A CDC25 family protein phosphatase gates cargo recognition by the Vps26 retromer subunit
- Yale School of Medicine, United States
- University of Iowa, United States
Figures
Figure 1 with 1 supplement
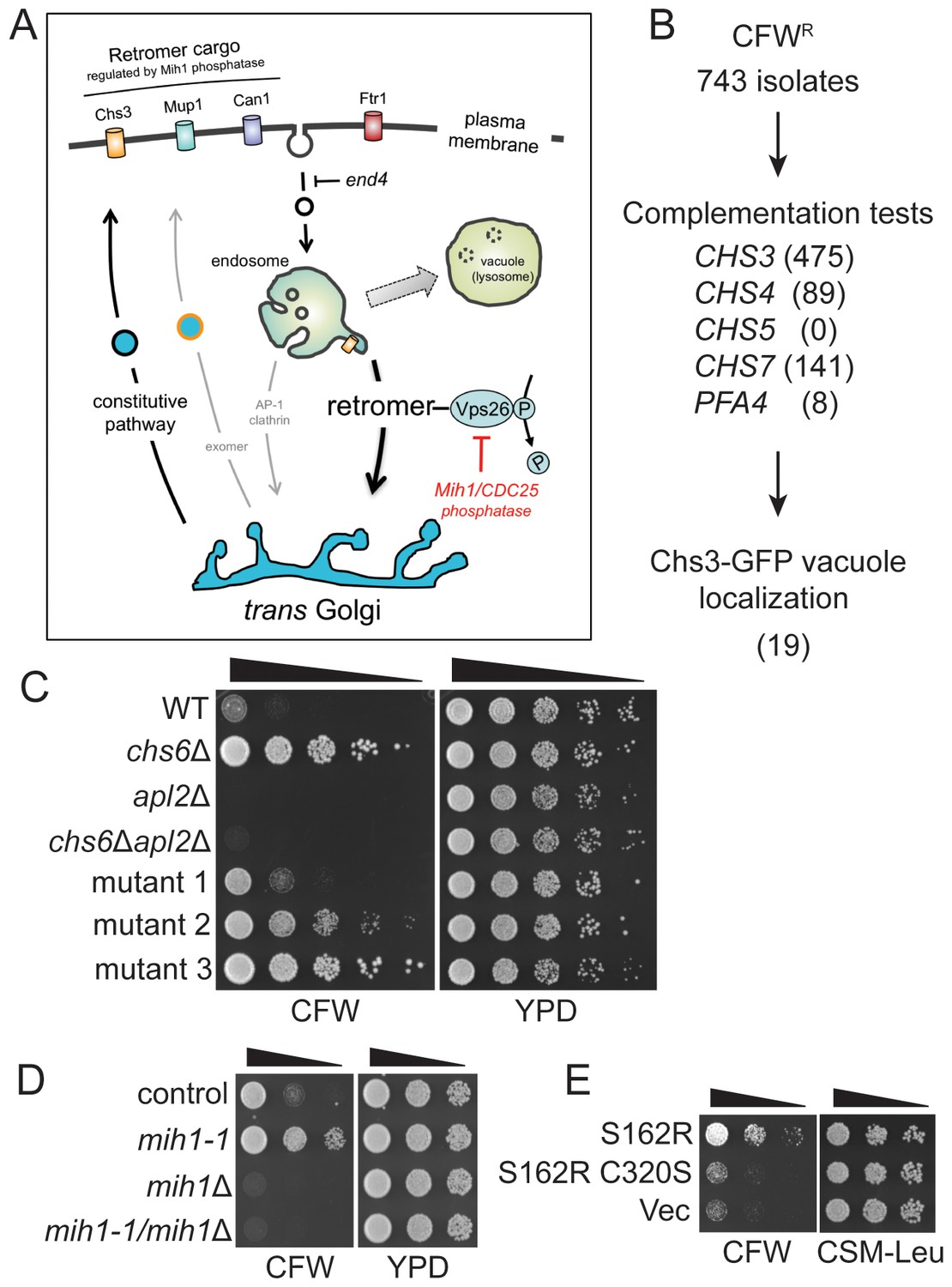
Selection of yeast mutants resistant to calcofluor white.
(A) Schematic diagram of post-Golgi trafficking pathways. In wild-type cells, Chs3 is retained at the Golgi-endosome interface via AP-1 dependent retrieval from the endosome, and is delivered to the plasma membrane via Golgi-derived exomer coated vesicles (gray arrows). In cells used for the genetic selection in this study, chitin synthase 3 (Chs3) is diverted into the constitutive secretory pathway due to deletions of the Chs6 exomer subunit and the Apl2 AP-1 subunit (chs6Δapl2Δ). Endocytosis of Chs3 and other plasma membrane proteins, which is attenuated by the end4 mutation, delivers the internalized proteins to the endosome, where some are sorted by retromer into a recycling pathway, while others are retained in the endosome and delivered to the lysosome-like vacuole and degraded. (B) Strategy to identify novel mutations affecting Chs3 trafficking. The flowchart lists the number of mutants remaining after each step of selection. (C) Representative calcofluor white (CFW) resistant strains. Serial dilutions (1:10; indicated by the black triangle above the photograph) of the indicated strains were spotted onto YPD medium with or without 100 μg/ml CFW. The plates were incubated for three days at 30°C and then photographed. (D) Mutations in MIH1 alter sensitivity of growth to CFW. Serial dilutions (indicated by a triangle above the photograph) of control cells of the indicated genotypes were spotted onto rich medium with or without 50 μg/ml CFW for three days at 30°C. The genotype of the control strain is: chs6Δapl2Δ. The chs6Δapl2Δmih1-1 strain was constructed by introducing the mih1-1 lesion de novo in the original chs6Δapl2Δ parent strain. The mih1Δ designation indicates that the wild-type MIH1 locus was deleted in the original parent strain (chs6Δapl2Δmih1Δ). The mih1-1/mih1Δ designation indicates that the mih1-1 allele was deleted in the original mih1-1 mutant strain. (E) Mih1 phosphatase activity is required for CFW resistance conferred by the mih1-1 mutation. Serial dilutions of chs6Δapl2Δmih1Δ cells containing plasmids expressing the mih1-1 allele (‘S162R’), a double mutant protein (S162R C320S) in which a second mutation was introduced that ablates catalytic activity, or empty vector (‘Vec’), were spotted onto complete synthetic medium with or without 50 μg/ml CFW for three days at 30°C. The host strain is the chs6Δapl2Δmih1Δ strain background.
