Spatial patterning of P granules by RNA-induced phase separation of the intrinsically-disordered protein MEG-3
- Howard Hughes Medical Institute, Johns Hopkins University School of Medicine, United States
Figures
Figure 1 with 1 supplement
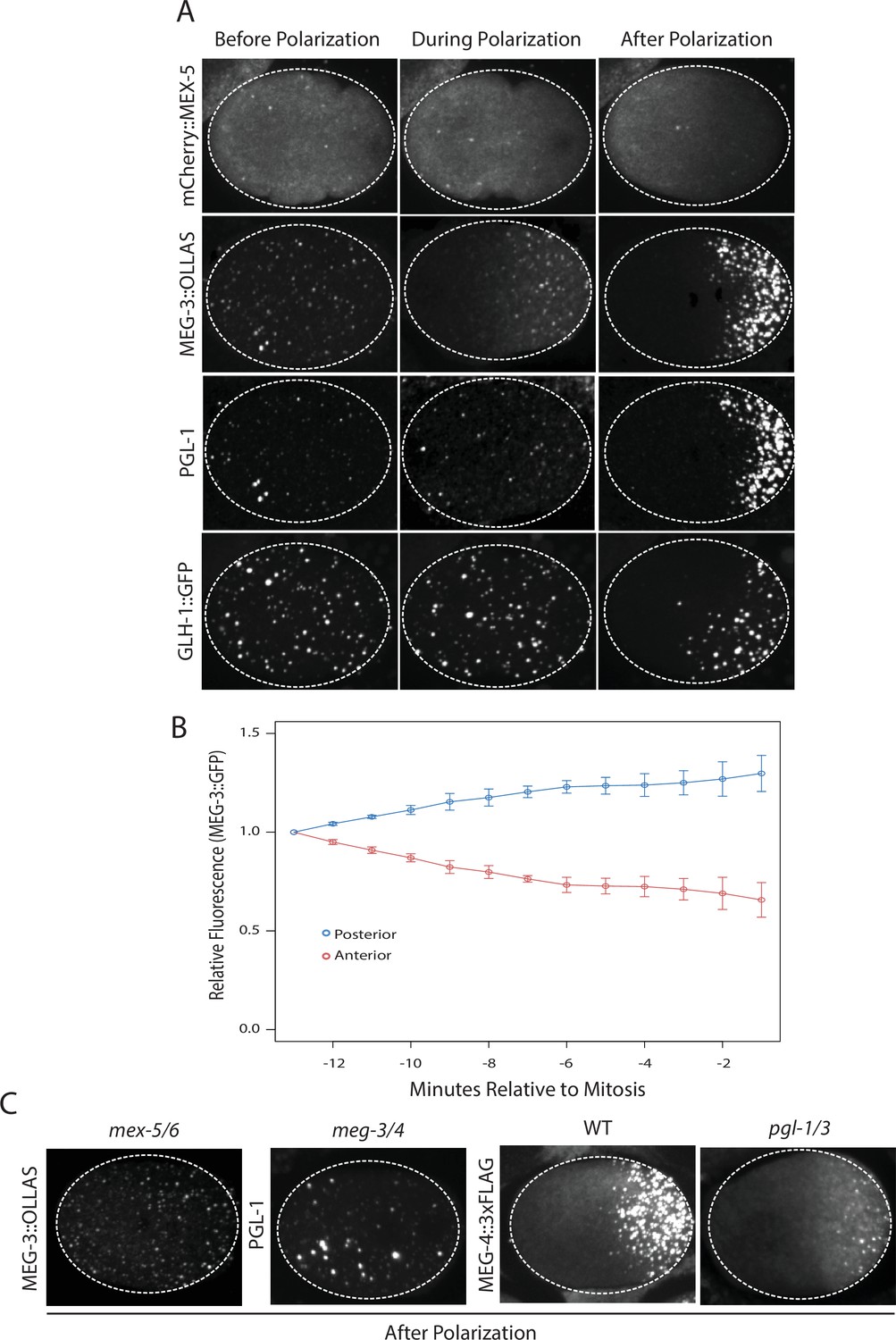
Localization of P granule proteins during zygote polarization.
(A) Photomicrographs of live wild-type (mCherry::MEX-5 and GLH-1::eGFP) or fixed meg-4 zygotes (MEG::3 OLLAS and PGL-1) at three different stages: before polarization (pronuclear formation), during polarization (pronuclear migration) and after polarization (mitosis). meg-4(ax3052) zygotes were co-immunostained for MEG-3::OLLAS (anti-OLLAS, Novus Biological), and PGL-1 (K76, DSHB). meg-4 is required redundantly with meg-3 for P granule assembly, and each is sufficient to support localized granule assembly (Wang et al., 2014). In this and subsequent figures, dashed lines outline each embryo, embryos are oriented with anterior to the left and posterior to the right and are ~50 μM long. At least three embryos were examined per genotype shown. (B) MEG-3::meGFP levels in the anterior and posterior halves of the zygote during polarization. Values represent average fluorescence intensity over time (relative to initial levels) in the anterior (red) and posterior (blue). Averages come from values measured from three different embryos. Error bars represent standard deviation of the mean. (C) Photomicrographs of fixed zygotes after polarization immunostained for OLLAS, PGL-1, or FLAG. mex-5/6 zygotes were derived from wild-type hermaphrodites treated with mex-5 and mex-6 RNAi. meg-3/4 zygotes were derived from meg-3(ax3055); meg-4(ax3052) hermaphrodites. pgl-1/3 zygotes were derived from pgl-3(bn104) hermaphrodites treated with pgl-1 RNAi (see Figure 1—figure supplement 1 for additional examples of pgl-1(RNAi);pgl-3(bn104) zygotes also stained for PGL-1 to verify loss of PGL-1).
