Post-translational thioamidation of methyl-coenzyme M reductase, a key enzyme in methanogenic and methanotrophic Archaea
- University of Illinois, United States
Figures
Figure 1 with 1 supplement
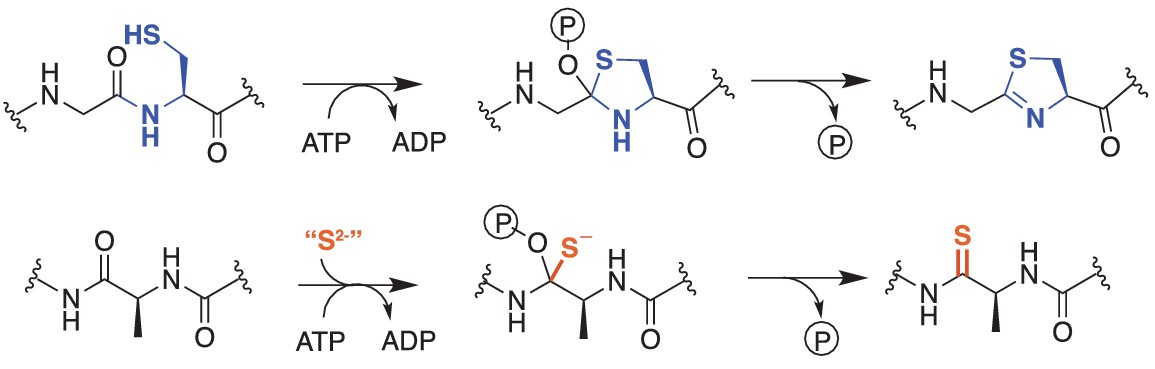
Comparison of reactions catalyzed by YcaO proteins.
Top, Biochemically characterized YcaO proteins involved in the synthesis of azol(in)e-containing ribosomal natural products catalyze the ATP-dependent cyclodehydration of cysteine, serine, and threonine residues. Shown is the conversion of peptidic cysteine to thiazoline. This reaction proceeds via an O-phosphorylated hemiorthoamide intermediate. Bottom, An analogous reaction is believed to occur in the biosynthesis of the thioamide bond in thioviridamide. Rather than an adjacent cysteine acting as the nucleophile, an exogenous source of sulfide (S2-) is required for this reaction.
Figure 1—figure supplement 1
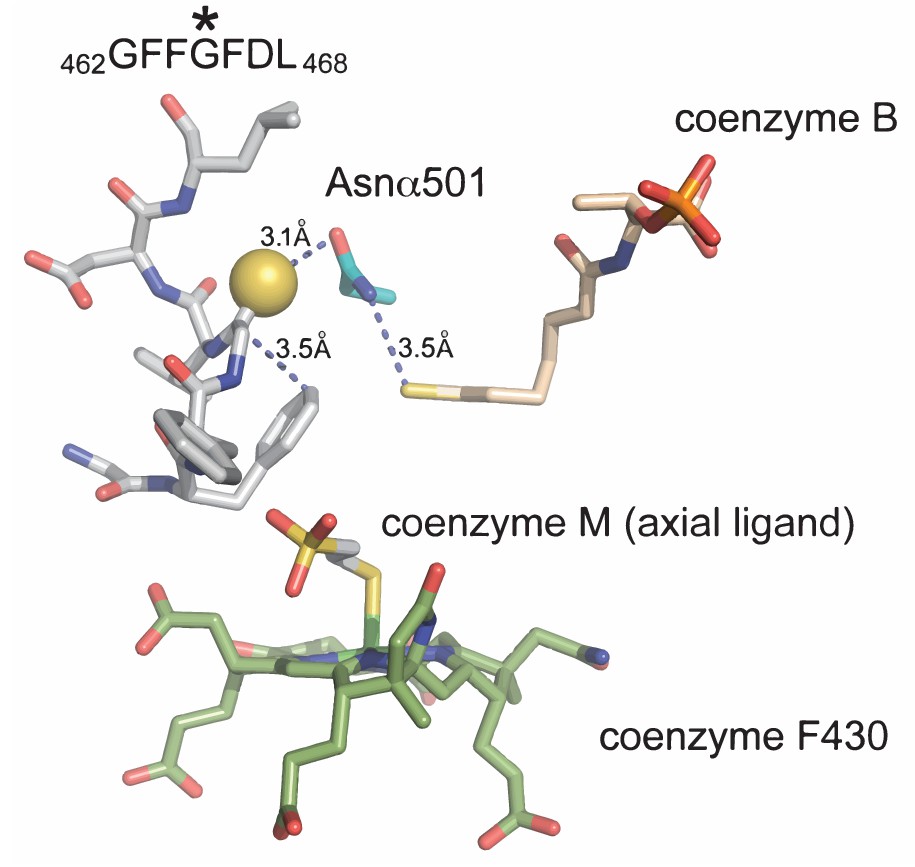
A view of the MCR active site using the crystal structure of M. barkeri (Protein DataBank entry 1E6Y).
The peptide backbone of McrA in the vicinity of the thioglycine (Gly462-Leu468) is shown in gray sticks with the thioamide sulfur of Gly465 shown as a yellow sphere. Coenzymes B, M, and F430 are shown with several interatomic distances to McrA-Asn501.
Figure 2 with 2 supplements

A maximum-likelihood phylogenetic tree of YcaO homologs in archaea.