Figure 1—figure supplement 1
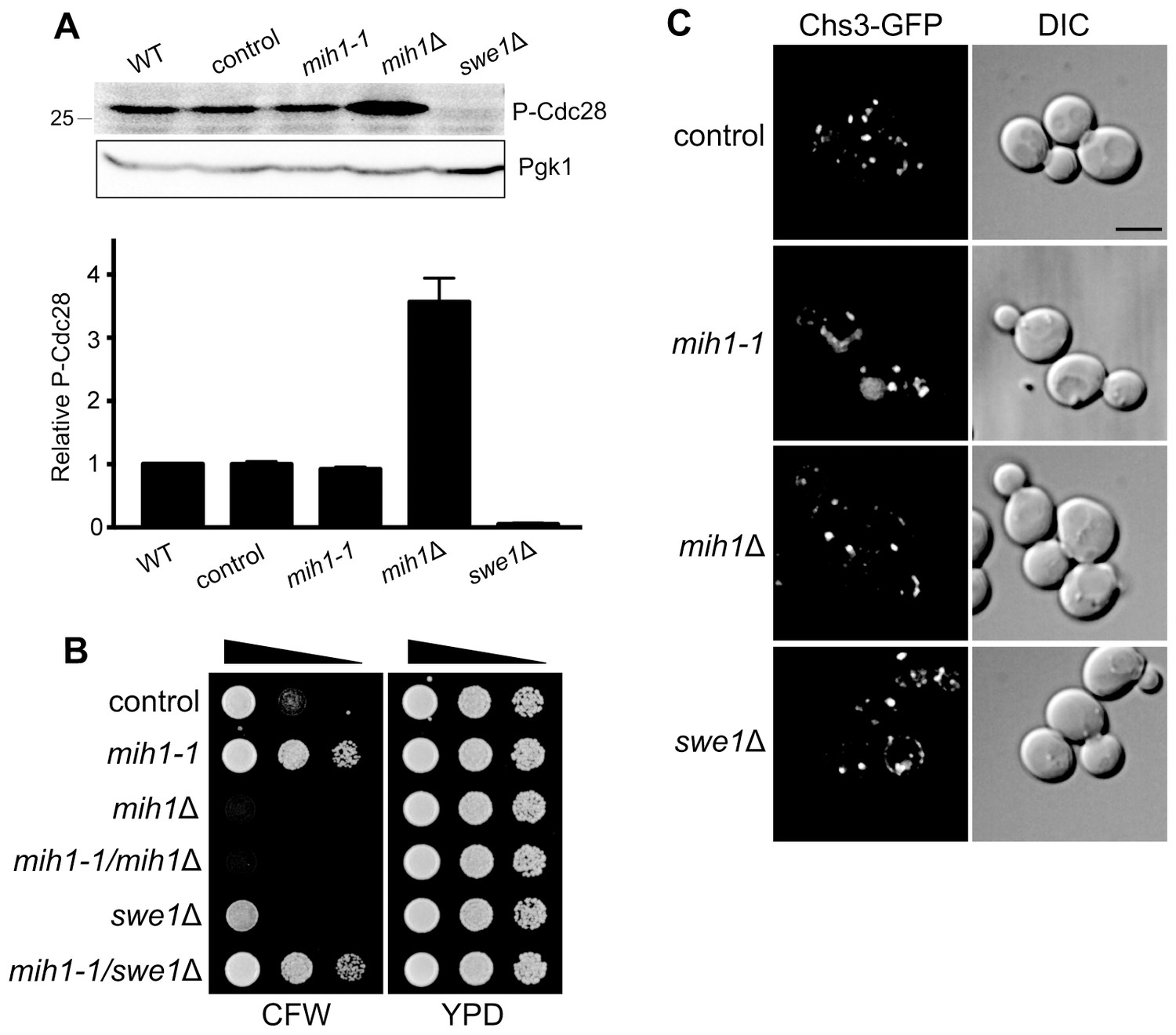
Regulation of membrane trafficking by Mih1 is not dependent on activated Cdc28.
