Distinct memory engrams in the infralimbic cortex of rats control opposing environmental actions on a learned behavior
- The Scripps Research Institute, United States
- University of Sussex, United Kingdom
- University of California San Diego, United States
- National Institute on Drug Abuse/National Institutes of Health/Intramural Research program, United States
Figures
Figure 1
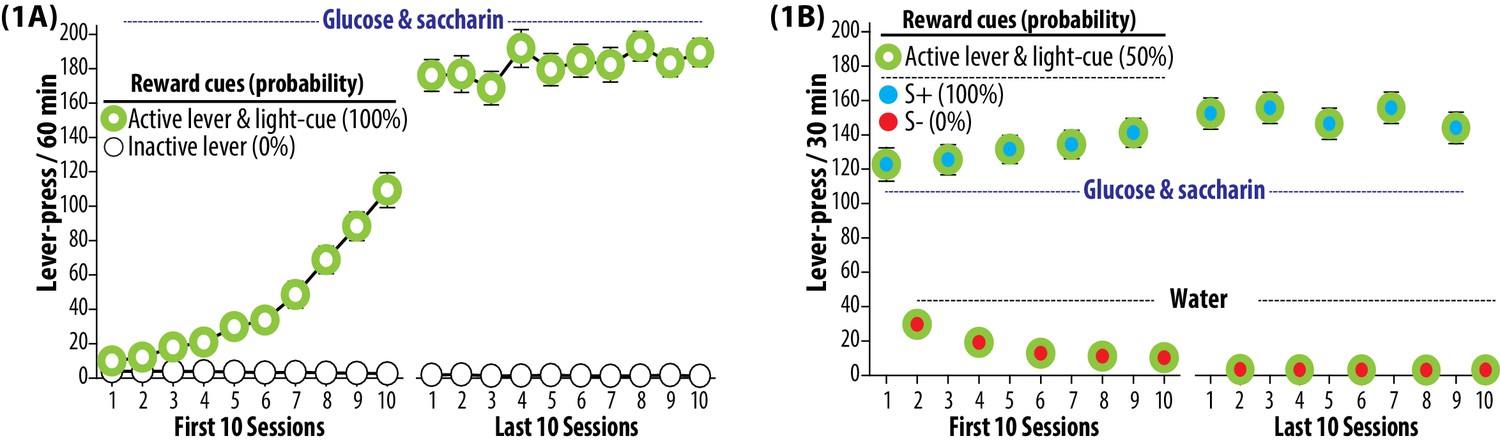
Trainings.
(1A) Self-administration training to establish operant response for a palatable reward. The rats were subjected to once daily operant conditioning sessions (60 min, each) to press an ‘active lever’ for the purpose of gaining a sweet solution (3% glucose and 0.125% saccharin dissolved in water). Each delivery of the sweet solution was paired with a ‘light-cue’. Presses on an ‘inactive lever’ were without a scheduled consequence. Each rat was required to satisfy preset training criteria, and underwent a total of 35 to 42 sessions. Group means of the total number of lever presses per session (±SEM) during the first and last 10 sessions are depicted. During this phase, both active lever and light-cue predicted the availability of sweet solution 100% of the time. N = 90. (1B) Discrimination training to establish two distinct auditory cues (white noise and beeping tone) as the discriminative stimuli predictive of reward availability (S+) and omission (S-). Each rat was subjected to alternating once daily operant conditioning sessions (30 min, each) to press the active lever for the purpose of gaining the sweet solution (preceded and accompanied by the S+ auditory cue) or plain water (preceded and accompanied by the S- auditory cue). Each delivery of either sweet solution or water was paired with the light-cue. During this phase, each experimentally manipulated stimulus was conditioned to predict glucose and saccharin at the following probabilities: active lever and light-cue (50%), S+ (100%) and S- (0%). Each rat was required to satisfy three preset training criteria, and underwent a total of 92 to 98 sessions (46 to 49 S+ sessions and 46 to 49 s- sessions). Group means of the total numbers of lever presses (±SEM) recorded during the first and last 10 sessions are depicted. N = 90.
