The synaptonemal complex has liquid crystalline properties and spatially regulates meiotic recombination factors
- University of California, Berkeley, United States
- Howard Hughes Medical Institute, United States
- University of Utah, USA
- Lawrence Berkeley National Laboratory, United States
- California Institute for Quantitative Biosciences, United States
Figures
Figure 1 with 2 supplements
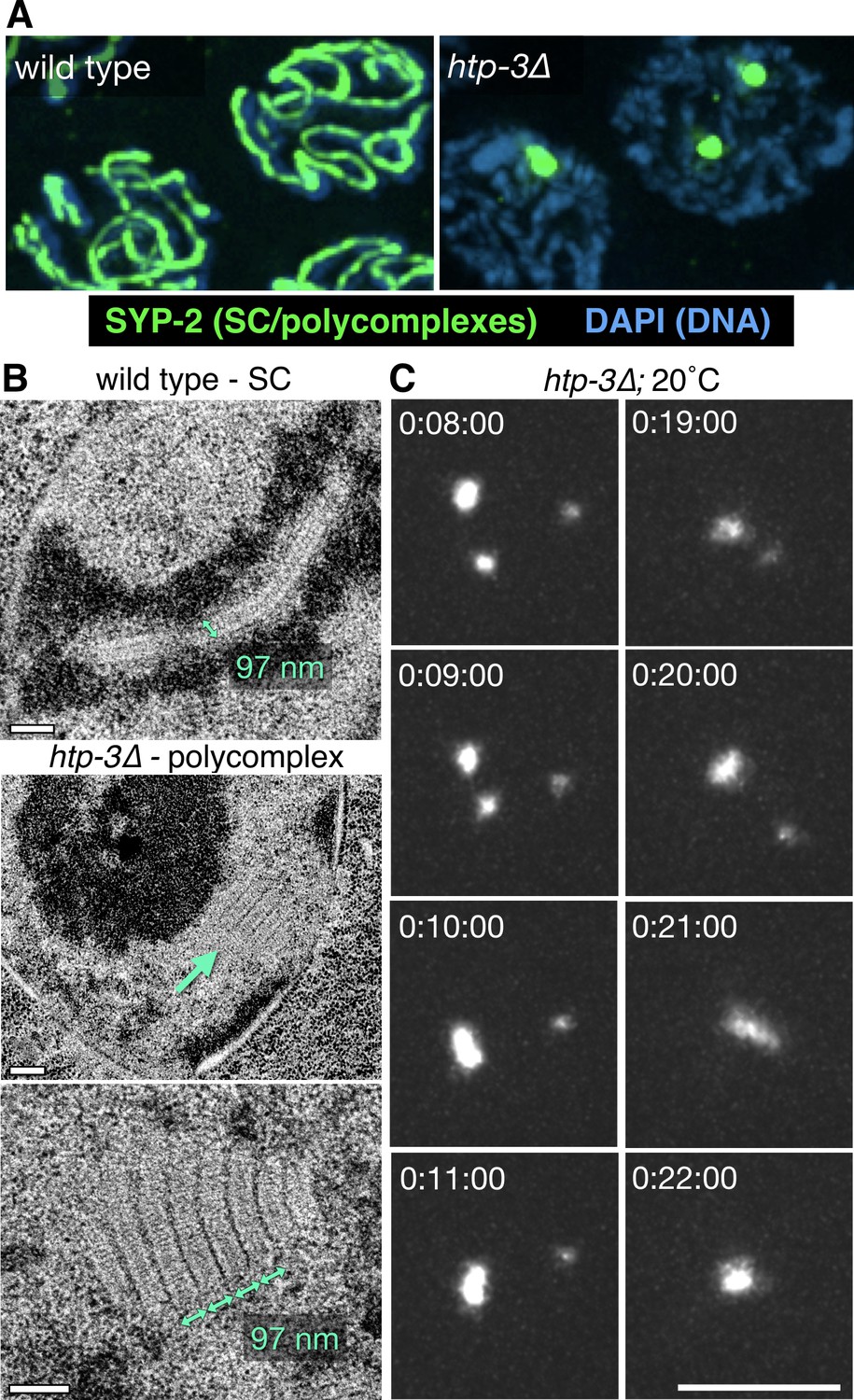
Polycomplexes are 3D lattices of SC proteins that exhibit liquid-like behaviors.
(A) Fluorescence micrograph showing mid-prophase oocyte nuclei from wild type and htp-3(tm3655) hermaphrodites, immunostained for SYP-2. Whereas in wild type animals SCs load between homologous chromosomes and appear as long filaments, in the absence of HTP-3 SC proteins form one or more large bodies that contain all of the known SC central region proteins. See Figure 1—figure supplement 1A for images of the entire gonads. (B) Representative transmission electron micrographs. The top image shows a single nucleus from a wild type hermaphrodites, with a synapsed chromosome pair (darkly-staining material flanking the SC is chromatin). The middle image shows a single nucleus from a htp-3(tm3655) hermaphrodite, with a polycomplex indicated by the green arrow. The darkly staining region in the center of the nucleus is the nucleolus. The bottom image shows a higher-magnification view of a polycomplex from a different nucleus. The distance between parallel darkly-staining bands is 97 nm, identical to the width of SCs that normally form between homologous chromosomes. Notably, these polycomplexes do not contain any of the known chromosome axis components, including cohesins and the HORMA domain proteins HTP-3, HIM-3, HTP-1 and HTP-2 (Severson et al., 2009 and our observations). This implies that the electron-dark lateral bands of the SC do not correspond to the chromosome axis, as has been long presumed, but are instead part of the structure formed by the central region proteins. See Figure 1—figure supplement 2A for serial sections of the polycomplex in the lower panel. Scale bars = 0.2 μm. (C) Projection images showing a single nucleus from a live recording of a htp-3(tm3655); GFP-SYP-3 hermaphrodite, at selected time points. Elapsed times are indicated as hours:minutes:seconds. Polycomplexes continually undergo deformations and fusions. Here, fusion of two initially separate polycomplexes is observed between 0:09:00 and 0:10:00, and again between 0:20:00 and 0:21:00. The full recording is shown in Video 1. Scale bar = 5 μm.
Figure 1—figure supplement 1
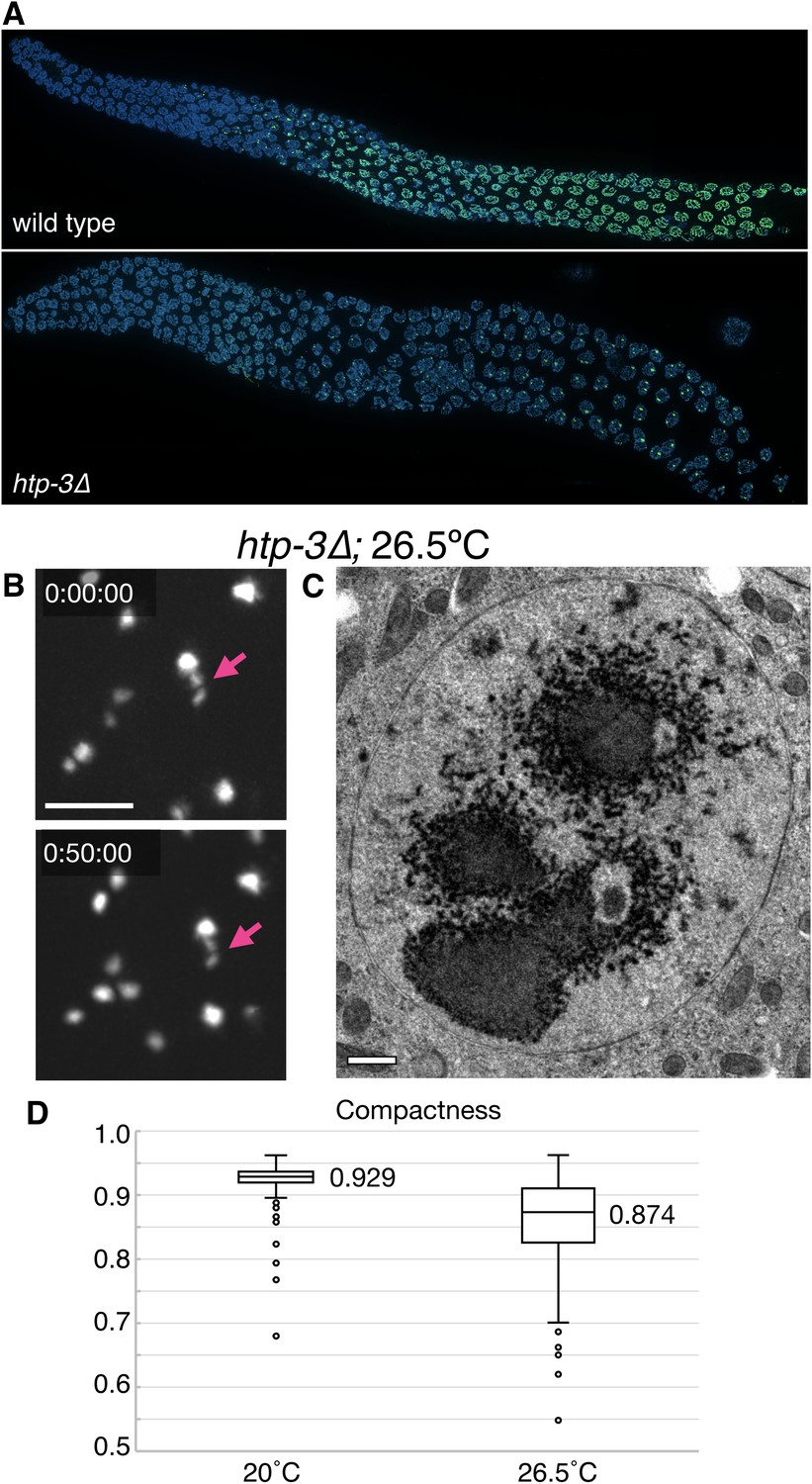
Further characterization of polycomplexes and heat-induced SC aggregates.