(A) Quantitation of phospho-Cdc28 levels. Cell lysates of the indicated strains were probed by immunoblot with an anti-phospho-cdc2 (Tyr15) antiserum (one example is shown in the inset, top). The intensity of the signal corresponding to Cdc28 (the S. cerevisiae homolog of Cdc2) was normalized for loading. The graph shows the mean intensities (+ s.e.m.) for three independent experiments, relative to the mean intensity measured for wild-type cells. The parent genotype of all strains (except wild-type) is chs6Δapl2Δ; additional mutations are indicated. (B) Deletion of SWE1 does not affect growth in the presence of calcofluor white. Serial dilutions of control (chs6Δapl2Δ), mih1-1 (chs6Δapl2Δmih1-1), mih1Δ (chs6Δapl2Δmih1Δ), mih1-1/mih1Δ (chs6Δapl2Δmih1-1mih1Δ), swe1Δ (chs6Δapl2Δswe1Δ), and mih1-1/swe1Δ (chs6Δapl2Δmih1-1swe1Δ) cells were grown on YPD medium with or without 100 μg/ml CFW for three days at 30°C. This is the uncropped image of Figure 1D. (C) Localization of Chs3-GFP in control (chs6Δapl2Δ), mih1-1 (chs6Δapl2Δmih1-1), mih1Δ (chs6Δapl2Δ mih1Δ) and swe1Δ (chs6Δapl2Δswe1Δ) cells. A representative medial optical plane of a z-series is shown. Scale bar, 5 µm.
Figure 2 with 1 supplement
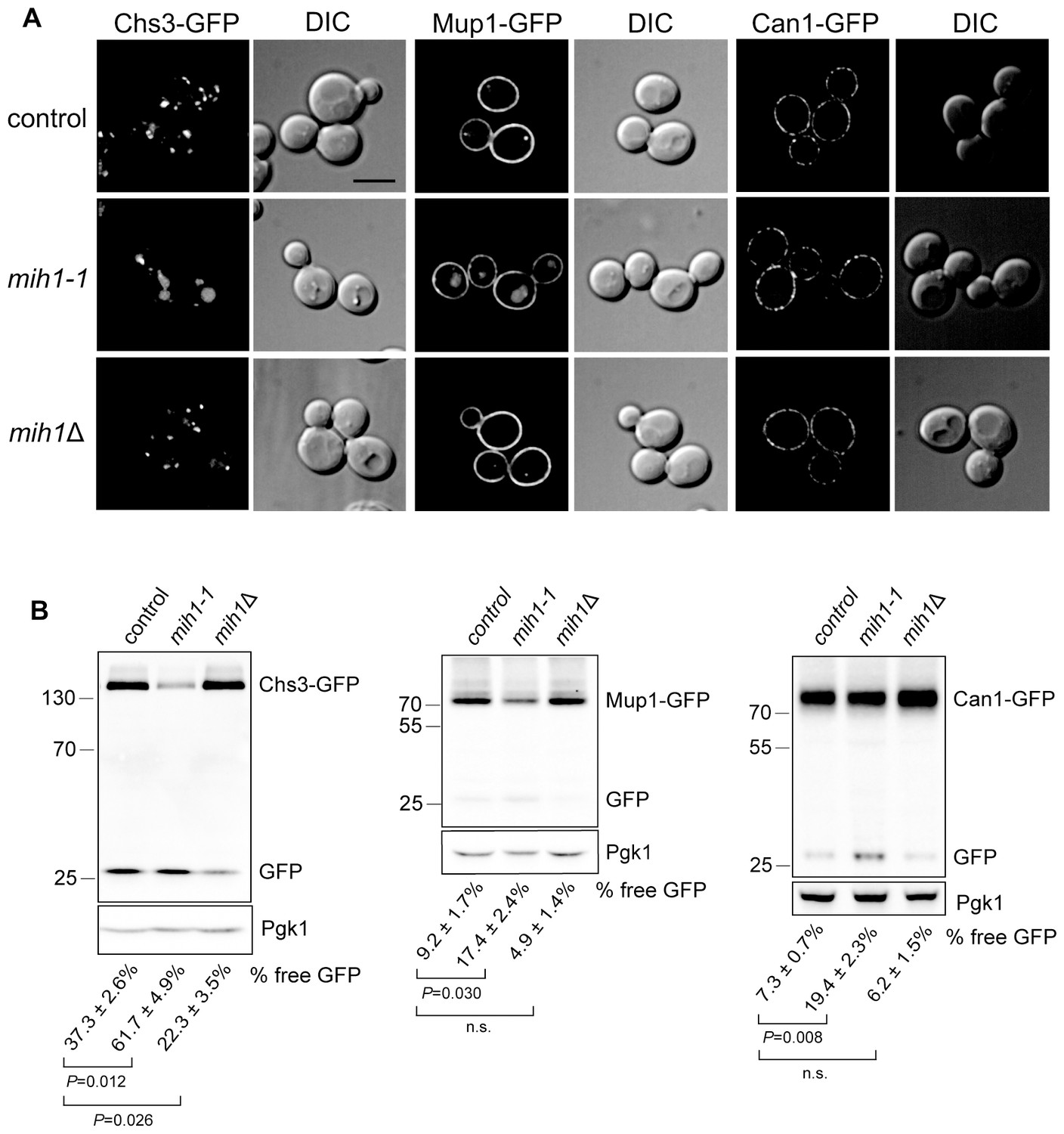
Mih1 regulates turnover of a subset of plasma membrane proteins.