Figure 2
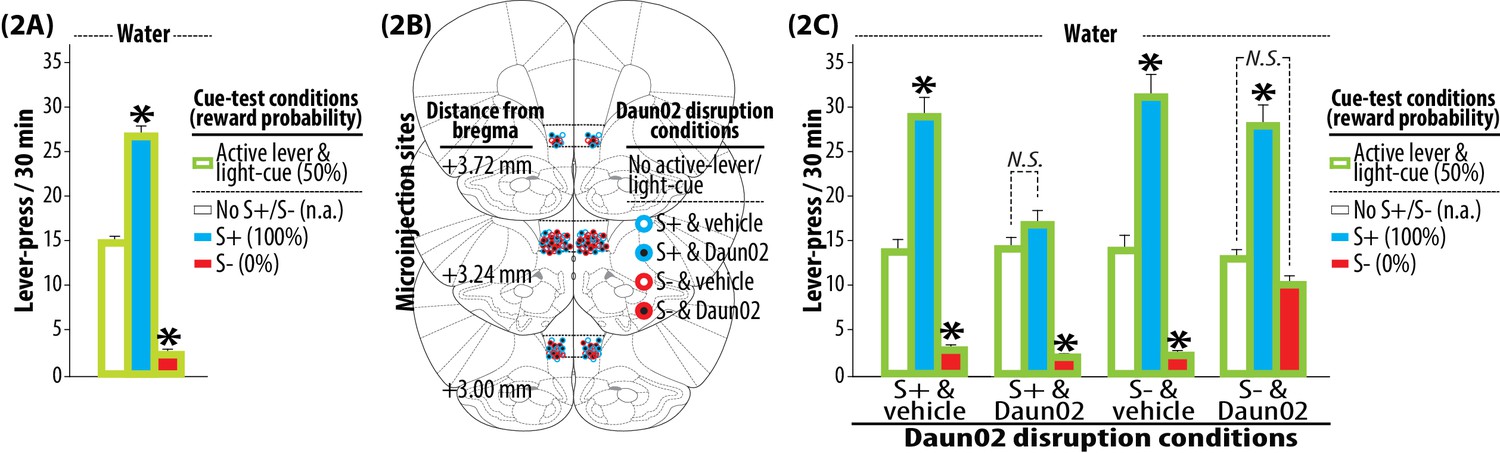
Tests and Daun02 disruption.
(2A) Pre-disruption tests for the bidirectional modulation of reward seeking by S+ or S-. Each rat was subjected to three once daily cue-test conditions (30 min, each): (1) active lever and light-cue without S+ or S- (No S+/S-), (2) active lever, light-cue and S+, (3) active lever, light-cue and S-. Under all cue-test conditions, a press on the active lever delivered water and illuminated the light-cue. The sweet solution was not available. Group means of the total number of active lever-presses under each cue-test condition (+SEM) are depicted. N = 90. *p<0.001 (vs. No S+/S-). (2B) Daun02 disruption for the activity-targeted inhibition of neural ensembles in IL reactive to S+ or S-. The rats were randomly assigned to one of four experimental groups defined by the type of disruption cue (S+ or S-) and microinjection (Daun02 or vehicle). The rats were exposed to S+ or S- (90 min), and then received a bilateral microinjection of Daun02 (2.0 µg/0.5 µl/side) or vehicle (0.5 µl/side) into IL. In order to disrupt neurons specifically activated by S+ or S-, both active lever and light-cue were withheld (‘No active lever/light-cue’). Representative sites of microinjection are depicted. (2C) Post-disruption tests for the bilateral modulation of reward seeking by S+ or S-. Each rat was subjected to three once daily cue-test conditions. The experimental schedule effective during the Pre-disruption tests (2A) was applied. Group means of the total number of active lever-presses under each cue-test condition (+SEM) are depicted. N = 21–24, each. *p<0.01–0.001 (vs. No S+/S-).
