Mouse Tmem135 mutation reveals a mechanism involving mitochondrial dynamics that leads to age-dependent retinal pathologies
- University of Wisconsin-Madison, United States
- Morgridge Institute for Research, United States
- Northwestern University, United States
- University of Iowa, United States
- Howard Hughes Medical Institute, University of Texas Southwestern Medical Center, United States
Figures
Figure 1
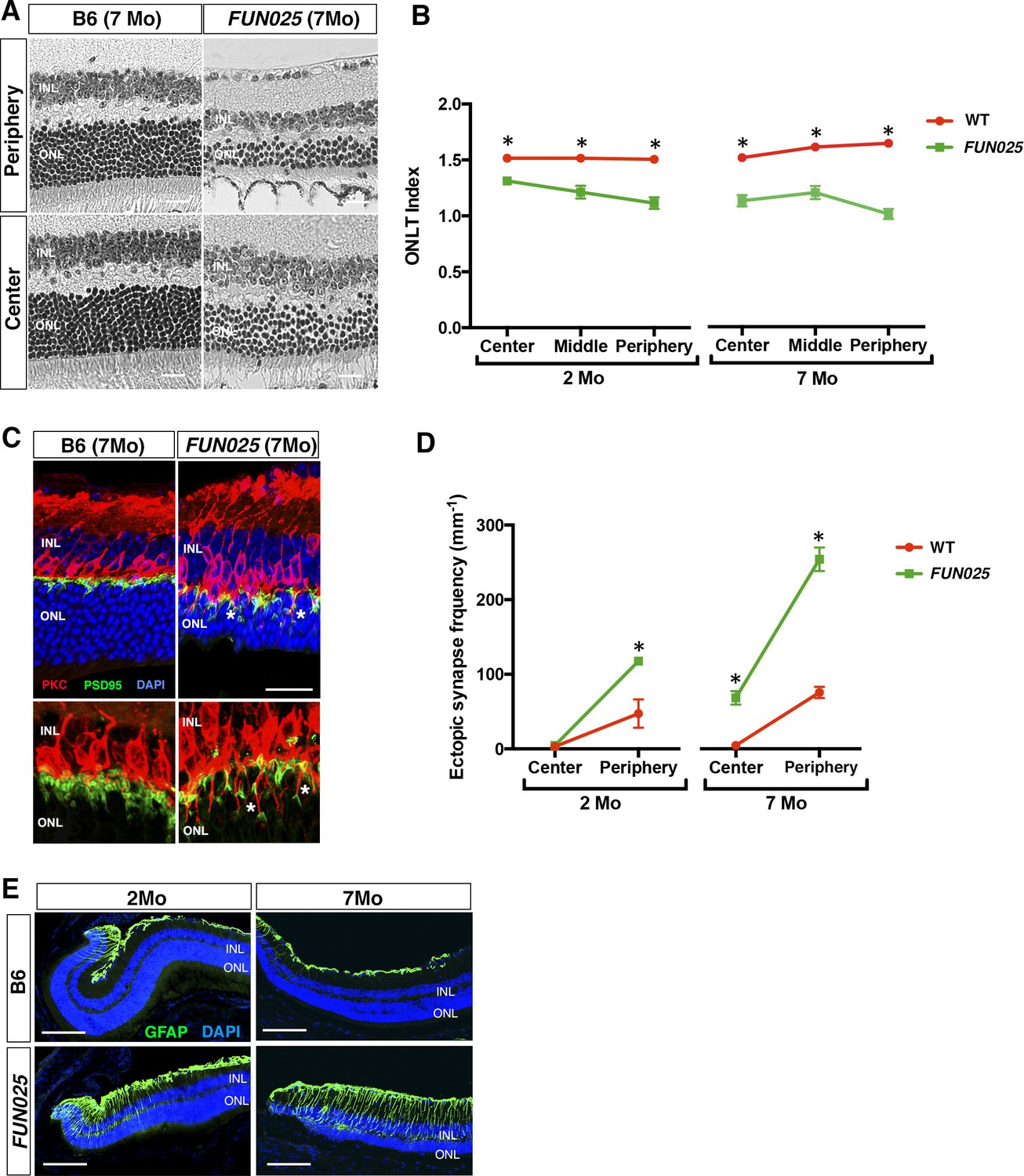
Age-dependent retinal abnormalities in FUN025 mice.
(A–B) A significant decrease of the ONLT index occurred by two months of age in FUN025 retina. Mo = months. Data from n = 10 WT (2 Mo), n = 4 FUN025 (2 Mo), n = 20 WT (7 Mo), n = 8 FUN025 (7 Mo) mice. Scale bar = 20 μm. (C–D) Ectopic synapses were observed as bipolar cell neurites (PKC, red) and photoreceptor synaptic terminals (PSD95, green) extending into the ONL indicated by asterisks (C). Scale bar = 10 μm. Significant increase of ectopic synapses were found earlier in the peripheral retina, and later in the central retina of FUN025 compared to WT mice. Data for central retina from n = 3 WT (2 Mo), n = 3 FUN025 (2 Mo), n = 3 WT (7 Mo), n = 3 FUN025 (7 Mo) mice; data for peripheral retina from n = 5 WT (2 Mo), n = 6 FUN025 (2 Mo), n = 6 WT (7 Mo), n = 6 FUN025 (7 Mo) mice. (E) GFAP (green) upregulation was progressively observed in the FUN025 retina. ONL: outer nuclear layer. INL: inner nuclear layer. Outer nuclear layer thickness (ONLT) index = ONL thickness/INL thickness. *p<0.05, Student’s t-test. All data are mean ± s.e.m. Scale bar = 50 μm.
