Doc2B acts as a calcium sensor for vesicle priming requiring synaptotagmin-1, Munc13-2 and SNAREs
- University of Copenhagen, Denmark
- VU Medical Center, Netherlands
- AstraZeneca R&D, United Kingdom
- VrijeUniversiteit, Netherlands
Figures
Figure 1 with 2 supplements
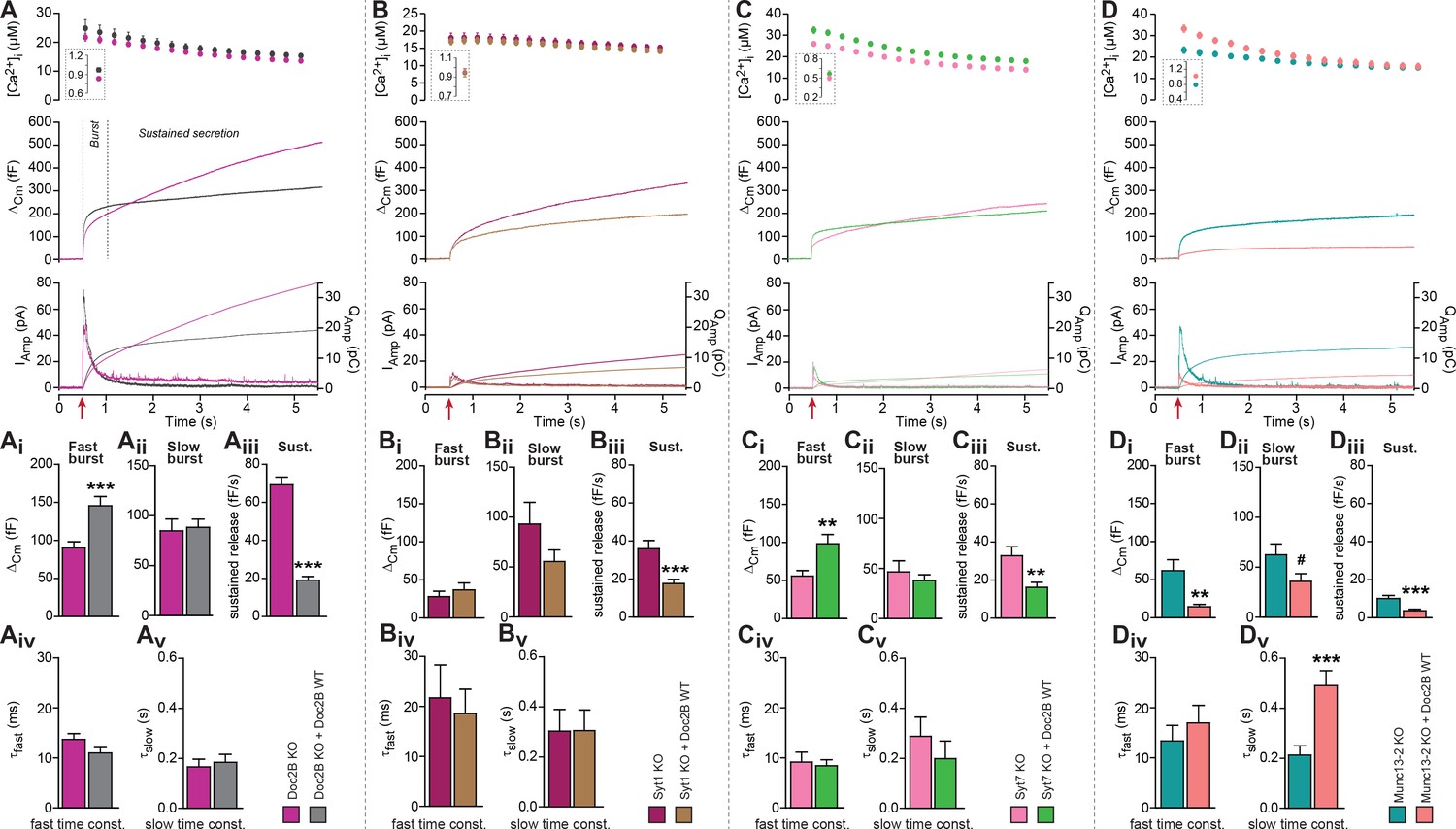
Doc2B requires synaptotagmin-1 and Munc13-2 to increase the Readily Releasable Pool (RRP) size.
(A) Expression of Doc2B in Doc2B knockout (KO) mouse adrenal chromaffin cells. Top panels: Intracellular [Ca2+] (mean ±SEM) after and (insert) shortly before Ca2+ uncaging (uncaging light for 1–2 ms at red arrow). Middle panel: average capacitance traces from Ca2+ uncaging experiments in Doc2B KO cells (magenta traces, n = 40 cells) and after overexpression of Doc2B WT in Doc2B KO cells (grey traces, n = 36 cells). Bottom panel: amperometric measurements (mean traces) from the same cells. Traces showing a release transient after the uncaging stimulus are the amperometric currents (left ordinate axes). Monotonically increasing traces display the time-integral of the amperometric current (i.e. the charge), scaled to the right ordinate axis. Note that the time-integrals display the same general shape as the capacitance traces, as expected. (Ai) The amplitude (mean ±SEM) of the fast burst was increased by Doc2B (***p<0.001, Mann-Whitney test), whereas (Aii) the amplitude of the slow burst component remained constant. Fast and slow burst correspond to the sizes of the RRP and the SRP, respectively. (Aiii) The rate of the near-linear sustained component (D) was strongly reduced by Doc2B (***p<0.001, Mann-Whitney test). (Aiv) The fusion time constant for the fast burst, and (Av) fusion time constant for the slow burst were unaffected by Doc2B. (B) Ca2+ uncaging experiment in syt-1 KO cells (n = 18) and syt-1 KO cells overexpressing Doc2B WT (n = 20). Panels (B–Bv) are arranged as in A. Doc2B expression led to a highly significant reduction in sustained release, but no significant changes in the fast or slow burst sizes. ***p<0.001, Mann-Whitney test. (C) Ca2+ uncaging experiment in syt-7 KO cells (n = 17) and syt-7 KO cells overexpressing Doc2B WT (n = 17). Panels (C–Cv) are arranged as in A. Doc2B expression led to a significant increase in fast burst size (**p<0.01, Mann Whitney test), and a reduction in sustained release (**p<0.01, unpaired, two-tailed Student’s t test), as in control experiments (panel A). (D) Ca2+ uncaging experiment in Munc13-2 KO cells (n = 16) and Munc13-2 KO cells overexpressing Doc2B WT (n = 18). Panels (D–Dv) are arranged as in A. All components of release were reduced by overexpression of Doc2B WT in the absence of Munc13-2 (**p<0.01, ***p<0.001, Mann Whitney test; # p=0.0575, unpaired two-tailed Student’s t-test). The time constant of the slow component was increased (***p<0.001, unpaired two-tailed Student's t-test).
Figure 1—figure supplement 1
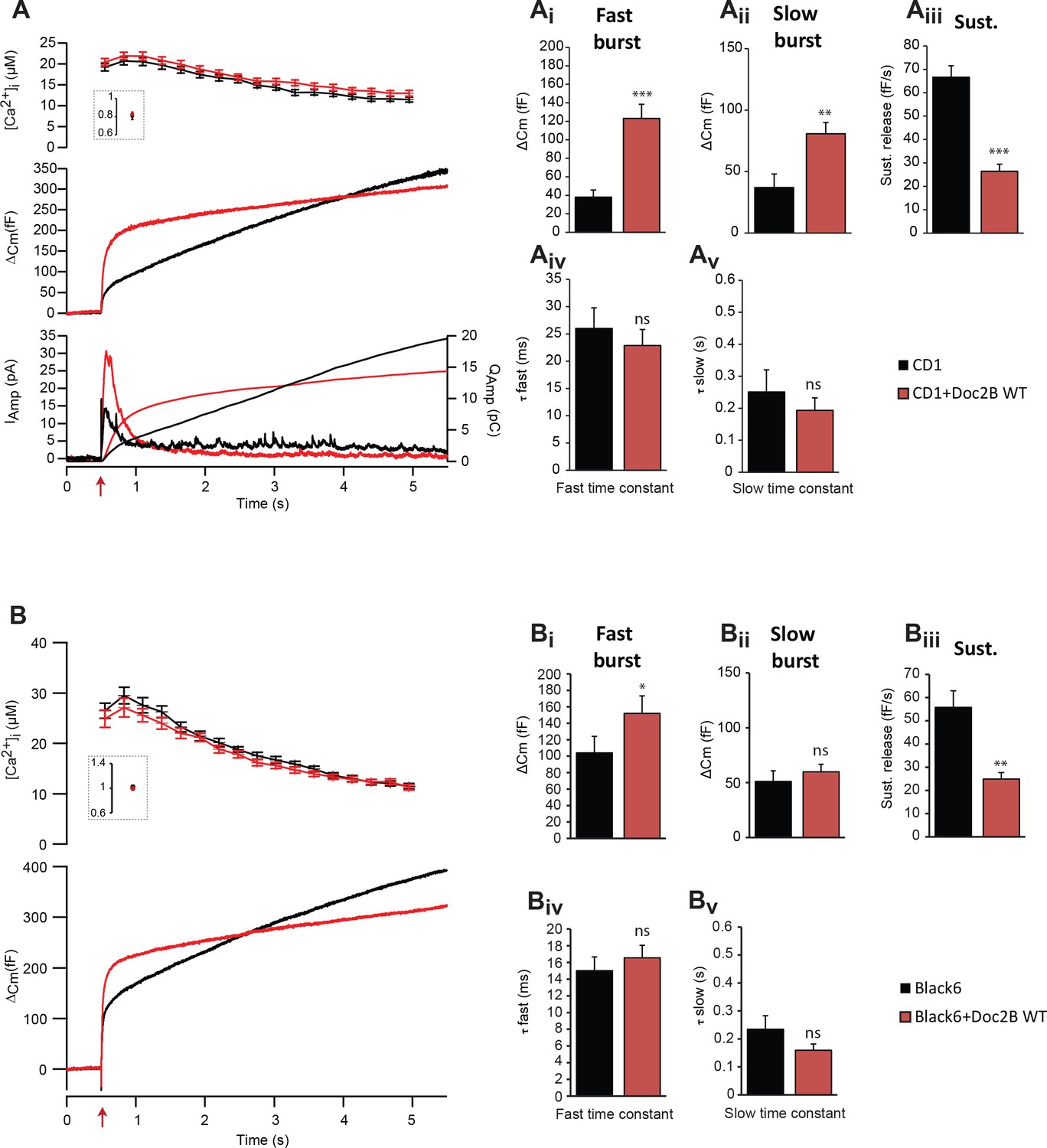
Overexpression of Doc2B in wild-type cells increases the burst size and decreases the sustained release.