Taxa in green, blue, and orange depict methanogens and anaerobic methane-oxidizing archaea (ANMEs) with sequenced genomes. Taxa shown in green contain a single copy of ycaO and tfuA, those in blue contain two copies of ycaO and one copy of tfuA, while those in orange contain one copy of ycaO, but do not encode tfuA. Taxa shown in pink are other archaea that also encode ycaO and tfuA homologs. The gene neighborhoods of selected taxa are also depicted, showing the common co-localization with genes annotated as having a role in sulfur metabolism or methanogenesis. The node labels indicate support values calculated using the Shiomdaira-Hasegawa test using 1000 resamples. Support values less than 0.5 have not been shown. The outgroup displayed derives from a variety of bacterial YcaO proteins.
Figure 2—figure supplement 1
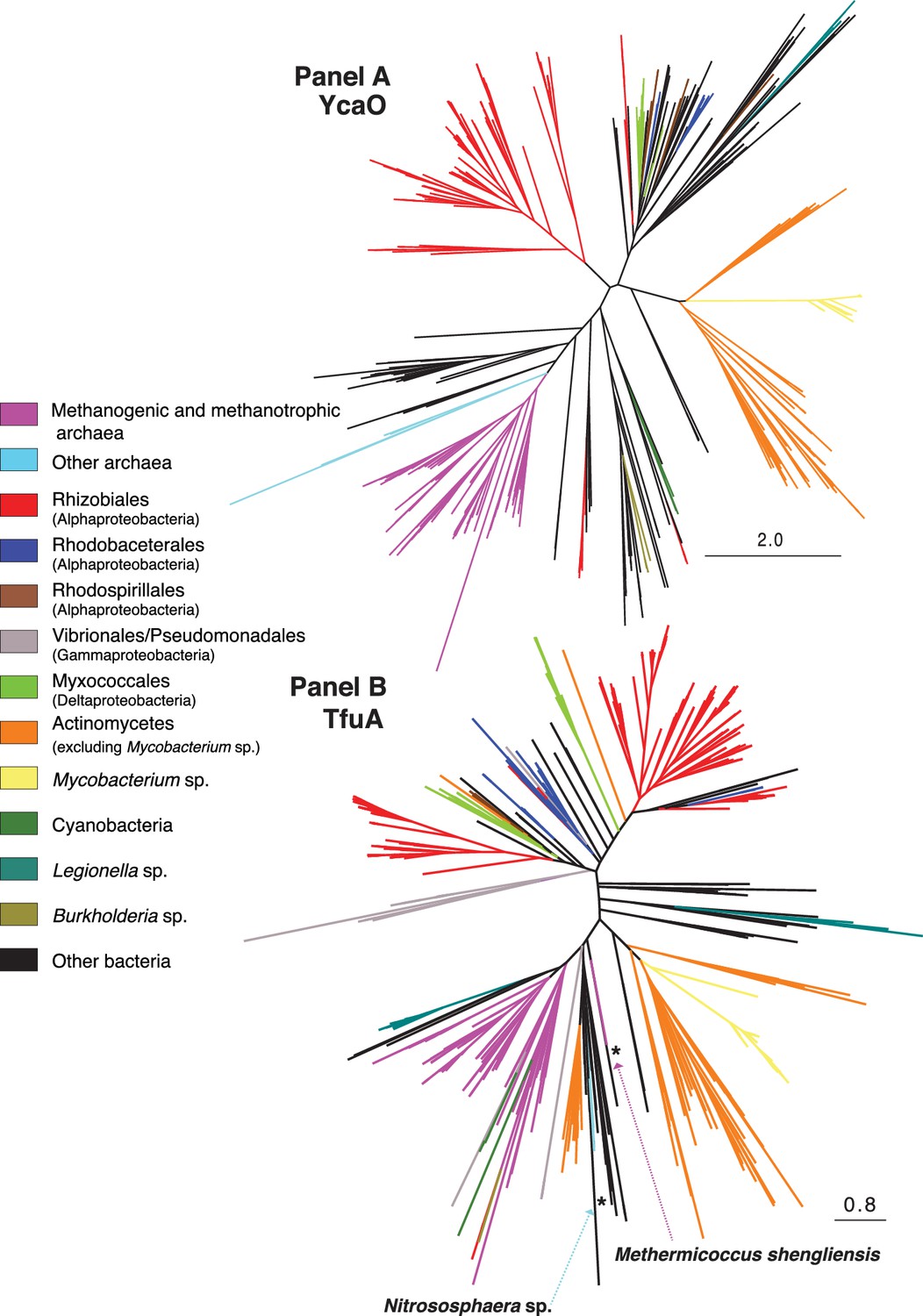
(Panel A) Unrooted maximum-likelihood phylogeny of 1,000 YcaO sequences retrieved from the NCBI non-redundant protein sequence database using the corresponding sequence from Methanosarcina acetivorans as a search query.
(Panel B) Unrooted maximum-likelihood phylogeny of 1,000 TfuA sequences retrieved from the NCBI non-redundant protein sequence database using the corresponding sequence from M. acetivorans as a search query. Divergent archaea-derived sequences in the TfuA tree are denoted with an *.