Figure 1—figure supplement 1
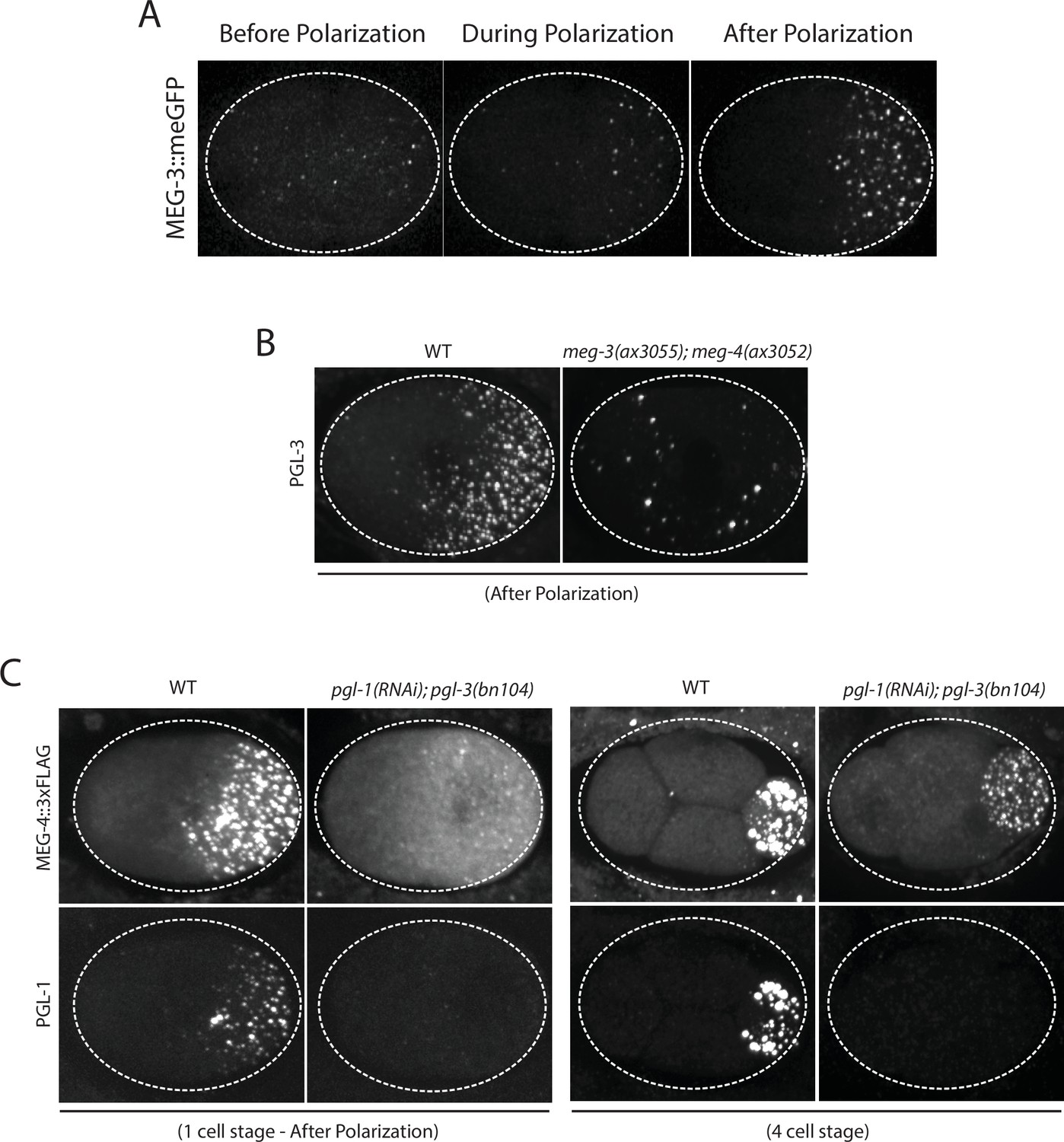
MEG-3 localizes before PGL-1 and does not require PGL-1 or PGL-3 to assemble granules.
(A) Photomicrographs of live MEG-3::meGFP in zygotes at three different stages: before polarization (pronuclear formation), during polarization (pronuclear migration) and after polarization (mitosis). These and similar images taken from 3 zygotes at 14 time points were used to quantify MEG-3::meGFP fluorescence levels over time as shown in Figure 1B. (B) Photomicrographs of fixed wild-type and meg-3/4 zygotes after polarization immunostained for PGL-3. meg-3/4 zygotes were derived from meg-3(ax3055); meg-4(ax3052) hermaphrodites. PGL-3 staining was done using the KT3 antibody (DSHB). (C) Photomicrographs of fixed zygotes stained with anti-FLAG (top) and K76 anti-PGL-1 (bottom) to show MEG-4 localization at the 1-cell (left) and 4-cell (right) stages. pgl-1/3 zygotes were derived from pgl-3(bn104); meg-4(ax2080FLAG tag) hermaphrodites treated with pgl-1 RNAi.