(A) Calcium uncaging experiments in CD1 mouse adrenal chromaffin cells (n = 20 cells) and CD1 cells overexpressing Doc2B (n = 20 cells). Top panel: intracellular [Ca2+] (mean ±SEM) obtained by microfluorimetry just after and (insert) before calcium uncaging (uncaging light for 1–2 ms at red arrow). Middle panel: Average capacitance traces of CD1 adrenal chromaffin cells (black trace) and CD1 cells overexpressing Doc2B WT (red trace). Bottom panel: amperometric measurements (average traces) from the same cells. The left ordinate axis represent the amperometric current, the right axis represent amperometric charged. (Ai) The amplitude of the fast burst (mean ±SEM) and the slow burst (Aii) were increased by overexpression Doc2B WT in CD1 cells (Mann-Whitney test, **p<0.01; ***p<0.001). The rate of the sustained component (Aiii) was strongly reduced by overexpressing Doc2B WT in CD1 cells (Mann-Whitney test, ***p<0.001). The fusion time constant of the fast (Aiv) and slow (Av) burst of release were unaffected. (B) Calcium uncaging experiments in Black6 mouse adrenal chromaffin cells (n = 25 cells) and Black6 cells overexpressing Doc2B (n = 26 cells). (Bi) The amplitude of the fast burst (mean ±SEM) was increased by overexpression of Doc2B WT in Black6 cells (Mann-Whitney test, *p<0.05) while the slow burst (Bii) was unchanged. In Black6 mouse chromaffin cells, the sustained rate was also dramatically reduced by overexpressing Doc2B WT (Mann-Whitney test, **p<0.01) and the fast (Biv) and slow (Bv) burst time constants were unaffected.
Figure 1—figure supplement 2
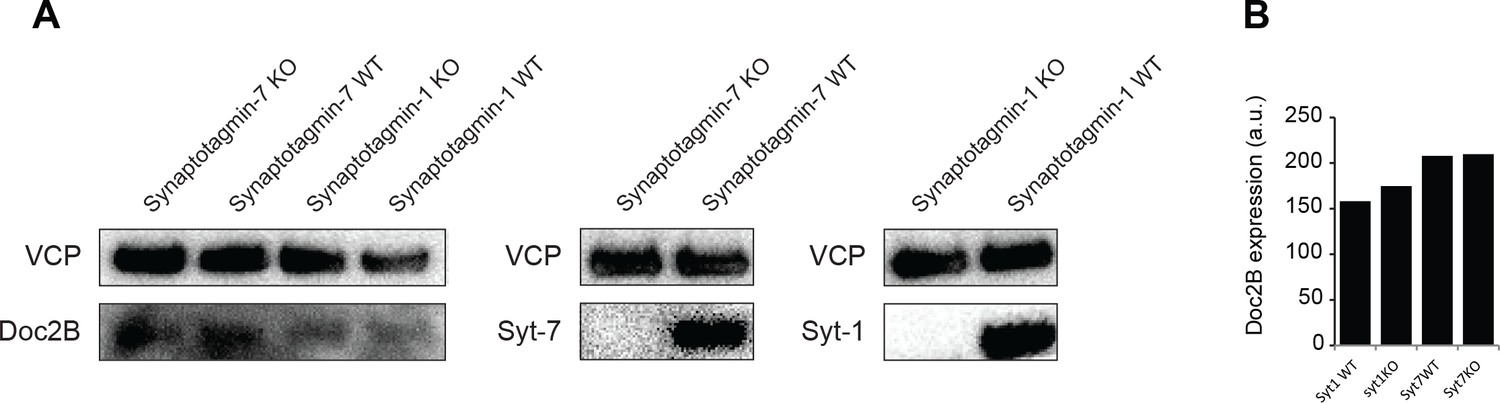
No difference of expression of Doc2B in Syt-1 and Syt-7 WT vs KO mouse adrenal glands.
(A) Western blot analysis of Doc2B expression in Syt-1 and Syt-7 (WT and KO) adrenal glands. Syt-1 and Syt-7 antibodies were used as a control for the genotype. VCP antibody was used as a loading control. (B) Quantification of Doc2B signal in Syt-1 and Syt-7 mouse adrenal glands showing no difference of Doc2b expression. (n = 1 animal/condition, no statistical test).
Figure 2
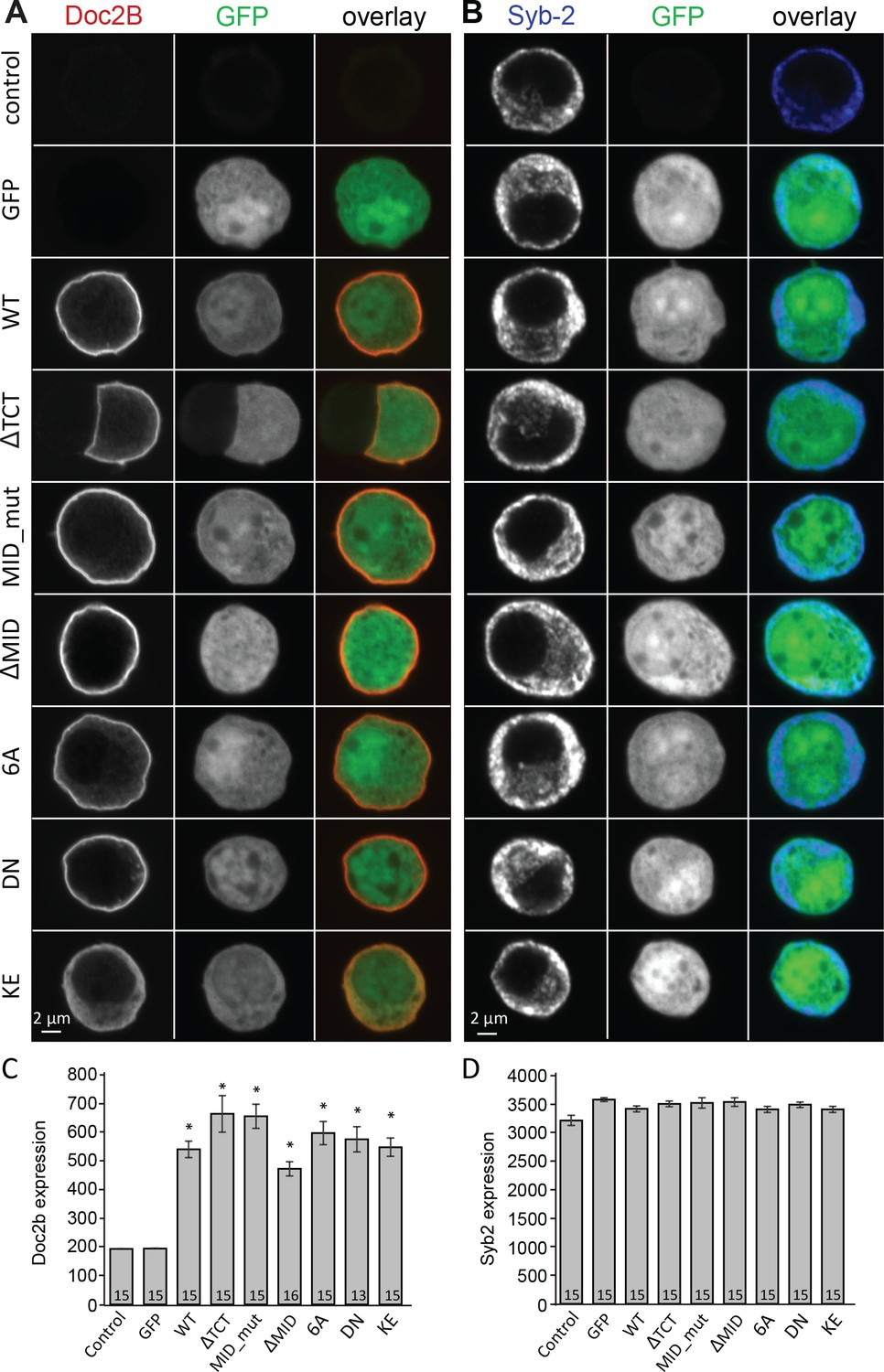
Overexpression of Doc2B in Doc2A/B deficient chromaffin cells transfected with semliki vectors.