Figure 2
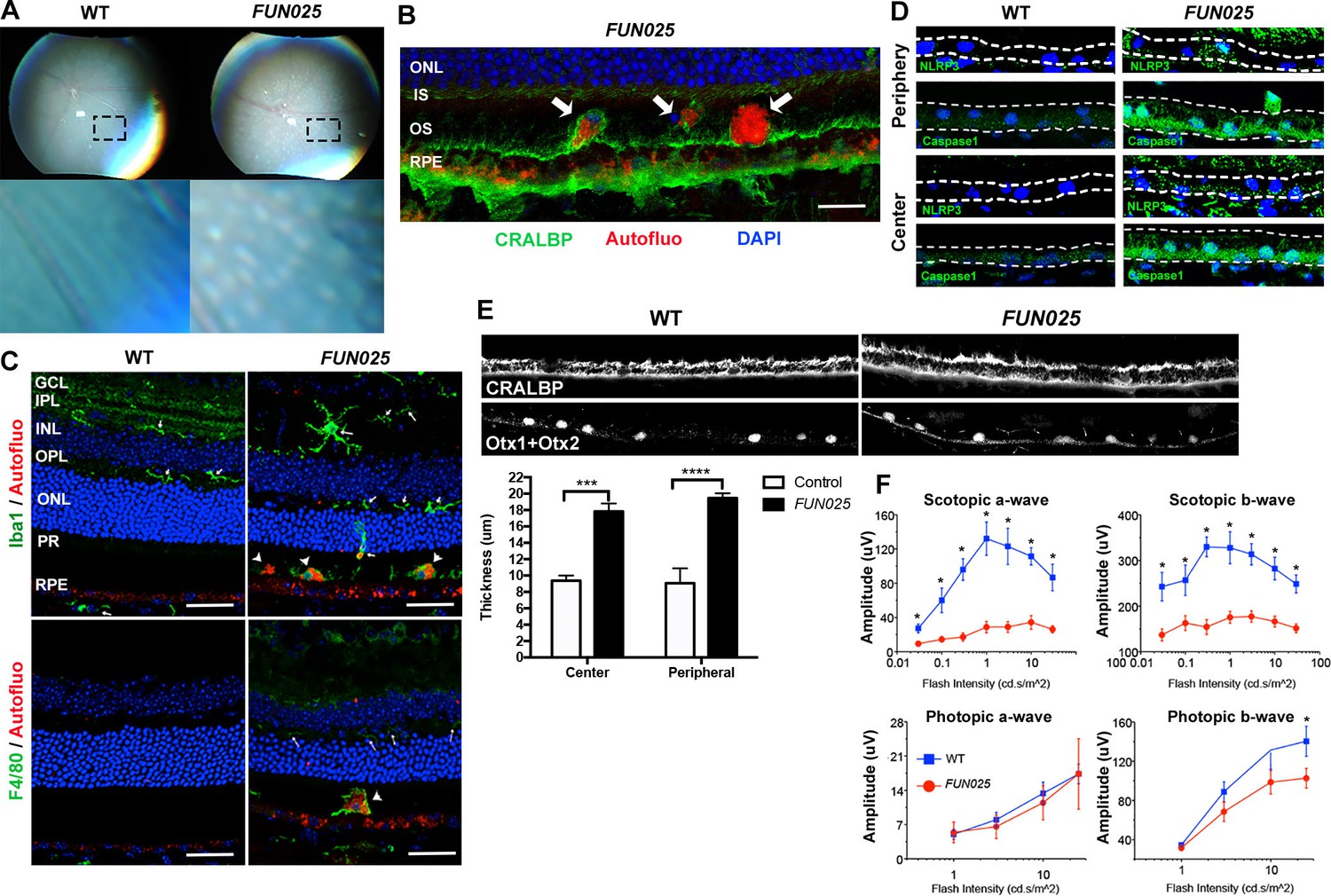
FUN025 mice show AMD-like pathologies.
(A) Punctate light deposits were found in the fundus photography of the eyes from WT and FUN025 mice. (B) Autofluorescent cells/aggregates (indicated by arrows) and lipofuscin-like autofluorescence were observed in proximity to the apical surface of the RPE in FUN025 mice. Scale bar = 60 μm. (C) Iba1 (microglia/ macrophage marker) and F4/80 (macrophage marker) positive cells were found in the FUN025 retina at seven months, whereas very few Iba1 positive cells were found in the WT retina at seven months. Scale bar = 20 μm. (D) Signals for inflammasome markers, NLRP3 and caspase1, increased in the RPE in both peripheral and central retina from FUN025 mice compared to WT control. (E) At seven months of age, the RPE (highlighted by CRALBP staining) thickness is significantly increased in both central and peripheral retina of FUN025 mice compared to control mice. The RPE nuclei were highlighted with Otx1+Otx2. Data from n = 4 mice per genotype. (F) Transcorneal electroretinograms (ERG) recordings from seven-month-old FUN025 mice and their WT littermates. Both scotopic (dark-adapted) ERG a- and b-waves from the rod pathway were markedly reduced in FUN025 mice. A reduction was also observed in photopic (light-adapted) ERG b-wave from the cone pathway with higher flash intensity, while no difference between FUN025 and WT was observed in the photopic a-wave, majority of which is postreceptoral in origin. Data from n = 5 mice per genotype. *p<0.05, two-way analysis of variance (ANOVA). All data are mean ± standard error of the mean (s.e.m.).