Figure 2 with 1 supplement

MEX-5 is necessary and sufficient to disassemble MEG-3 granules in vivo.
(A) Photomicrographs of mCherry tagged live zygotes [wild-type and mex-5(S404A)] or fixed zygotes expressing MEX-5 tagged with an OLLAS epitope [mex-5(ZF-);mex-6(RNAi)] at pronuclear meeting to show MEX-5 localization. Wild-type MEX-5 is in an anterior-rich gradient, whereas MEX-5(S404A) and MEX-5(ZF-) are uniformly distributed. Numbers indicate number of zygotes exhibiting phenotype shown / total number of zygotes examined. (B) Photomicrographs of live embryos expressing MEG-3::meGFP. Genotypes at the mex-5 locus are as indicated. Numbers indicate the numbers of zygotes examined as in A. In 1/10 mex-5(ZF-, S404A); mex-6(RNAi) zygotes, MEG-3 granules were asymmetric possibly due to incomplete depletion of MEX-6 by RNAi.
Figure 2—figure supplement 1
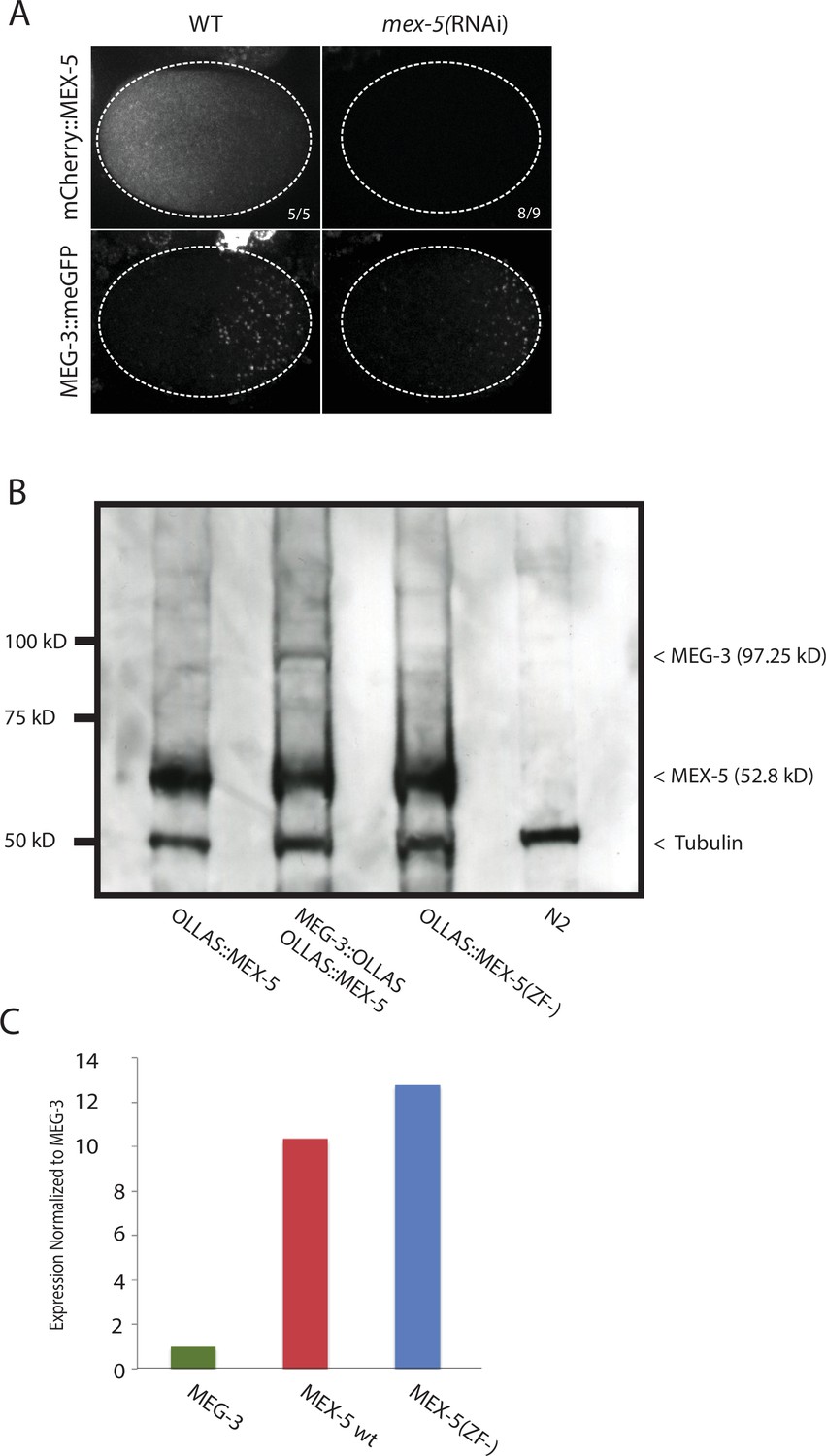
Expression of MEX-5, and MEX-5(ZF-), MEG-3, and MEG-3IDR.