(A–B) Typical chromaffin cells stained for (A) Doc2B or (B) VAMP2/synaptobrevin-2 to mark chromaffin granules. EGFP was co-expressed from an internal ribosome entry site (IRES) to mark semliki infection. ‘Control’ indicates non-infected cells. WT: wildtype Doc2B; ΔTCT: deletion of aa. 2–9 to block Tctex-1 binding; MID_mut: substitution of aa. 15–20 (QEHMAI to YKDWAF) to block Munc13 interaction; ΔMID: deletion of aa. 14–41 to block Munc13 interaction; 6A: D163,218,220,303,357,359A; DN: D218,220N, KE: K237,319E. (C–D) Average staining intensity ±sem for (C) Doc2B and (D) VAMP2/synaptobrevin-2 from the indicated number of cells. *p<0.001 (t-test for independent samples with Bonferroni correction).
Figure 3
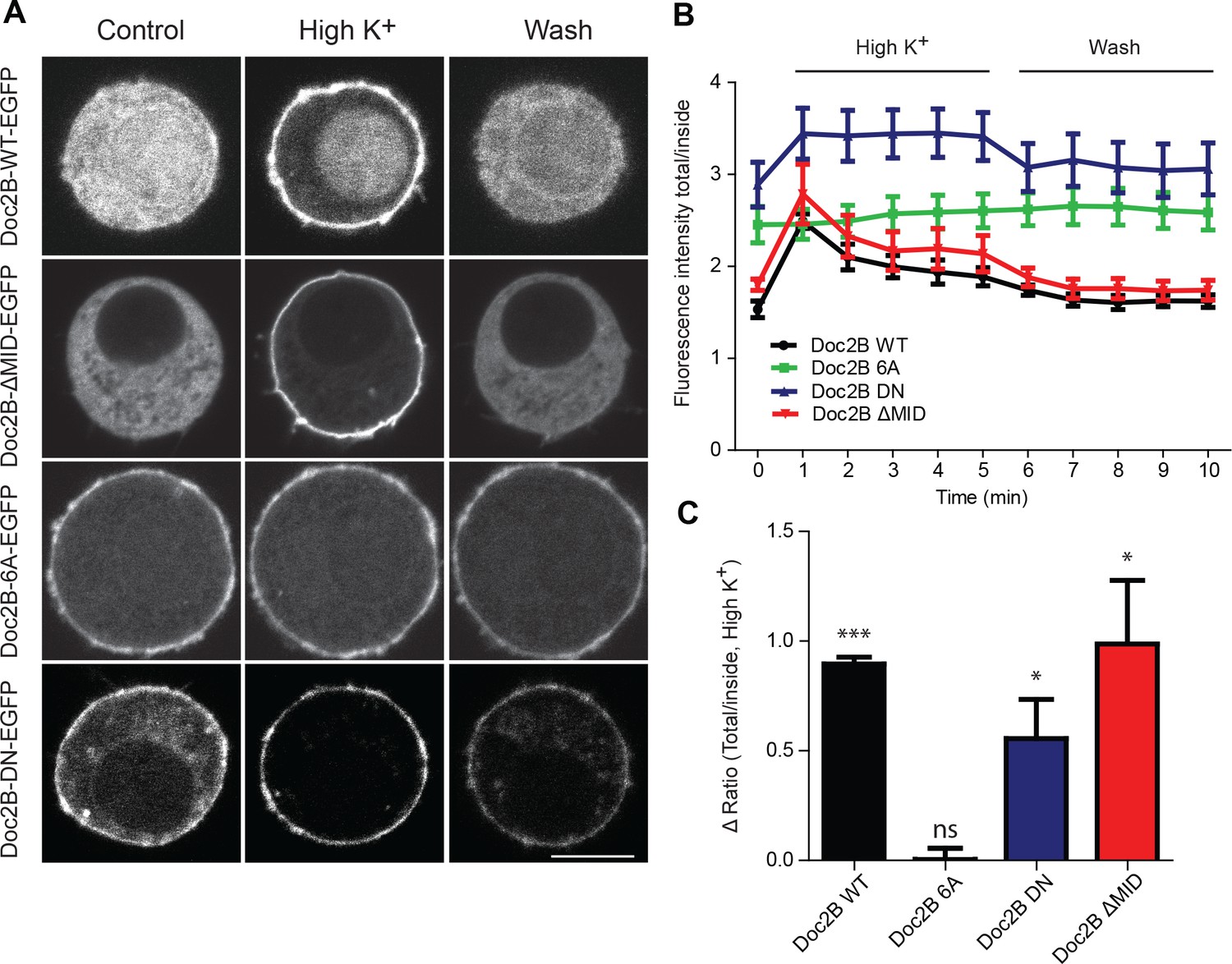
Doc2B translocates to the plasma membrane in a calcium-dependent manner.
(A) Live confocal imaging experiments in CD1 mouse adrenal chromaffin cells expressing Doc2B WT, Doc2B ΔMID, Doc2B 6A or Doc2B DN fused to EGFP. Cells depolarization with High K+ solution (59 mM) triggers the recruitment of Doc2B WT, Doc2b ΔMID and in a lesser extent Doc2B DN to the plasma membrane. Mutations of the calcium binding residues of both C2 domains of Doc2B (Doc2B 6A) led to a permanent localization at the plasma membrane. Washing with extracellular solution (Wash) permits Doc2B WT and ΔMID to re-localize in the cytosol. Scale bar = 5 µm (B) Quantification of Doc2B intensity (ratio total intensity/cytosol intensity) during live imaging experiment. Cells were imaged at resting and after 1 to 5 min stimulation (high K+) every minute, and then washed with extracellular solution and imaged every minute for 5 min. (C) Quantification of the plasma membrane recruitment of doc2B WT and mutants after 1 min stimulation with high K+ solution (Difference in ratio [total fluorescence intensity/cytosol fluorescence intensity] at 1 min compared to resting) showing that only Doc2b 6A does not traffic (n cells: Doc2B WT: 4; Doc2B 6A: 5; Doc2b DN: 11; Doc2B ΔMID: 8. Panel C: One-sample t-test comparing the difference in fluorescence ratio to zero; * p-value<0,05; *** p-value<0.001).
Figure 4
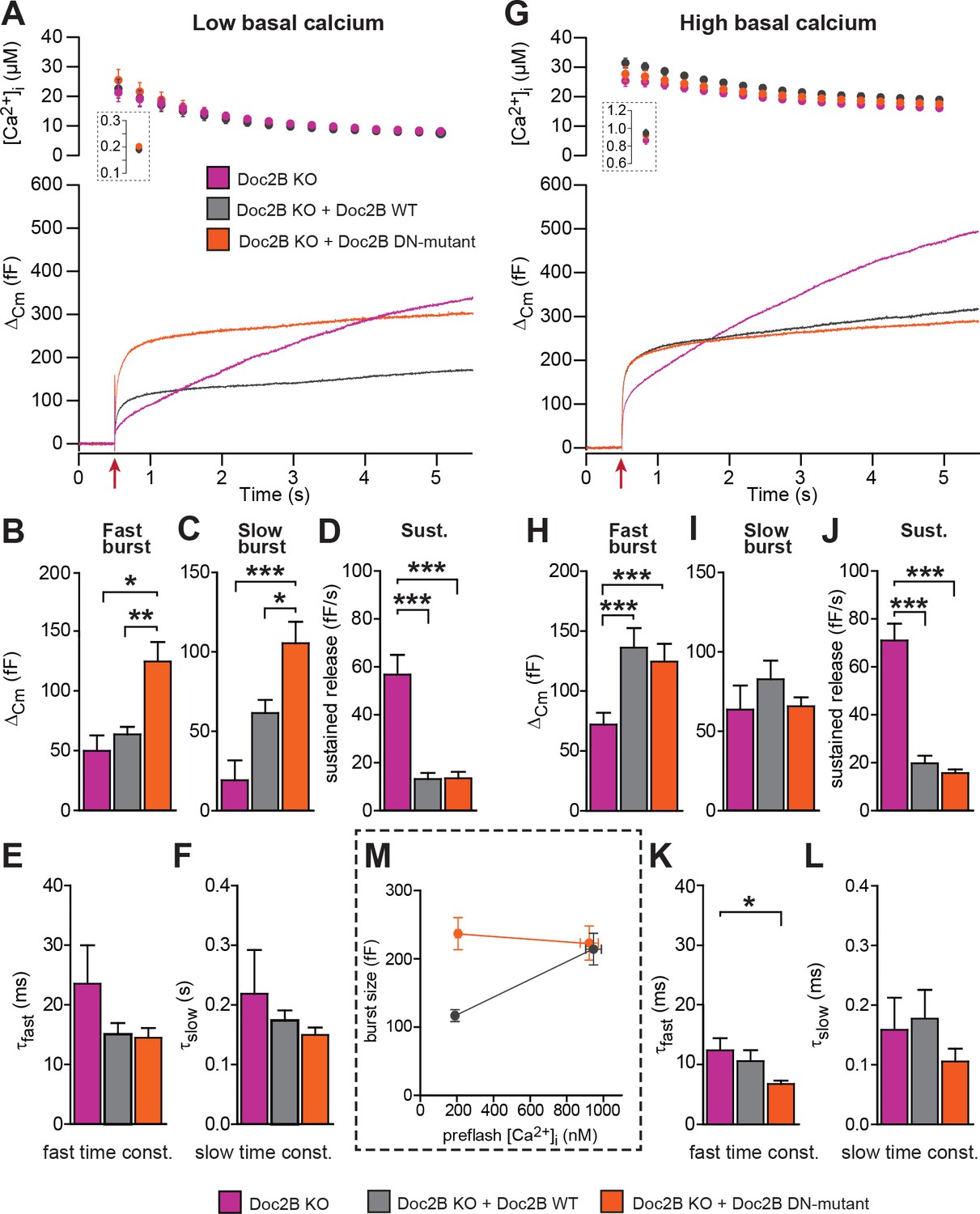
The Doc2B DN-mutant (D218,220N) supports maximal vesicle priming independently of the Ca2+ level.