(A) Fluorescence micrograph showing gonads from wild type and htp-3(tm3655) hermaphrodites, immunostained for SYP-2. Whereas in wild type animals SCs load between homologous chromosomes and appear as long filaments, in the absence of HTP-3 one or more large bodies that contain all of the known SC central region proteins are observed in each nucleus. See Figure 2B for an explanatory diagram of the C. elegans gonad and Figure 1A for high magnification images of pachytene nuclei. (B–D) Heat-induced SC aggregates are distinct from polycomplexes. (B) Projection images from a real-time recording from a live htp-3; GFP-SYP-3 hermaphrodite that was incubated for 24 hr at 26.5°C prior to imaging. Several nuclei are seen in this field. Following extended exposure to heat, the SC protein aggregates undergo limited motion within the nucleus, and maintain their overall shapes throughout the time course (50 min). They were not observed to undergo fusions when in close proximity. The full recording is shown in Video 5. Scale bar = 5 μm. (C) Transmission electron micrograph showing a single mid-prophase meiotic nucleus from an htp-3(tm3655) mutant hermaphrodite incubated overnight at 26.5°C prior to high-pressure freezing. No striated polycomplexes were detected in these samples. The nucleoli appeared to be very electron-dark and fragmented, as also observed by fluorescence microscopy (Figure 4—figure supplement 4B). Scale bar = 0.5 μm. (D) Polycomplexes are more compact than heat-induced aggregates. Compactness was calculated by dividing the surface area of a sphere with the same volume as the given particle to the surface area of the particle, based on images of polycomplexes and aggregates from live htp-3; GFP-SYP-3 hermaphrodites grown at 20°C or 26.5°C for 24 hr. Compactness is overestimated in both cases due to the limited resolution of the microscope system; values are thus primarily useful for comparison. The boxes indicate the median and the interquartile range, and the whiskers 1.5 times the interquartile range. n = 234 and 289 nuclei for 20°C and 26.5°C, respectively. Student’s t-test: p<10−37.
Figure 1—figure supplement 2
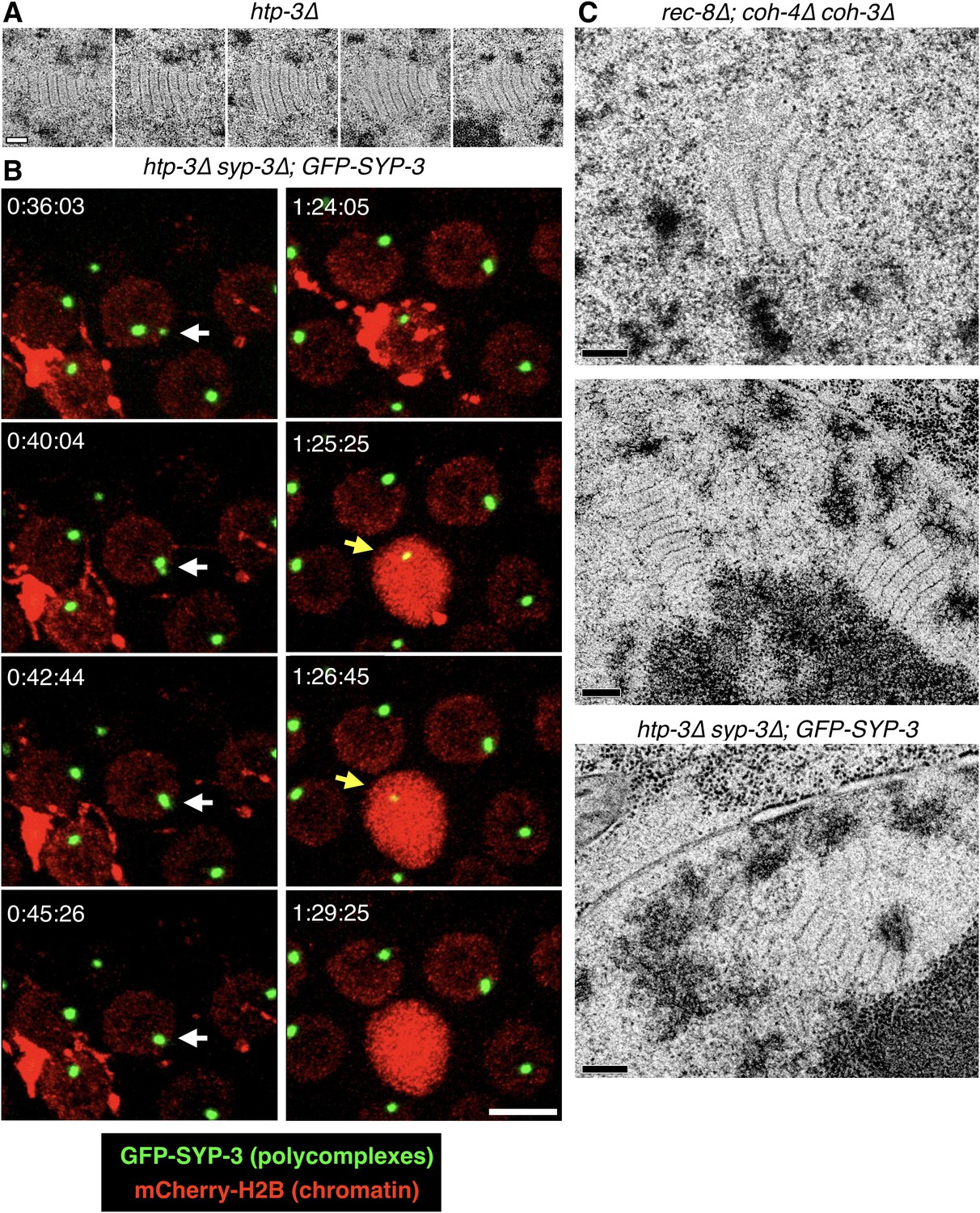
Further characterization of polycomplexes.
(A) Images from serial 70 nm sections through the polycomplex shown in Figure 1B (bottom panel). Scale bars = 0.2 μm. (B) Still images from a representative recording from an intact, live htp-3(tm3655) mutant hermaphrodite expressing GFP-SYP-3 (SC/polycomplexes; green) and mCherry-H2B (chromatin; red). When polycomplexes come into contact within the same nucleus, they frequently merge with one another (white arrows at t = 36:04–45:26). An example of an oocyte undergoing apoptosis is also observed in this recording. The mCherry-H2B reporter (or proteolytic fragments) localizes to the membranes of the engulfing somatic cell. A single large polycomplex persists for a few minutes following engulfment of the apoptotic corpse (yellow arrows at t = 1:25:25–1:26:45), and then disappears. The full recording is shown in Video 2. Scale bar = 5 μm. (C) Polycomplexes are detected in triple kleisin mutants and in htp-3 mutants expressing only fluorescently tagged SYP-3. Top and middle: transmission electron micrographs showing mid-prophase meiotic nuclei from rec-8(ok978); coh-4(tm1857) coh-3(gk112) hermaphrodites, which lack meiotic kleisins. Middle, two polycomplexes within the same nucleus, both adjacent to the darkly-staining nucleolus (bottom). The darkly staining material around the polycomplexes is chromatin. Bottom, an oocyte nucleus from an htp-3(tm3655) syp-3(ok758); GFP-SYP-3 hermaphrodite. The darkly staining nucleolus is seen at the bottom right corner of the frame. Scale bars = 0.2 μm.
Figure 2 with 1 supplement
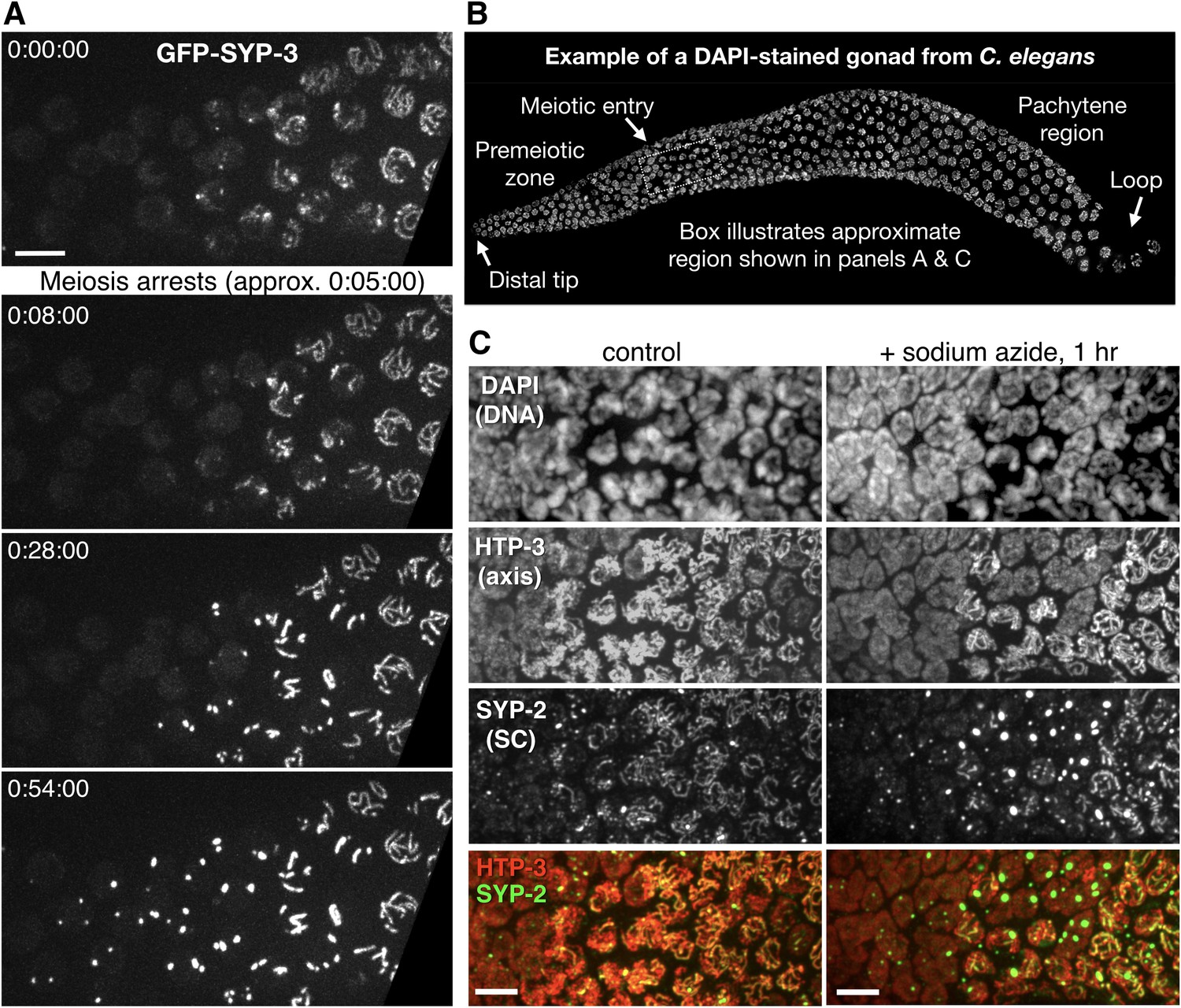
Condensation of SC proteins following meiotic arrest or ATP depletion.