(A) MEX-5 functions redundantly with MEX-6 to localize MEG-3: Photomicrographs of live wild-type and mex-5(RNAi) zygotes co-expressing mCherry:MEX-5 and MEG-3::meGFP. Loss of mCherry fluorescence in the mex-4(RNAi) zygotes was used to confirm efficient loss of mCherry::MEX-5. Numbers indicate number of zygotes exhibiting phenotype shown / total number of zygotes examined. (B) Western blot of embryo lysates co-blotted with α OLLAS and α tubulin (loading control). Expected sizes for protein-epitope fusions are indicated on the right by a < symbol. N2 lysate is a control lysate expressing no tagged proteins, all others are lysates from embryos expressing tagged proteins as indicated. (C) Bar graph showing relative expression of MEG-3 and MEX-5. Western blot in Figure 2—figure supplement 1 was quantified using Image J. MEX-5 is 10X more abundant than MEG-3.
Figure 3 with 1 supplement
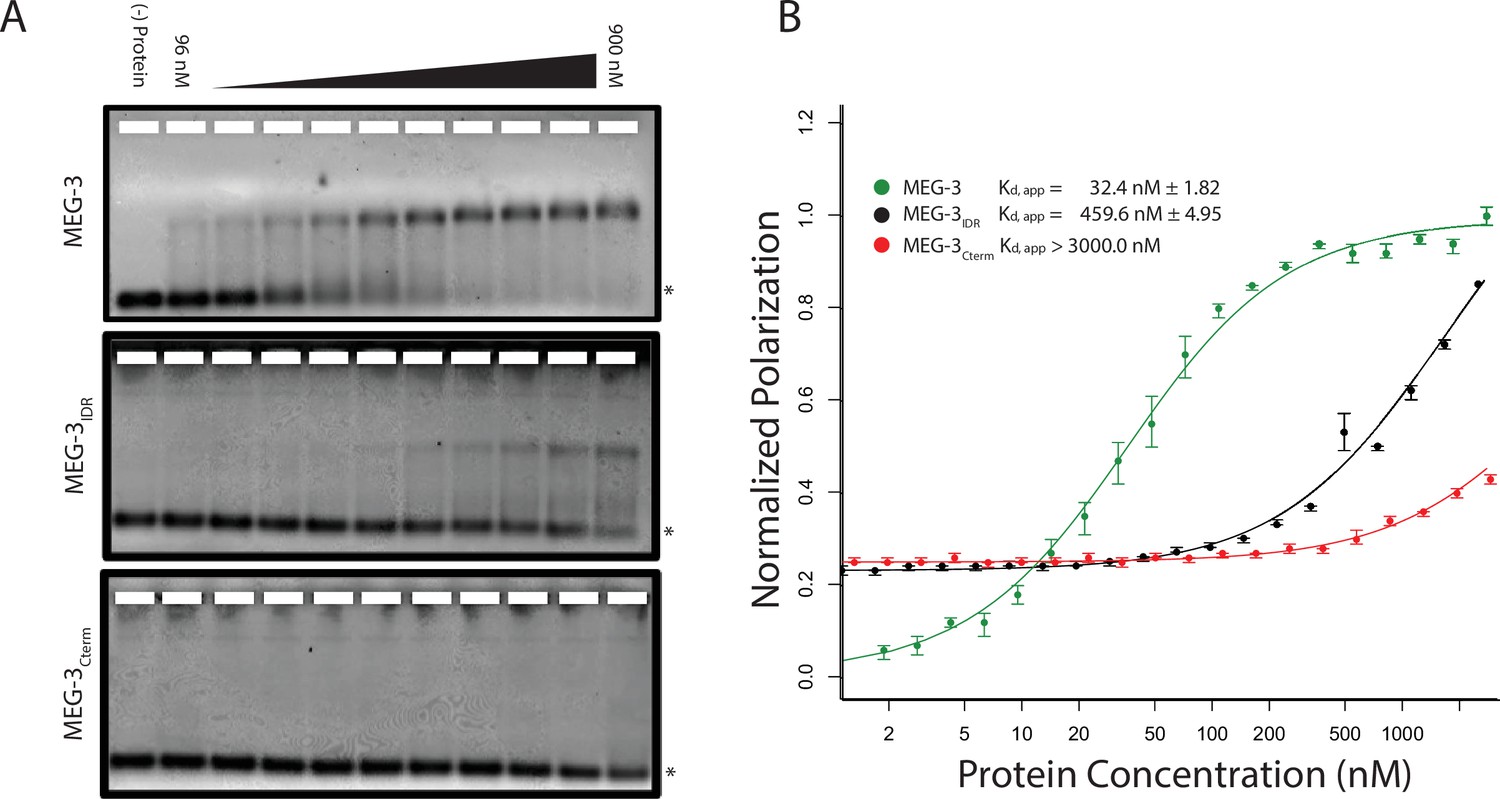
MEG-3 Binds RNA in vitro.