Figure 3 with 2 supplements
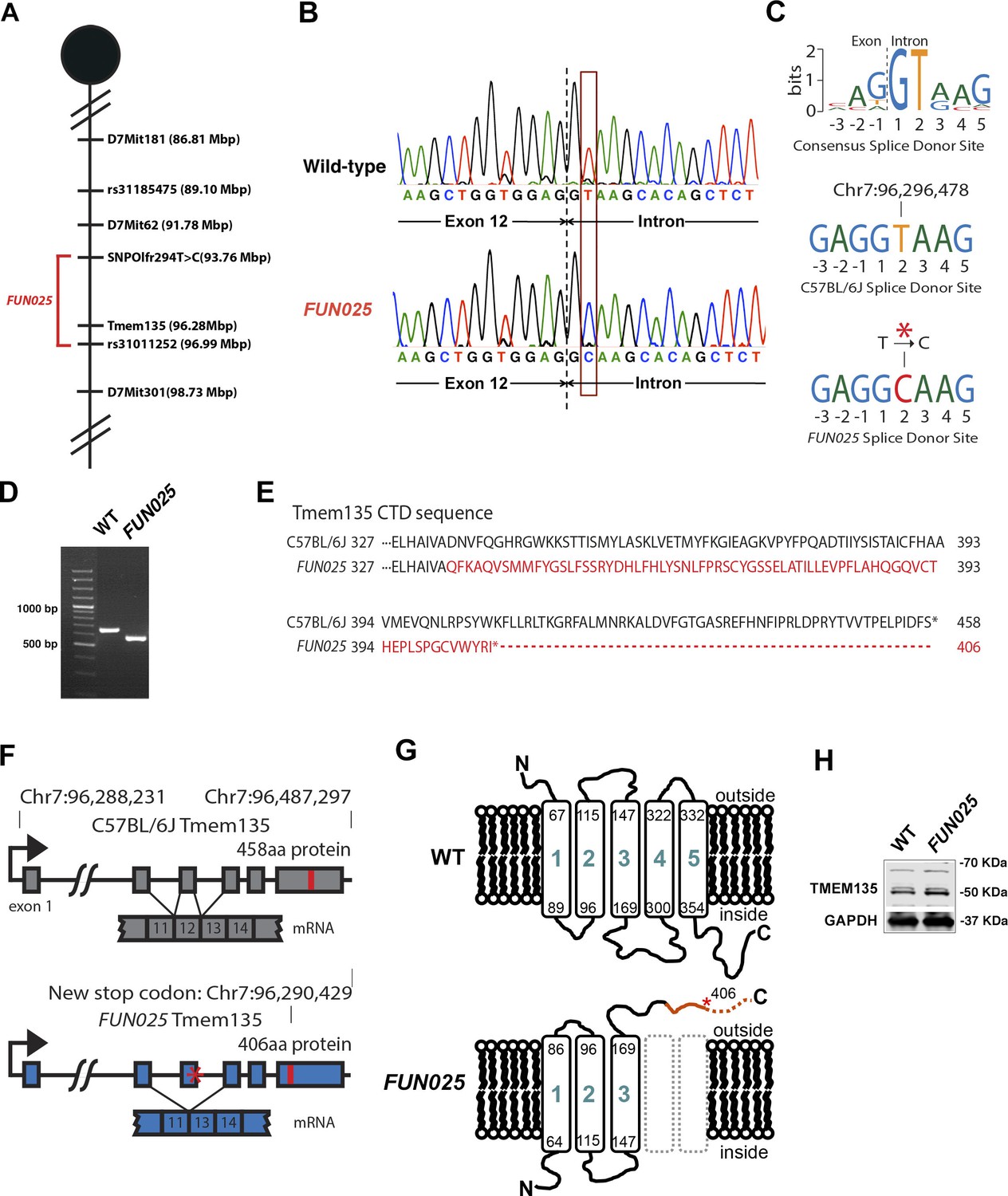
Identification of a Tmem135 mutation in FUN025 mice.
(A) Minimal genetic region of FUN025 on chromosome 7 determined by genetic mapping. (B) A point mutation (T > C) in the splice-donor site adjacent to exon 12 of Tmem135 in FUN025 mice. (C) The consensus sequence of mouse splice donor sites, depicting the necessity of the GT sequence at positions 1 and 2 downstream of the exon boundary for the functionality of the site. The C57BL/6J and FUN025 sequences are shown below, demonstrating the disrupted site in FUN025 mice. (D) RT-PCR spanning exon 12 and sequencing the product revealed the absence of this exon in the FUN025 retina. (E) Amino acid sequences of the C-terminus of TMEM135 in C57BL/6J and FUN025 mice. The WT protein is 458 amino acids long, whereas the truncated mutant protein is 406. The change in amino acid sequence is highlighted in red. (F) Consequences of the mutation in the genomic sequence of Tmem135. The mutation adjacent to exon 12 of Tmem135 (red star) results in a non-functional splice donor site, causing skipping of exon 12. This results in a frameshift and an early stop codon (chr7: 96,290,429, NCBI build 37). Locations of the stop codons are highlighted in red. (G) A predicted structure of TMEM135 having five transmembrane domains. The FUN025 mutation is predicted to result in a protein with only three transmembrane domains, whose orientation in the membrane is reversed. The rest of the c-terminal region is absent due to the early stop codon (asterisk). (H) Western blot for TMEM135 in WT and FUN025 brains. GAPDH was used as a loading control.
Figure 3—figure supplement 1

Identification of the causative gene, Tmem135, for the FUN025 mutation.
Wild-type TMEM135 protein is predicted to form five transmembrane helices by the program TMHMM (v.1.0) (http://www.cbs.dtu.dk/). The FUN025 mutation decreases the probability of forming the 4th and 5th transmembrane helix.
Figure 3—figure supplement 2
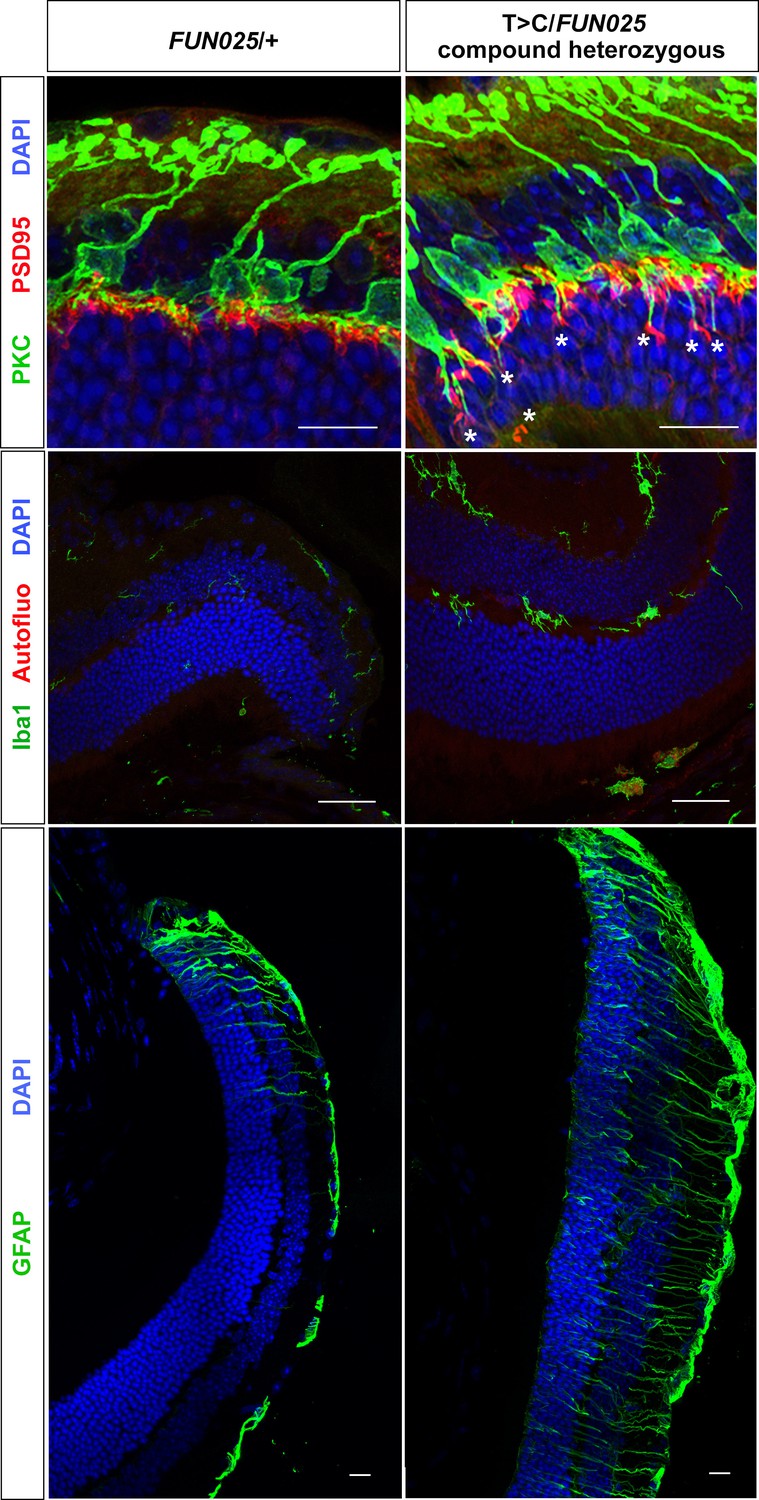
A T > C mutation in Tmem135 fails to complement FUN025.