Figure 2—figure supplement 2
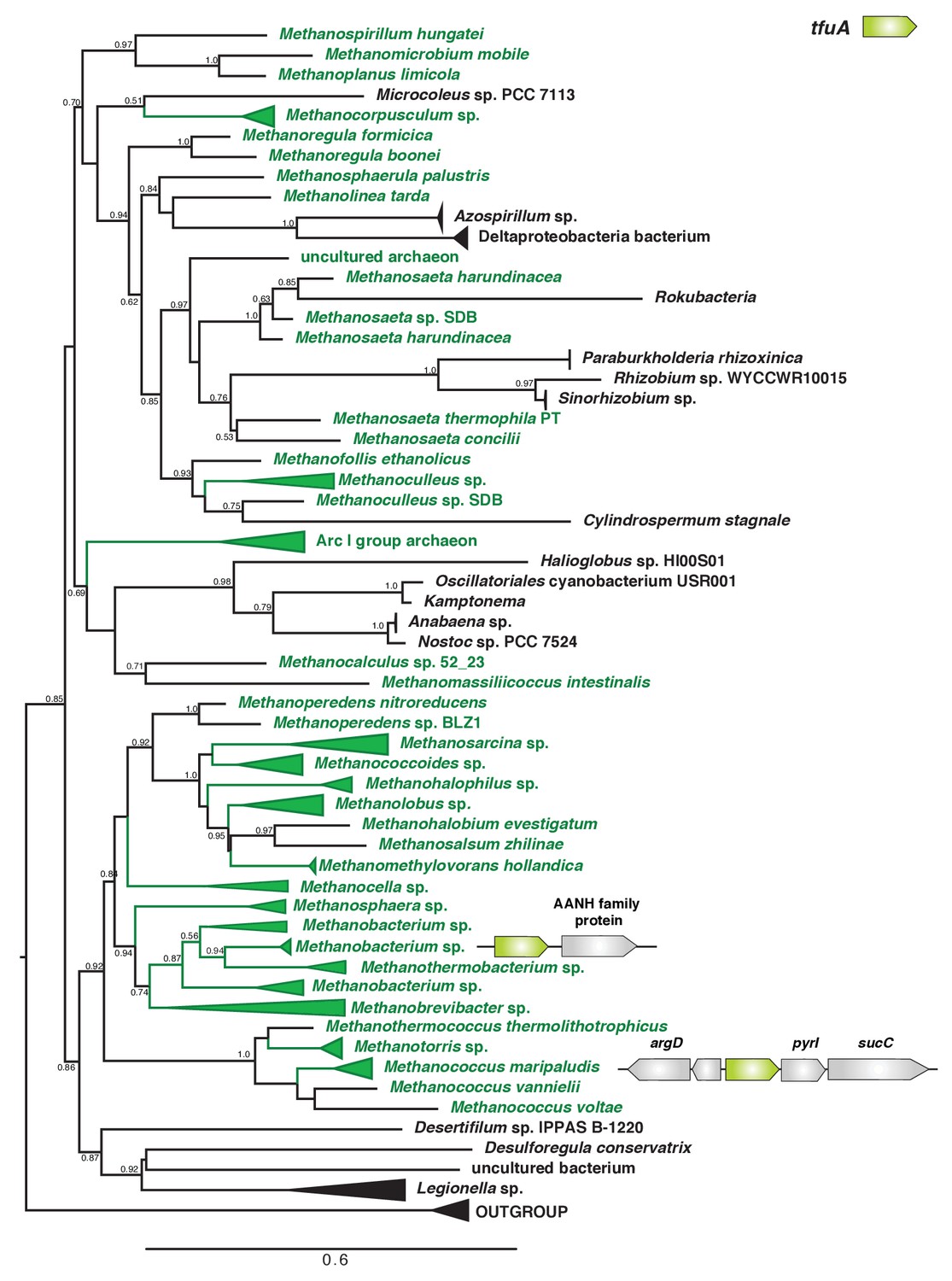
Unrooted maximum-likelihood phylogeny of 1000 TfuA sequences retrieved from the NCBI non-redundant protein sequence database using the corresponding sequence from Methanosarcina acetivorans as a search query.
For taxa where tfuA and ycaO are not adjacently encoded genes a gene architecture diagram is shown. The node labels indicate support values calculated using the Shiomdaira-Hasegawa test using 1000 resamples. Support values less than 0.5 have not been shown. The outgroup is comprised of various bacterial TfuA protein sequences.
Figure 3 with 2 supplements
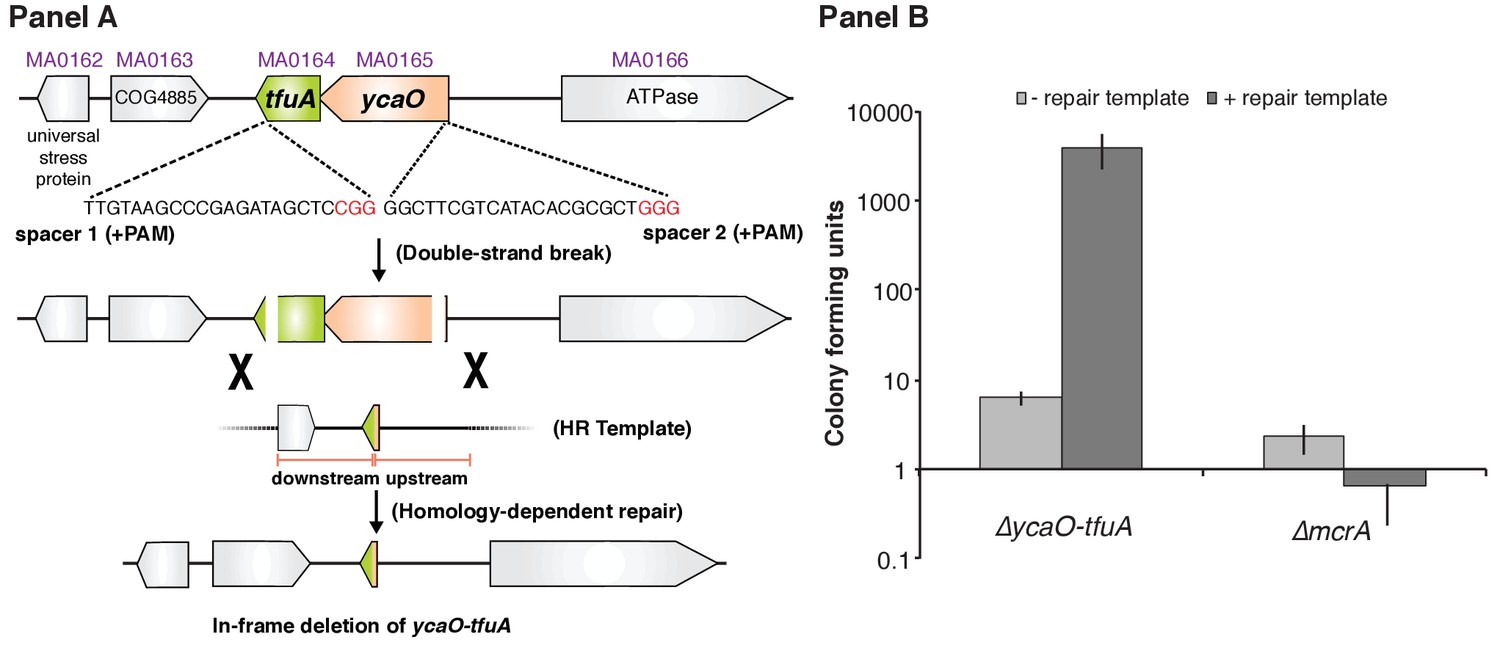
Panel A, Experimental strategy for deletion of the ycaO-tfuA locus in M. acetivorans using a Cas9-mediated genome editing technique.