(A) Localization of Chs3-GFP, Mup1-GFP, and Can1-GFP in control cells (chs6Δapl2Δ), mih1-1 (chs6Δapl2Δmih1-1) and mih1Δ (chs6Δapl2Δmih1Δ). A representative medial optical plane of a z-series is shown. Scale bar, 5 µm. (B) Increased proteolysis of Chs3-GFP, Mup1-GFP, and Can1-GFP in mih1-1 (chs6Δapl2Δmih1-1) cells and decreased proteolysis in mih1Δ (chs6Δapl2Δmih1Δ) cells. Mup1-GFP and Can1-GFP expressing cells were grown in methionine- or arginine-deficient medium, respectively, to promote plasma membrane recycling. A representative anti-GFP immunoblot of cell lysates of the indicated strains is shown. The number below each lane indicates the mean proportion (+s.e.m.) of free GFP to the total GFP signal in the lane calculated from a minimum of three independent measurements. Anti 3-phosphoglycerate kinase (Pgk1) blotting was used to control for loading. Statistical significance for pairwise comparisons is indicated; n.s., not statistically significant. The positions of molecular weight (kDa) protein standards is indicated on the left side of the gel.
Figure 2—figure supplement 1
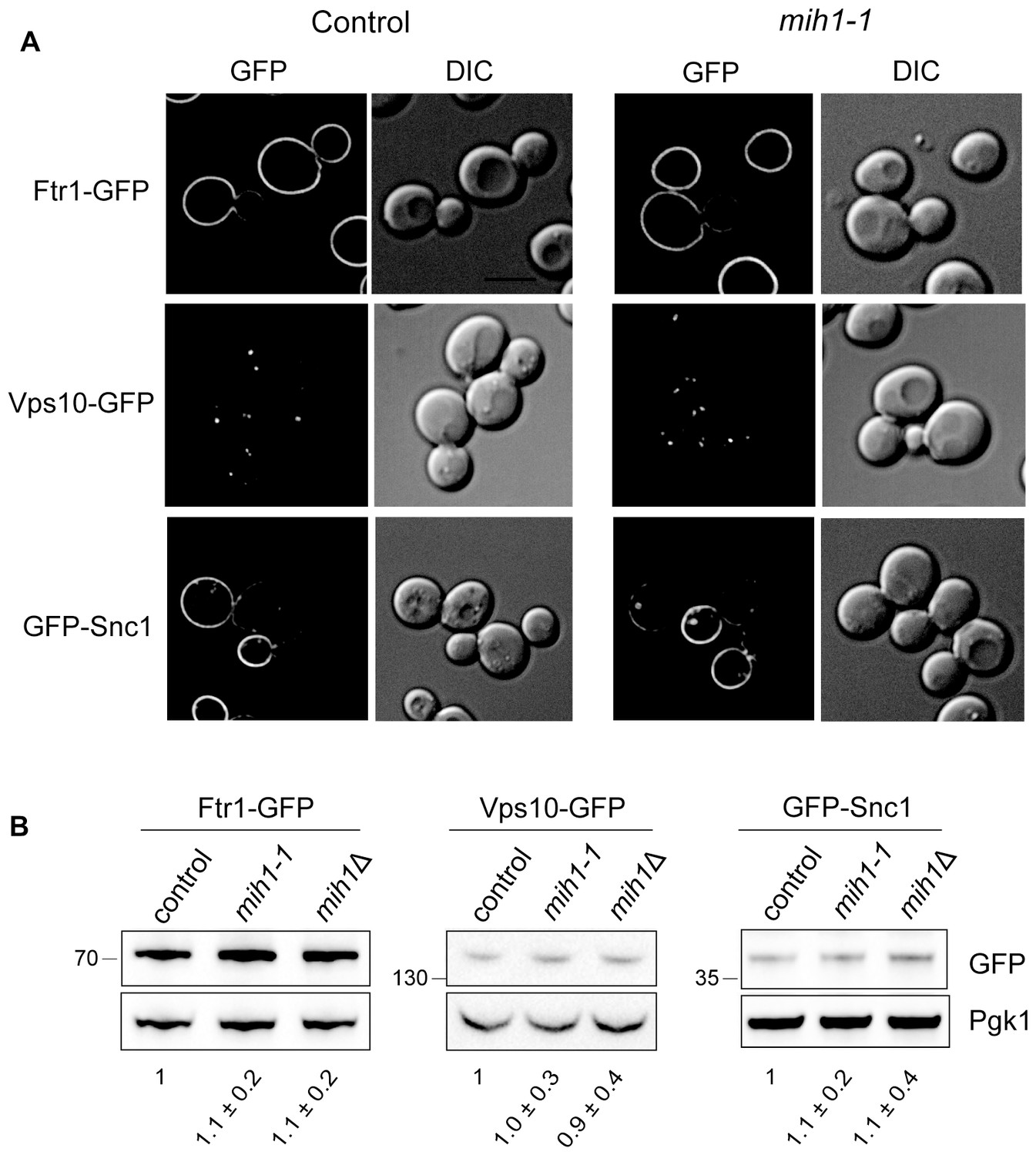
Localization and steady state levels of Ftr1-GFP, Vps10-GFP and GFP-Snc1 in control (chs6Δapl2Δ), mih1-1 (chs6Δapl2Δmih1-1), and mih1Δ (chs6Δapl2ΔmihΔ) cells.
