Coordinated control of terminal differentiation and restriction of cellular plasticity
- Howard Hughes Medical Institute, Columbia University, United States
Figures
Figure 1 with 3 supplements
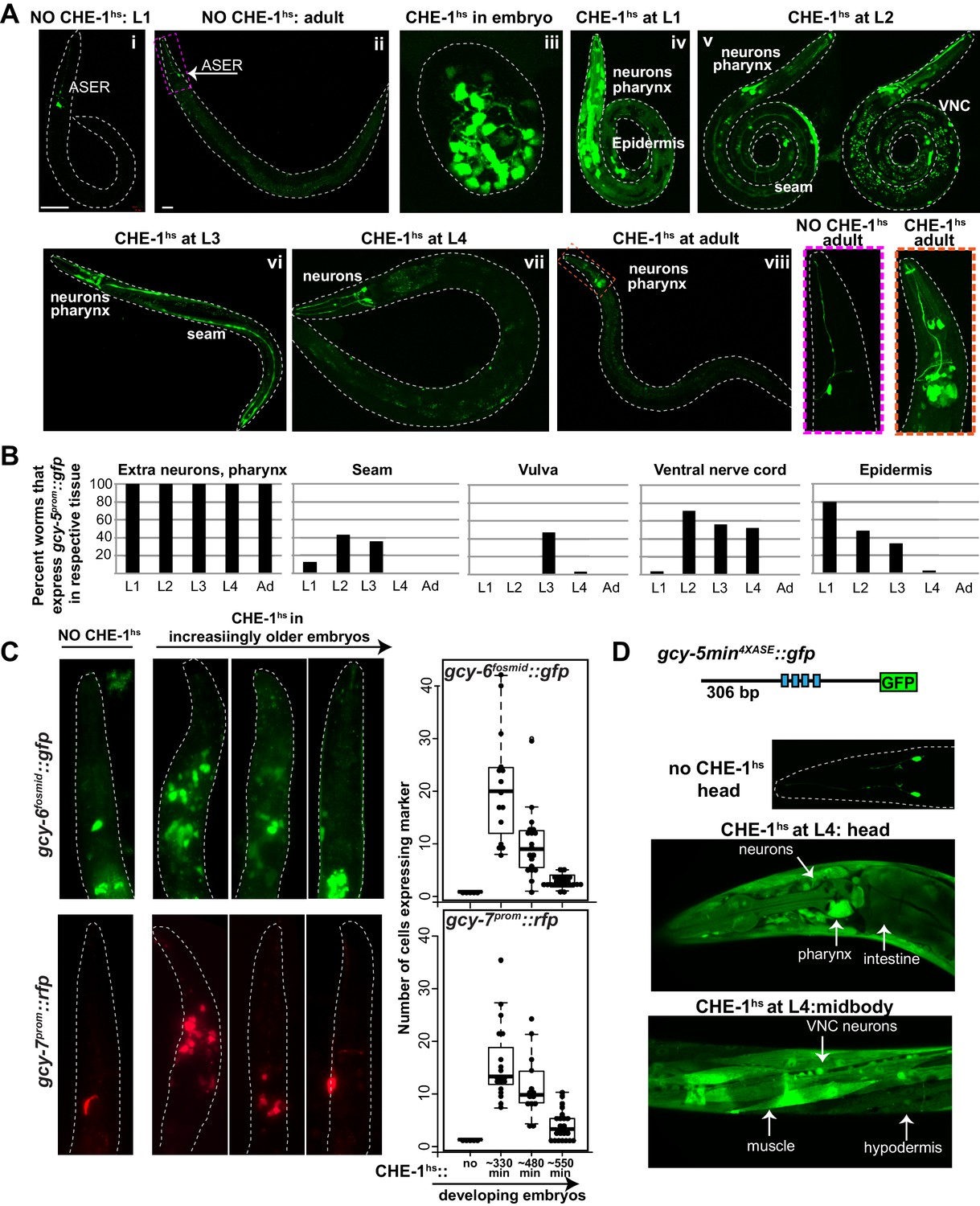
Expression of ASE markers in response to CHE-1hs at different developmental stages.
(A) Expression of gcy-5prom::gfp becomes less broad as CHE-1hs is induced at later stages. (i) L1 worm without CHE-1hs, bright expression in ASER; (ii) adult without CHE-1hs, expression in ASER (arrow); (iii) embryonic CHE-1hs, broad gcy-5prom::gfp induction in deformed embryos; (iv) L1 CHE-1hs,expression of gcy-5prom::gfp in various tissues including hypodermis, various neurons, muscles, pharynx. (v) L2 CHE-1hs at two different focal points, gcy-5prom::gfp in seam, muscles, ventral nerve cord, other neurons and pharynx; (vi) L3 CHE-1hs, gcy-5prom::gfp in seam, vulva cells, neurons and pharynx; (vii) L4 and (viii) adult CHE-1hs, gcy-5prom::gfp is seen only in neurons and pharynx. Higher magnification of the heads in (i) and (viii). (B) Quantification of the expression of gcy-5prom::gfp in various tissues after CHE-1hs induction at various ages, as listed on the X-axis. n => 20 for each stage. (C) Expression of gcy-6fosmid and gcy-7prom in response to CHE-1hs at different stages. Quantification of the ectopic expression: every dot in this graph represents a worm and the number on the y-axis represents the total number of marker-expressing neurons counted. Ectopic expression decreases as the age of the embryos being heatshocked increases. (D) A synthetic ~300 bp region of the gcy-5 promoter in which the ASE motif has been multimerized four times can be broadly induced by CHE-1hs at all stages, including L4s, as shown here. Some of the tissues that express gcy-54XASE::gfp are labeled. gcy-54XASE::gfp is normally expressed only in the two ASEs.