Co-expression of the Cas9 protein along with a single guide RNA (sgRNA) with a 20 bp target sequence flanked by a 3’ NGG protospacer adjacent motif on the chromosome (PAM; in red) generates double-stranded breaks at the ycaO-tfuA locus. Repair of this break via homologous recombination with an appropriate repair template (HR Template) generates an in-frame deletion on the host chromosome. Panel B, A Cas9-based assay for gene essentiality. Mean transformation efficiencies of plasmids with (dark gray) or without (light gray) an appropriate repair template are shown. The known essential gene mcrA is included as a positive control. The error bars represent one standard deviation of three independent transformations.
Figure 3—figure supplement 1

A PCR-based screen to genotype the ycaO-tfuA locus in Methanosarcina acetivorans.
The PCR amplicon from three independently derived mutants containing an unmarked in-frame deletion of ycaO-tfuA (Lanes 2–4) is 2980 bp whereas the corresponding amplicon from a strain containing the wild-type locus is 4742 bp (Lane 5). Lanes 1 and 6 contain a 1 kb DNA ladder.
Figure 3—figure supplement 2
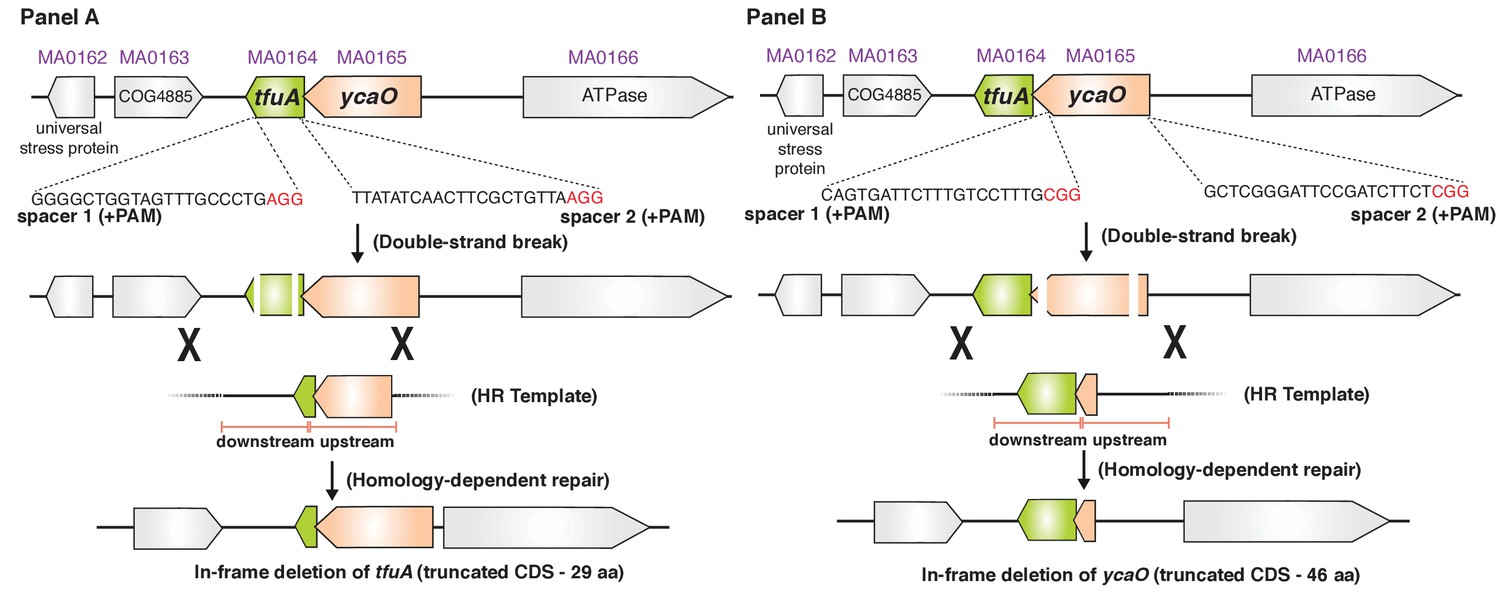
The experimental strategy for generating an in-frame deletion of either the tfuA locus (Panel A) or the ycaO locus (Panel B) in M. acetivorans using a Cas9-mediated genome editing technique.