(A) Micrographs of cells expressing the indicated proteins as fusions to GFP are shown. Cells expressing Ftr1-GFP were cultured in iron-deficient medium to promote recycling. A representative medial optical plane of a z-series is shown. Scale bar, 5 µm. (B) Steady state levels of Ftr1-GFP, Vps10-GFP, and Snc1-GFP are unaffected in mih1-1 or mih1Δ cells. Cell lysates of cells expressing the indicated GFP fusion proteins, which all undergo plasma membrane recycling, were probed with anti-GFP antibody and quantified. Ftr1-GFP expressing cells were grown in iron-deficient medium to promote plasma membrane recycling. The means of three independent measurements (+s.e.m.), normalized to the mean of control cells, are indicated below each lane. None of the differences in abundances are statistically significant.
Figure 3
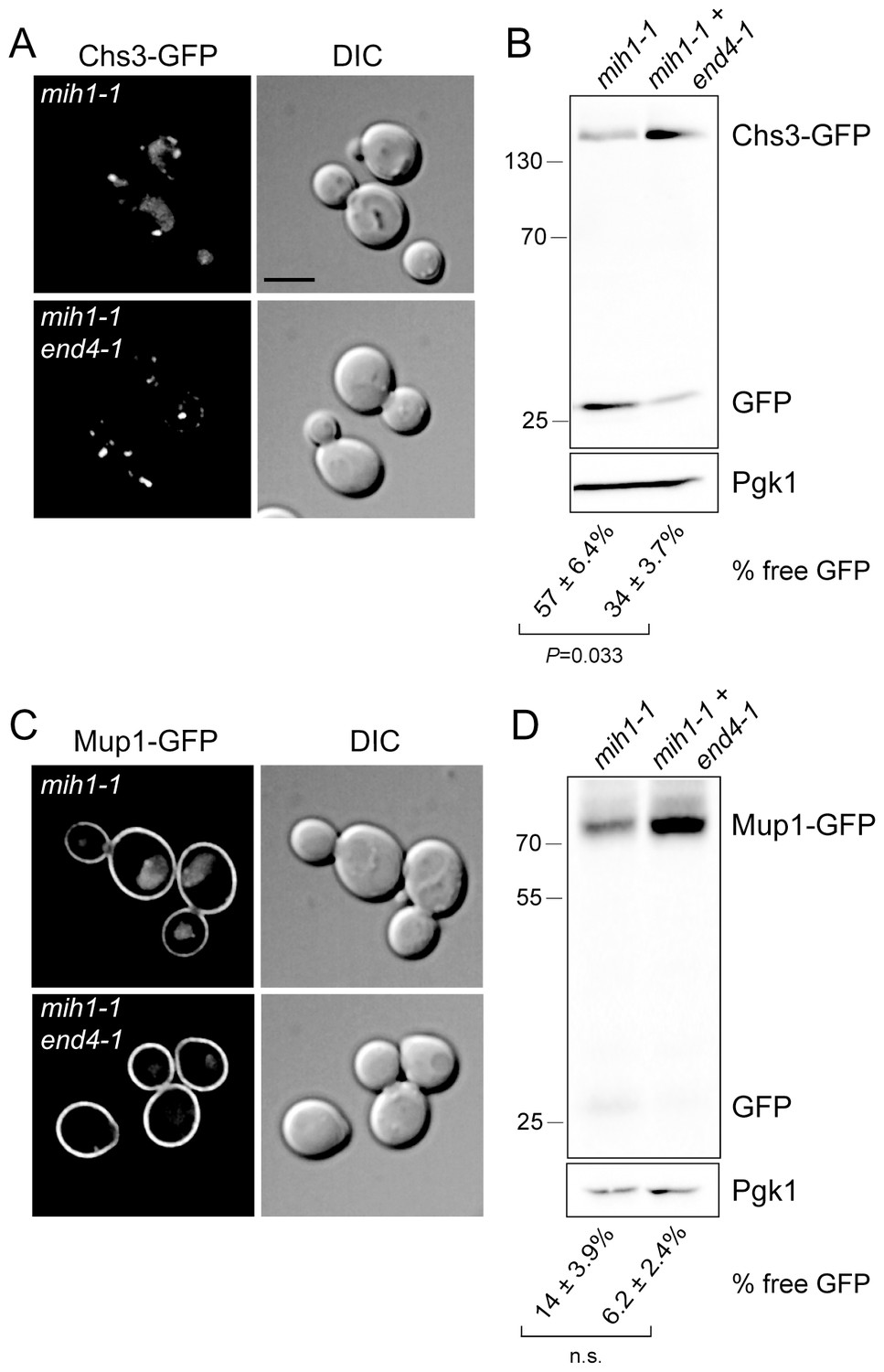
Mih1 regulates protein sorting at the endosome.