Figure 1—figure supplement 1
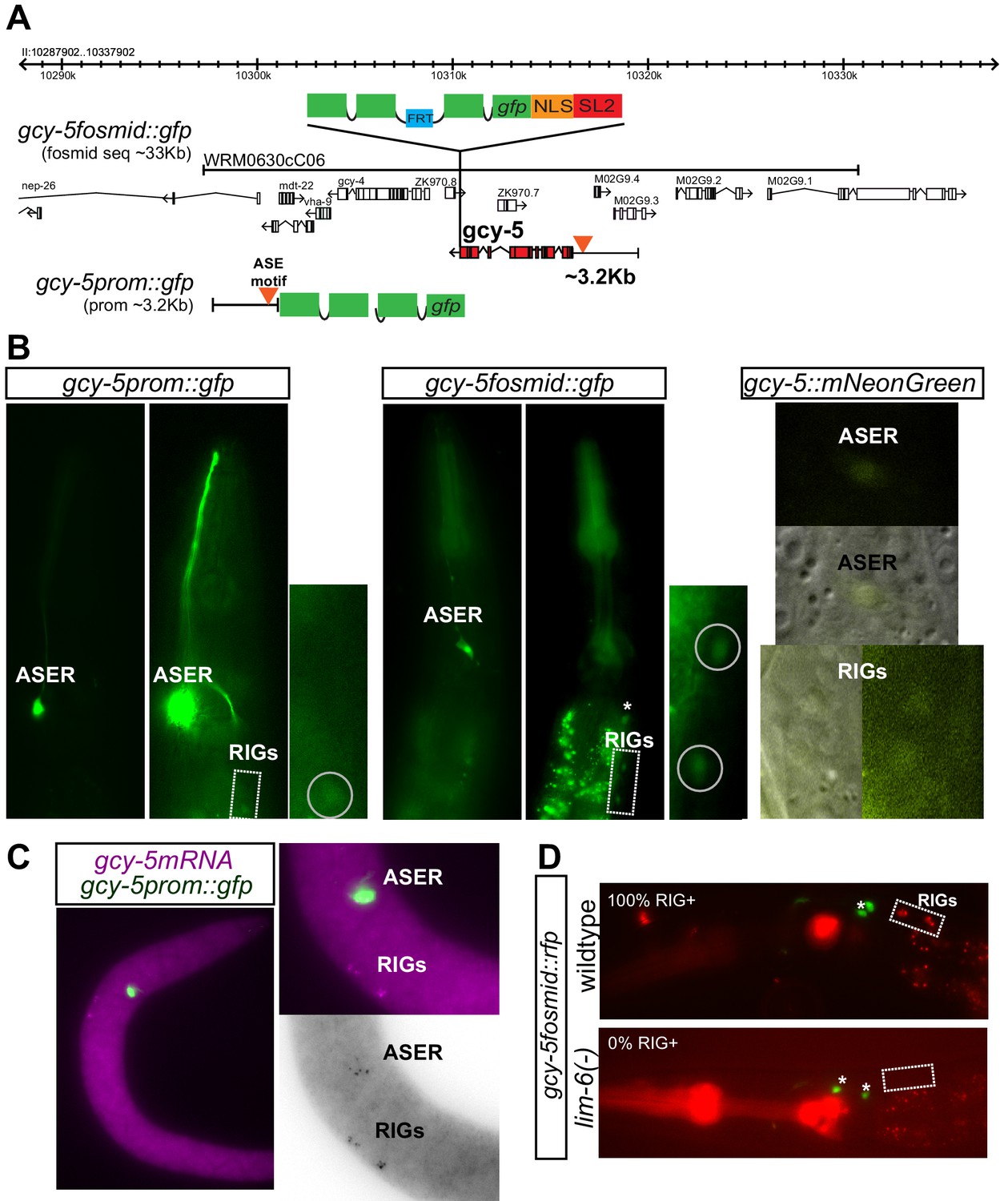
Tools used for the detection of gcy-5 expression.
(A) Schematic representation of the fosmid and promoter reporters of gcy-5. The fosmid contains ~33 kb of DNA surrounding the gcy-5 locus. A trans-splicing sequence, SL2, and GFP are recombineered after the stop codon of gcy-5. This ensures that gcy-5 and gfp get transcribed together but get spliced and translated separately. The promoter reporter contains 3.2 kb of information upstream of the start codon of the gcy-5 gene fused to gfp. (B) Images of heads of transgenic neurons containing the gcy-5prom::gfp, gcy-5fosmid::gfp, and a CRISPR-engineered allele containing an SL2::mNeonGreen inserted into the genomic gcy-5 locus after the gcy-5 stop codon. RIG expression of the promoter-based reporter is not fully penetrant. Expression of the CRISPR allele is very dim. Asterisks indicate expression of co-injection marker. (C) FISH for gcy-5 mRNA also shows expression in the ASER and RIG neurons. (D) Expression of gcy-5 in the RIGs is dependent on lim-6, not che-1. Two RIG neurons can be seen in 100% of the wildtype worms. 0 RIGs can be seen in 100% of lim-6 mutant worms (n = 20 each). Asterisks indicate expression of co-injection marker.
Figure 1—figure supplement 2

Expression of gcy-5 in response to CHE-1hs at different developmental stages.
(A) Expression of gcy-5fosmid in the VNC after CHE-1hs at L2 is enriched in newly born neurons based on overlap with rab-3low (born at L2) and rab-3high (born during embryonic stages). (B) Ectopic induction of endogenous gcy-5 in response to heat-shock. FISH for gcy-5 mRNA after CHE-1hs induction in MiniMos strain containing a single copy hsprom::che-1 transgene integrated into the genome and gcy-5prom::gfp. Expression of gcy-5 mRNA overlaps with gcy-5prom::gfp. (C) che-1 mRNA induction after heatshock. Heatshock induction results in robust che-1 mRNA expression at the adult stage. Pictures show smFISH of che-1 mRNA in heads of adult worms before and after heat-shock. In the absence of heat-shock, a few mRNA molecules can be detected, however after heat-shock the expression is much more robust. (D) CHE-1 protein induction after heatshock. The heatshock-induced CHE-1 contains an HA tag. Antibody staining for HA tag shows the presence of protein in all cell types. Mid-bodies of adult worms are shown. No CHE-1::HA protein is detected without heat-shock, while nuclear localized protein is clearly visible after heat- shock. Some tissues, such as hypodermis and muscle, show stronger induction of protein than other tissues.