Figure 3
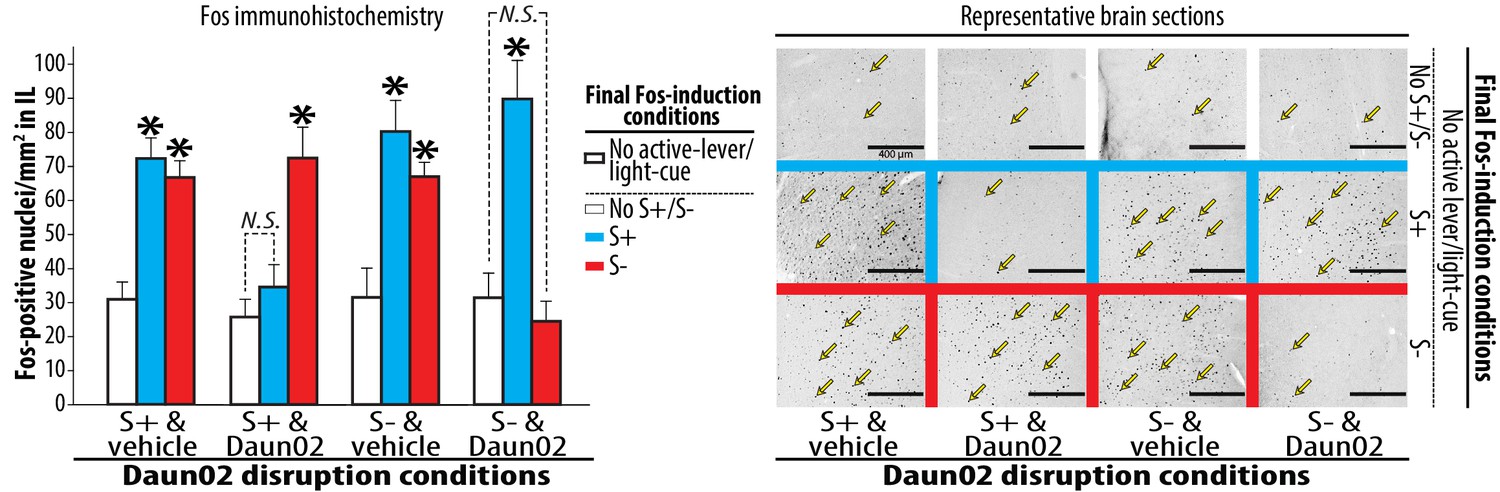
Neural ensembles in IL reactive to S+ or S- following the activity-targeted disruption by Daun02.
Each of the four experimental groups, prepared and used for the Daun02 disruption (Figure 2B) and Post-disruption tests I (Figure 2C), was further randomly divided into three groups defined by the type of final Fos induction condition (S+, S- or No S+/S-). The rats were exposed to S+, S- or No S+/S- (control), then deeply anesthetized and euthanized. Brains were collected, sectioned (40 µm), processed for Fos immunohistochemistry. Fos-positive nuclei from sampling areas around the IL microinjection sites were quantified double-blindly. The average numbers of Fos-positive nuclei per mm2 were calculated for each rat, and used for statistical analyses. Yellow arrows represent typical Fos-positive nuclei. Group means of these average numbers (+SEM) are depicted. N = 7–9, each. *p<0.05–0.01 (vs. No active lever/light-cue).