(A) Endocytosis promotes vacuolar localization of Chs3-GFP in mih1-1 cells. Micrographs showing Chs3-GFP in mih1-1 (chs6Δapl2Δmih1-1) and mih1-1end4-1 (chs6Δapl2Δmih1-1end4-1) cells are shown. A representative medial optical plane of a z-series is shown. Scale bar, 5 µm. (B) Cleavage of GFP from Chs3-GFP is attenuated by the end4-1 mutation. Anti-GFP blotting of cell lysates was used to quantify the pools of full length Chs3-GFP and free GFP. The number below each lane indicates the mean (+s.e.m.) proportion of free GFP to the total GFP signal in the lane calculated from a minimum of three independent measurements. Statistical significance is indicated. Anti-Pgk1 blotting was used to control for loading. The positions of molecular weight (kDa) of protein standards are indicated on the left side of the gel. (C) Vacuolar localization of Mup1-GFP requires endocytosis. Micrographs showing Mup1-GFP in mih1-1 (chs6Δapl2Δmih1-1) and mih1-1end4-1 (chs6Δapl2Δmih1-1end4-1) cells cultured in medium lacking methionine are shown. A representative medial optical plane of a z-series is shown. Scale bar, 5 µm. (D) Cleavage of GFP from Mup1-GFP is attenuated by end4-1 mutation. The data are presented as described in the legend to panel B. Due to the small amount of free GFP, the difference is not statistically significant (n.s.).
Figure 4

Mih1 regulates the phosphorylation state of Vps26 retromer subunit.
(A) Growth of strains lacking genes encoding endosomal sorting nexins and retromer deletions in the presence of CFW. Serial dilutions of control (chs6Δapl2Δ), mih1-1 (chs6Δapl2Δmih1-1), snx3Δ (chs6Δapl2Δsnx3Δ), snx4Δ (chs6Δapl2Δsnx4Δ), vps5Δ (chs6Δapl2Δvps5Δ), vps17Δ (chs6Δapl2Δvps17Δ), mvp1Δ (chs6Δapl2Δmvp1Δ), vps26Δ (chs6Δapl2Δvps26Δ), vps29Δ (chs6Δapl2Δvps29Δ) and vps35Δ (chs6Δapl2Δvps35Δ) cells were spotted onto rich medium with or without 50 μg/ml CFW for three days at 30°C. (B) Localization of Chs3-GFP in control (chs6Δapl2Δ), mih1-1 (chs6Δapl2Δmih1-1) and vps29Δ (chs6Δapl2Δvps29Δ) (as a representative retromer deletion). A representative medial optical plane of a z-series is shown. Scale bar, 5 µm. (C) Endogenously expressed Vps35-GFP, Vps29-GFP, Vps26-GFP, Vps17-GFP or Vps5-GFP fusion proteins were immunopurified from chs6Δapl2Δmih1Δ cells and samples of the collected material were analyzed by immunoblotting with a pan phosphoryl amino acid antiserum (top) or anti-GFP (bottom). An amount of cell lysate equivalent to 0.5% of the starting material for the purifications was run in the ‘load’ lanes. The positions of molecular weight (kDa) protein standards are indicated on the left side of the gels. (D) Vps26 phosphorylation status. Endogenously expressed Vps26-GFP fusion protein