Using the CRISPR/Cas9/sgRNA system, we generated C57BL/6J mice that carry a point mutation (T > C) in intron 12 of Tmem135 that is the same as observed in FUN025 mice (see Materials and methods). We refer to these mice as T > C /+ mice. The T > C heterozygous mice were crossed with the FUN025 homozygous mice to produce F1 (T > C/FUN025 compound heterozygous) mice, which were analyzed for retinal phenotypes. At 11 weeks, T > C/FUN025 mice showed increased ectopic synapses (upper panel), increased Iba1+ cells (middle panel), and increased GFAP immunoreactivity (lower panel) in peripheral retina compared to the age-matched FUN025 heterozygous control mice, which are similar to those observed in FUN025 homozygous mice. These results indicate non-complementation between FUN025 and T > C mutations, demonstrating that the point mutation (T > C) in Tmem135 is the FUN025 causative mutation. Scale bar = 20 μm.
Figure 4

Localization of TMEM135 to the mitochondria.
(A) Mitochondrial localization of TMEM135 in MFs co-transfected with GFP tagged TMEM135 vector (green) and DsRed2 tagged mitochondria vector (red). GFP-TMEM135 signals were detected as puncta to the mitochondria as well as in the cytoplasm. Colocalization of TMEM135 and mitochondria in MFs transfected with AcGFP1 tagged mitochondria vector (green) and immunostained with anti-TMEM135 antibody (red). Scale bar = 10 μm. (B) Colocalization of TMEM135 (anti-TMEM135 antibody, green) and mitochondira (MitoTracker, red) in wild-type and FUN025 mouse fibroblasts. Scale bar = 10 μm. (C) Immuno-EM revealed localization of GFP-tagged TMEM135 to the mitochondria. (D–E) Colocalization of TMEM135 (anti-TMEM135 antibody, green) and mitochondria (MitoTracker and TOMM20, red) in Cos-7 cells and primary mouse hippocampal neuron. Scale bar = 10 μm. (F) The mitochondrial fraction isolated from the WT mouse brain show TMEM135 signals by immunoblotting. Following proteins were used as organelle markers: MFN2–mitochondria; LAMP2–lysosome; Lamin B1– nucleus; PDI– endoplasmic reticulum (ER). (G) Strong TMEM135 signals (green) in GCL, IPL, OPL, inner segments of photoreceptor cells, and RPE from wild-type and FUN025 mouse retina. Throughout the retina, TMEM135 is colocalized with mitochondria (anti-TOMM20 antibody, red). Scale bar = 10 μm. (H) Colocalization of TMEM135 (anti-TMEM135 antibody, green) and mitochondria (MitoTracker, red) in wild-type and FUN025 primary mouse RPE cell culture. Scale bar = 5 μm.
Figure 5 with 3 supplements
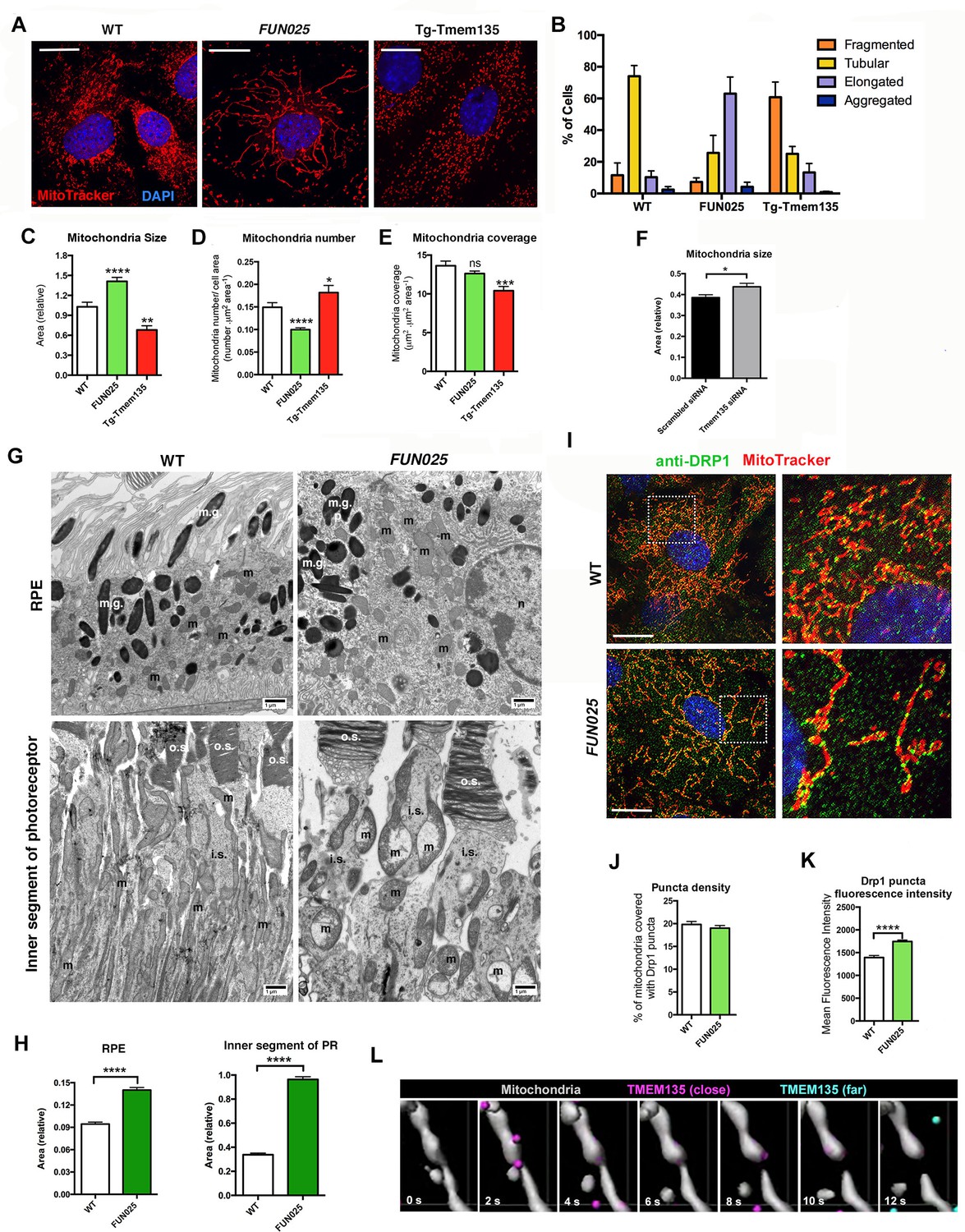
TMEM135 is involved in the balance of mitochondrial fission and fusion.