(A) Still images from a time-lapse recording of an otherwise wild-type hermaphrodite expressing GFP-SYP-3, sampled at 1 min intervals. A portion of the germline containing early meiotic nuclei, corresponding approximately to the boxed region in panel (B), is shown. During this recording, chromosome movements abated at approximately 0:05:00. Following this arrest, gradual condensation of fluorescent bodies is observed in nuclei lacking preexisting stretches of SC (left side of frame), while existing SC segments become brighter, but do not elongate (near the right side of frame). Scale bar = 5 μm. The full recording is shown in Video 3. (B) The proximal region of a gonad from a C. elegans hermaphrodite, fixed and stained with DAPI, is shown as a reference for other figures and recordings. Each gonad contains a complete progression of meiotic stages; here, nuclei enter and advance through meiotic prophase from left to right. A signal from the distal tip inhibits meiotic entry; once cells move far enough from this signal they enter meiosis and progress through prophase. At the loop region of the gonad, a spatially regulated signal triggers exit from pachytene. (C) Projection images from deconvolved 3D image stacks, showing premeiotic and early meiotic nuclei from wild-type adult hermaphrodites that were incubated for 1 hr on plates containing 0.5% w/v sodium azide before fixation, and control (non-azide-treated) animals. Gonads were dissected and fixed immediately following azide treatment, and stained with antibodies against SYP-2 and HTP-3 to localize SCs and chromosome axes, respectively. Small polycomplexes that contain SYP-2 but not HTP-3 are observed in some early meiotic nuclei in the absence of any treatment, as previously described (Goldstein, 2013), but become much larger and more abundant in response to azide exposure. Note that transition zone nuclei maintain their polarized chromatin morphology during arrest. Scale bars = 5 μm.
Figure 2—figure supplement 1
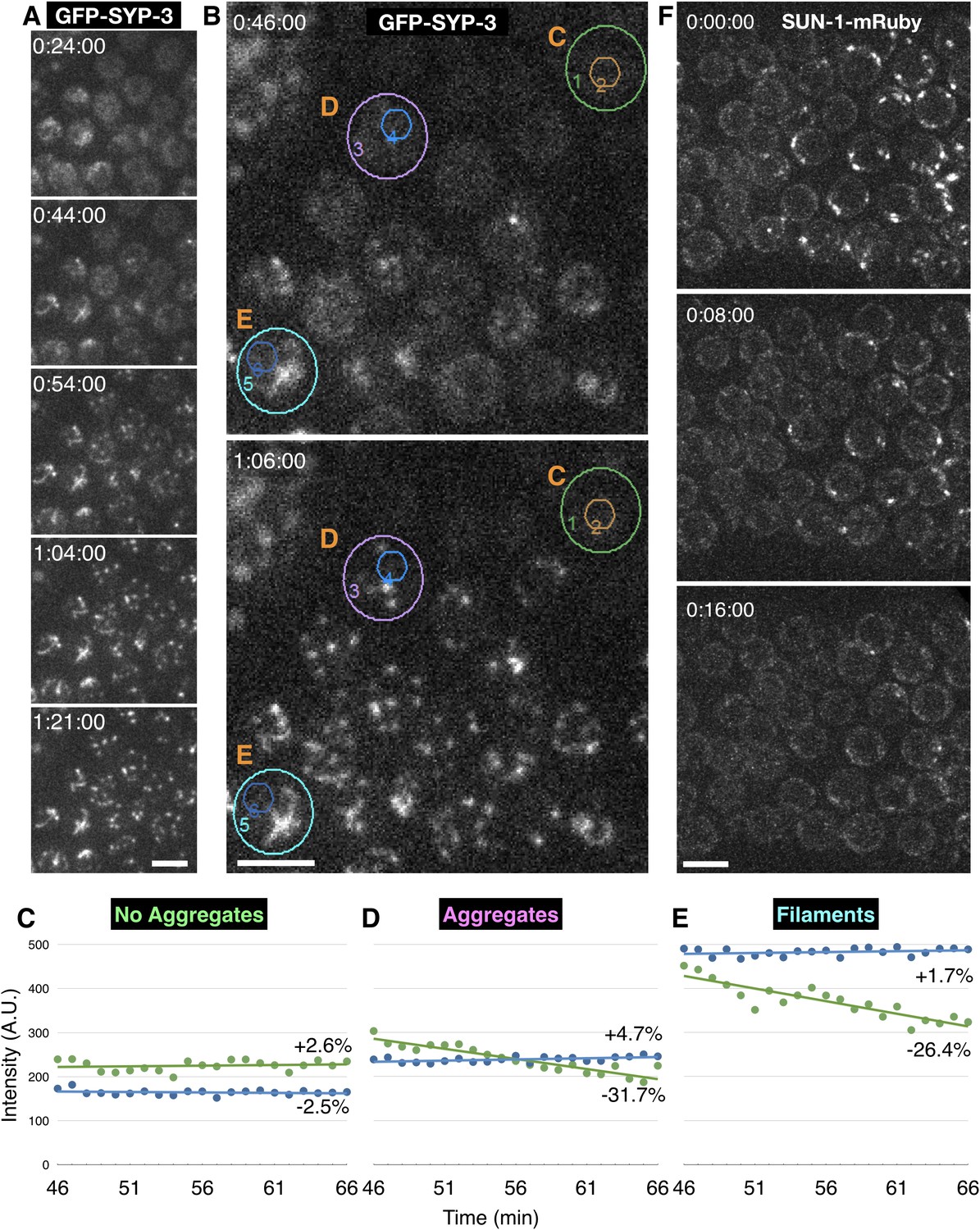
Condensation of polycomplexes from the nucleoplasm following meiotic arrest.
(A–E) Time-lapse recording showing the coalescence of SC proteins to existing SCs and polycomplexes following cessation of meiotic chromosome movements. Images were acquired from hermaphrodites expressing GFP-SYP-3 at 1 min intervals. In these images the direction of meiotic progression is from top right to bottom left. Meiotic chromosome movements cease at approximately 0:20:00. The full recording is shown in Video 4. Scale bars = 5 μm. (A) Additive Z projections at the indicated times. (B–E) Coalescence of SC proteins results in loss of diffuse fluorescence from the nucleoplasm. Additive Z projections from the indicated times are shown. Autofluorescence of lipid droplets in the intestine is seen at the upper left corner. (C–E) show plots of integrated fluorescence intensity, measured within the regions marked by concentric circles in (B) as a function of time. The larger circles encompass whole nuclei, while the smaller circles mark nucleoplasmic regions that lack discrete aggregates or filaments. All intensity values were background-subtracted based on an area of the same size outside the gonad. The difference between the final and initial value, based on linear regression, is indicated. (C) Nucleoplasmic fluorescence remains stable over time in nuclei where no aggregation is seen (green circle). (D) In nuclei with new aggregates there is a small increase in overall intensity, but a marked decrease in the intensity of nucleoplasmic regions without aggregates, indicating depletion of nucleoplasmic SC proteins (lilac circle). (E) In nuclei that contain SC filaments at the time when chromosome motions cease, filaments become progressively brighter (cyan circle). As in (D), the overall fluorescence increases marginally, while regions without filaments exhibit marked decrease in fluorescence, indicating depletion of SC proteins from the nucleoplasm. (F) Aggregates of SUN-1 at the nuclear periphery disperse following meiotic prophase arrest, by contrast to the behavior of SC proteins, which aggregate in response to arrest. Adult hermaphrodites expressing SUN-1-mRuby were immobilized in 1% w/v sodium azide to deplete ATP. Meiosis in these images progresses from left to right. Additive Z projections of premeiotic transition zone nuclei at the indicated times are shown. Scale bar = 5 μm.
Figure 3 with 1 supplement
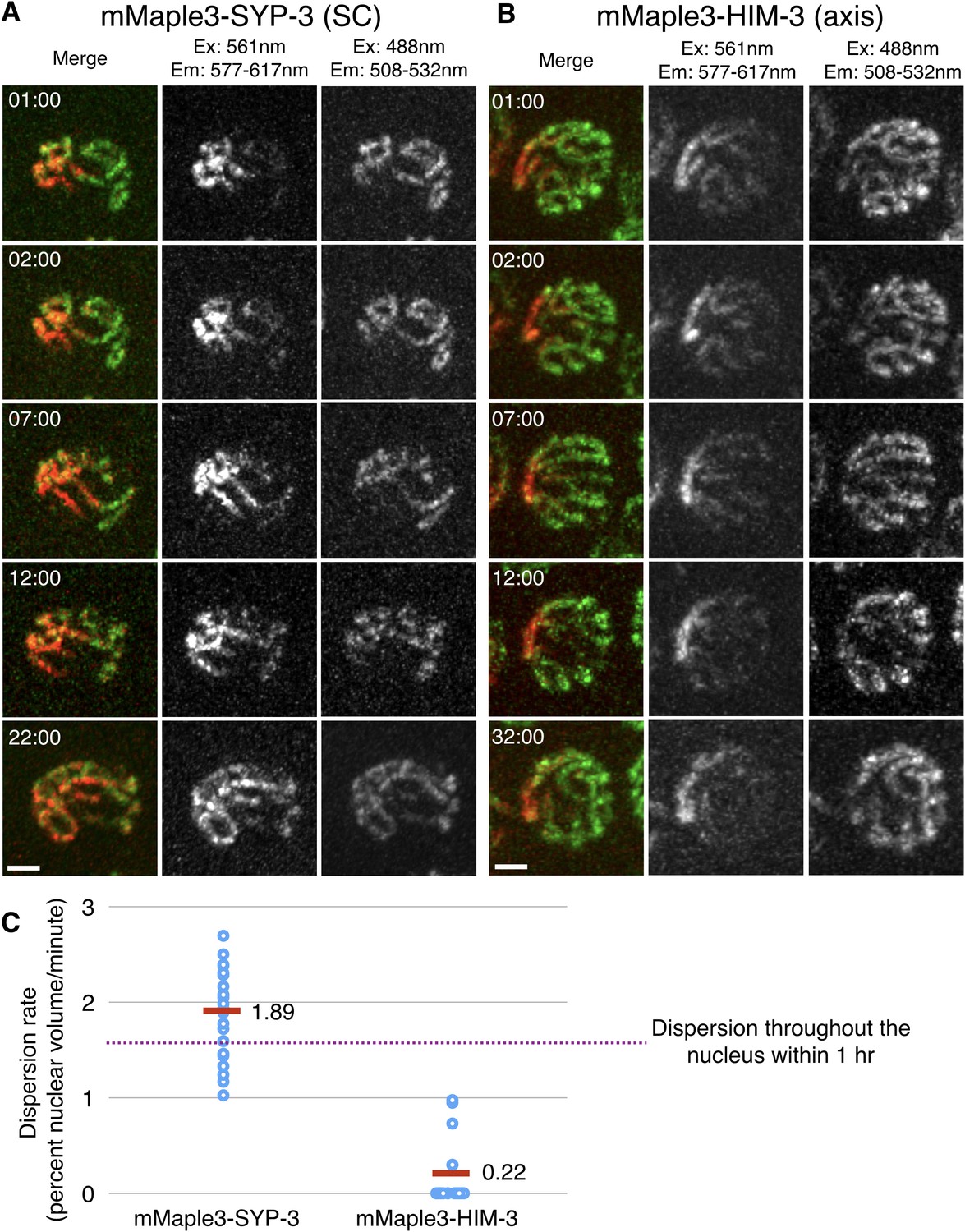
SC proteins, but not axis components, are highly dynamic.