was immunopurified from chs6Δapl2Δ (control), chs6Δapl2Δmih1-1 (mih1-1) and chs6Δapl2Δmih1Δ (mih1Δ) cells and subjected to immunoblot assay with a pan phosphoryl amino acid antiserum (top) or anti-GFP (bottom). The molecular weight (kDa) of protein standards is indicated on the left side of the gels. (E) Purified Mih1 proteins. The indicated Mih1 proteins were purified from yeast cell lysates. A coomassie blue stained gel of aliquots of the eluted fractions used for activity assays is shown. (F) In vitro assay of Vps26 dephosphorylation by Mih1. Purified Mih1 proteins or lambda protein phosphatase were incubated with immunopurified Vps26-GFP. Equivalent portions of each reaction were examined by immunoblot with a pan phosphoryl amino acid antiserum (top) or anti-GFP (bottom). ‘Mock’ indicates a reaction that received no purified phosphatase.
Figure 5
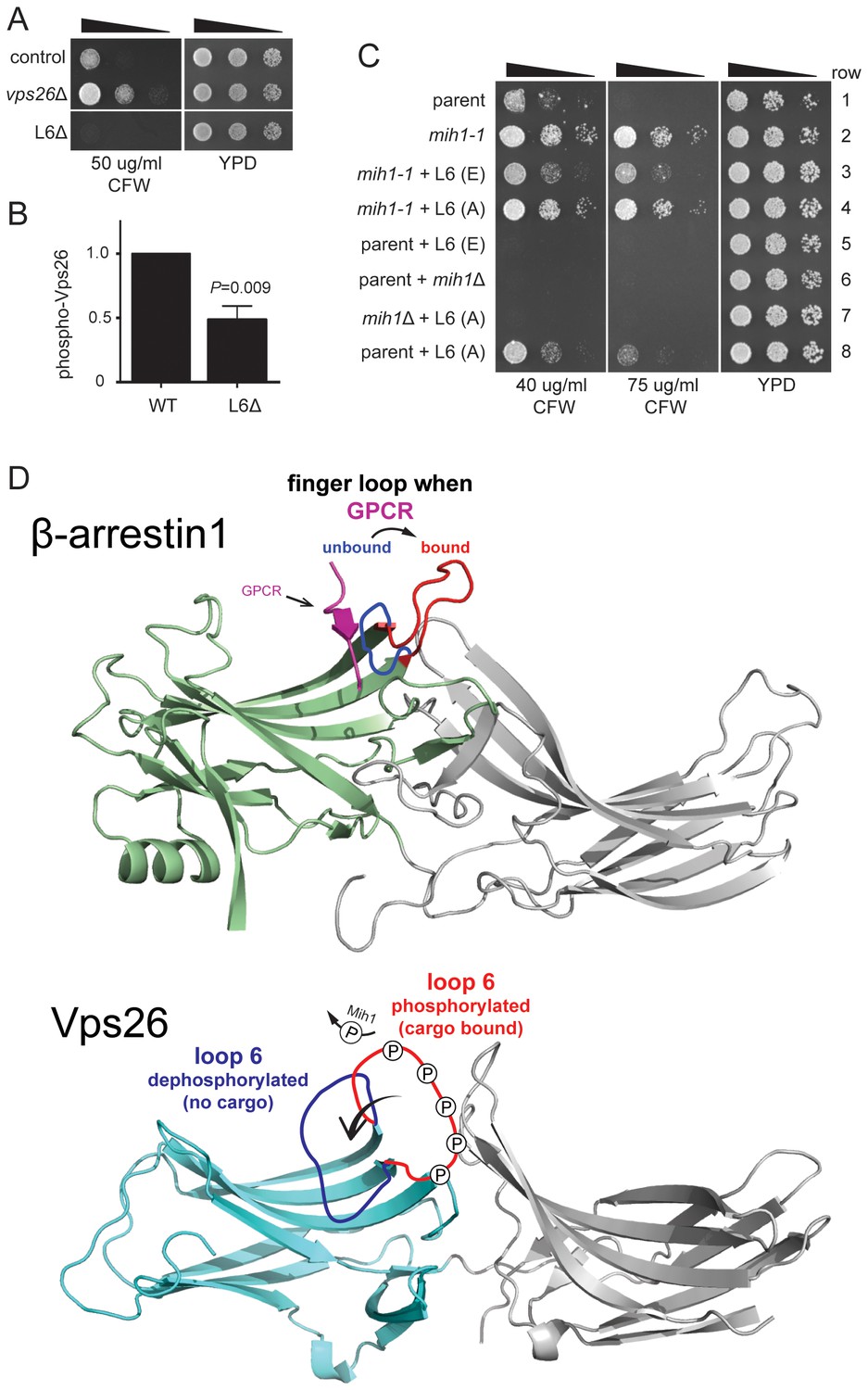
Substitutions within Vps26 loop 6 influence cargo recycling.
(A) Deletion of loop 6 of Vps26 confers CFW sensitive growth. Serial dilutions of control (chs6Δapl2Δ), vps26Δ (chs6Δapl2Δvps26Δ) and L6Δ (chs6Δapl2Δvps26ΔL6) cells were spotted on YPD medium with or without CFW at the indicated concentration for three days at 30°C. (B) Vps26 loop 6 is de-phosphorylated by Mih1 in vivo. The amount of phosphorylated Vps26ΔL6 -GFP relative to native Vps26-GFP is shown. The mean (+s.e.m.) of three independent determinations is plotted. Statistical significance is indicated. (C) Phospho-mimetic substitutions in loop 6 of Vps26 confer sensitivity to CFW. Serial dilutions of control (chs6Δapl2Δ), mih1-1 (chs6Δapl2Δmih1-1), mih1−1 + L6 (E) (chs6Δapl2Δmih1-1vps26-loop 6 (E)), mih1−1 + L6 (A) (chs6Δapl2Δmih1-1vps26-loop 6 (A)), L6 (E) (chs6Δapl2Δvps26-loop 6 (E)), mih1Δ (chs6Δapl2Δmih1Δ), mih1Δ + L6 (A) (chs6Δapl2Δmih1Δvps26-loop 6 (A)) and L6 (A) (chs6Δapl2Δvps26-loop 6 (A)) cells were spotted on YPD medium with or without CFW at the indicated concentration for three days at 30°C. (D) Top: Structure of human β-arrrestin1 bound to a peptide derived from the sequence of the V2 vasopressin receptor (Shukla et al., 2013) (PDB: 4JQI). The ‘finger loop’, which corresponds to loop 6 of Vps26, is shown in the inactive (i.e., GPCR unbound) and active (GPCR bound) conformations to illustrate the manner in which it occludes GPCR binding in the inactive state. Bottom: Structure of human Vps26a (Shi et al., 2006) (PDB: 2FAU) with the extended loop 6 of yeast modeled in the dephosphorylated and phosphorylated conformations. We suggest that phosphorylation relieves an auto-inhibited conformation of the loop that antagonizes cargo recognition and that dephosphorylation by Mih1 restores the auto-inhibited state.