(A-F) at low basal (preflash) calcium, the DN-mutant supports more secretion than WT Doc2B. (A) Ca2+ uncaging experiments in Doc2B KO cells (n = 12), Doc2B KO cells overexpressing Doc2B WT (n = 19) and Doc2B KO cells overexpressing Doc2b DN (n = 20 cells). Top panel shows the average (±SEM) post-flash [Ca2+]i measured by microfluorimetry; inset shows the pre-flash [Ca2+]i. Bottom panel shows the changes in cell membrane capacitance (mean traces for all cells). (B–F) Kinetic analysis revealed that both the fast and slow bursts were larger upon overexpression of Doc2B DN than of Doc2B WT, leading to a gain of function phenotype at low basal cell calcium. The sustained component was similarly decreased by overexpression of wildtype and DN-mutated Doc2B (*p<0.05, **p<0.01, ***p<0.001, Dunn’s multiple comparison test). (G), similar experiment to (A) but in high basal pre-flash Ca2+ (Doc2B KO, n = 20 cells; Doc2B KO expressing Doc2B WT, 20 cells; Doc2B KO expressing Doc2B DN, 32 cells). (H–L) Kinetic analysis shows that overexpression of either Doc2B DN or Doc2B WT causes a similar increase in the fast burst. Similar to the low preflash [Ca2+]i experiment, the sustained component was decreased to the same levels by overexpression of both proteins. (K) Overexpression of Doc2B DN caused a significant acceleration of the fast burst fusion kinetics. (M) Plot of the total burst size (fast + slow) as a function of the basal cell calcium reveals that Doc2B DN supports maximal LDCV priming irrespective of the cell Ca2+. *p<0.05, ***p<0.001; Dunn’s multiple comparison test.
Figure 5
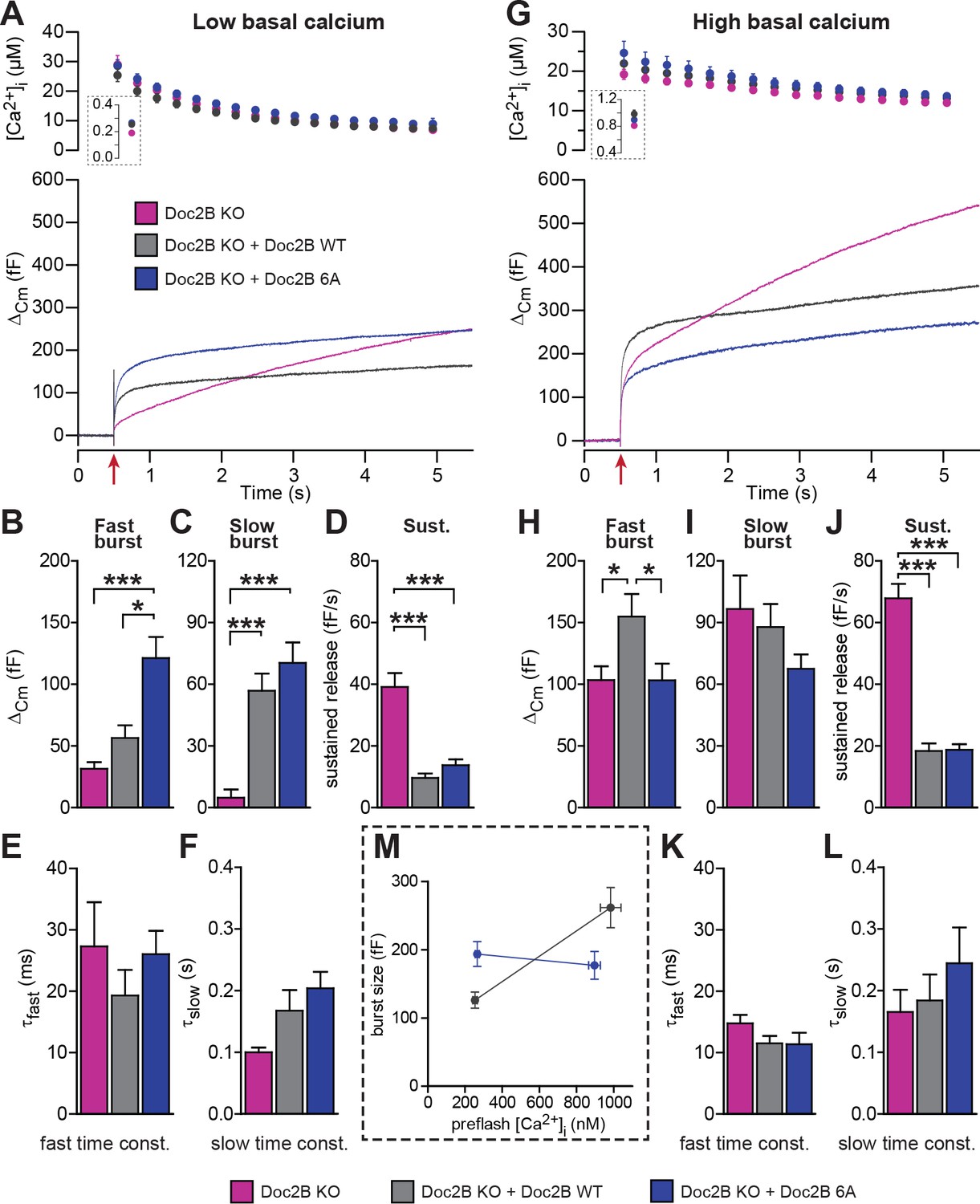
Mutation of the Ca2+binding sites of Doc2B leads to permanent membrane localization and Ca2+-independent priming.
(A-F) When stimulated from low basal calcium, the Ca2+-binding mutant Doc2B 6A supports more secretion that Doc2B WT. (A) Ca2+ uncaging in Doc2b KO cells (n = 23), KO cells overexpressing Doc2B WT (n = 19) and KO cells overexpressing Doc2B 6A (n = 18). Panels are arranged as in Figure 4. (B–F) Kinetic analysis revealed that the fast burst was increased more by overexpression of Doc2B 6A than by Doc2B WT, showing a gain of function of the 6A mutant at low basal cell calcium. The sustained component was similarly decreased by overexpression of either protein. *p<0.05, ***p<0.001; Dunn’s multiple comparison test. (G) similar experiment to (A) but stimulated from high basal pre-flash [Ca2+]i (Doc2B KO, n = 25 cells; Doc2B WT, n = 21 cells; Doc2B 6A, n = 22 cells). (H–L) Kinetic analysis shows that overexpression of Doc2B WT caused a marked increase in the fast burst while Doc2B 6A did not, demonstrating a loss of function when studied from a high basal [Ca2+]. Similar to the low [Ca2+]i experiment, the sustained component was decreased to the same levels by overexpression of both proteins. *p<0.05, Tukey’s multiple comparison test; ***p<0.001, Dunn’s multiple comparison test. (M) Plot of the total burst size (fast + slow) as a function of the basal cell calcium reveals the Ca2+-independent priming reaction in the presence of the 6A mutant.