Co-expression the Cas9 protein along with single guide RNAs (sgRNAs) containing a 20 bp target sequence flanked by a 3’ NGG protospacer adjacent motif on the chromosome (PAM; in red) generates double-stranded breaks. Repair of this break via homologous recombination with an appropriate repair template (HR Template) generates an in-frame deletion on the host chromosome.
Figure 4 with 3 supplements
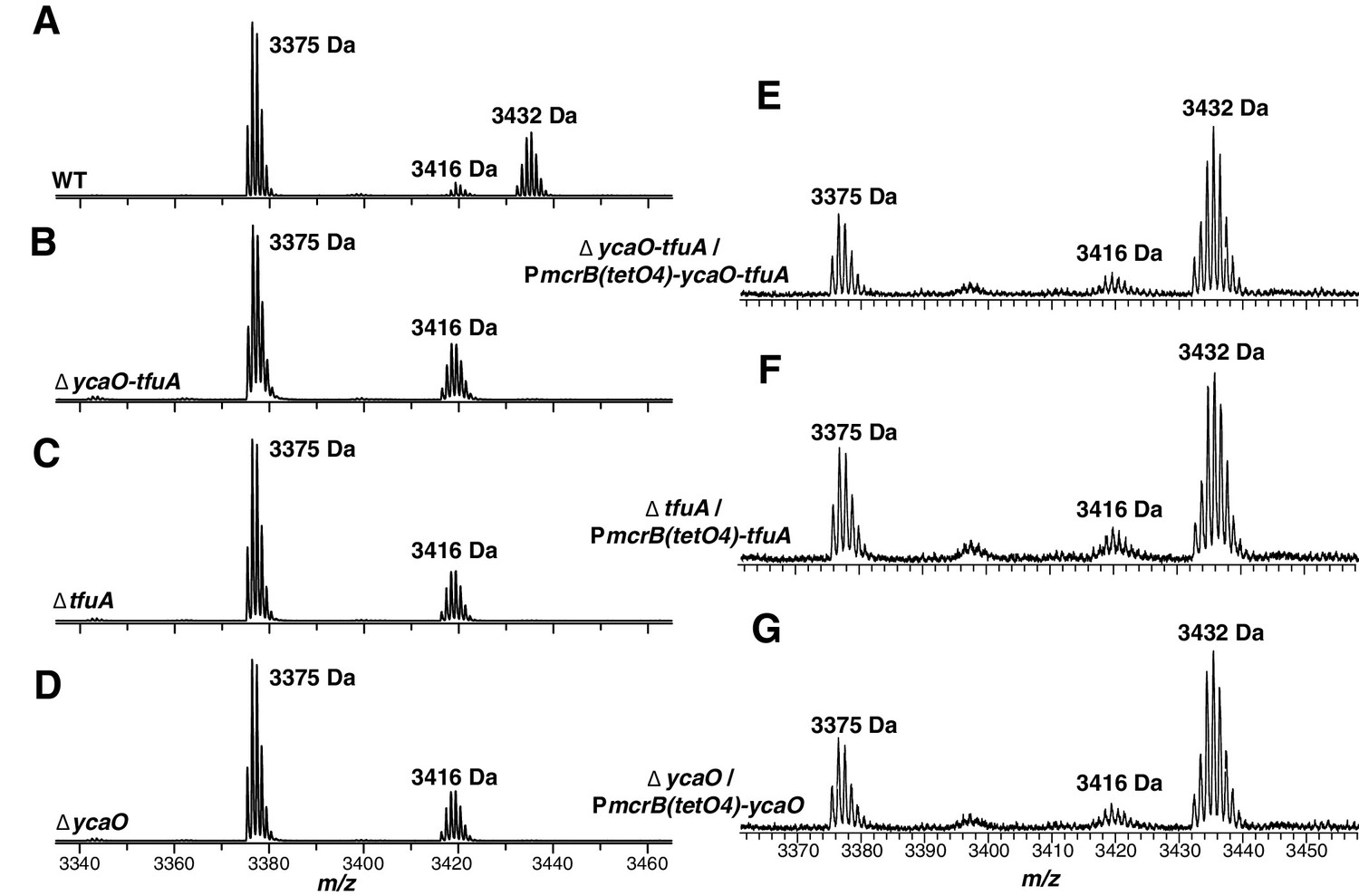
MALDI-TOF MS analysis of McrA.
Panel A, Spectrum obtained from trypsinolysis of WT MCR, which contains the thioglycine-modified L461-R491 peptide (m/z 3432 Da). A small amount of the unmodified L461-R491 peptide (m/z 3416 Da) is also observed, which probably arises via non-enzymatic hydrolysis of the thioamide bond as previously reported (Kahnt et al., 2007). Panels B, C, D, Spectrum obtained from trypsinolysis of McrA from the ∆ycaO-tfuA, ∆tfuA, ∆ycaO strains, respectively, containing only the unmodified L461-R491 peptide (m/z 3416 Da). Panels E, F, G. Spectrum obtained from trypsinolysis of MCR from the ∆ycaO-tfuA mutant complemented with PmcrB(tetO4)-ycaO-tfuA, ∆tfuA mutant complemented with PmcrB(tetO4)-tfuA, ∆ycaO mutant complemented with PmcrB(tetO4)-ycaO, respectively. Tetracycline was added to a final concentration of 100 µg/mL to the growth medium to induce expression from the PmcrB(tetO4) promoter. All three spectra contain the thioglycine-modified Leu461-Arg491 peptide (m/z 3432 Da). The m/z 3375 Da species present in all samples corresponds to McrA residues D392-R421.