Figure 1—figure supplement 3
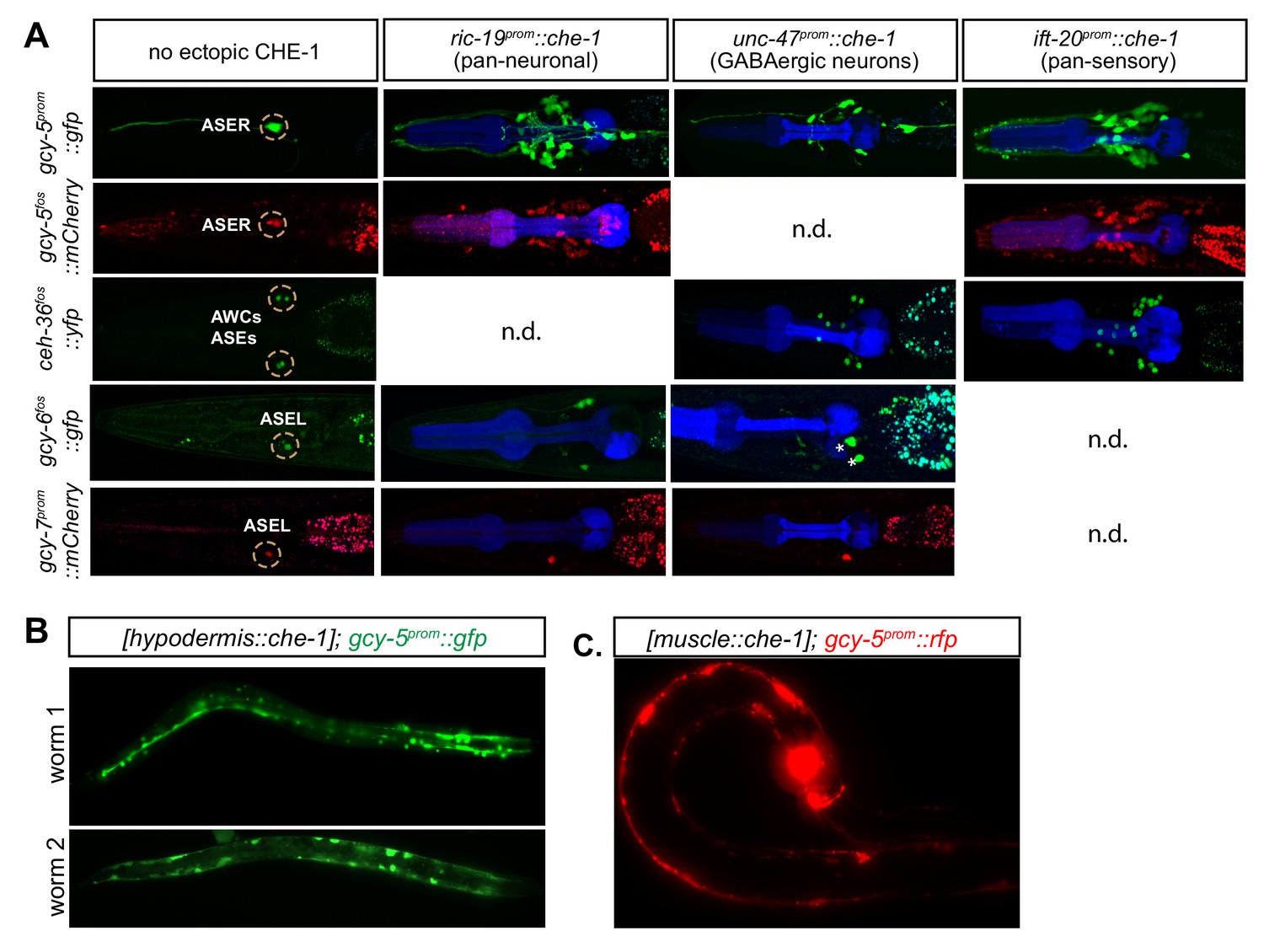
Expression of ASE markers in response to cell type-specific, but not temporally controlled, che-1 expression.
(A) For each worm, promoters driving post-mitotic che-1 expression are listed on the horizontal axis and ASE markers used are listed on the vertical axis. gcy-5prom::gfp, gcy-5fosmid::rfp, and ceh-36fosmid::yfp are broadly expressed in response to che-1, whereas gcy6fosmid::gfp, and gcy-7prom::rfp are not. Some marker, promoter combinations were not determined (n.d.) because of unsuccessful crosses. (B) gcy-5prom is broadly expressed in the hypodermis and (C) post mitotic muscles in response to CHE-1 induction in these tissues.
Figure 2
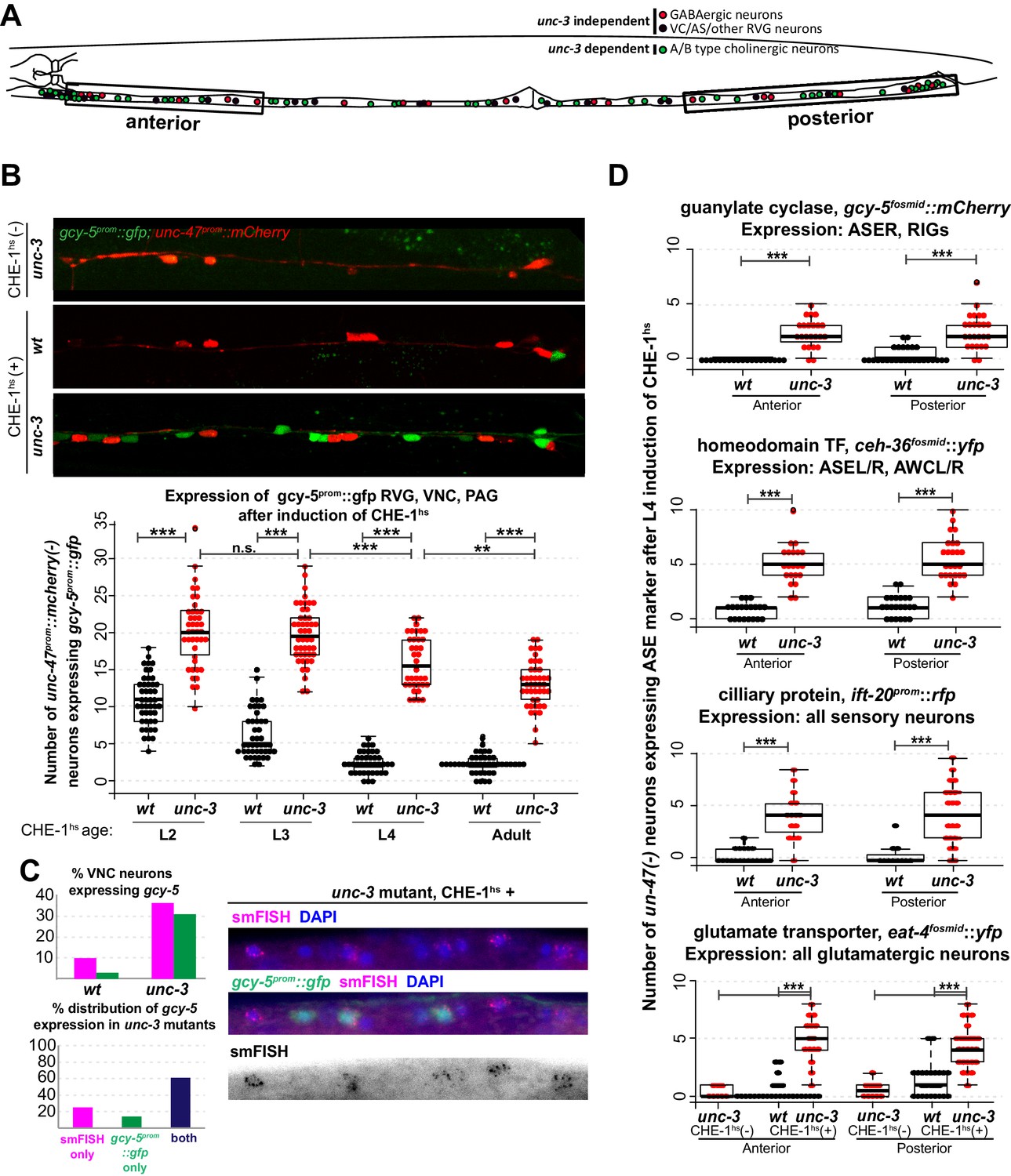
Several CHE-1 targets retain their CHE-1 responsiveness in unc-3 mutant cholinergic MNs.