(A–B) Images of representative nuclei from time-lapse recordings of hermaphrodites expressing either the SC protein SYP-3 or the axis component HIM-3 fused to the photoconvertible fluorescent protein mMaple3. A subnuclear volume was photoconverted using 405 nm laser illumination at t = 00:00. The mMaple3-SYP-3 signal spread throughout the nucleus and to all chromosomes by 22 min (A), whereas the mMaple3-HIM-3 signal remained confined to a small region throughout the time course (32 min; B). Scale bars = 2 μm. Elapsed times are indicated as min:sec. (C) The mobility of HIM-3 (axis) and SYP-3 (SC) were quantified by estimating the volume of individual nuclei containing photoconverted (red) signal as a function of time (see Materials and methods for details); SYP-3 within assembled SCs is far more mobile than HIM-3 (Student’s t-test: p<10−13). The horizontal dashed line represents the expansion rate for a point source that becomes homogeneously distributed throughout the nuclear volume in one hour.
Figure 3—figure supplement 1
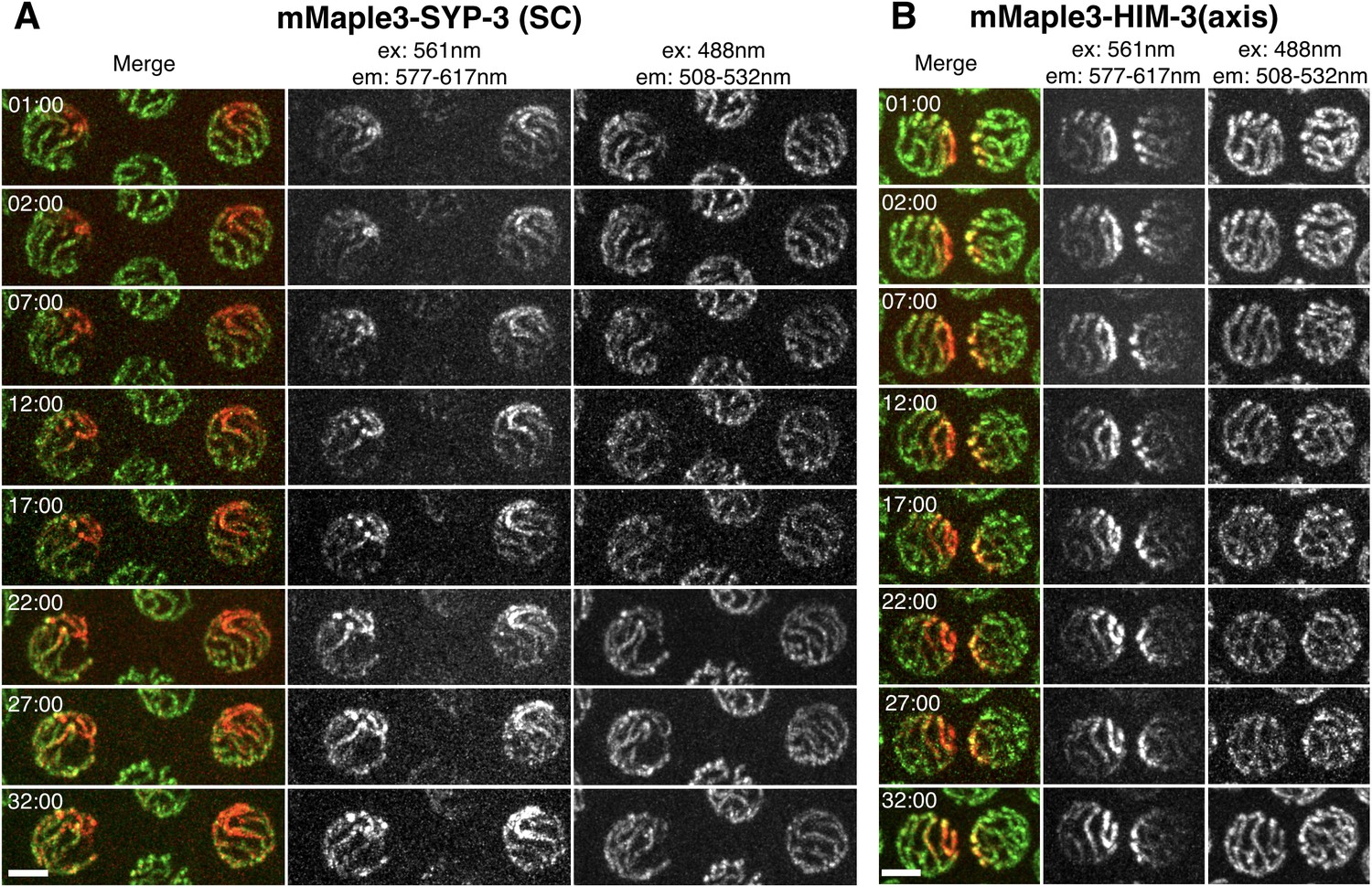
SC proteins, but not axis components, are highly dynamic.
(A) Time-lapse images of representative nuclei from adult hermaphrodites expressing mMaple3-SYP-3 (A) or mMaple3-HIM-3 (B). The fluorescent protein was photoconverted within a small region of individual nuclei at t = 00:00 using focused 405 nm light. Scale bars = 3 μm. (A) Photoconverted SYP-3 spreads throughout the nucleus and to all chromosome during the time course (32 min). (B) Photoconverted HIM-3 remains confined to a region of similar volume throughout the time course (32 min).
Figure 4 with 4 supplements
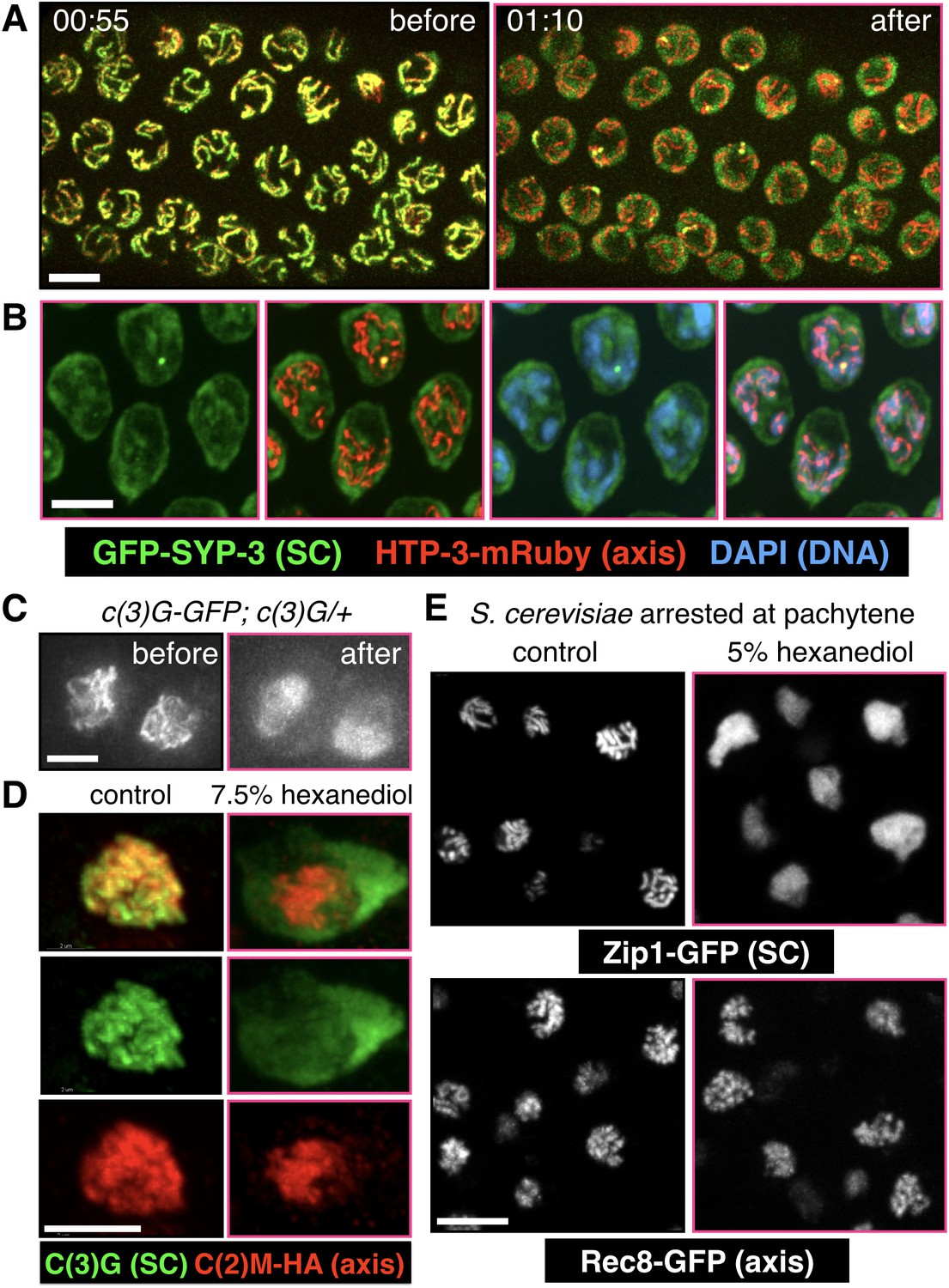
SCs in C.elegans, S. cerevisiae and Drosophila melanogaster dissolve in the presence of 1,6-hexanediol.