(A) Binding of MEG-3 to poly-uridine 30 (poly-U30) is shown by electrophoretic mobility shift assay (EMSA) using fluorescein-labeled poly-U. EMSAs are shown for (top to bottom) full length MEG-3, MEG-3IDR, and MEG-3Cterm. Unbound poly-U30 is denoted by an asterisk (*). For each image shown, n ≥ 3 technical replicates. (B) Fluorescence Polarization of poly-U30 by MEG-3. Fluorescence polarization values normalized relative to saturation are shown for full length MEG-3 (green), MEG-3IDR (black), and MEG-3Cterm (red). Values represent averages of ≥3 technical replicates. A fit of the polarization as a function of protein concentration is plotted and used to calculate the given Kd,app. Error bars report S.E.M. Expanded graphs are shown in Figure 3—figure supplement 1.
Figure 3—figure supplement 1
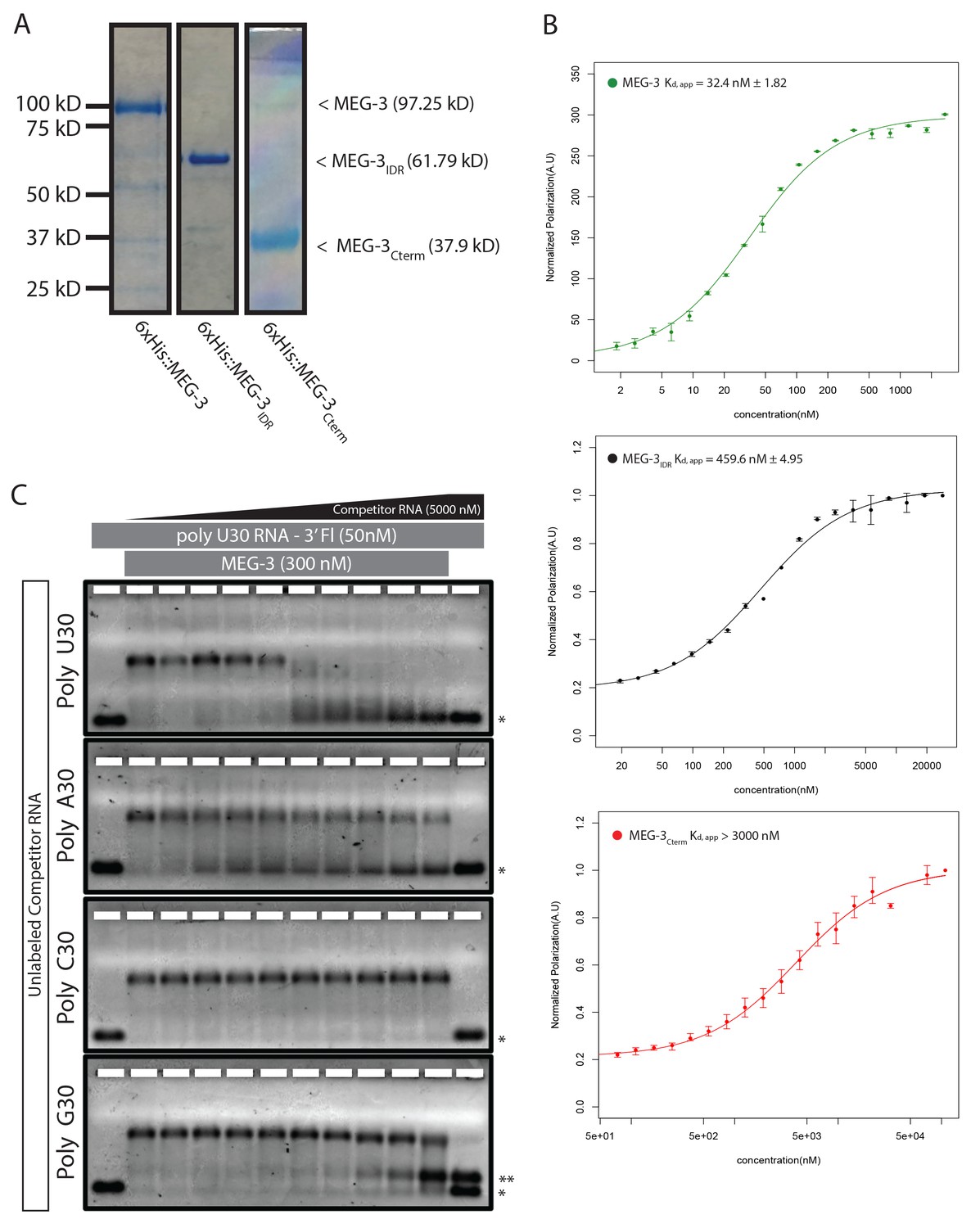
MEG-3 RNA binding.
(A) Coomassie stained gel showing His-tagged MEG-3 proteins. Expected sizes are indicated on the right. Each lane is from a separate gel. (B) RNA-RNA competition EMSA. 300 nM MEG-3 (300 nM) was incubated with 50 nM poly-U30 and increasing amounts of unlabeled 30-mer RNAs as indicated to the left of each panel. The first lane of each gel contains only labeled poly-U RNA. The last lane of each gel contains poly-U RNA and the highest concentration of unlabeled competitor RNA (5000 nM). The unbound poly-U RNA is denoted by an asterisk (*). Double asterisks (**) in the poly-G assay denote a band that arises due to an interaction between poly-G and poly-U (independent of MEG-3). For each image shown, n = 2 technical replicates. (C) Fluorescence Polarization of polyuridine by MEG-3 (expanded from Figure 3). Fluorescence polarization values normalized relative to saturation are shown for full length MEG-3 (top, green), MEG-3IDR (middle, black), and MEG-3Cterm (bottom, red). Values represent averages of ≥3 technical replicates. A fit of the polarization as a function of protein concentration is plotted and used to calculate the given Kd,app. Error bars report S.E.M.