For all heatshock experiments, three biological replicates were performed. The student t-test is used for statistical comparisons, *p<0.05, **p<0.01, ***p<0.001, n.s.= p>0.05. (A) A schematic representation of the neurons in the RVG, VNC, and PAG of the worm. In heatshock experiments represented in B, C, and D, the number of ASE marker expressing cells in the RVG, VNC, and PAG were scored. In B and D, unc-47, a marker of GABAergic identity (is used in the background and only unc-47(-) neurons are scored. This strategy ensures proper scoring of the ~50 unc-3 dependent cholinergic MNs . (B) Images of wildtype and unc-3 worms with and without CHE-1hs and a quantification of gcy-5prom induction at various stages of development. Every dot in this plot represents an individual worm. The entire RVG, VNC, and PAG of the worm is scored. A significantly higher number of gcy-5prom::gfp neurons are seen in unc-3 mutants as compared to wildtype in response to CHE-1hs induction. No gcy-5prom::gfp neurons are seen in unc-3 mutant worms that get heatshocked but do not contain the heatshockprom::che-1 array (0 cells in >20 worms). (C) gcy-5prom transgene (green bar) and gcy-5 mRNA (smFISH; pink bar) induction in heatshocked worms carrying a single copy insertion of heatshockprom::che-1. Induction of gcy-5 endogenous mRNA (smFISH) is similar to the induction of the gcy-5prom reporter. (D) Expression of various other ASE markers is seen in a significantly higher number of unc-47(-) RVG, VNC, and PAG neurons in unc-3 mutants as compared to wildtype after CHE-1hs induction. The scored markers and their wildtype (without CHE-1hs) expression patterns are listed above each plot. Every dot in these plots represents an individual worm. Markers are scored only in the anterior and posterior regions of the worm, as represented in A. These worms were heatshocked at the L4 stage.
Figure 3 with 1 supplement
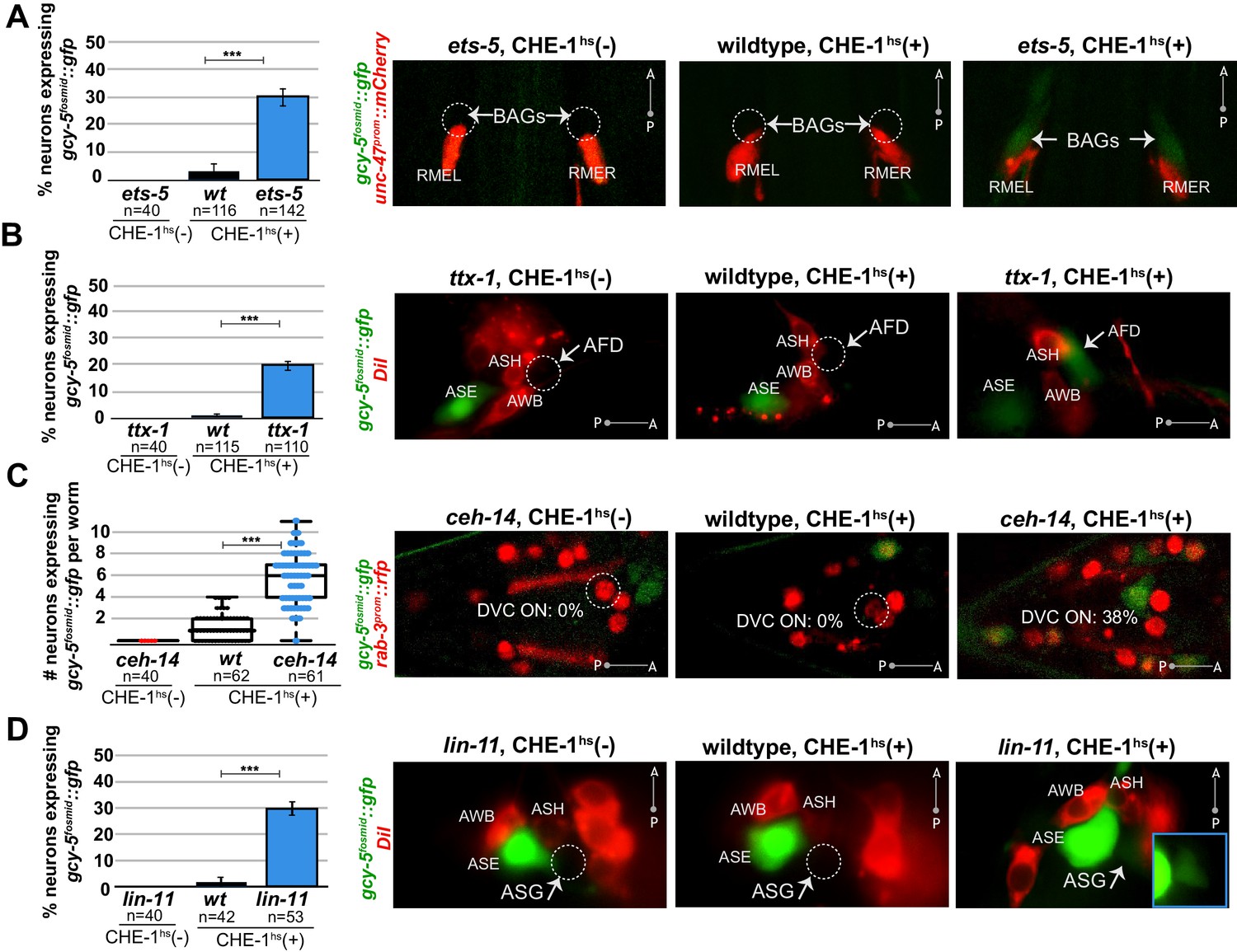
Several distinct terminal selectors control restriction of cellular plasticity in distinct neuron types.