(A) Morphology of mitochondria (MitoTracker; red) in WT, Tmem135FUN025/FUN025 (FUN025) and Tg-Tmem135 MFs. Scale bar = 10 μm. (B) Scoring of mitochondrial network morphologies in WT, FUN025 and Tg-Tmem135 MFs. Data from n = 290 WT, n = 304 FUN025, and n = 372 Tg-Tmem135 cells; 3 mice per genotype. (C–E) Quantification of size, number and coverage of mitochondria in WT, FUN025 and Tg-TMEM135 MFs. Data from n = 49 WT, n = 104 FUN025, and n = 29 Tg-Tmem135 cells; 3 mice per genotype. (F) Knocking down Tmem135 by siRNA against Tmem135 results in increased mitochondrial size in WT MFs. Data from n = 72 cells with scrambled siRNA, n = 81 cells with Tmem135 siRNA. Two tailed, unpaired, Student’s t-test. (G) EM revealed the morphology of mitochondria in RPE and inner segments of photoreceptor cells from wild-type and FUN025 mice. m = mitochondria; m.g. = melanin granules; n = necleus; o.s. = outer segments; i.s. = inner segments. Scale bar = 1 μm. (H) Quantification of mitochondria size in RPE and inner segments of photoreceptor cells from wild-type and FUN025 mice. RPE data from 300 mitochondria from three wild-type mice and 400 mitochondria from four FUN025 mice. Inner segments data from 200 mitochondria from four wild-type mice and 200 mitochondria from four FUN025 mice. (I) Immunofluorescence of DRP1 (anti DRP1 antibody, green) and mitochondria (MitoTracker, red) in WT and FUN025 MFs. Scale bar = 10 μm. (J–K) Quantification of DRP1 puncta density and fluorescence intensity on mitochondria. (L) Time-lapse fluorescence imaging (modified as described in Materials and methods) of a living HT22 cell expressing TMEM135-GFP and labeled with MitoTracker at the indicated time points. TMEM135 puncta that are located relatively