(A) Projection images showing selected time points from time-lapse recordings of extruded gonads from C. elegans hermaphrodites expressing GFP-SYP-3 and HTP-3-mRuby to mark SCs and axes, respectively. Upon exposure to 5% 1,6-hexanediol, SYP-3 immediately dispersed throughout the nucleoplasm, while HTP-3 remained associated with chromosomes. Most of the GFP-SYP-3 fluorescence remained in the nucleus for the duration of our experiments. The full recording is shown in Video 6. Scale bar = 5 μm. (B) Gonads were treated with 1,6-hexanediol as in (A), but then fixed and counterstained with DAPI. Scale bar = 5 μm. (C–D) The SC in Drosophila is also sensitive to 1,6-hexanediol. (C) Projection images showing GFP fluorescence in two adjacent oocytes from a Drosophila female expressing GFP-C(3)G. C(3)G) is an essential component of the SC central region. Ovarioles were dissected in buffer, and imaged before and after exposure to 7.5% 1,6-hexanediol. Scale bar = 5 μm. (D) Projection images showing a single oocyte within a germarium from a fly expressing HA-tagged C(2)M (a component of the chromosome axis). Ovarioles were dissected, exposed to 7.5% hexanediol (or not), and fixed with formaldehyde, then stained with antibodies against C(3)G (green) and HA (red). C(3)G was solubilized by hexanediol, while C(2)M remained associated with chromosome axes. Similar results were obtained with flies expressing GFP-C(3)G and stained with antibodies against GFP and C(2)M (data not shown). Scale bar = 5 μm. (E) Diploid ndt80 S. cerevisiae strains expressing the indicated GFP fusion proteins were sporulated and arrested in mid-pachytene. Following exposure to 1,6-hexanediol, cells were imaged immediately without fixation. The central region component Zip1p dispersed throughout the nuclei upon 1,6-hexanediol treatment, while the cohesin subunit Rec8p remained along chromosomes. Scale bar = 5 μm.
Figure 4—figure supplement 1

SC proteins, but not axis components, are highly dynamic.
(A) Electrostatic and hydrophobic interactions contribute to SC stability. As in Figure 4E, diploid ndt80 S. cerevisiae cells expressing Zip1p-GFP were sporulated, arrested in mid-pachytene, and treated with various concentrations of 1,6-hexanediol and KCl in buffer. SCs were scored as ‘dissolved’ if no distinct structures remained. (B) Effect of various di-alcohols on SC stability. As in Figure 4E, diploid ndt80 S. cerevisiae cells expressing Zip1p-GFP were sporulated, arrested in mid-pachytene, and treated with various concentrations of di-alcohols in buffer. For each alcohol, the lowest concentration necessary to completely dissolve the SC is shown. (C) Explanatory diagram of the effects of SC dissolution by 1,6-hexanediol on meiocytes from S. cerevisae, D. melanogaster and C. elegans. Prior to treatment the SC (green) and axes (red) colocalize on chromosomes in the nucleus (nucler envelope, blue; plasma membrane, lilac). Upon treatment with 1,6-hexanediol (right), SC material becomes diffuse in the nucleus and the cytoplasms, while the chromosome axes remain intact.
Figure 4—figure supplement 2
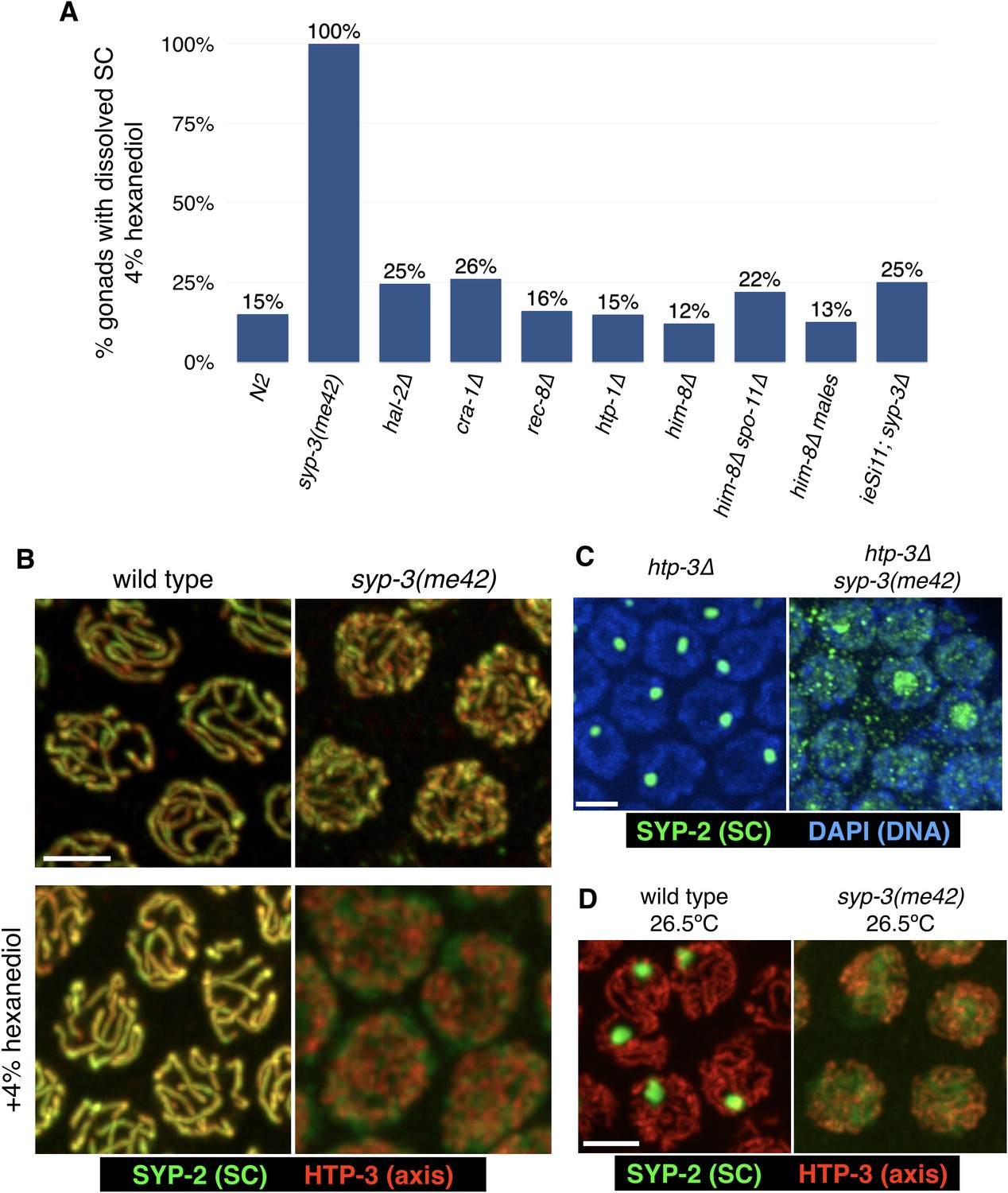
The syp-3(me42) mutation results in hypersensitivity of SCs to 1,6-hexanediol and prevents aggregation in the absence of axes or at high temperatures.
(A) The sensitivity of SCs to hexanediol was compared for various mutants that affect SC assembly and/or structure. Extruded gonads were exposed to 4% 1,6-hexanediol, which disrupts only a minor fraction (15%) of gonads in wild-type animals. In syp-3(me42) mutants, in which the C-terminus of one SC component is truncated by 11 amino acids, resulting in aberrant synapsis (Smolikov et al., 2007), 4% hexanediol completely disrupted the SCs (see images in (B)). Other mutations that affect CO formation or SC structure did not markedly affect hexanediol sensitivity, including hal-2 mutants, which show defects in SC assembly similar to syp-3(me42) mutants (Zhang et al., 2012). (B) Nuclei from wild-type and syp-3(me42) animals were dissected, treated with 4% hexanediol where indicated, fixed with formaldehyde, and stained with antibodies against SYP-2 (SC; green) and HTP-3 (axes; red). Scale bar = 3 μm. (C–D) The syp-3(me42) mutation prevents polycomplex formation and heat-induced aggregation. (C) Gonads from htp-3 and htp-3 syp-3(me42) hermaphrodites were fixed and stained with DAPI (chromatin; blue) and with antibodies against the SC component SYP-2 (green). SC proteins in htp-3 syp-3(me42) animals remain diffuse in the nucleoplasm, or concentrate in the nucleoli of some nuclei. Scale bar = 3 μm. (D) Wild-type and syp-3(me42) animals were incubated for 24 hr at 26.5°C before fixation and stained with antibodies against SYP-2 (green) HTP-3 (red). Under these conditions, SC proteins form nuclear aggregates in wild-type animals, as reported (Bilgir et al., 2013), but remain diffuse in the nucleoplasm of syp-3(me42) animals. Scale bar = 3 μm.