CHE-1hs was induced in adults for all experiments except lin-11, for which L4s were heatshocked (lin-11 mutant adults are very short-lived due to internal hatching of progeny). The markers in red are used to help identify the neuron of interest. gcy-5fosmid::gfp is more frequenctly activated by CHE-1hs in BAG neurons of ets-5 mutants (panel A), in AFD neurons of ttx-1 mutants (panel B), in DVC and a few other unidentifiable neurons in the tail of ceh-14 mutants (panel C), and in ASG neurons of lin-11 mutants (panel D). All error bars represent SEM. Data are accumulated from three independent heatshock experiments. The student t-test is used, ***p<0.0001, n.s.= p>0.01.
Figure 3—figure supplement 1
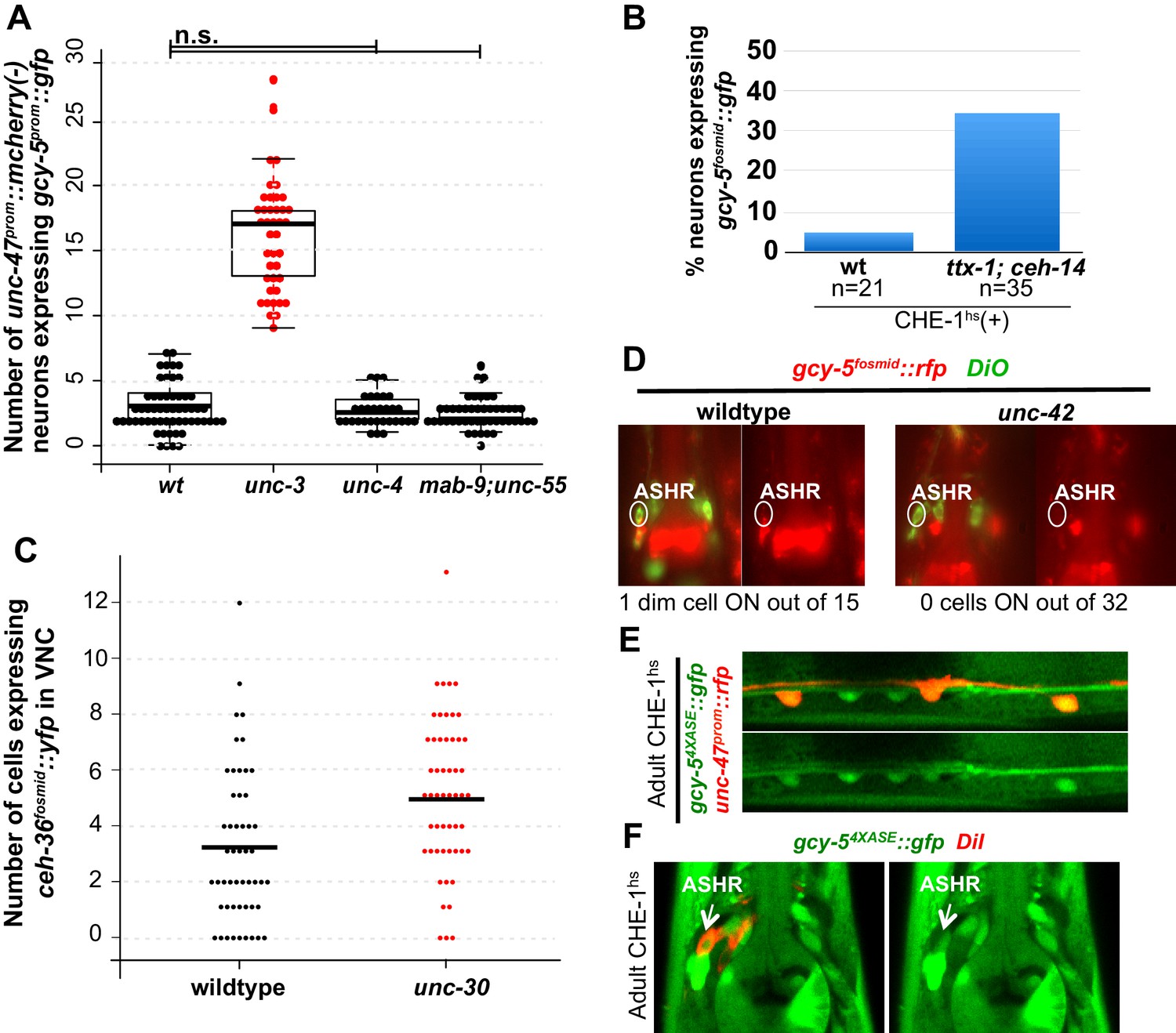
Phenotypes of additional mutants.
CHE-1hs was induced in adults for all experiments. Every dot in (A) and (B) represents an individual worm. The student t-test is used for statistical comparisons, *p<0.05, **p<0.01, ***p<0.001, n.s.= p>0.05. (A) mab-9;unc-55 double mutants and unc-4 mutants are not significantly different from wildtype and are significantly different from unc-3. (B) The phenotype in the AFD neurons remains partially penetrant in ttx-1; ceh-14 double mutants, similar to ttx-1 single mutants. (C) An increase of ceh-36fosmid::yfp expression is seen in 2 neurons in unc-30 compared to wildtype worms in unc-30 mutants. As there are 19 total unc-30 dependent neurons, this difference does not seem biologically significant. Every dot in the graph represents an individual worm. (D) No gcy-5fosmid::rfp expression is seen in the ASH neurons of unc-42 mutants. (E) gcy-54XASE::gfp can be induced in wildtype GABAergic neurons after CHE-1hs induction in adults, showing that CHE-1hs is functional in these neurons. (F) gcy-54XASE::gfp can also be induced in wildtype ASH neurons after CHE-1hs induction in adults. All error bars represent SEM.