close to the mitochondrial surface are shown in magenta, whereas those relatively far from the mitochondrial surface are shown in turquoise. Mitochondria (gray) was made partial transparent in order to see TMEM135 puncta located on the back of mitochondria. *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001, two-way ANOVA. All data are mean ± s.e.m.
Figure 5—figure supplement 1

Generation of Tg-TMEM135 mice.
(A) Transgenic construct for generating the transgenic mice overexpressing Tmem135 (Tg-TMEM135). (B) Western blot analysis of TMEM135 expression in WT and Tg-TMEM135 (TG) mouse brain. GAPDH was used as a loading control.
Figure 5—figure supplement 2
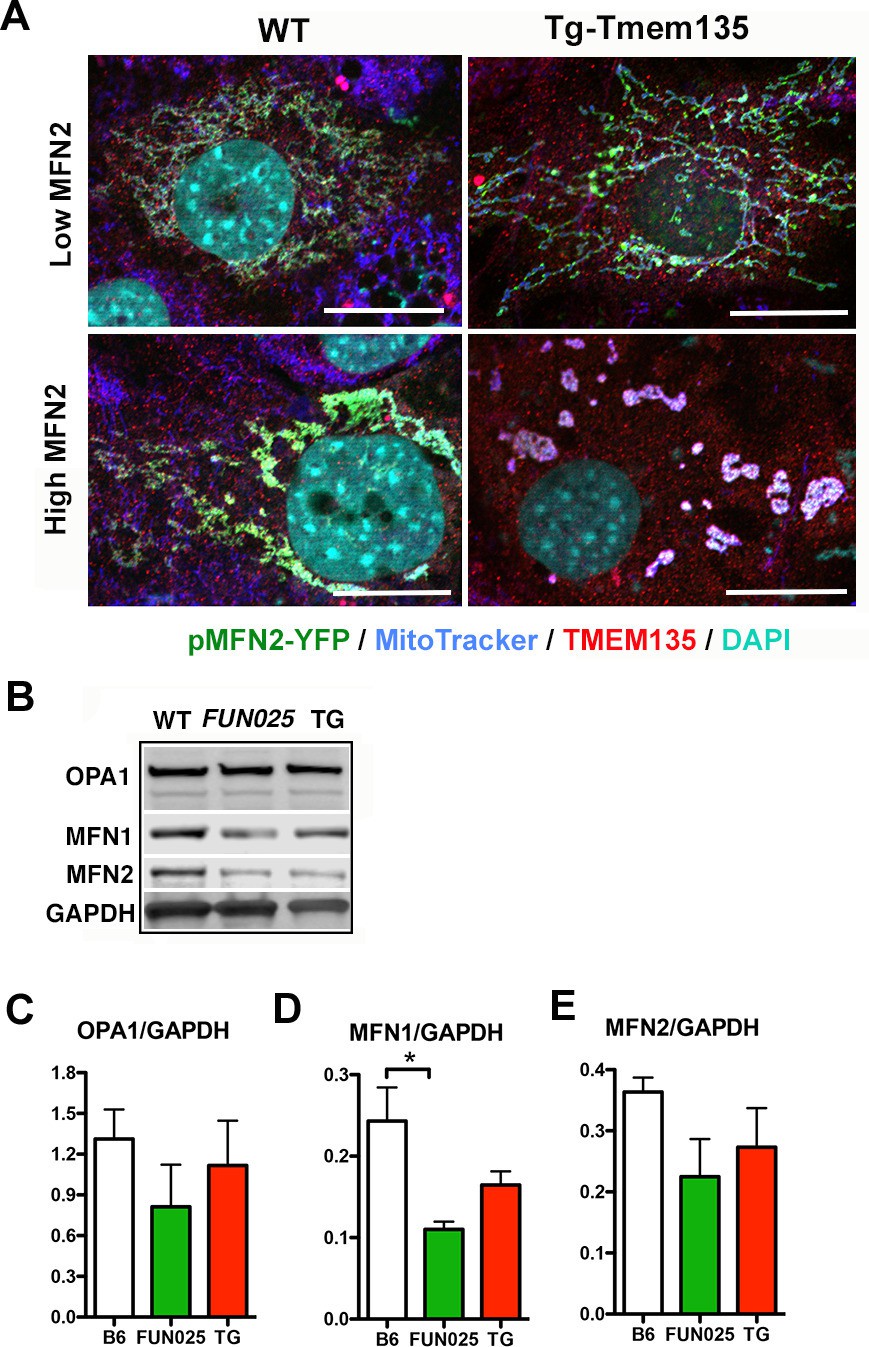
TMEM135 does not inhibit mitochondrial fusion.
(A) WT and Tg-Tmem135 MFs were transfected with expression vectors encoding MFNs-YFP (green) at low and high levels and immunostained with TMEM135 antibody (red). Mitochondria were highlighted by MitoTracker (blue). Nuclei were labeled by DAPI (cyan). Scale bar = 10 μm. (B) Western blot analysis of OPA1, MFN1, and MFN2 in WT, FUN025 and Tg-TMEM135 (TG) fibroblasts. GAPDH was used as a loading control. (C–E) Quantification of western blotting results from (B). Protein levels were normalized using GAPDH expression levels.
Figure 5—figure supplement 3

A portion of TMEM135 is colocalized with DRP1 on the mitochondria.
Confocal fluorescence microscopy of TMEM135 localization (red) and DRP1 localization (green) on mitochondria (blue). Some of mitochondrial-bound DRP1 puncta colocalize with mitochondrial-bound TMEM135 puncta indicated by arrows. Scale bar = 2 μm.
Figure 6 with 2 supplements
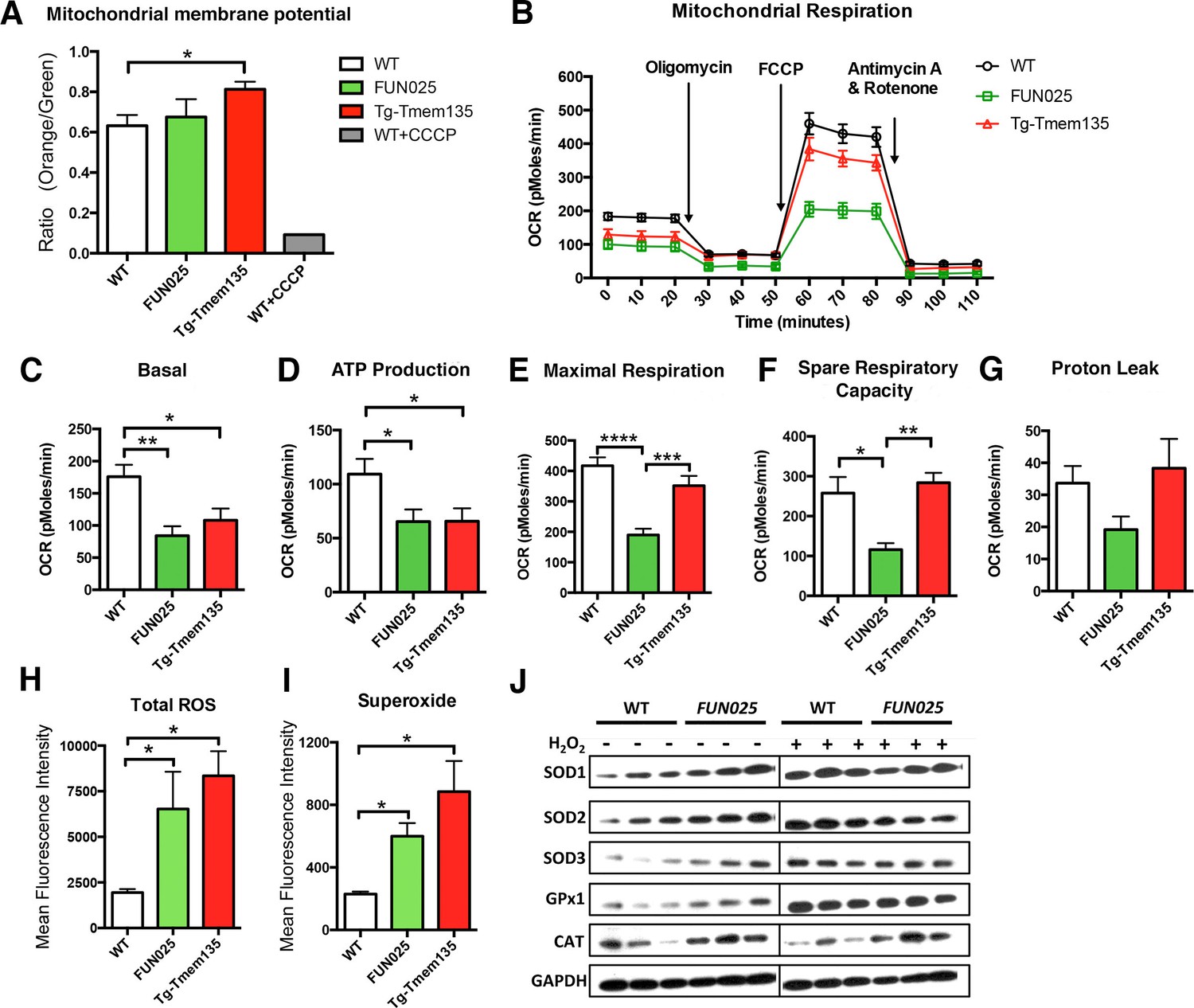
Tmem135 plays a role in mitochondrial metabolism.