Figure 4
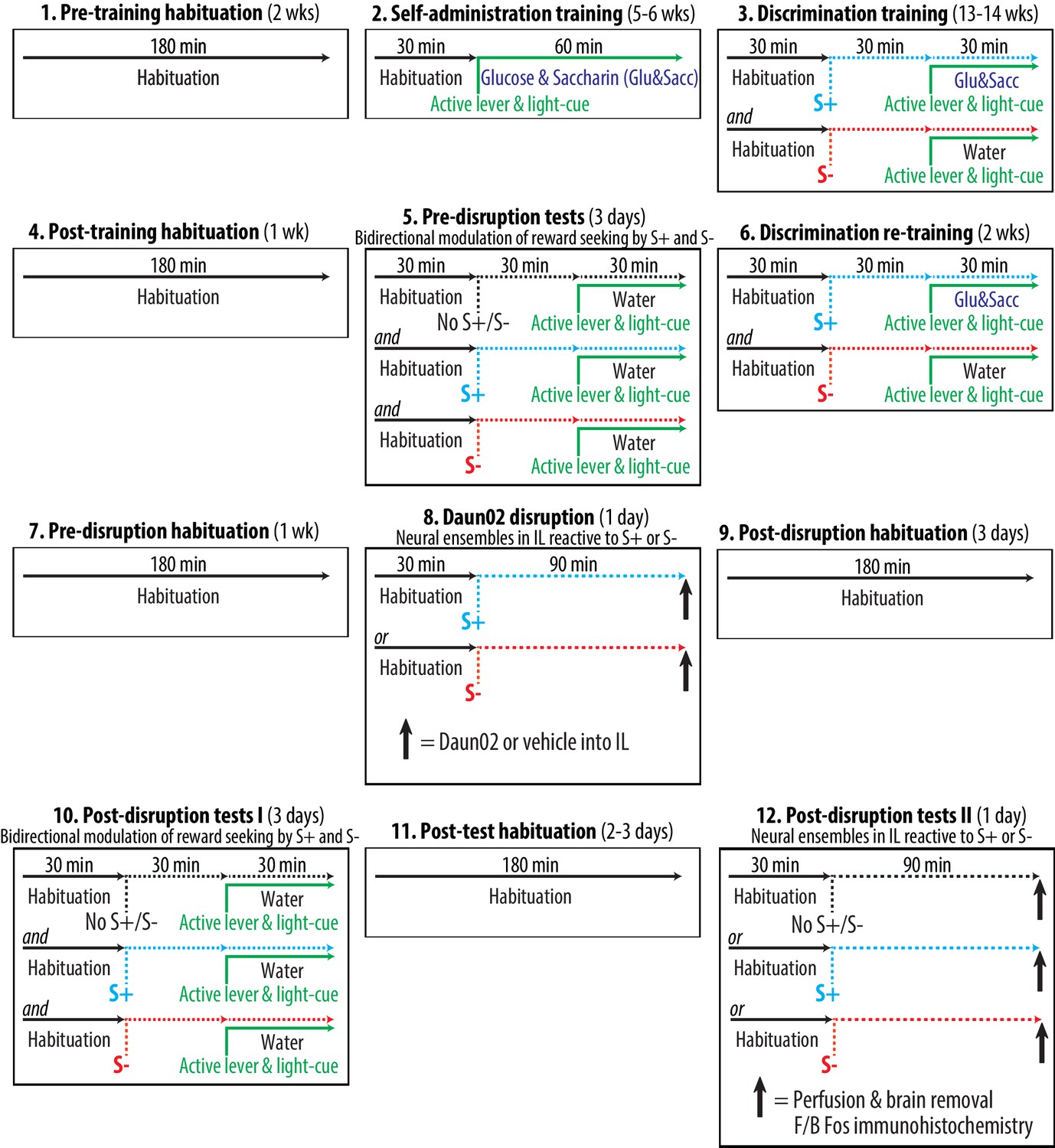
Experimental time course and schedule.
https://doi.org/10.7554/eLife.21920.005Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Distinct memory engrams in the infralimbic cortex of rats control opposing environmental actions on a learned behavior
eLife 5:e21920.
https://doi.org/10.7554/eLife.21920




{kind=link}
{kind=link}
{kind=link}
{kind=link}