Figure 6
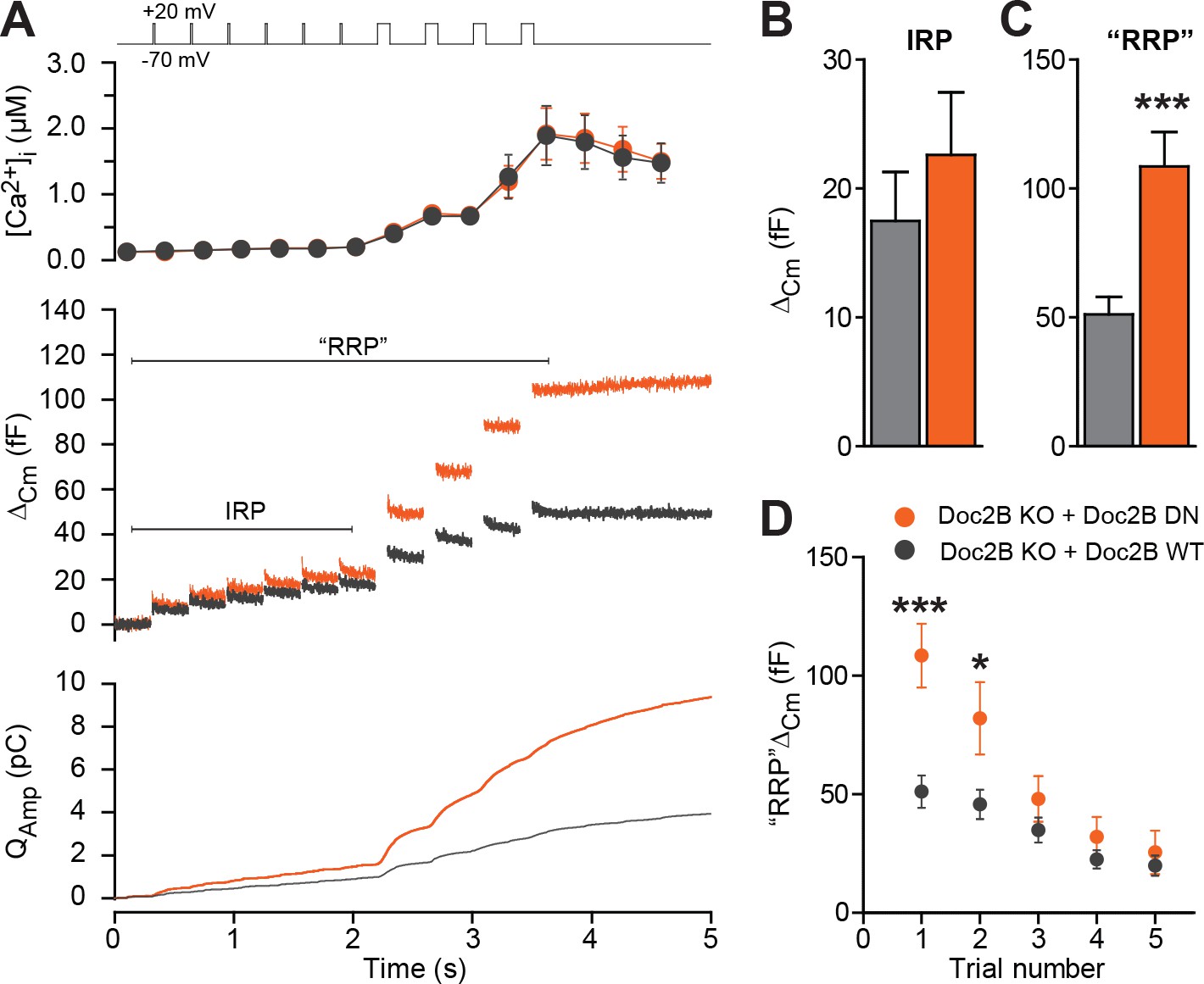
The Doc2B DN-mutant promotes the filling of the Readily Releasable vesicle Pool (RRP), but not the Immediately Releasable Pool (IRP).
(A) A depolarization protocol, consisting of 6 brief (10 ms) and four long (100 ms) depolarizations from −70 mV to +20 mV, was repeated a total of 5 times (trials 1–5), separated by 60 s recovery periods. Only the first trial is depicted. A pipette solution with no added calcium and containing 0.5 mM EGTA was used (n = 21 cells for Doc2B WT; n = 22 cells for Doc2B DN). Top panel shows average (±SEM) [Ca2+]i. Middle panel shows the changes in cell membrane capacitance and bottom panel shows the integrated amperometric current (mean traces for all cells). (B) The size of the Immediately Releasable Pool (IRP) was not significantly affected by overexpression of Doc2B DN while (C) the size of the RRP was significantly enhanced by the Doc2B DN mutant, compared to Doc2B WT. (D), Plot of the mean (±SEM) RRP size over the five trials - the responses decrease because of secretory run-down. *p<0.05, ***p<0.001; unpaired, two-tailed Student’s t-test.
Figure 7
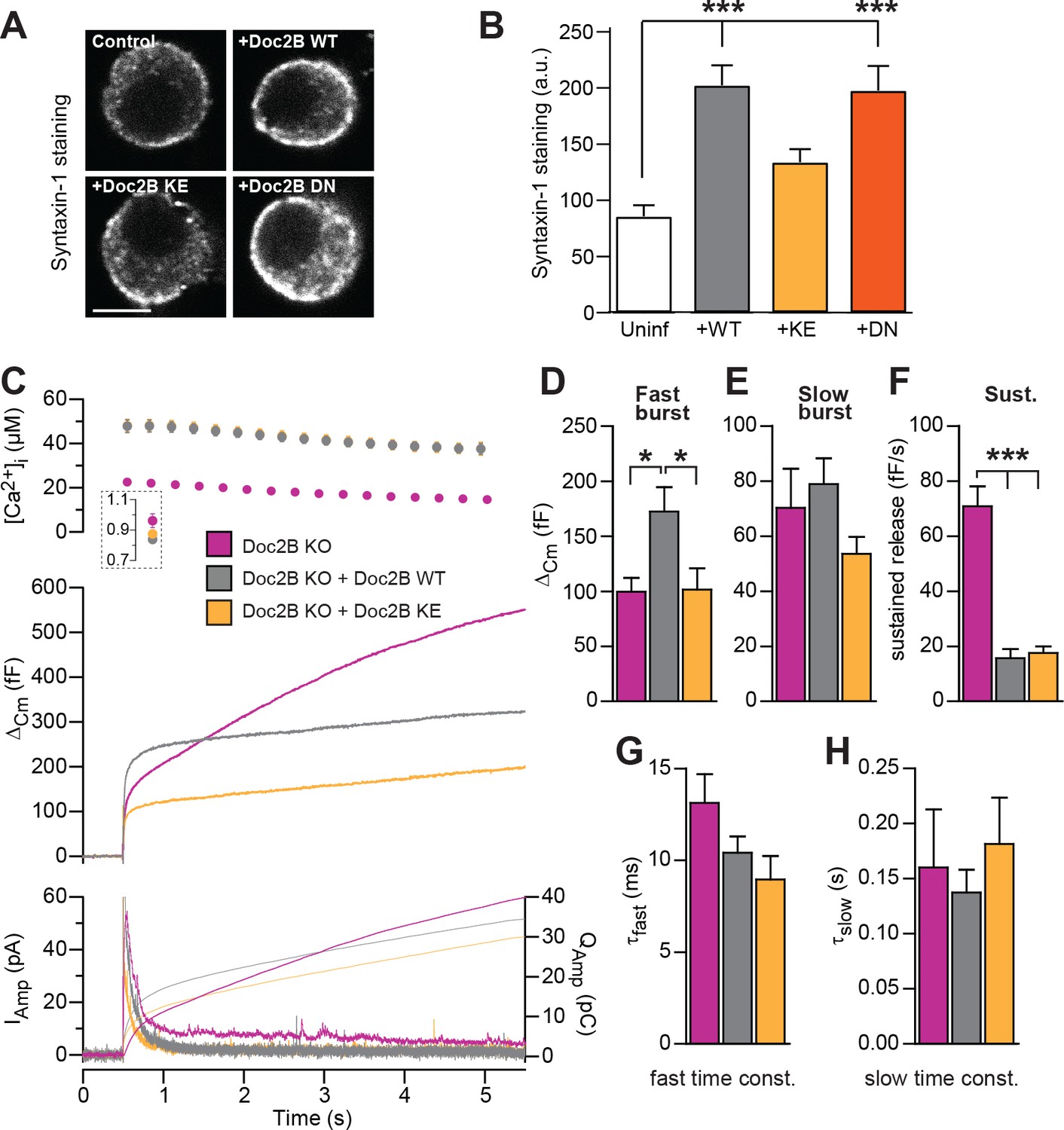
SNARE-binding of Doc2B is required for boosting the RRP, but not for inhibiting the sustained phase.