Figure 4 with 2 supplements

Stimulation of MEG-3 phase separation by RNA.
(A) Bar graph showing the number of condensates formed by 0.5 μM MEG-3 or MEG-3IDR in the presence of increasing poly-U30. Values represent averages from three technical replicates. Error bars indicate S.E.M. (B) Photomicrographs of phase separation assay showing condensate formation of 5 μM full length MEG-3 incubated with poly-U RNA and/or MEX-5 as indicated.(C) Violin plots showing condensate size and number for each experiment represented in B. The height of the plot shows the area of condensates in μm2. The width of the plot correlates to the proportion of condensates of that size. Numbers inside each violin plot are the total number of condensates pooled from three technical replicates for each condition. An additional control is shown in Figure 4—figure supplement 2.
Figure 4—figure supplement 1

Phase Separation assay titration.
DIC images of MEG-3 solutions in the presence of increasing concentrations of protein and RNA. MEG-3 was diluted into phase separation buffer (25 mM HEPES, pH7.4, NaCl adjusted to a final concentration of 150 mM) at room temperature. The solution was transferred to a glass bottom dish and the dish surface was photographed using an inverted DIC microscope 10 min after the initial dilution (room temperature).
Figure 4—figure supplement 2
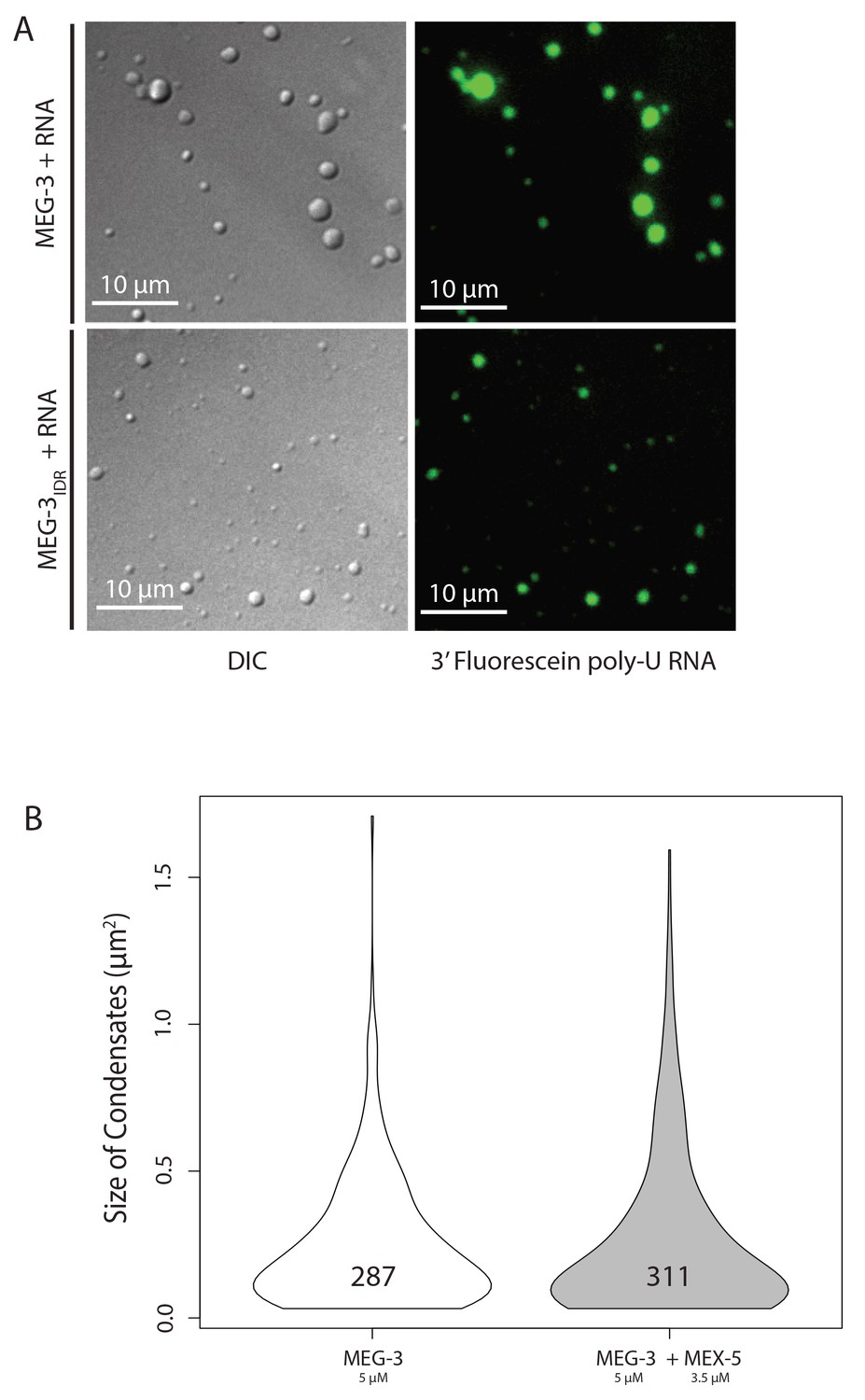
Phase Separation assay controls.