Figure 4—figure supplement 3
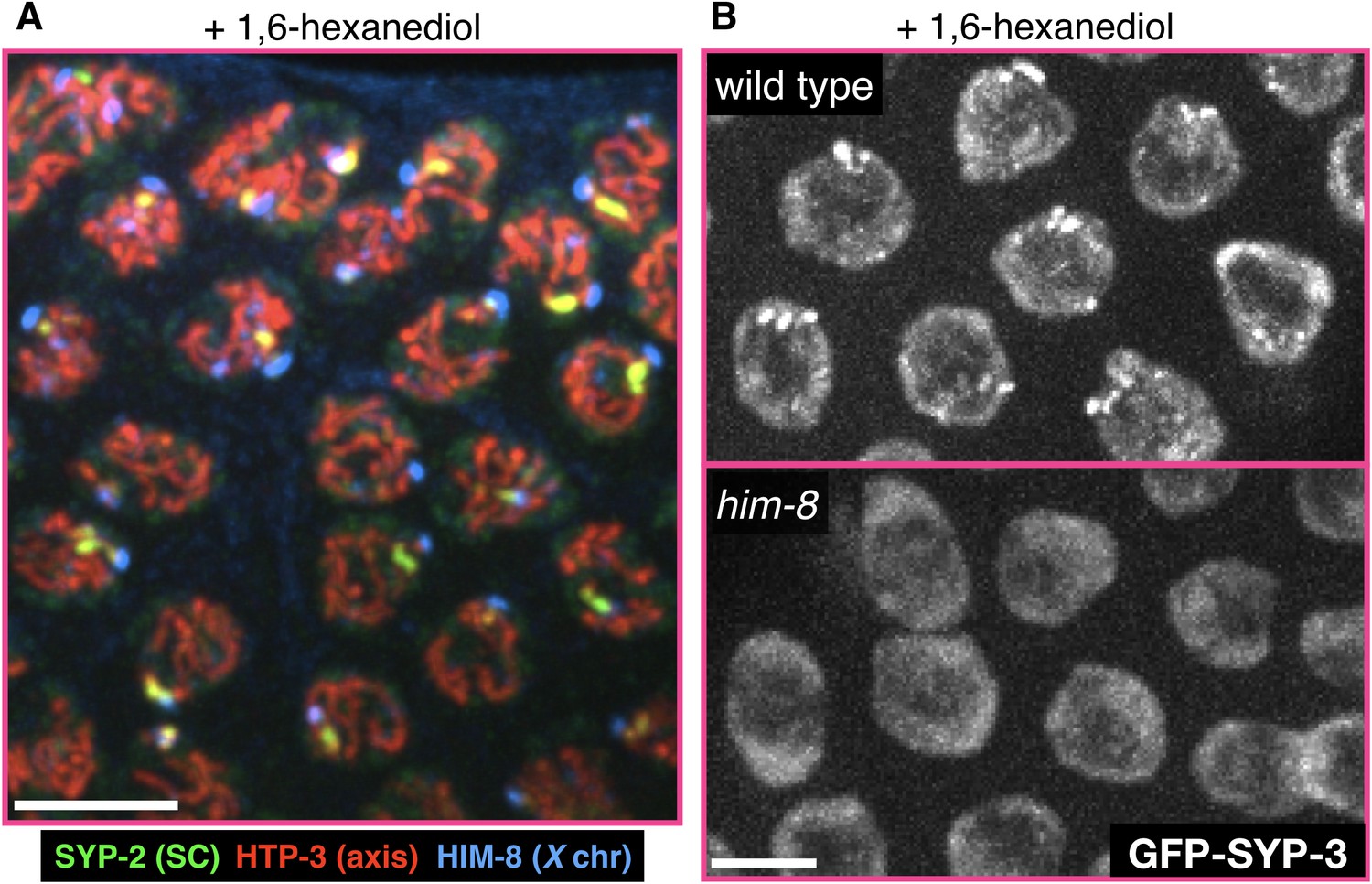
The X chromosome pairing center protein HIM-8 and a some of the SC on the X chromosome are refractory to 1,6-hexanediol.
(A) A representative gonad from a wild-type adult hermaphrodite treated with 7.5% 1,6-hexanediol prior to fixation, and stained with antibodies against the SC component SYP-2 (green), the axis component HTP-3 (red), and the X chromosome pairing center protein HIM-8 (blue). While most of the SC is dissolved, the chromosomal axes and the X chromosome pairing center remain intact. The small hexanediol-resistant stretch of SC is adjacent the chromosome X pairing center. Scale bar = 5 μm. (B) Images of meiotic nuclei from adult hermaphrodites expressing GFP-SYP-3 and treated with 7.5% 1,6-hexanediol prior to fixation. The small hexanediol-resistant SC stretches and puncta are not present in him-8 mutant worms (bottom), in which the X chromosomes do not pair or synapse. Scale bar = 5 μm.
Figure 4—figure supplement 4
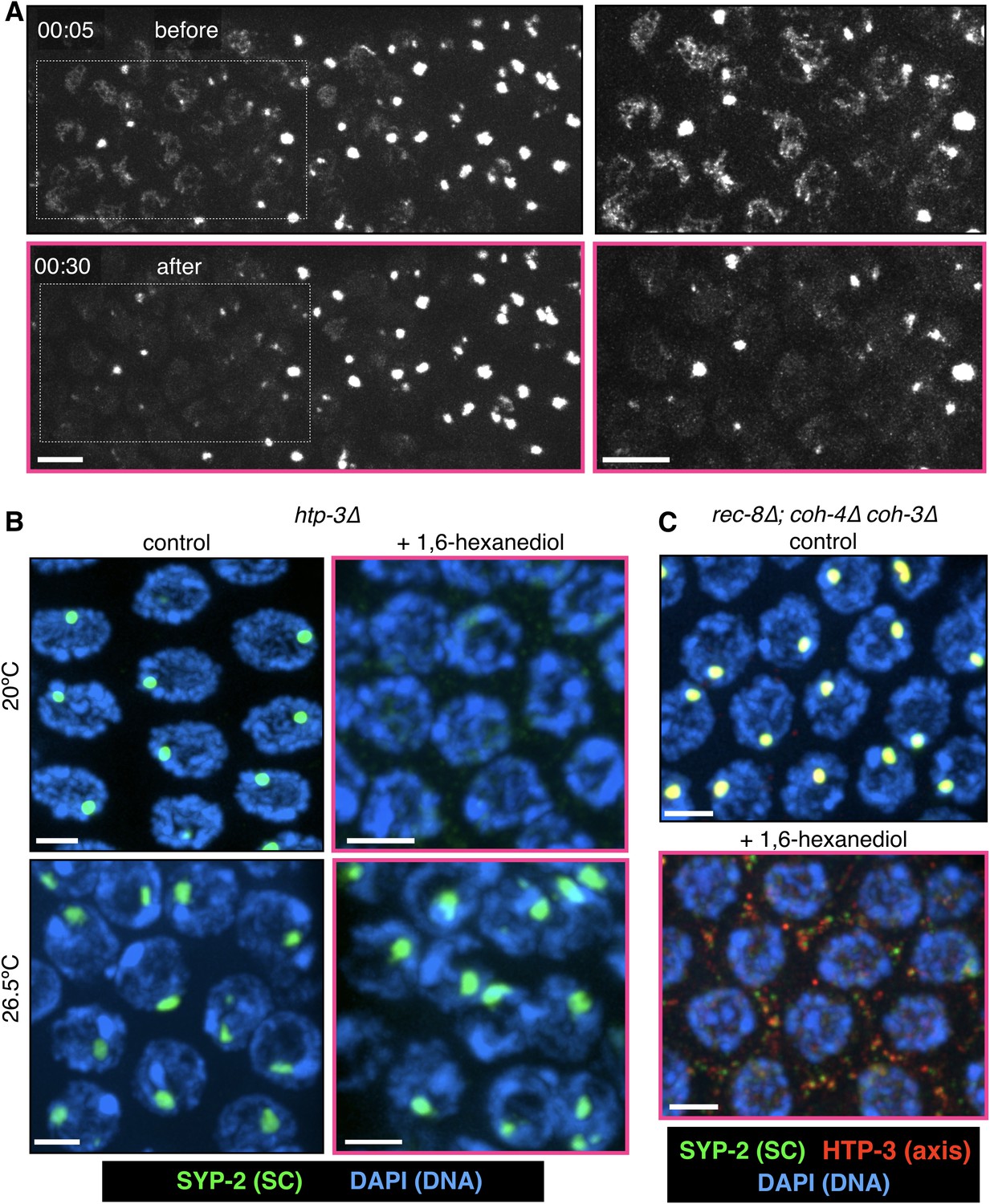
Polycomplexes, but not heat-induced aggregates, are dissolved by 1,6-hexanediol.
(A) Time-lapse images of gonads from GFP-SYP-3 hermaphrodites grown for 24 hr at 26.5°C and then treated with 7.5% 1,6-hexanediol. The large SC aggregates were resistant to hexanediol, while the few assembled SCs present in some nuclei were readily dissolved. The outlined regions are shown at larger magnification on the right; rescaled to highlight SC filaments, which are dimmer than the aggregates. The full recording is shown in Video 8. Scale bars = 5 μm. (B–C) Projections of 3D fluorescence optical sections. (B) Extruded gonads from htp-3(tm3655) hermaphrodites were incubated with 7.5% 1,6-hexanediol where indicated, then fixed and stained with antibodies against SYP-2 (green) and with DAPI (blue). Polycomplexes in samples grown at 20°C are sensitive to 1,6-hexanediol. SC aggregates following prolonged incubation at 26.5°C are resistant to 1,6-hexanediol. Scale bars = 3 μm. (C) Extruded gonads from rec-8(ok978); coh-4(tm1857) coh-3(gk112) hermaphrodites were incubated with 7.5% 1,6-hexanediol where indicated, then fixed and stained with antibodies against the SC component SYP-2 (green) and the axis component HTP-3 (red), and counterstained DAPI (blue). Polycomplexes in this triple kleisin mutant contain the axis protein HTP-3, which disperses (along with the SC components) upon 1,6-hexanediol exposure. Scale bars = 3 μm.
Figure 5
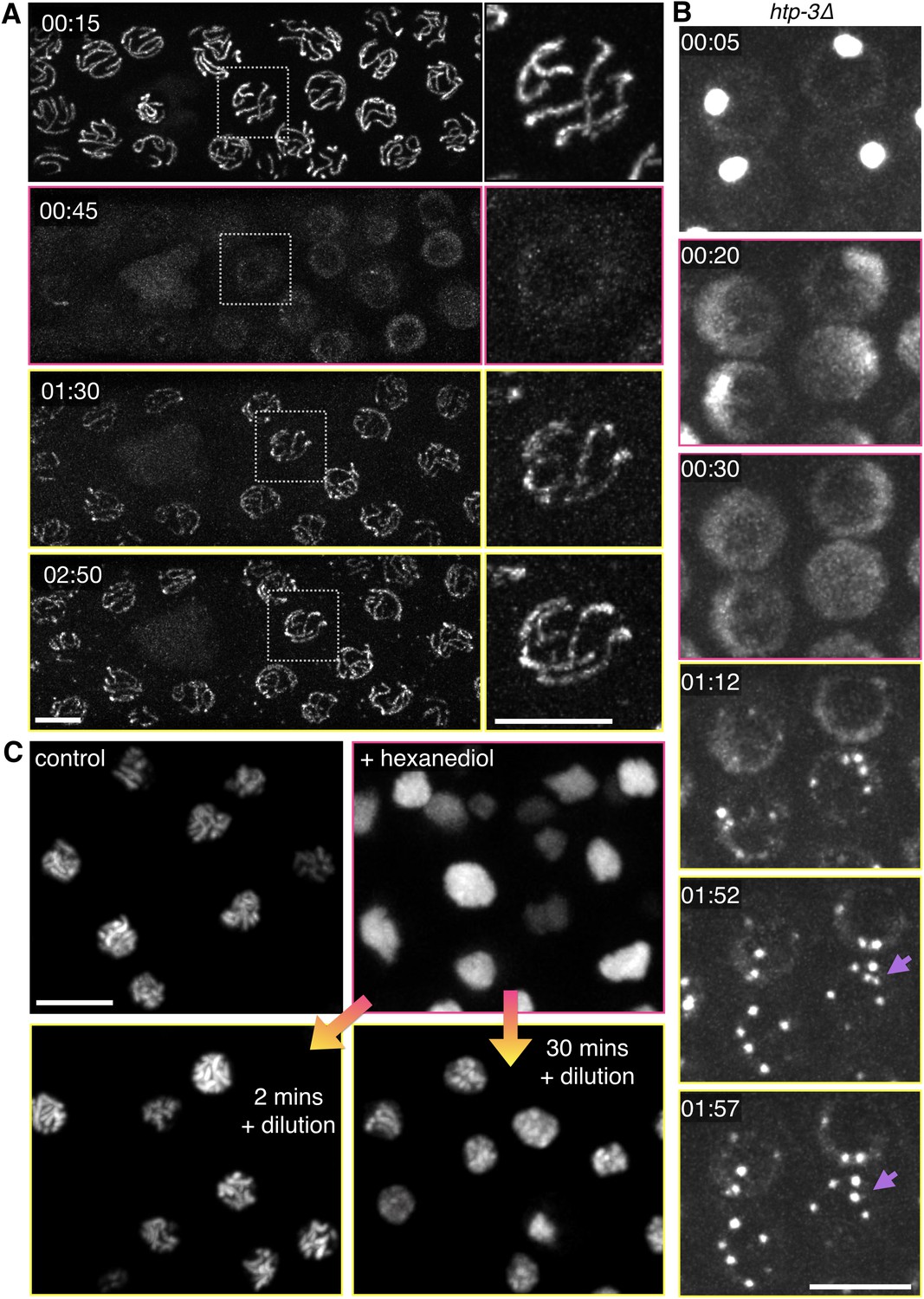
Dissolution of SCs and polycomplexes by 1,6-hexanediol is reversible.