Figure 4—figure supplement 1

(Panel A) High-resolutions electrospray ionization tandem mass spectrometry (HR-ESI MS/MS) analysis of a wild-type tryptic peptide from Methanosarcina acetivorans McrA (Leu461-Arg491, m/z 3432 Da).
The b6 ion localizes the thioamide modification to Gly465. No b5 ion was detected suggesting negligible fragmentation at the thioamide bond. Predicted masses and associated errors are shown. (Panel B) HR-ESI-MS/MS analysis of the corresponding McrA tryptic peptide from the ∆ycaO-tfuA deletion strain of M. acetivorans (m/z 3416 Da). The b5 and b6 ion localizes the amide functionality at Gly465. Predicted masses and associated errors are shown.
Figure 4—figure supplement 2
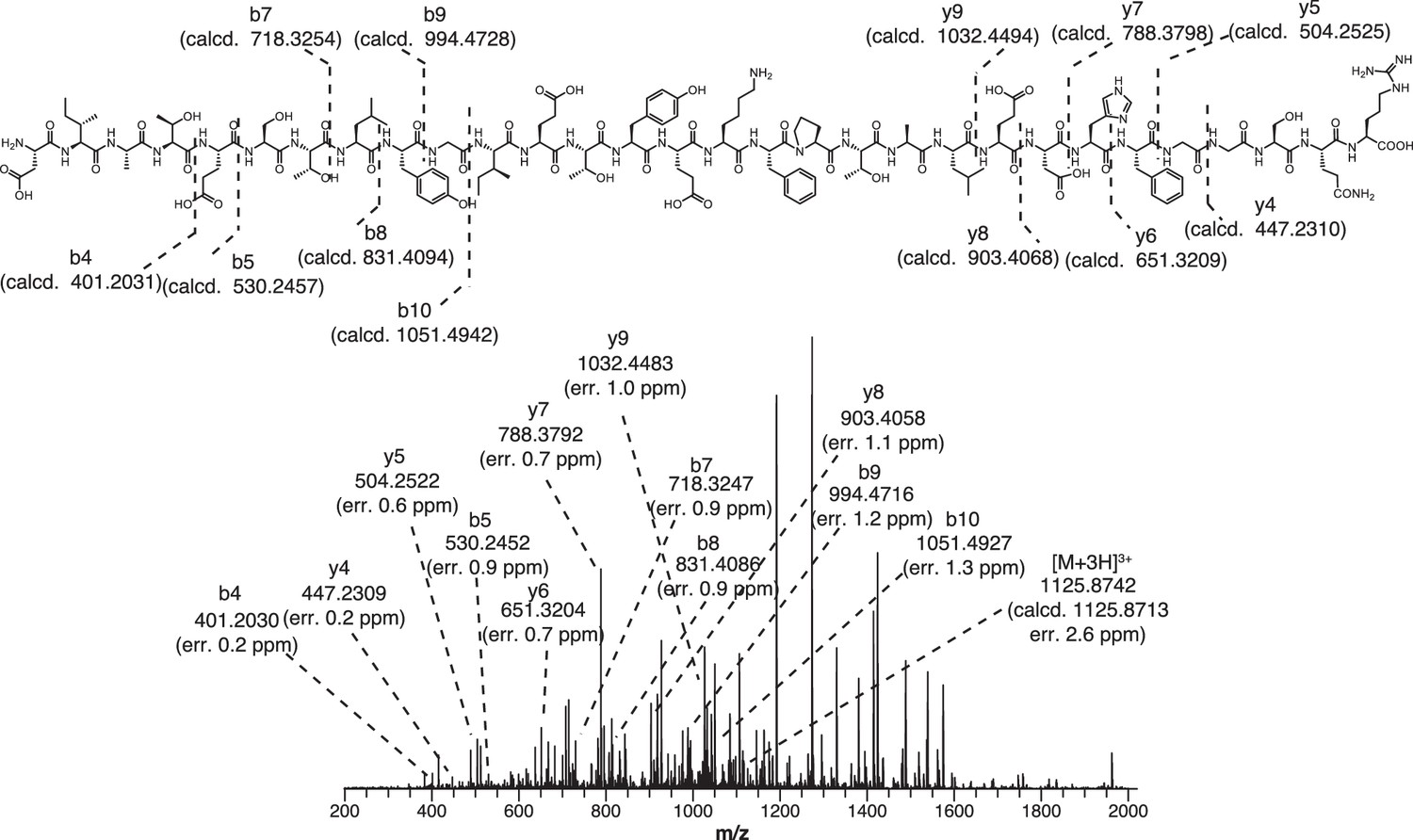
HR-ESI-MS/MS of the tryptic peptide from Methanosarcina acetivorans McrA (Asp392-Arg421, m/z 3375 Da) present in the wild-type and ∆ycaO-tfuA, ∆tfuA, ∆ycaO deletion strains.
Predicted masses and associated errors are shown.