(A) Same as above except 3’ fluorescein labeled poly-U RNA was included in the buffer. DIC images (left) show the MEG-3 condensates and fluorescence images (488 channel) show the 3’ fluorescein labeled poly-U RNA concentrated in the condensates. (B) Violin plots showing condensate size and number comparing MEG-3 alone and MEG-3 + MEX-5 (Additional data). As in Figure 4C, the height of the plot shows the area of condensates in μm2. The width of the plot correlates to the proportion of condensates of that size. Numbers inside each violin plot are the total number of condensates pooled from three technical replicates for each condition. Data for the MEG-3 alone sample are the same as in Figure 4C and shown here only for reference.
Figure 5 with 1 supplement
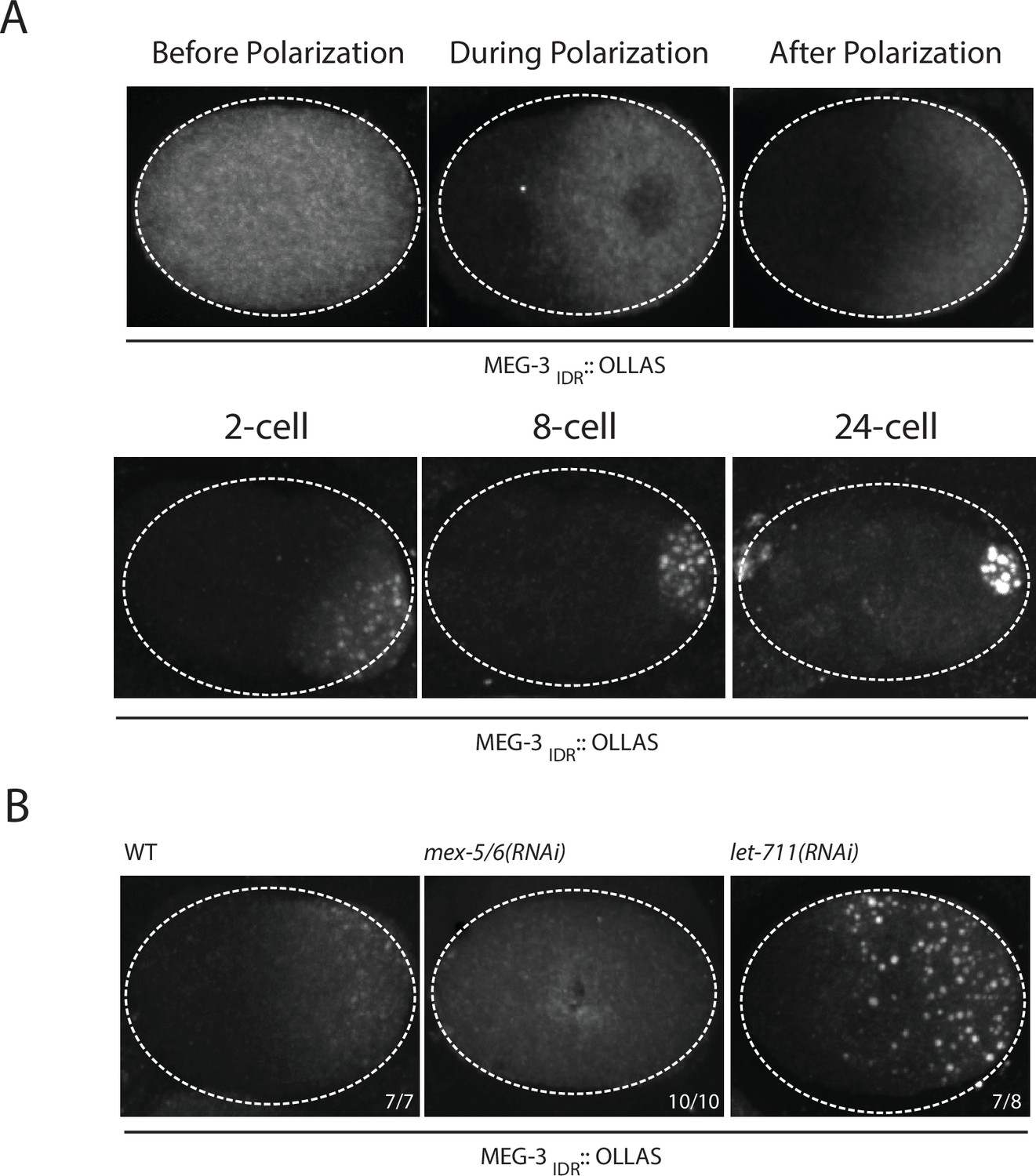
Coalescence of MEG-3IDR can be stimulated by blocking mRNA turnover in vivo.
(A) Photomicrographs of fixed embryos expressing MEG-3IDR tagged with OLLAS epitope. First row are zygotes (one-cell stage) and second row are later stage embryos as indicated. (B) Photomicrographs of fixed zygotes expressing MEG-3IDR tagged with OLLAS epitope. Genotypes are indicated above each embryo (left to right: wild-type, mex-5/6(RNAi), and let-711(RNAi)). Numbers indicate number of zygotes exhibiting phenotype shown / total number of zygotes examined. In 1/8 let-711(RNAi) zygotes, MEG-3IDR formed granules but these were smaller and confined to the posterior half of the zygote, possibly due to incomplete depletion of let-711.