(A) Projection images from a time-lapse recording showing GFP-SYP-3 before and after dispersion by 5% 1,6-hexanediol, followed by dilution with buffer. Pink borders around images indicate times after hexanediol addition, and yellow borders indicate times after dilution of the hexanediol with buffer. Upon dilution, fluorescence reappears along chromosomes. Some small puncta are also observed in the cytoplasm. Chromosome association of SC proteins after longer exposures to 1,6-hexanediol (>2 min) was not reversible (data not shown), perhaps due to irreversible perturbations of chromosome structure or SC subunits. The full recording is shown in Video 9. Elapsed time is indicated as mm:ss. Larger-magnification images of a representative nucleus are shown on the right. Scale bars = 5 μm. (B) Projection images from a time-lapse recording of a htp-3(tm3655) mutant hermaphrodite expressing GFP-SYP-3. Exposure to 7.5% 1,6-hexanediol (pink image borders) induces rapid dispersion of the SC proteins from polycomplexes. Upon dilution of 1,6-hexanediol with buffer (yellow image borders), SYP-3 coalesces into smaller bodies that fuse with each other upon contact (lilac arrows). The full recording is shown in Video 10. Scale bar = 5 μm. (C) A diploid ndt80 S. cerevisiae strain expressing Zip1p-GFP was sporulated, arrested in mid-pachytene, and treated with 5% 1,6-hexanediol. 10 volumes of buffer were added after the indicated incubation times with hexanediol, and cells were then mounted and imaged without fixation. Reassociation of Zip1 with chromosomes was observed when 1,6-hexanediol was diluted after 2 min or 30 min, but appeared to be less robust after the longer incubation. Scale bar = 5 μm.
Figure 6 with 1 supplement
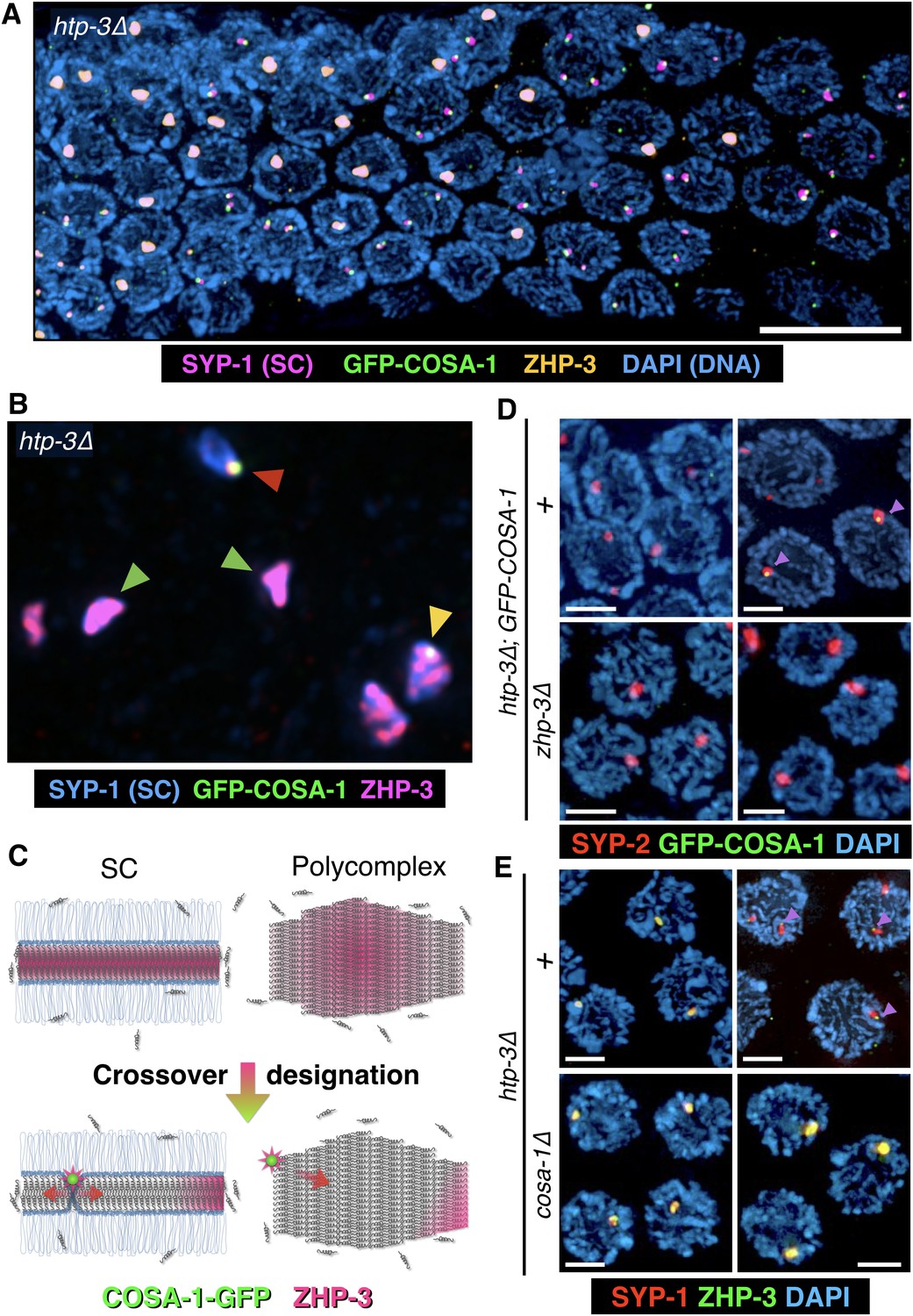
The localization of CO factors to polycomplexes recapitulates their dynamic behavior and interdependencies upon CO designation.
(A–B) Dynamic relocalization of ZHP-3 and COSA-1 during meiotic progression. Fluorescence micrograph of a representative gonad from htp-3(tm3655) hermaphrodite expressing GFP-COSA-1 stained with antibodies against SYP-1 (SC/polycomplexes; magenta), ZHP-3 (orange), and GFP (COSA-1; green), and counterstained with DAPI (blue). ZHP-3 is detected throughout the volume of polycomplexes in the mid-pachytene region of the gonad, while GFP-COSA-1 is not detected. In nuclei at the ‘loop’ region of the gonad, where nuclei normally exit pachytene (see Figure 2B for reference), ZHP-3 staining becomes confined to small foci abutting polycomplexes, where it colocalizes with GFP-COSA-1. An intermediate stage in which COSA-1 and ZHP-3 colocalize at the surface but ZHP-3 also remains throughout the polycomplex is also seen in some nuclei. Scale bar = 10 μm. (B) A higher magnification image of a similarly stained gonad, except SYP-1 is shown in blue and ZHP-3 is shown in magenta (DAPI is not shown). Early, late and intermediate stage polycomplexes are marked with green, red and yellow arrowheads, respectively. (C) A model of ZHP-3 and COSA-1 dynamic localization to SCs and polycomplexes. See main text for details. (D) Early (left) and late (right) stage pachytene nuclei from adult htp-3 or htp-3 zhp-3 hermaphrodites expressing GFP-COSA-1. Gonads were dissected and stained with antibodies against SYP-2 (SC/polycomplexes; red) and GFP (COSA-1; green) and counterstained with DAPI (blue). GFP-COSA-1 does not form foci associated with polycomplexes in the absence of ZHP-3. Scale bars = 2 μm. (E) Early (left) and late (right) stage pachytene nuclei from adult htp-3 or htp-3; cosa-1 hermaphrodites. Gonads were dissected and stained with antibodies against SYP-1 (SC/polycomplexes; red) and ZHP-3 (green), and counterstained with DAPI (blue). In the absence of COSA-1, ZHP-3 remains localized throughout polycomplexes even at late stage of meiotic prophase. Scale bars = 2 μm. The entire gonads for (D) and (E) are shown in Figure 6—figure supplement 1C–D.
Figure 6—figure supplement 1

Further characterization of the localization and 1,6-hexanediol sensitivity of ZHP-3 and COSA-1.