Figure 4—figure supplement 3
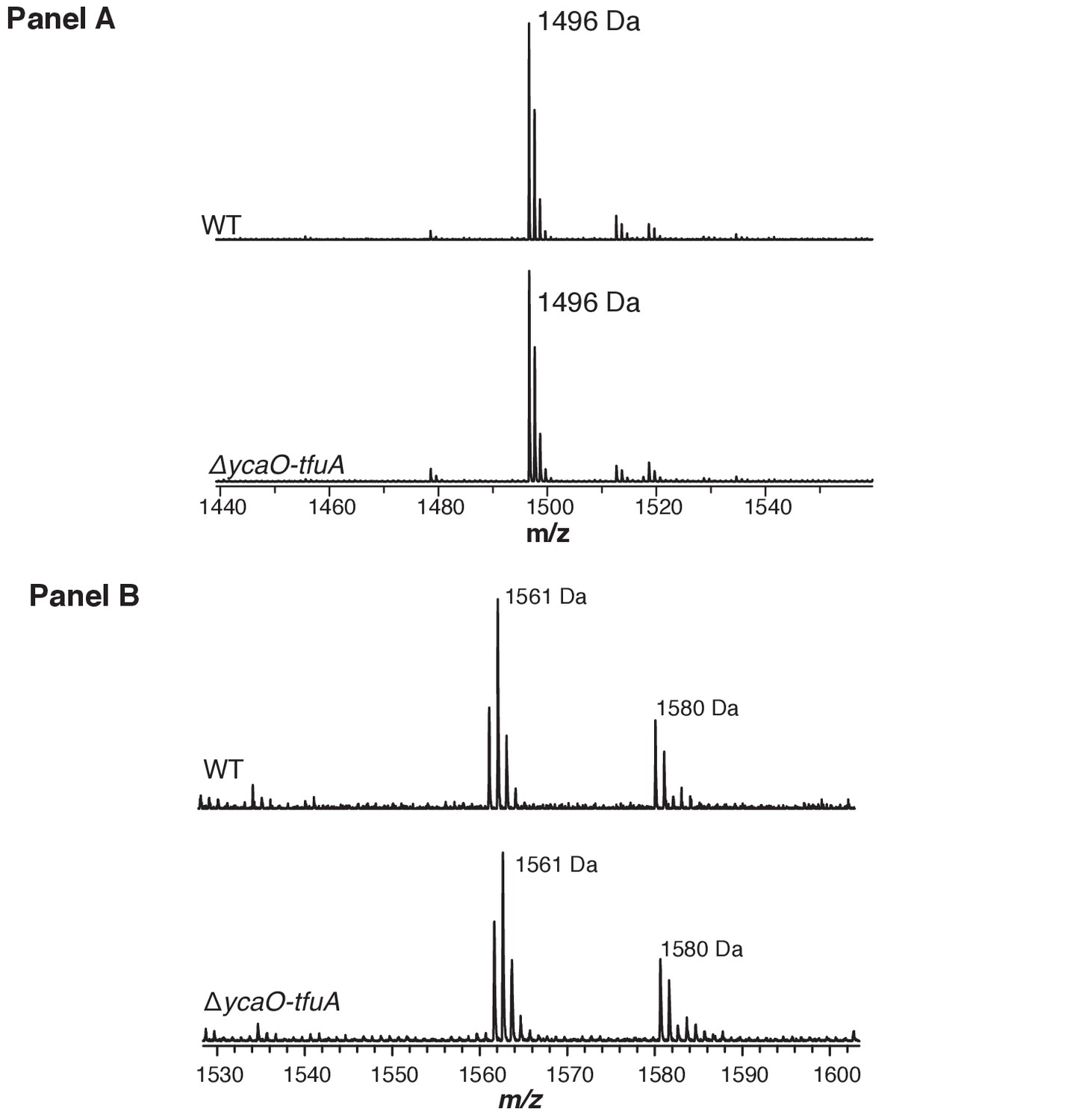
(Panel A) Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS) analysis of the wild-type (WT, top) and ∆ycaO-tfuA deletion strain (bottom) spectra identify the McrA tryptic peptide, His271-Arg284 (m/z 1496 Da), containing the known N-methylhistidine modification.
This suggests that the thioglycine formation is not a prerequisite for this methylation event. (Panel B) MALDI-TOF MS analysis of the WT (top) and ∆ycaO-tfuA deletion strain (bottom) spectra identify the McrA tryptic peptide, Phe408-Arg421, (m/z 1561 Da). This peptide has been reported to contain a β-methylation at Gln420 in other methanogens (Kahnt et al., 2007), however, it remains unmodified in M. acetivorans.
Figure 5 with 2 supplements
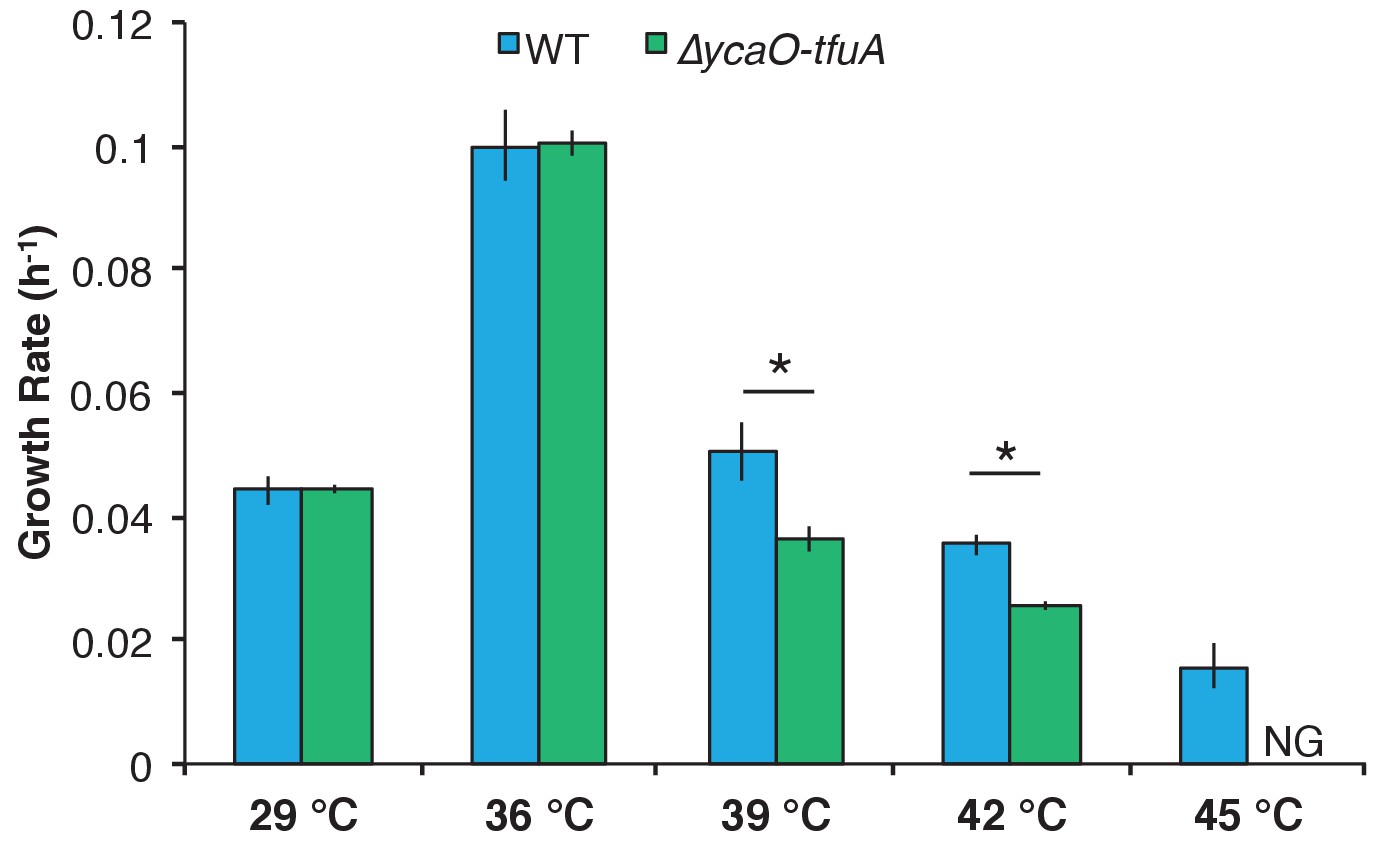
Growth rate of WT (blue) and the ∆ycaO-tfuA mutant (green) in HS medium with 125 mM methanol at the indicated temperatures.