Figure 5—figure supplement 1
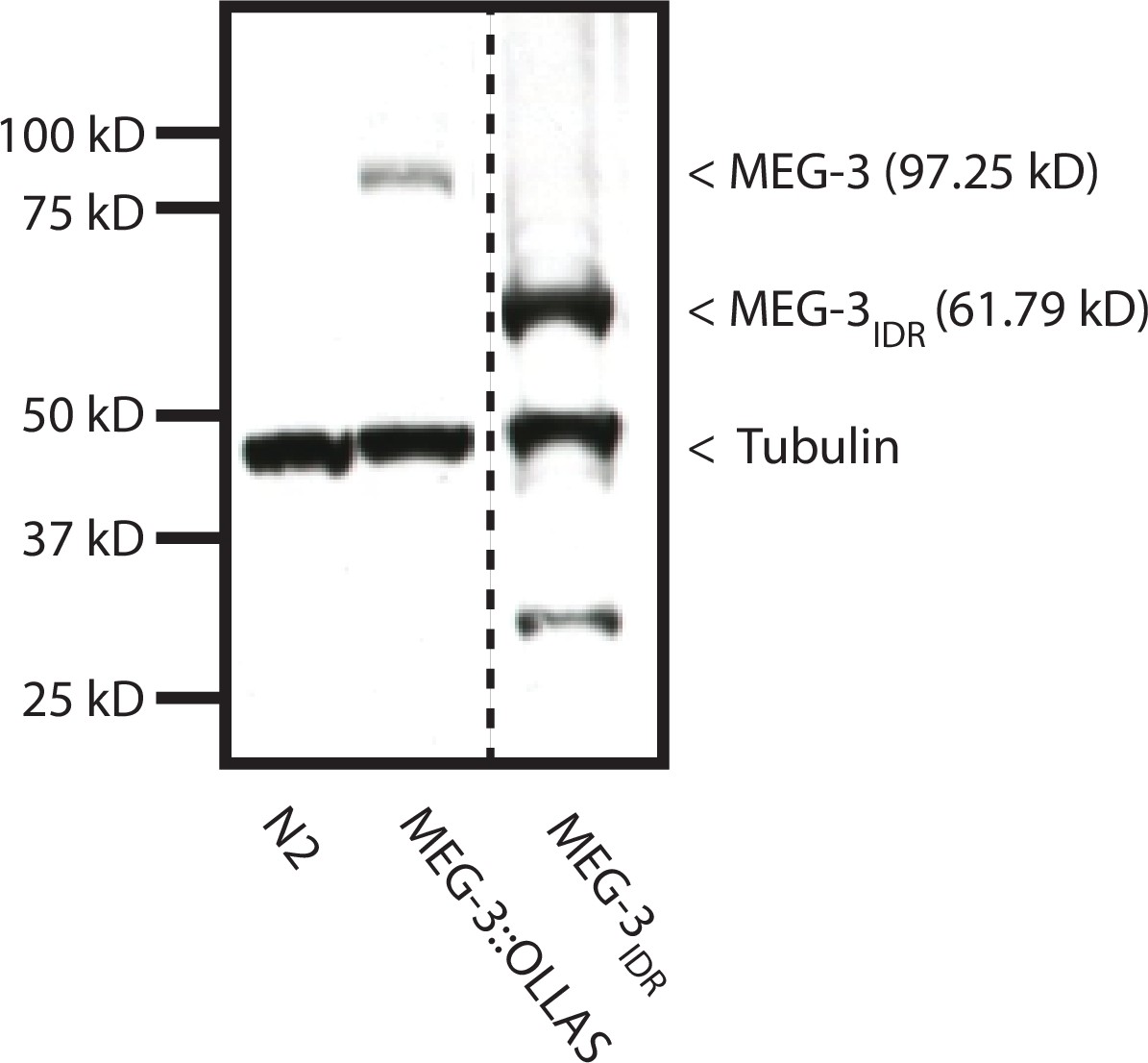
MEG-3IDR in vivo.
Western blot of mixed-stage embryo lysates co-blotted with α OLLAS and α tubulin. Expected sizes for protein-epitope fusions are indicated on the right by a < symbol. Dashed line indicates a break in the original gel. Genotype is indicated on the bottom. The MEG-3IDR is present at higher level than MEG-3, likely due to the fact that unlike MEG-3, MEG-3IDR persists longer in somatic cells (see Figure 5 and data not shown). The significance of this difference is not known. n = 3 technical replicates.
Additional files
-
Supplemental file 1
Strains used in this study.
All strains were generated in this study by genome editing or crossing. No transgenic lines were used. Independent edits displayed the same phenotypes. The mex-5(S404A) lines could not be maintained due to semi-dominant maternal-effect sterility (91.6%) and recessive maternal-effect lethality (100%).
- https://doi.org/10.7554/eLife.21337.014
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Spatial patterning of P granules by RNA-induced phase separation of the intrinsically-disordered protein MEG-3
eLife 5:e21337.
https://doi.org/10.7554/eLife.21337




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}