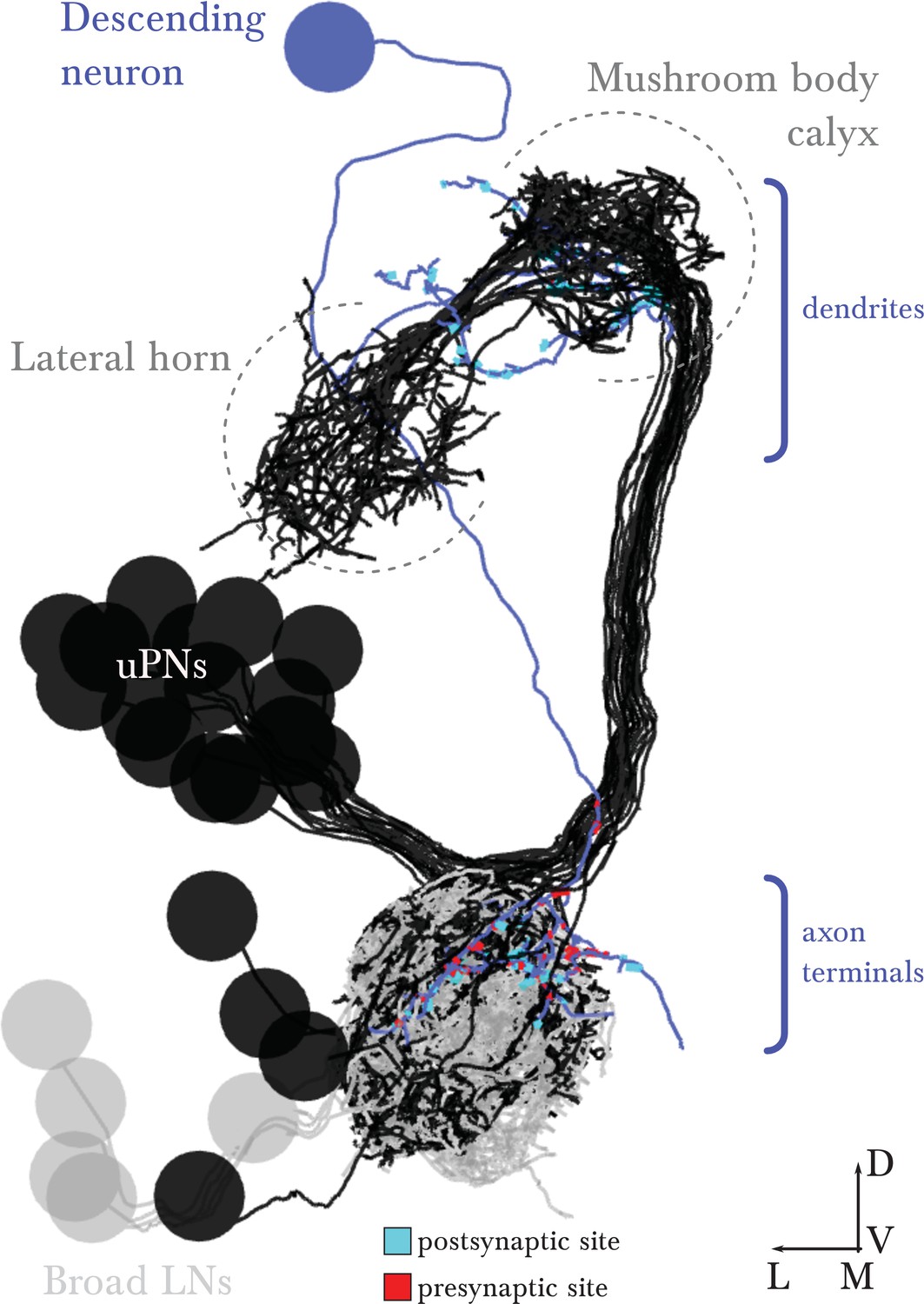
EM-reconstructed arbor of the descending neuron.
Renderings of the left antennal lobe, posterior view. This identified neuron exists in the left and right antennal lobes, presenting similar morphology and connectivity in the right antennal lobe. Broad LNs and uPNs are shown for reference. Scale bar: a cell body measures about 4 micrometers in diameter.
Additional files
-
Supplementary file 1
Adjacency matrices with the complete synaptic connectivity of the wiring diagram of the left antennal lobe.
- https://doi.org/10.7554/eLife.14859.019
-
Supplementary file 2
Adjacency matrices with the complete synaptic connectivity of the wiring diagram of the right antennal lobe.
- https://doi.org/10.7554/eLife.14859.020
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
The wiring diagram of a glomerular olfactory system
eLife 5:e14859.
https://doi.org/10.7554/eLife.14859




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}