(A) Fixed and Immunostained wildtype chromaffin cells stained against syntaxin-1. (B) Quantification of the intensity of syntaxin-1 staining shows that expression of Doc2B WT (n = 12 cells) lead to an increase in intensity compared to non-expressing control cells (‘Uninf’, n = 11). The KE mutation (K237,319E; n = 15) did not significantly affect syntaxin-1 staining, whereas the DN mutant increased staining (n = 12). ***p<0.001, Dunn’s multiple comparison test. (C) Ca2+ uncaging experiment in Doc2B KO cells overexpressing Doc2B WT (n = 16) or Doc2B KE (K237,319E; n = 19). Recordings from another experiment in uninfected Doc2B KO cells (pink; same data as in Figure 9) are shown in parallel (n = 25); (D-H) Kinetic analysis of the capacitance traces. The size of the fast burst upon Doc2B KE overexpression was similar to Doc2B KO cells and smaller that in cells overexpressing Doc2B WT (D) while the slow burst showed a tendency for reduced size (E). The sustained phase was similarly affected by expression of either Doc2B KE or Doc2B WT (F). No differences were identified in the release time constants (G,H). *p<0.05, ***p<0.001; One-way ANOVA with Tukey's multiple comparison test.
Figure 8
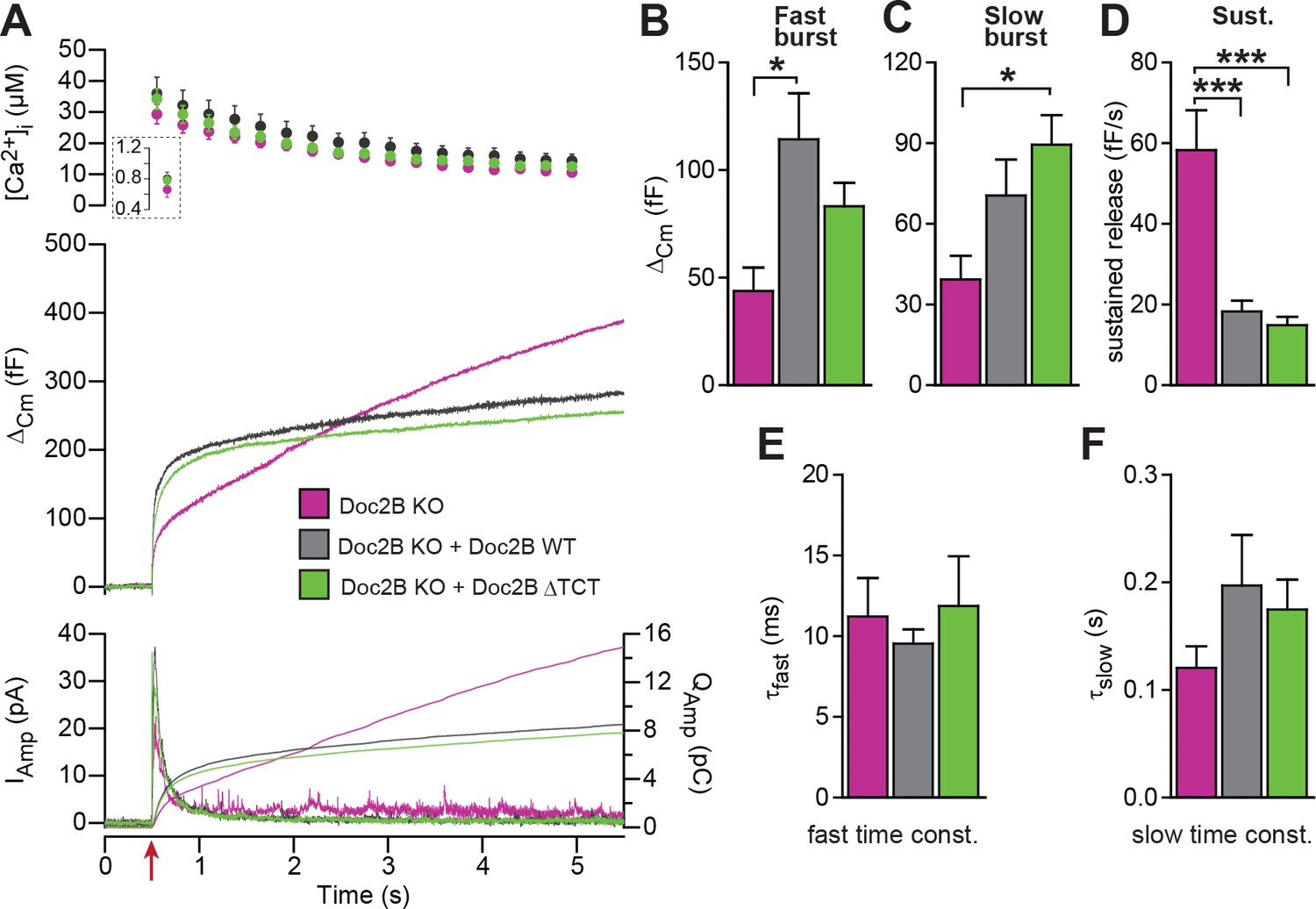
A Doc2B mutant lacking the binding sequence for Tctex-1 (Doc2BΔTCT) supports normal secretion.
(A) Ca2+ uncaging experiment in Doc2B KO cells (n = 13) and KO cells overexpressing Doc2B WT (n = 15) or Doc2BΔTCT (n = 14). (B–F) Kinetic analysis revealed that both vesicle pool sizes and kinetics of release were not significantly different between Doc2B WT and Doc2BΔTCT. *p<0.05; ***p<0.001, Dunn’s multiple comparison test.
Figure 9
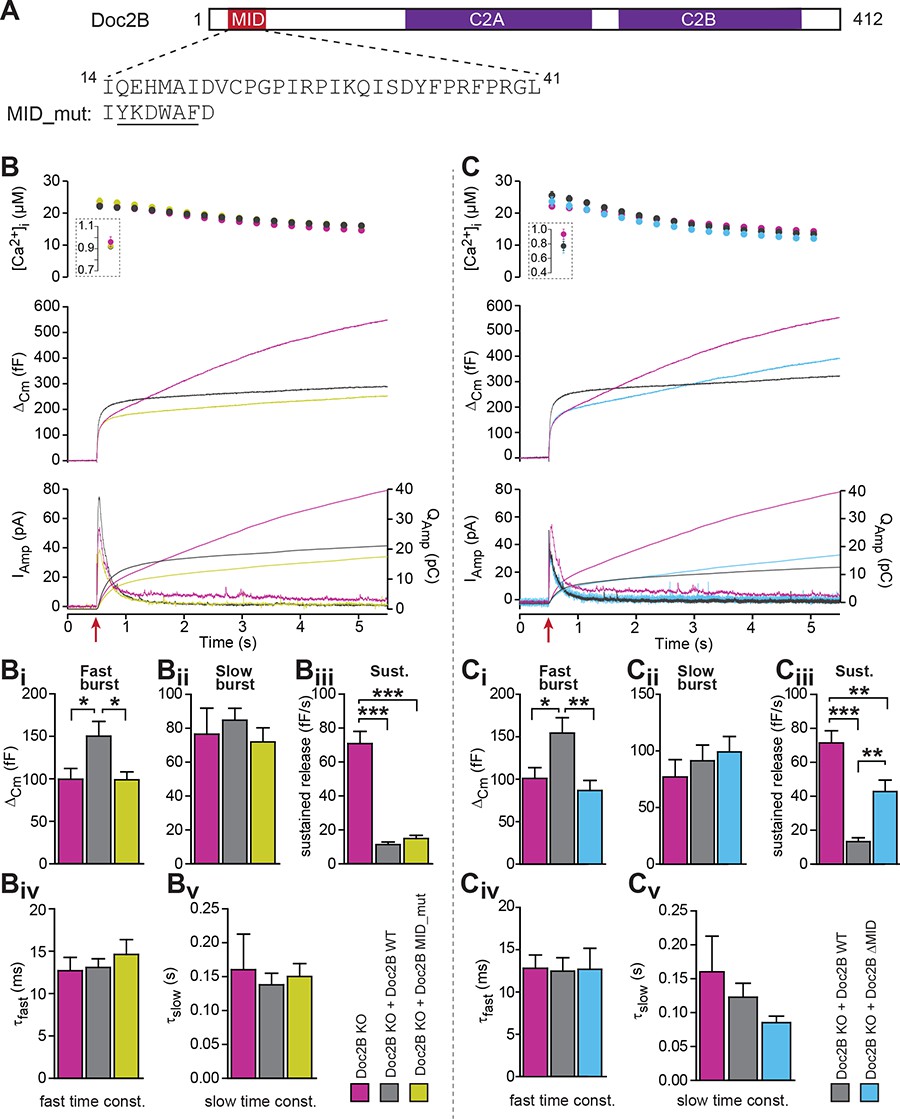
The MID-domain is required for both positive and negative functions of Doc2B in vesicle priming.