(A) Mitochondrial membrane potential (ΔΨM) in Tg-Tmem135 MFs was higher than that in WT MFs, whereas ΔΨM in Tmem135FUN025/FUN025 (FUN025) MFs was comparable to that in WT MFs. Data from n = 3 mice per genotype. n = 1 for WT+CCCP. (B–G) Oxygen consumption rates (OCR) of MFs were determined with a Seahorse XFe24 Extracellular Flux Analyzer in basal and stimulated conditions (n = 3 mice per genotypes). The areas under the curve from different sections of the experiment (B) are shown as individual histograms for basal respiration, ATP production, maximal respiration, spare respiratory capacity, and proton leak (C–G). Data from n = 3 mice per genotype. (H–I) Total ROS and superoxide in FUN025 and Tg-Tmem135 MFs were higher compared to WT cells. Data from n = 3 mice per genotype. (J) Western blot analysis for SOD1, SOD2, SOD3, GPx1and CAT in WT and FUN025 MFs with and without the hydrogen peroxide treatment. Protein levels were normalized using GAPDH expression levels. *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001, two-way ANOVA. All data are mean ± s.e.m..
Figure 6—figure supplement 1

Mitochondrial membrane potential in FUN025 and Tg-Tmem135 MFs.
(A–D) Representative flow cytometry data showing ΔΨM differences between MFs derived from WT (A), FUN025 (B), and Tg-Tmem135 (C) mice. CCCP treatment caused decrease of fibroblast ΔΨM and therefore serves as the negative control for fibroblast ΔΨM (D). Unstained fibroblasts were used to gate the quadrants 1–4 (Q1-Q4) which Q4 represents background fluorescence intensity. (D) Population with detectable ΔΨM was gated according to CCCP treated WT MFs.
Figure 6—figure supplement 2
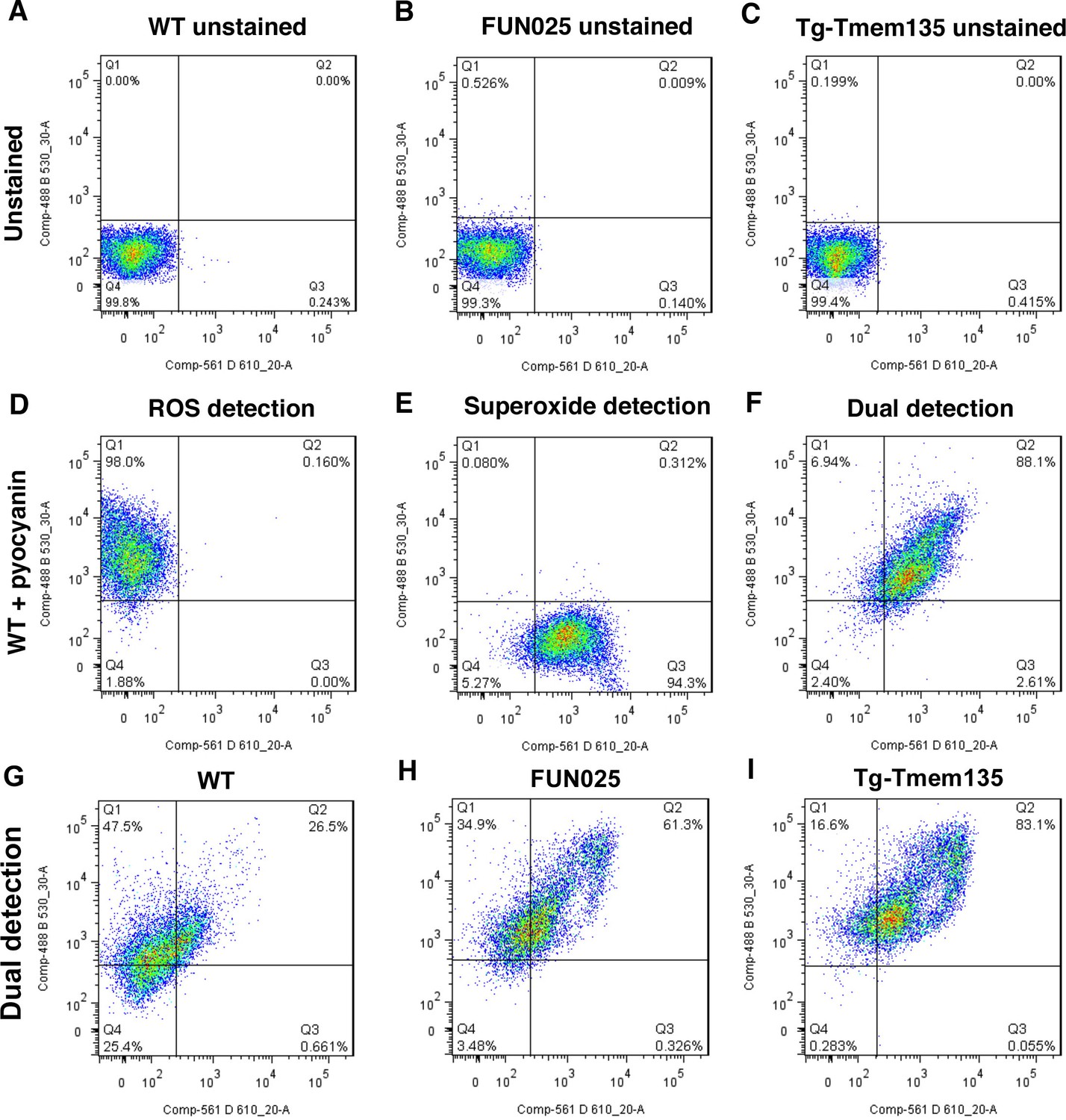
Increased ROS and superoxide in FUN025 and Tg-Tmem135 MFs.