Figure 6
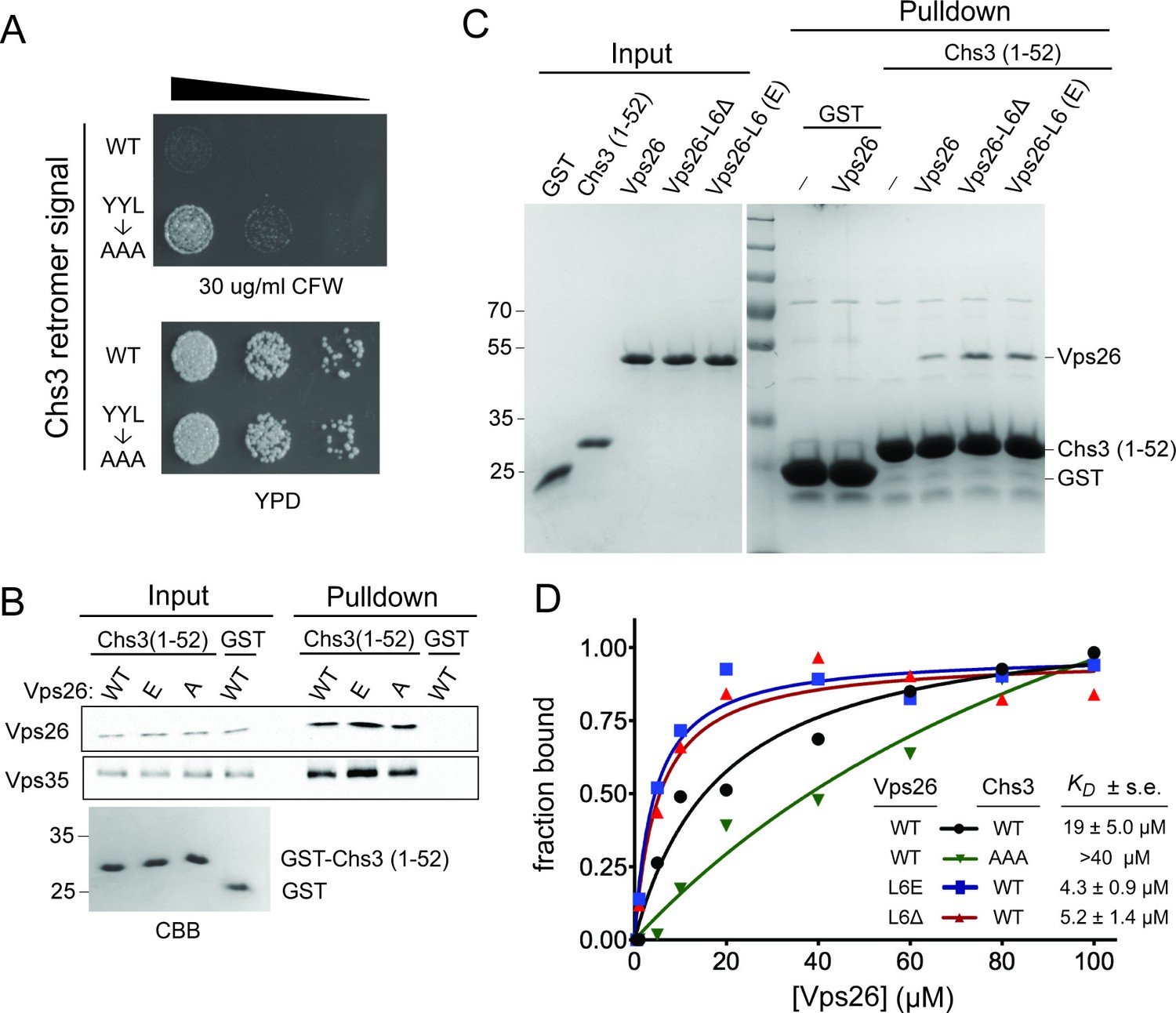
Vps26 loop 6 controls affinity for a retromer sorting signal.
(A) Substitution of a putative retromer sorting motif in Chs3 confers resistance CFW. Chs3 residues Y12YLwere changed to alanine in chs6Δapl2Δ cells. Serial dilutions of control (chs6Δapl2Δ) and mutant cells were spotted on calcofluor plates (30 μg/ml) and grown for three days at 30°C. (B) The Vps26 and Vps35 retromer subunits are captured by beads presenting Chs3 residues 1–52. Lysates from cells expressing GFP tagged Vps26 (or its variants) or Vps35-FLAG were incubated with immobilized GST-Chs3 (amino acids 1–52; ‘GST-1–52’) or GST. An amount of cell lysate equivalent to 0.5% of the starting material for the purifications was run in the ‘load’ lanes. The relative enrichment of each protein in the pulldown fraction (means and standard deviations of three independent experiments) is indicated. A coomassie blue-stained gel of 3% of each starting affinity resins is shown below the immunoblots. The molecular weight (kDa) of protein standards is indicated on the left side of the gels. (C) Vps26 binds directly to the YYL Chs3 sorting signal. Binding assays of pure, 6xHis-tagged wild-type Vps26, Vps26 loop 6 deletion (Vps26ΔL6) and loop 6 phosphomimetic Vps26 (Vps26-L6(E) to GST-Chs3(1-52) or the same portion of Chs3 with alanines substituted for the YYL sequence (GST-Chs3(YYL->AAA)) are shown. The input fractions show one microgram of the starting material used for each assay. The positions of molecular weight (kDa) protein standards are indicated on the left side of the gel. (D) Loop 6 controls affinity of Vps26 for a retromer sorting signal. Equilibrium binding measurements of pure, recombinant wild-type Vps26, Vps26L6Δ, and Vps26-L6(E) for the GST-Chs3(1-52) fusion protein, and wild-type Vps26 for the GST-Chs3(YYL->AAA) fusion protein, are shown. The points plotted in the graphs represent the mean fraction of bound Vps26 proteins, calculated from single measurements using three independent protein preparations of each Vps26 protein. The calculated KD values (±s.e.) are listed. Saturation binding to the GST-Chs3(YYL->AAA) fusion protein was not observed, so a dissociation constant could not be calculated.
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
A CDC25 family protein phosphatase gates cargo recognition by the Vps26 retromer subunit
eLife 6:e24126.
https://doi.org/10.7554/eLife.24126




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}