(A) Domain structure of mouse Doc2B. C2A and C2B are the two C2-domains. The MID-domain encompasses amino acids 14–38. We mutated the N-terminal part of the MID-domain (MID_mut, (Mochida et al., 1998)), or deleted it entirely (deletion of amino acids 14–41: ΔMID). (B) Ca2+ uncaging experiment in Doc2B KO cells (n = 25) and KO cells overexpressing Doc2B WT (n = 26) or Doc2B MID_mut (n = 26). (Bi-Bv) Kinetic analysis showed that mutating a stretch of the Doc2B MID domain abolishes the facilitating effect that Doc2B WT has in potentiating the RRP size (i.e. the size of the fast burst) (Bi) while causing a similar reduction in the rate of sustained release as the wildtype protein (Biii). (C), Full deletion of the Munc13-interacting domain of Doc2B (amino acids 14–41; Doc2B ΔMID) interferes with the function of Doc2B in both the fast burst and the sustained phase of release. Ca2+ uncaging experiment in Doc2B KO cells overexpressing Doc2B WT (n = 18) or Doc2B ΔMID (n = 20). (Ci-Cv) Kinetic analysis showing that removal of the full MID region of Doc2b interferes both with the promoting action on the filling of the fast pool of vesicles (Ci) and alleviates the suppression of the sustained phase of release caused by Doc2B WT overexpression (Ciii). The data from recordings of uninfected Doc2B KO cells used in panels C is the same as the one in panels B. *p<0.05, **p<0.01, ***p<0.001; Dunn’s multiple comparison test.
Figure 10
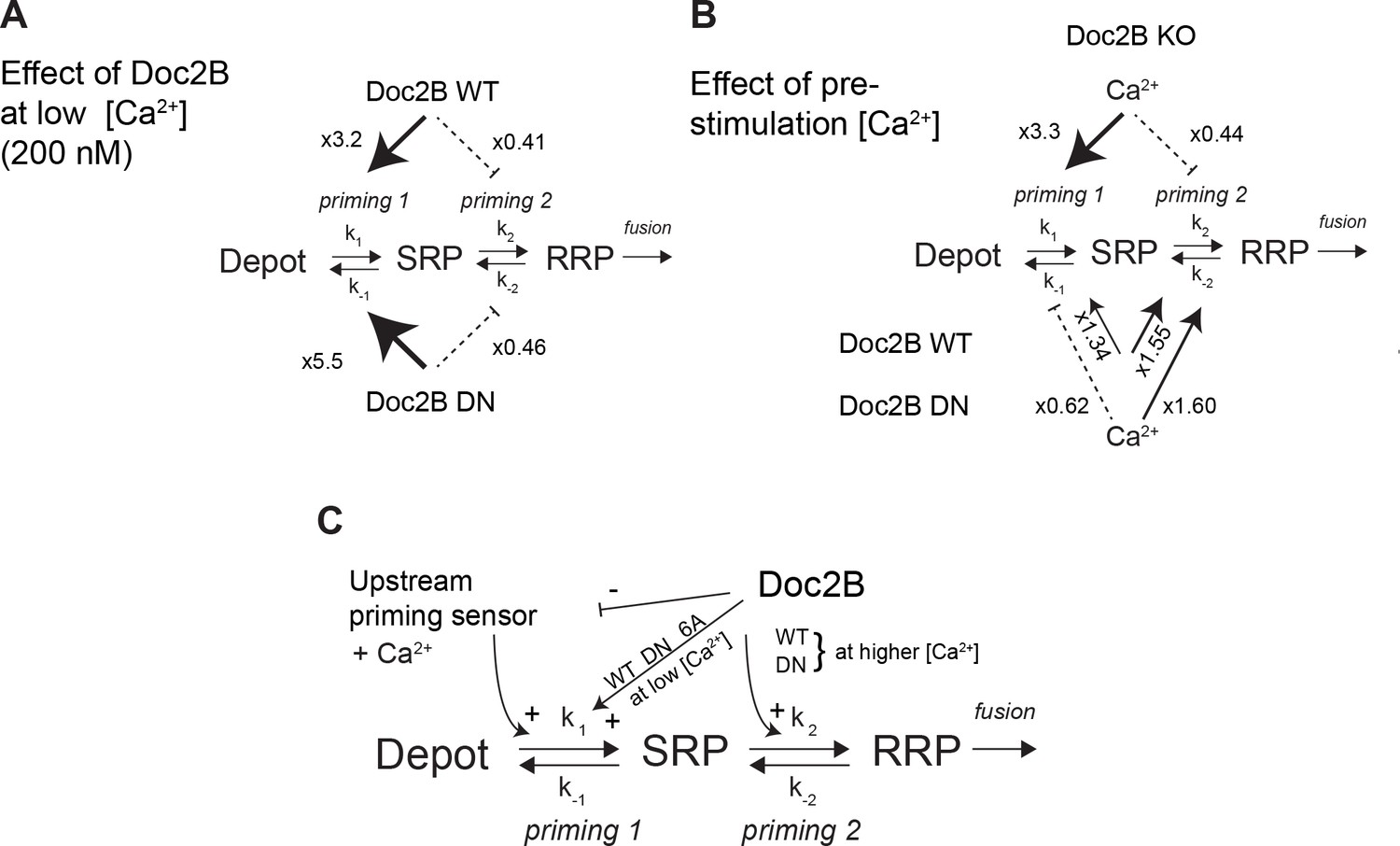
Dissecting the effect of Doc2B on two sequential priming steps.
(A) Chromaffin cells feature two sequential priming steps: priming 1, which recruits vesicles to the SRP, and priming 2, which recruits them to the RRP. At equilibrium, the overall propensity for forward priming is k1[Depot]/k-1 and k2/k-2, respectively (see Materials and methods). Shown is the multiplicative effect of expressing Doc2B WT and DN mutant on priming 1 and priming 2 propensities at low [Ca2+]. Both proteins stimulate priming 1, but inhibit priming 2. It has been proposed that SRP-vesicles fuse directly (Voets, 2000), or, as indicated here, that they mature to RRP-vesicles before they fuse (Walter et al., 2013); the experiments we present here do not distinguish between two possibilities, but see (Walter et al., 2013). (B) The effect of increasing [Ca2+] from 200 to ~ 800 nM, in the Doc2B KO, or after expression of Doc2B WT or DN. Ca2+ stimulates priming 1 strongly in the Doc2B KO, but inhibits priming 2, whereas with Doc2B, priming 2 is stimulated by Ca2+. (C) Overall model for the effect of Doc2B on priming 1 and priming 2. Doc2B (and the DN and 6A mutants) stimulates priming 1, but also occludes its Ca2+-dependence, presumably by inhibiting or competing with another Ca2+-dependent priming protein (upstream priming sensor) that normally acts at priming 1. Ca2+ increases the effect of Doc2B on priming 2; this effect is seen also in the DN-mutant, but not in the 6A-mutant; therefore, it depends on Ca2+ binding to the C2B-domain.
-
Figure 10—source data 1
Values for pool sizes and priming 1 and priming 2 propensity from experiments in Doc2B KO, and after overexpression of Doc2B WT and DN-mutant (see also Figure 4).
The propensity for upstream priming, priming 1, k1[Depot]/k-1, is simply equal to the slow burst (SRP) size, whereas the propensity for priming 2, k2/k-2, is equal to the ratio between fast and slow burst (RRP/SRP). To determine the effect of increasing basal [Ca2+] from ~200 nM to ~800 nM, we calculated the ratio of the propensities between Ca2+ concentrations for a given genotype (two columns on the right). To calculate the effect of expressing Doc2B and the Doc2B DN mutant at ~200 nM [Ca2+] we formed the ratio of the propensities between expressed and KO cells for a given [Ca2+] (six bottom rows).