Figure 4
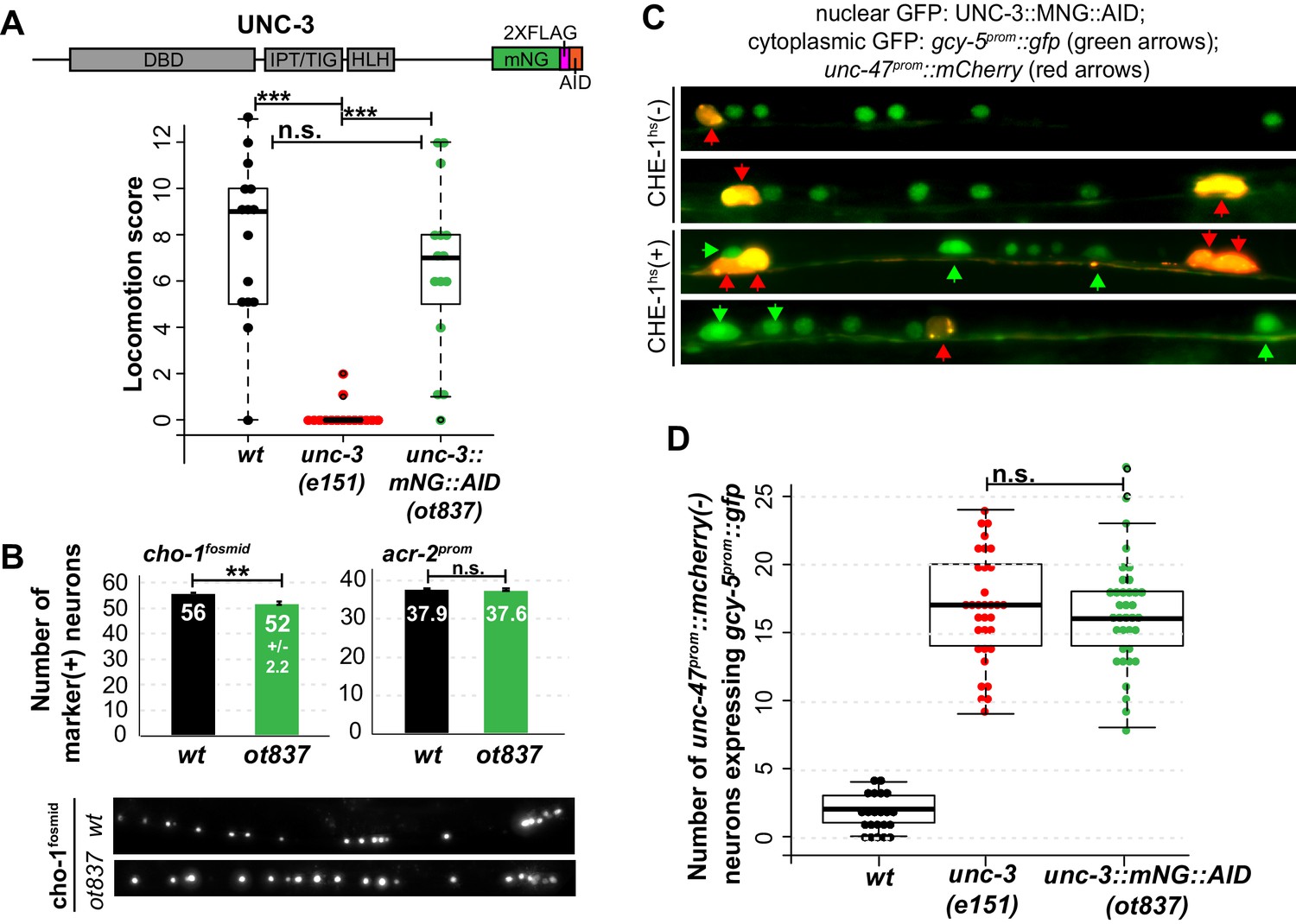
Genetic separation of identity-specification and plasticity restriction.
All error bars represent SEM. The student t-test is used, *p<0.05, **p<0.01, ***p<0.001, n.s.= p>0.05. (A) An mNeonGreen::AID tagged allele of unc-3, ot837, does not have the locomotion defects of an unc-3 null allele. (B) ot837 worms show near wildtype and wildtype induction of cho-1fosmid and acr-2prom respectively, whereas in unc-3 null mutants these genes are strongly repressed (Kratsios et al., 2011). cho-1fosmid expression is missing in ~4 neurons in ot837, but levels of expression in the neurons that retain expression look wildtype. (C) ot837 behaves like the unc-3 null with respect to restriction of plasticity. Induction of gcy-5prom is indistinguishable in ot837 and the unc-3 null allele after CHE-1hs induction. (D) Images of gcy-5prom expression in ot837 worms after CHE-1hs induction. In these images the nuclear green signal comes from the mNG tagged UNC-3 protein, the cytoplasmic GFP from gcy-5prom (green arrows), and the red cells are GABAergic (red arrows).
Figure 5

Assessing chromatin state of a gcy-5 transgenic locus with the LacI/LacO dot assay.
The student t-test is used for statistical comparisons, *p<0.05, **p<0.01, ***p<0.001, n.s.= p>0.05. (A) Schematic representation of the spot assay. A transgene containing the gcy-5prom::rfp sequence and LacO binding sites is integrated into the genome of worms that ubiquitously express LacI::GFP. In the resulting transgenic worm, every nucleus contains two GFP spots representing the LacI::GFP bound to the gcy-5 LacO array. The structure and location of this locus can be visualized by GFP. (B) The gcy-5 arrays localize to the nuclear periphery in all cells in which expression was checked, including the ASEs. The array is transcriptionally active in the ASER, as evident by the presence of RFP, suggesting that this locus localizes to the nuclear periphery regardless of its transcriptional state. An identically built ace-2prom::rfp LacO array shows sub-nuclear localization that correlates to its transcriptional state. In cells that express ace-2prom::rfp, as seen by the presence of RFP, the GFP spots are diffused and in the nuclear lumen, suggesting that the locus is decompacted. In cells that do not express ace-2prom::rfp, the GFP spots are compact and localized to the nuclear periphery. Two independent array integrants examined for both ace-2 and gcy-5 showed similar results and one is shown here. (C) Worms were fixed and stained with DAPI for quantification of the GFP spots as an estimation of array structure. (D) ace-2 arrays were significantly larger when transcriptionally active as opposed to inactive in the VNC neurons of wildtype worms compared to unc-3 mutants. In (D, E, F) each dot represents a single nucleus and the number on the y-axis is the total pixel count occupied by the LacI/O spots or H3K27me3 per neuron. (E) gcy-5 arrays were significantly larger in unc-3 mutant neurons than in wildtype. (F) Control staining for H3K27me3 in gcy-5 LacO array containing worms showed no difference between wildtype and unc-3 and the size of the gcy-5 LacI/O spots do not change in the hypodermal nuclei of wildtype vs. unc-3 worms.