Representative flow cytometry data of ROS and superoxide levels in MFs derived from WT (A,D,G), FUN025 (B,C,H), and Tg-Tmem135 (C,F,I) mice. Panels A–C show unstained controls for each condition. WT MFs treated with pyocyanin were used as a positive control for ROS and superoxide detection (D–F). Both FUN025 (H) and Tg-Tmem135 (I) MFs showed increased levels of ROS and superoxide when compared with WT MFs (G).
Figure 7
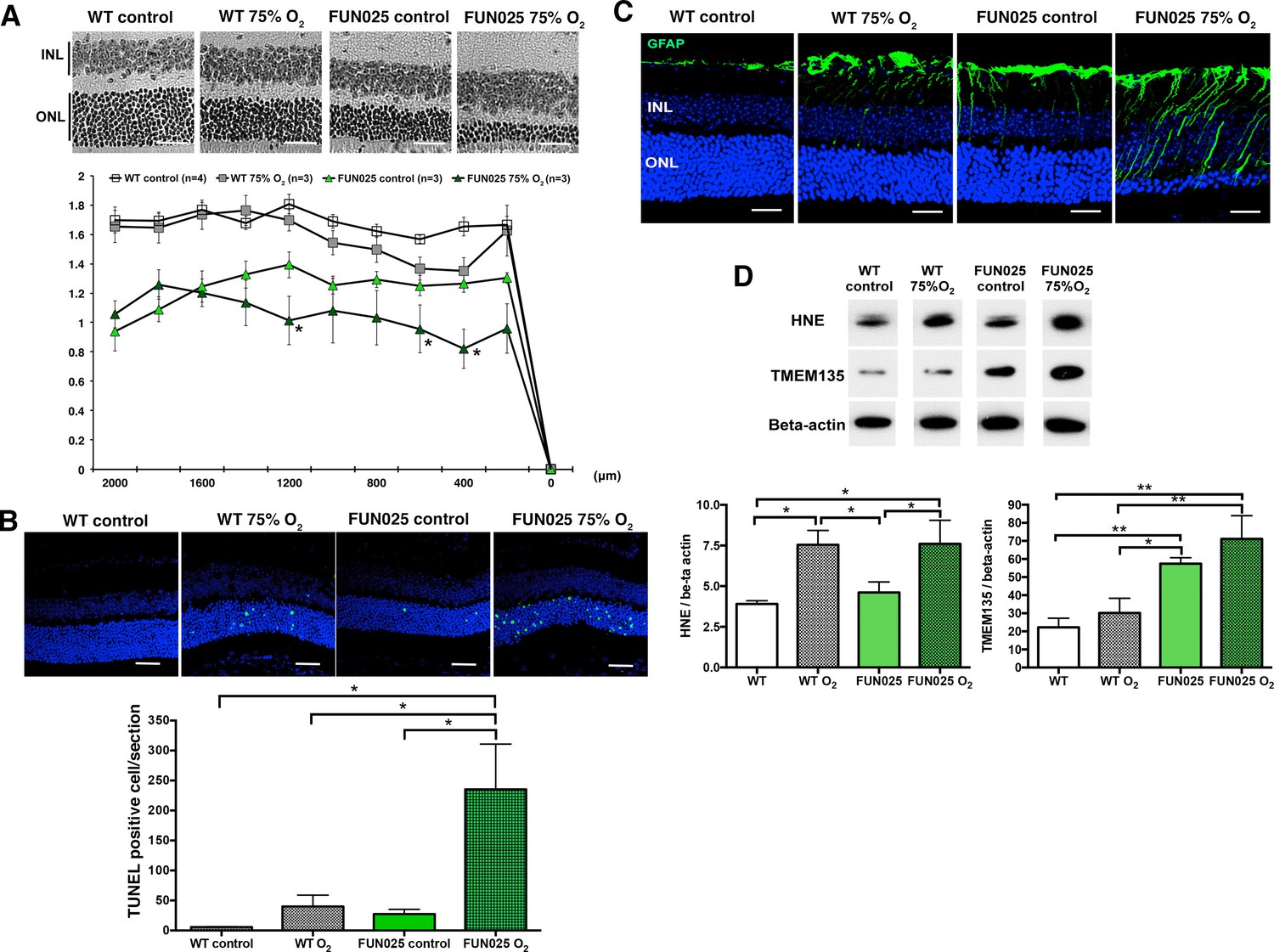
Tmem135FUN025/FUN025 mice are more sensitive to hyperoxic condition.
(A–B) Tmem135FUN025/FUN025 (FUN025) mice raised in 75% O2 for two weeks show significant decrease of ONLT and increase of TUNEL positive cells, indicating accelerated photoreceptor cell death by apoptosis. Data from n = 4 WT mice in control air, n = 3 WT mice in 75% O2, n = 3 FUN025 mice in control air, and n = 3 FUN025 mice in 75% O2. Scale bar = 20 μm. (C) Upregulation of GFAP (green) indicating retinal stress is observed in FUN025 mice raised in the normal air, as well as WT mice and FUN025 mice raised in 75% O2 for two weeks. FUN025 mice raised in 75% O2 have the highest increase of GFAP signals in the retina. Scale bar = 20 μm. (D) Western blotting showing that hyperoxia results in upregulation of 4-HNE in both WT and FUN025 retina, and an increase of TMEM135 in FUN025 retina but not in WT retina. Data from n = 3 WT mice in control air, n = 3 WT mice in 75% O2, n = 3 FUN025 mice in control air, and n = 3 FUN025 mice in 75% O2. *p<0.05, **p<0.01, two-way ANOVA. All data are mean ± s.e.m.
Videos
Video 1
Live imaging of HT22 cells expressing TMEM135-GFP.
Live imaging of a HT22 cell expressing TMEM135-GFP and labeled with MitoTracker Red. TMEM135 puncta that are located relatively close to the mitochondrial surface are shown in magenta, whereas those relatively far from the mitochondrial surface are shown in turquoise. Mitochondria (gray) was made partial transparent in order to see TMEM135 puncta located on the back of mitochondria.
Video 2
Live imaging of WT fibroblasts expressing TMEM135-eGFP.
Live imaging of fibroblasts derived from B6 (WT) mice transfected with TMEM135-eGFP plasmid and labeled with MitoTracker Red. TMEM135-eGFP are shown in green and mitochondria are shown in red.
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Mouse Tmem135 mutation reveals a mechanism involving mitochondrial dynamics that leads to age-dependent retinal pathologies
eLife 5:e19264.
https://doi.org/10.7554/eLife.19264




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}