- https://doi.org/10.7554/eLife.27000.014
Tables
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| strain, strain background (M. musculus) | C57BL/6 | |||
| strain, strain background (M. musculus) | CD1 | |||
| genetic reagent (M. musculus) | Doc2a null allele | Sakaguchi G, Manabe T, Kobayashi K, Orita S, Sasaki T, Naito A, Maeda M, Igarashi H, Katsuura G, Nishioka H, Mizoguchi A, Itohara S, Takahashi T, Takai Y. Doc2alpha is an activity- dependent modulator of excitatory synaptic transmission. Eur J Neurosci. 1999 Dec;11(12):4262–8. | PMID: 10594652 | |
| genetic reagent (M. musculus) | Doc2b null allele | Groffen AJ, Martens S, Díez Arazola R, Cornelisse LN, Lozovaya N, de Jong AP, Goriounova NA, Habets RL, Takai Y, Borst JG, Brose N, McMahon HT, Verhage M. Doc2b is a high-affinity Ca2+ sensor for spontaneous neurotransmitter release. Science. 2010 Mar 26;327 (5973):1614–8. | PMID: 20150444 | |
| genetic reagent (M. musculus) | Synaptotagmin-7 (syt7) null allele | Maximov A, Lao Y, Li H, Chen X, Rizo J, Sørensen JB, Südhof TC. Genetic analysis of synaptotagmin-7 function in synaptic vesicle exocytosis. Proc Natl Acad Sci U S A. 2008 Mar 11;105(10):3986–3991. | PMID: 18308933 | |
| genetic reagent (M. musculus) | Synaptotagmin-1 (syt1) null allele | Geppert M, Goda Y, Hammer RE, LI C, Rosahl TW, Stevens CF, Südhof TC. 1994. Synaptotagmin I: a major Ca2 + sensor for transmitter release at a central synapse. Cell 79(4): 717–727. | PMID: 7954835 | |
| genetic reagent (M. musculus) | Unc13b null allele | Varoqueaux F, Sigler A, Rhee J-S, Brose N, Enk C, Reim K, Rosenmund C. Total arrest of spontaneous and evoked synaptic transmission but normal synaptogenesis in the absence of Munc13-medicated vesicle priming. Proc Natl Acad Sci U S A. 2002. Jun 25; 99(13):9037–9042. | PMID: 12070347 | |
| transfected construct (R. norvegicus) | pSFV-Doc2b-WT- IRES-EGFP | Friedrich R, Groffen AJ, Connell E, van Weering JR, Gutman O, Henis YI, Davletov B, Ashery U. DOC2B acts as a calcium switch and enhances vesicle fusion. J Neurosci. 2008 Jul 2;28(27):6794–806. | Genbank Accession: EU635444.1 | local reference: SG#334 |
| transfected construct (R. norvegicus) | pSFV-Doc2b-DN- IRES-EGFP | Friedrich R, Groffen AJ, Connell E, van Weering JR, Gutman O, Henis YI, Davletov B, Ashery U. DOC2B acts as a calcium switch and enhances vesicle fusion. J Neurosci. 2008 Jul 2;28(27):6794–806. | Genbank Accession: EU635445.1 | derived from EU635444.1 with indicated mutations |
| transfected construct (R. norvegicus) | pSFV-Doc2b-6A- IRES-EGFP | this paper | derived from EU635444.1 with indicated mutations | |
| transfected construct (R. norvegicus) | pSFV-Doc2b-KE- IRES-EGFP | this paper | derived from EU635444.1 with indicated mutations | |
| transfected construct (R. norvegicus) | pSFV-Doc2b-TCT- IRES-EGFP | this paper | derived from EU635444.1 with indicated mutations | |
| transfected construct (R. norvegicus) | pSFV-Doc2b-MID (14-41)-IRES-EGFP | this paper | derived from EU635444.1 with indicated mutations | |
| transfected construct (R. norvegicus) | pSFV-Doc2b-MID (Scrambled)- IRES-EGFP | this paper | derived from EU635444.1 with indicated mutations | |
| transfected construct (R. norvegicus) | pLenti- Doc2bEGFP-WT | this paper | derived from EU635444.1; local reference: J274#2 | |
| transfected construct (R. norvegicus) | pLenti- Doc2bEGFP-DN | this paper | local reference: J274#7 | |
| transfected construct (R. norvegicus) | pLenti- Doc2bEGFP-6A | this paper | local reference: K102#33 | |
| transfected construct (R. norvegicus) | pSFV-EGFP- Doc2B-DeltaMID | this paper | local reference: #480 | |
| transfected construct () | ||||
| transfected construct () | ||||
| transfected construct () | ||||
| biological sample () | ||||
| antibody | anti-syntaxin | Synaptic System | SySy: 110011 | dil. 1:1000; Overnight/room temperature |
| antibody | anti-GFP | Abcam | Ab13970 | dil. 1:1000; Overnight/room temperature |
| antibody | secondary Goat anti-mouse Alexa 488 | Invitrogen | Invitrogen: A11029 | dil. 1:1000; Overnight/4 deg. |
| antibody | secondary Goat anti-chicken Alexa 488 | Abcam | Ab150169 | dil. 1:1000; Overnight/4 deg. |
| antibody | anti- synaptotagmin-1 | Synaptic System | SySy: 105011 | dil. 1:1000; Overnight/4 deg. |
| antibody | anti- synaptotagmin-7 | Synaptic System | SySy: 105173 | dil. 1:500; Overnight/4 deg. |
| antibody | anti-VCP | Abcam | Ab11433 | dil. 1:2000; 1h/room temperature |
| antibody | mouse mAb against Syb2, clone 69.1 | Synaptic Systems GmbH | cat#104211 | 1:2500 diluted in PBS+ 2% BSA, incubated 2 hr at RT |
| antibody | rabbit pAb against Doc2b | Groffen AJ, Brian EC, Dudok JJ, Kampmeijer J, Toonen RF, Verhage M. Ca(2+)-induced recruitment of the secretory vesicle protein DOC2B to the target membrane. J Biol Chem. 2004 May 28;279(22):23740–7. | PMID: 15033971 | 1:133 diluted in PBS+ 2% BSA, incubated 2 hr at RT (note: Doc2b and Syb2 stainings were performed separately on different coverslips to rule out potential spectral overlap)/For western blotting: dil. 1:500; Overnight at 4 deg |
| antibody | Goat anti-rabbit HRP | Agilent | Dako-P0448 | dil. 1:10000; 1h30/room temperature |
| antibody | Goat anti-mouse HRP | Agilent | Dako-P0447 | dil. 1:10000; 1h30/room temperature |
| antibody | Goat-anti-Rabbit Alexa546 | ThermoFisher Scientific | cat#A-11010 | 1:1000 diluted in PBS+ 2% BSA |
| antibody | Goat-anti-Mouse Alexa647 | ThermoFisher Scientific | cat#10739374 | 1:1000 diluted in PBS+ 2% BSA |
| antibody | ||||
| recombinant DNA reagent | ||||
| sequence-based reagent | ||||
| peptide, recombinant protein | ||||
| commercial assay or kit | BCA Protein assay kit | Pierce | Pierce: 23227 | |
| commercial assay or kit | ||||
| commercial assay or kit | ||||
| chemical compound, drug | NaCl | Sigma-aldrich | Sigma-aldrich: S9888 | |
| chemical compound, drug | KCl | Sigma-aldrich | Sigma-aldrich: P5405 | |
| chemical compound, drug | NaH2PO4 | Sigma-aldrich | Sigma-aldrich: S8282 | |
| chemical compound, drug | Glucose | Sigma-aldrich | Sigma-aldrich: G8270 | |
| chemical compound, drug | DMEM | Gibco | Gibco: 31966047 | |
| chemical compound, drug | L-cysteine | Sigma-aldrich | Sigma-aldrich: C7352 | |
| chemical compound, drug | CaCl2 | Sigma-aldrich | Sigma-aldrich: 499609 | |
| chemical compound, drug | EDTA | Sigma-aldrich | Sigma-aldrich: E5134 | |
| chemical compound, drug | papain | Worthington Biochemical | Worthington Biochemical: LS003126 | |
| chemical compound, drug | albumin | Sigma-aldrich | Sigma-aldrich: A3095 | |
| chemical compound, drug | trypsin-inhibitor | Sigma-aldrich | Sigma-aldrich: T9253 | |
| chemical compound, drug | penicillin/ streptomycin | Invitrogen | Invitrogen: 15140122 | |
| chemical compound, drug | insulin-transferrin- selenium-X | Invitrogen | Invitrogen: 51500056 | |
| chemical compound, drug | fetal calf serum | Invitrogen | Invitrogen: 10500064 | |
| chemical compound, drug | MgCl2 | Sigma-aldrich | Sigma-aldrich: 449172 | |
| chemical compound, drug | HEPES | Sigma-aldrich | Sigma-aldrich: H3375 | |
| chemical compound, drug | Nitrophenyl- EGTA (NPE) | Synthesized at the Max-Planck-Institut for biophycial chemistry, Göttingen. | ||
| chemical compound, drug | Fura-4F | Invitrogen | Invitrogen: F14174 | |
| chemical compound, drug | Furaptra | Invitrogen | Invitrogen: M1290 | |
| chemical compound, drug | Mg-ATP | Sigma-aldrich | Sigma-aldrich: A9187 | |
| chemical compound, drug | GTP | Sigma-aldrich | Sigma-aldrich: G8877 | |
| chemical compound, drug | Vitamin C | Sigma-aldrich | Sigma-aldrich: A5960 | |
| chemical compound, drug | EGTA | Sigma-aldrich | Sigma-aldrich: E4378 | |
| chemical compound, drug | Paraformaldehyde | Sigma-aldrich | Sigma-aldrich: P6148 | |
| chemical compound, drug | PIPES | Sigma-aldrich | Sigma-aldrich: 80635 | |
| chemical compound, drug | Triton X-100 | Sigma-aldrich | Sigma-aldrich: T8787 | |
| chemical compound, drug | BSA | Sigma-aldrich | Sigma-aldrich: A4503 | |
| chemical compound, drug | Prolong Gold | Invitrogen | Invitrogen: P36934 | |
| chemical compound, drug | Protease cocktail inhibitor | Invitrogen | Invitrogen: 87785 | |
| chemical compound, drug | RIPA buffer | Invitrogen | Invitrogen: R0278 | |
| chemical compound, drug | ECL plus western blotting substrate | Pierce | Pierce: 32132 | |
| chemical compound, drug | ||||
| chemical compound, drug | ||||
| chemical compound, drug | ||||
| chemical compound, drug | ||||
| software, algorithm | Igor | wavemetrics | ||
| software, algorithm | ImageJ | NIH software |
Additional files
-
Transparent reporting form
- https://doi.org/10.7554/eLife.27000.015
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Doc2B acts as a calcium sensor for vesicle priming requiring synaptotagmin-1, Munc13-2 and SNAREs
eLife 6:e27000.
https://doi.org/10.7554/eLife.27000




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}