(A) Projection images showing meiotic prophase progression in whole gonads from adult hermaphrodites stained with antibodies against SYP-1 (SC; red) and the RING finger protein ZHP-3 (green). The images in the lower panels (pink borders) are from a gonad that was exposed to 7.5% hexanediol just prior to fixation. As previously described (Bhalla et al., 2008), ZHP-3 localizes throughout SCs in early pachytene, and then concentrates at designated CO sites. When the SC is dissolved by hexanediol, the diffusely localized ZHP-3 is also solubilized, but after localization to COs the protein becomes resistant to solubilization by hexanediol. The higher-magnification views on the right show individual nuclei from before and after the relocalization of ZHP-3 upon CO designation (top and bottom, respectively). Scale bars correspond to 10 μm (left) and 2 µm (right). (B) COSA-1 localizes to CO sites and is not dissolved by hexanediol. These projection images are from an adult hermaphrodite expressing GFP-COSA-1 and mRuby-SYP-3. Extruded gonads were exposed to 7.5% 1,6-hexanediol prior to fixation, and counterstained with DAPI (blue). GFP-COSA-1 (green) and mRuby-SYP-3 (red) were visualized by their intrinsic fluorescence. Although SC proteins were dispersed by hexanediol, the 6 CO sites are still marked by GFP-COSA-1. Bottom, magnified view of a single nucleus. Scale bars are 8 μm and 2 μm, for the top and bottom panels, respectively. (C–D) COSA-1 and ZHP-3 are interdependent for their dynamic relocalization during meiotic progression. The outlined regions are shown at higher magnification in Figure 6D–E. (C) Adult htp-3 or htp-3 zhp-3 hermaphrodites expressing GFP-COSA-1 were dissected and stained with antibodies against SYP-2 (SC/polycomplexes; red) and GFP (COSA-1; green) and counterstained with DAPI (blue). GFP-COSA-1 does not form foci associated with polycomplexes in the absence of ZHP-3. Scale bars = 10 μm. (D) Adult htp-3 or htp-3; cosa-1 hermaphrodites were dissected and stained with antibodies against SYP-1 (SC/polycomplexes; red) and ZHP-3 (green), and counterstained with DAPI (blue). In the absence of COSA-1, ZHP-3 remains localized throughout polycomplexes throughout meiotic prophase. Scale bars = 10 μm. (E) Dynamic relocalization of ZHP-3 and COSA-1 is accompanied by different hexanediol sensitivity. Gonads from htp-3 hermaphrodites expressing GFP-COSA-1 were dissected and stained with antibodies against SYP-1 (SC/polycomplexes; magenta), ZHP-3 (orange), and GFP (COSA-1; green), and counterstained with DAPI (blue). Top, higher-magnification view of polycomplexes showing examples of early and late localization patterns. Bottom, upon dissolution of polycomplexes with 1,6-hexanediol, small foci positive for ZHP-3 and GFP-COSA-1 remain visible. The solubilized SC proteins are largely excluded from the volume occupied by chromosomes, resulting in a somewhat filamentous appearance. Scale bars = 2 μm.
Videos
Video 1
Polycomplexes fuse upon contacting each other.
Oocytes from the diplotene region of an htp-3(tm3655) hermaphrodite expressing GFP-SYP-3, corresponding to the stills shown in Figure 1C. Polycomplexes exhibit constant deformations and merge with one another (between t = 0:09:00 and t = 0:10:00 and between t = 0:20:00 and t = 0:21:00). Meiotic progression in this recording is right to left. Images were acquired every 1 min. Playback is 300x real-time. Scale bar = 5 μm..
Video 2
Polycomplexes merge upon contact.
Pachytene nuclei from an intact htp-3(tm3655) hermaphrodite expressing GFP-SYP-3 and histone-mCherry, corresponding to the stills in Figure 1—figure supplement 2B. Polycomplexes (green) merged with one another (between t = 0:40:04 and t = 0:42:44). An apoptotic nucleus was being engulfed between t = 1:24:05 and t = 1:25:25. A polycomplex remains visible until t = 1:26:45. Meiotic progression in this recording is from left to right. Images were acquired about every 1 min. Playback is 400x real-time. Scale bar = 4 μm.
Video 3
Aggregation of SC components following cessation of meiotic chromosome movements.
Meiotic prophase in a hermaphrodite expressing GFP-SYP-3. Premeiotic nuclei are at the left, and nuclei enter meiosis and progress from left to right. Chromosome motion ceases, indicative of meiotic arrest, at about t = 5 min. Images were acquired every 1 min. Playback speed is 300x real-time. Scale bar = 5 μm. Stills are shown in Figure 2A.
Video 4
Aggregation of SC components following cessation of meiotic chromosome movements.
Recording from a hermaphrodite expressing GFP-SYP-3. Premeiotic nuclei are at the lower right portion of the images, and meiotic entry/progression is observed from lower right to upper central portion of the images. Fluorescent lipid droplets within the intestine are seen to the left of the gonad. Images were acquired every 1 min. Playback is 300x real-time. Scale bar = 5 μm. Stills and analysis of this recording are shown in Figure 2—figure supplement 1.
Video 5
Heat-induced SC aggregates do not show liquid-like properties.
A htp-3(tm3655) mutant hermaphrodite expressing GFP-SYP-3 was incubated overnight at 26.5°C (corresponding to images in Figure 1—figure supplement 1B). SC aggregates show some mobility within the nucleus, but do not merge with each other, and maintain their irregular shapes over the time course. Meiotic progression in this recording is from left to right. Images were acquired every 1 min. Playback is 300x real-time. Scale bar = 5 μm.
Video 6
The central region of the SC, but not the chromosome axis, is dissolved by 1,6-hexanediol.
This gonad was extruded from a hermaphrodite expressing GFP-SYP-3 and HTP-3-mRuby. Partially synapsed zygotene nuclei are seen at the left, and fully synapsed pachytene nuclei at the right. 1,6-hexanediol was added to a concentration of 5% at t = 0:00:20. Images were acquired every 5 s. Playback is 50x real-time. Scale bar = 5 μm. Stills are shown in Figure 4A..
Video 7
1,6-hexanediol dissolves the SC central region.
Recording from a gonad extruded from a hermaphrodite expressing GFP-SYP-3 and HTP-3-mRuby. 5% 1,6-hexanediol was added at t = 0:00:35 (yellow flash). Meiotic progression in this recording is from right to left. Images were acquired every 5 s. Playback is 25x real-time. Scale bar = 10 μm.
Video 8
Heat-induced SC aggregates are resistant to 1,6-hexanediol.
Time-lapse movie of a gonad from GFP-SYP-3 hermaphrodite incubated overnight at 26.5°C shown in Figure 4—figure supplement 4A. 7.5% 1,6-hexanediol was added at t = 0:00:10. SC stretches in the region of the germline at the left side of the image are quickly dissolved, while the large, irregular SC aggregates remain intact. Meiotic progression in this recording is from left to right. Images were acquired every 5 s. Playback speed is 25x real-time. Scale bar = 5 μm.
Video 9
1,6-hexanediol reversibly dissolves the SC.
Time-lapse movie of a gonad from GFP-SYP-3 hermaphrodite shown in Figure 5A. 7.5% 1,6-hexanediol was added at t = 0:00:20, and diluted at t = 0:00:50 and 0:01:35. Meiotic progression in this recording is from right to left. Images were acquired every 5 s. Playback is 25x real-time. Scale bar = 5 μm.
Video 10
1,6-hexanediol dissolves polycomplexes.
Extruded gonad from an htp-3(tm3655) mutant expressing GFP-SYP-3, corresponding to the stills shown in Figure 5B. 7.5% 1,6-hexanediol was added at t = 0:00:10, and diluted at t = 0:00:45 and t = 0:01:17. Two small polycomplexes, reformed after 1,6-hexanediol addition and dilution, merge to form a single body at t = 0:01:52. Images were acquired about every 5 s. Playback is 25x real-time. Scale bar = 4 μm.
Tables
Table 1
Strains used in this study.
| Strain Name | Genotype |
| CA257 | him-8(tm611) IV |
| CA277 | unc-24(e138) him-8(e1489) spo-11(ok79) / mIs11 IV |
| CA795 | htp-1(gk174) / nT1 [unc-?(n754) let-?] (IV;V) |
| CA821 | htp-3(tm3655) I / hT2 [bli-4(e937) let-?(q782) qIs48] (I;III) |
| CA826 | cra-1(tm2144) III / hT2 [bli-4(e937) let-?(q782) qIs48] (I;III) |
| CA861 | syp-3(ok758) I / hT2 [bli-4(e937) let-?(q782) qIs48] (I;III) |
| CA899 | syp-3(me42) I / hT2 [bli-4(e937) let-?(q782) qIs48] (I;III) |
| CA904 | rec-8(ok978) / nT1 IV; coh-4(tm1857) coh-3(gk112) / nT1 [qIs51] V |
| CA1010 | meIs8 [pie-1p::GFP::cosa-1 + unc-119(+)] II |
| CA1095 | rec-8(ok978) IV / nT1[qIs51] (IV;V) |
| CA1122 | cosa-1(me13) / qC1 [dpy-19(e1259) glp-1(q339) qIs26] III |
| CA1234 | syp-3(ok758) I; ieSi63 [cbunc-119+, psyp-3::mMaple3::syp-3] II; unc-119(ed3) III. |
| CA1237 | htp-3(tm3655) I; cosa-1(ie98) III |
| CA1238 | htp-3(tm3655) I/hT2 [bli-4(e937) let-?(q782) qIs48] (I,III); meIs8 [pie-1p::GFP::cosa-1 + unc-119(+)] II |
| CA1239 | htp-3(tm3655) zhp-3(ie97) I; meIs8 [pie-1p::GFP::cosa-1 + unc-119(+)] II |
| CA1253 | syp-3 (ok758) I; ieSi11 [cbunc-119+, Psyp-3::EmeraldGFP::syp-3] II; unc-119(ed3) III |
| CA1255 | htp-3(tm3655) syp-3(ok758) I; ieSi11 [cbunc-119+, Psyp-3::EmeraldGFP::syp-3] II; ieSi17 [cbunc-119, Phtp-3::htp-3::mRuby] IV |
| CA1255 | htp-3(tm3655) syp-3(ok758) I; ieSi11 [cbunc-119+, Psyp-3::EmeraldGFP::syp-3] II; ieSi17 [cbunc-119, Phtp-3::htp-3::mRuby] IV |
| CA1297 | syp-3(ok857) I; ieSi11 [cbunc-119+, Psyp-3::EmeraldGFP::syp-3] II; him-8(tm611) IV |
| CA1298 | meIs9 [gfp::syp-3]; unc-119 |
| CA1299 | hal-2(me79) / qC1 [dpy-19(e1259) glp-1(q339) qIs26] III; meIs9 (gfp::syp-3); unc-119 |
| CA1300 | syp-3(ok857) htp-3(tm2655) I; ieSi11 [cbunc-119+, Psyp-3::EmeraldGFP::syp-3] II |
| CA1303 | syp-3 (ok857) I; ieSi11 [cbunc-119+, Psyp-3::EmeraldGFP::syp-3] II; htp-1(gk174)/nT1 [unc-?(n754) let-?] (IV;V) |
| CA1309 | unc-119(ed3) III; ieSi21[cbunc-119+, Psun-1::sun-1::mRuby] IV |
| CA1350 | him-3(ie34[mMaple3::him-3]) IV |
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
The synaptonemal complex has liquid crystalline properties and spatially regulates meiotic recombination factors
eLife 6:e21455.
https://doi.org/10.7554/eLife.21455




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}