Figure 6 with 1 supplement

Interactions of unc-3 with chromatin mutants.
CHE-1hs was induced at the L4 stage. For each graph, the data cumulatively represent three independent heatshock experiments in which all genotypes were scored in parallel. Every dot represents an individual worm. One way ANOVA with post-hoc Bonferroni and Scheffé tests for multiple comparisons were performed independently for each group of strains tested together. *p<0.05, **p<0.01, ***p<0.001, n.s.= p>0.05. (A) mes-2 mutants show increased induction of gcy-5prom::gfp in response to CHE-1hs and show an additive phenotype with unc-3, suggesting H3K27me3 regulates restriction in a parallel pathway with unc-3. The mes-2 and unc-4 mutations are genetically linked. unc-4 does not play a role in restriction, but it was used in all strains tested in this set of experiments to ensure similar genetic backgrounds. (B) Both met-2 and set-25 mutants have significantly more gcy-5prom::gfp expressing cholinergic MNs compared to wildtype. However, for set-25 the increase is only modest, and a met-2 set-25 double mutant is not significantly different from met-2, suggesting met-2, has a greater role in making the gcy-5prom::gfp locus inaccessible to CHE-1. An additive phenotype is not evident in met-2; unc-3, set-25; unc-3, or met-2 set-25; unc-3, suggesting that these factors might act in the same pathway to restrict the accessibility of gcy-5prom::gfp. No ectopic expression of gcy-5prom::gfp is seen in these mutants in the absence of CHE-1hs (not shown, n > 50 worms). (C) HP1 mutants also show increased induction of gcy-5prom::gfp, and slightly enhance the phenotype of unc-3 suggesting they have roles independent of met-2 set-25 dependent H3K9me. No gcy-5prom::gfp is induced in hpl-1;hpl-2 double mutants in the absence of CHE-1hs (not shown, n > 50 worms). (D) Representative images of scored worms. (E) Induction of CHE-1hs with the single copy transgene also results in increased induction of gcy-5prom::gfp in the VNC of met-2 set-25 mutants and the number of gcy-5prom::gfp and gcy-5 mRNA expression cells in the double mutant worms are similar showing that the phenotype in (B) is not simply a result of derepressed repetitive transgenes.
Figure 6—figure supplement 1
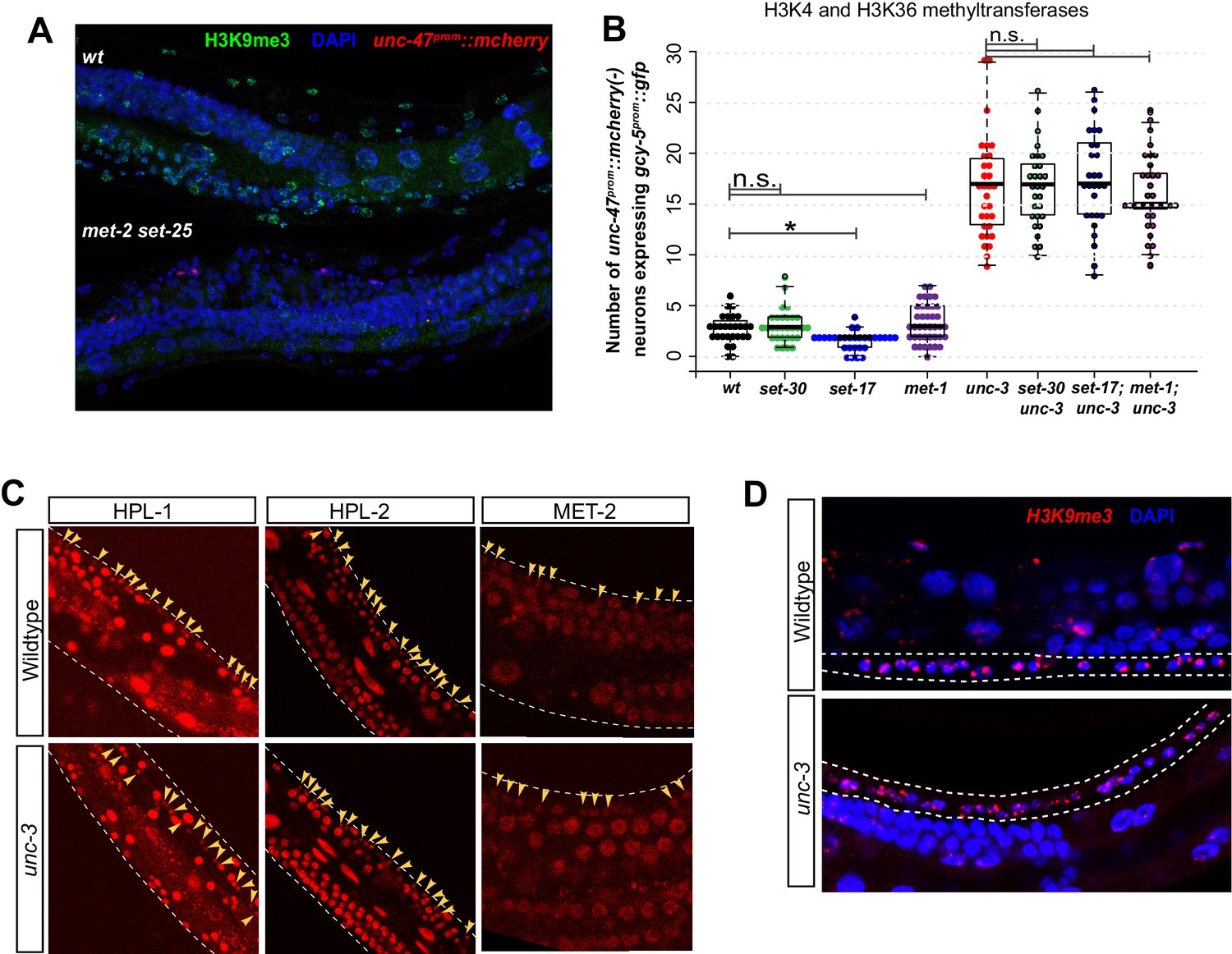
H3K9 in wildtype and unc-3 mutant worms.
(A) H3K9me3 is not detectable in met-2 set-25 double mutants at the L4/adult stage (n > 50 for each). Wildtype worms without unc-47prom::rfp and double mutant worms containing unc-47prom::rfp were fixed, stained, and imaged together. (B) Mutants of methyl transferases that deposit active histone marks do not show any increased or modified induction of gcy-5prom::gfp after CHE-1hs induction. (C) H3K9me3 staining is detectable in the VNC neurons of wildtype and unc-3 mutant worms. (D) CRISPR engineered fluorescently tagged HPL-1, HPL-2, and MET-2 proteins are detectable in the VNC neurons of wildtype and unc-3 mutant worms. Yellow arrows point at VNC neuronal nuclei.
Figure 7
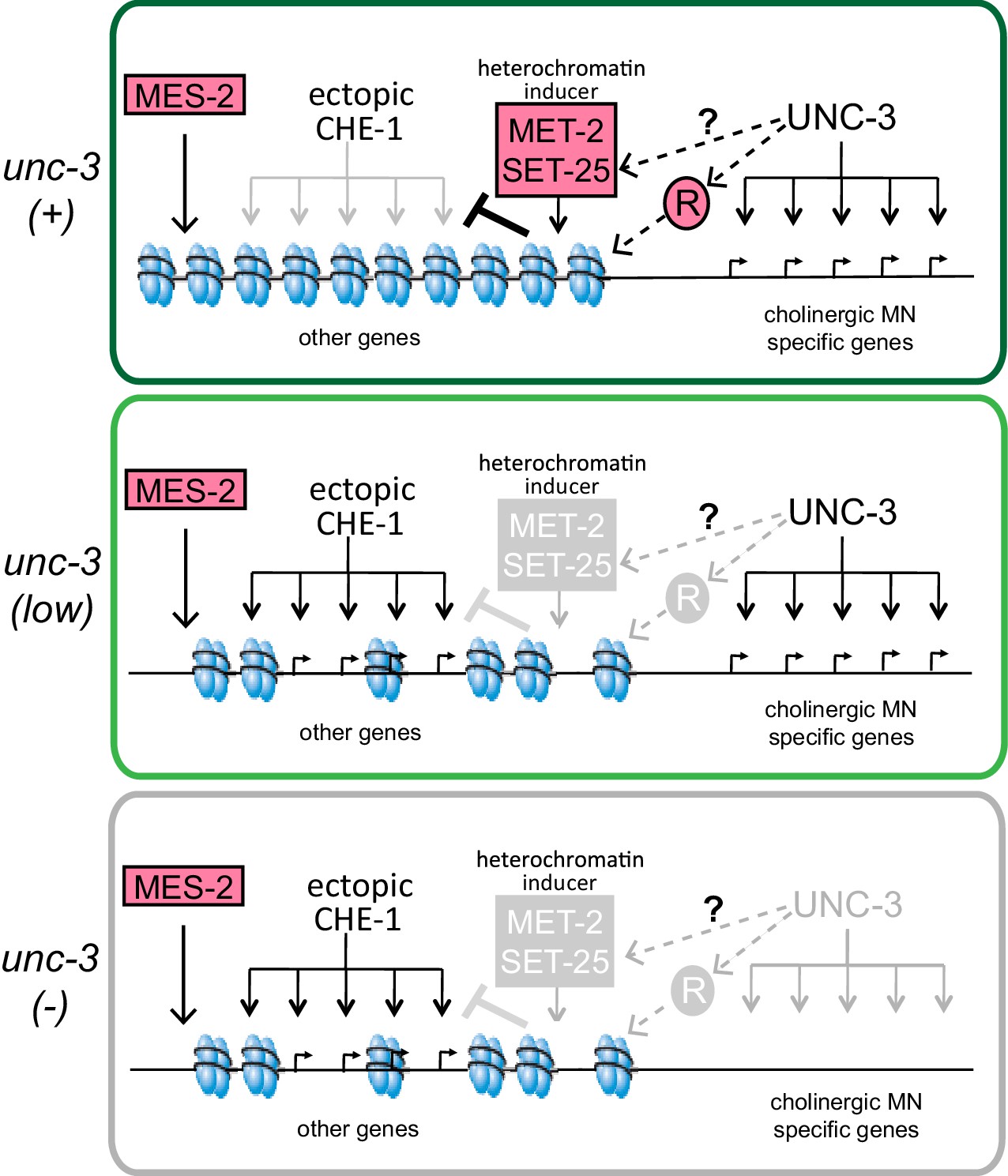
A model for the role of terminal selectors in restriction of plasticity.
Grey indicates lack of protein activity. In wildtype neurons, unc-3 activates expression of target genes to specify cholinergic MN identity but it also triggers the restriction of plasticity by activating pathways that repress genes not required for cholinergic MN identity. The repressed genes include ASE effector genes, which cannot be activated by ectopic CHE-1 in wildtype MNs. To restrict plasticity, unc-3 may regulate the correct distribution of H3K9 methylation through unknown pathways and may also activate additional repressors (R). These repressor mechanisms may then act to 'survey' the genome for genes that have not been activated during identity specification and transform them into a 'non-activatable' stage via H3K9-dependent heterochromatinization. unc-3 is required to trigger this surveillance mechanism by controlling the function or expression of the genes involved in this process. Independently, mes-2-dependent H3K27 methylation also plays a role in gene silencing. Upon lowering levels of unc-3 activity (ot837 allele) activation of cholinergic MN genes still takes place, however the unc-3 dependent pathways that repress other identity genes are not activated, as shown by the ability of CHE-1 to activate its target genes. When unc-3 is absent, neither specification nor restriction takes place so that cholinergic MN specific genes are not expressed and CHE-1 targets can be activated. A loss of both unc-3 and mes-2 results in even more open chromatin, and an increased activation of CHE-1 target genes.
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Coordinated control of terminal differentiation and restriction of cellular plasticity
eLife 6:e24100.
https://doi.org/10.7554/eLife.24100




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}