A statistically significant difference (p<0.01) using a two-tailed unpaired t-test is indicated with an *. NG indicates that no growth was detected after one month of incubation.
Figure 5—figure supplement 1

(Panel A) Nucleotide and amino acid sequence of the N-terminal TAP (tandem affinity purification) tag inserted immediately upstream of the start codon of mcrG (locus tag, MA4547) using the Cas9-mediated genome editing technique.
(Panel B), Affinity purified TAP-tagged MCR from WT and the ∆ycaO-tfuA mutant. A fourth band, identified by as a hypothetical protein encoded by MA3997, co-purified along with the three MCR subunits in each case.
Figure 5—figure supplement 2
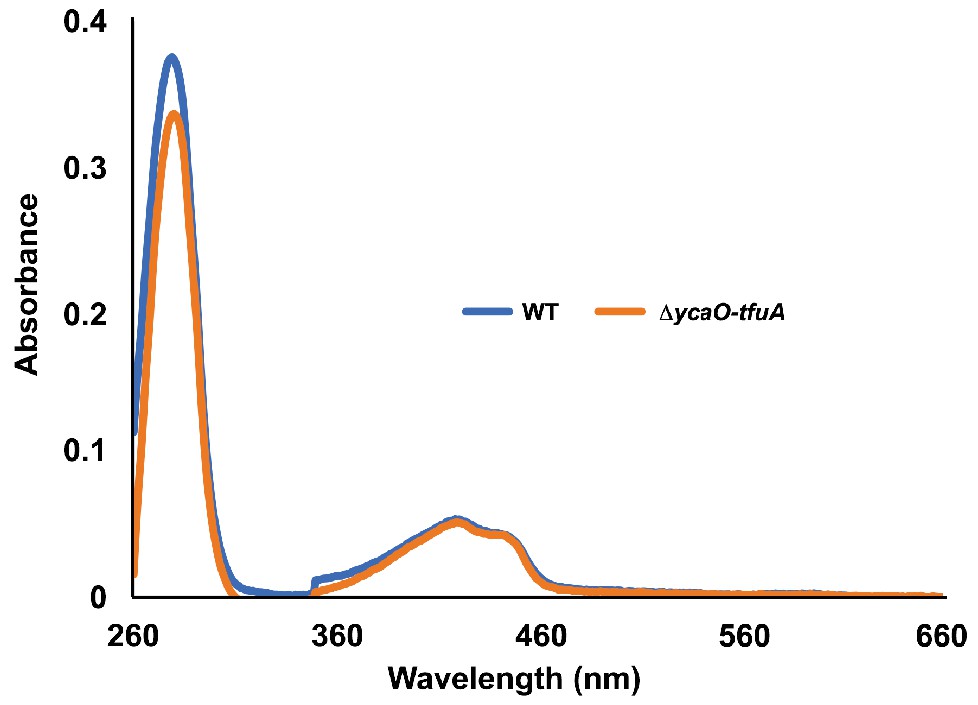
UV-Vis absorbance spectra of tagged-MCR purified from the WT strain (blue) and the ∆ycaO-tfuA mutant (orange).
The peaks at 280 nm and 430 nm respectively correspond to absorbances from the protein (Trp/Tyr) and F430. To facilitate comparison, each spectrum was normalized by setting the absorbance at 660 nm to 0.
Figure 6
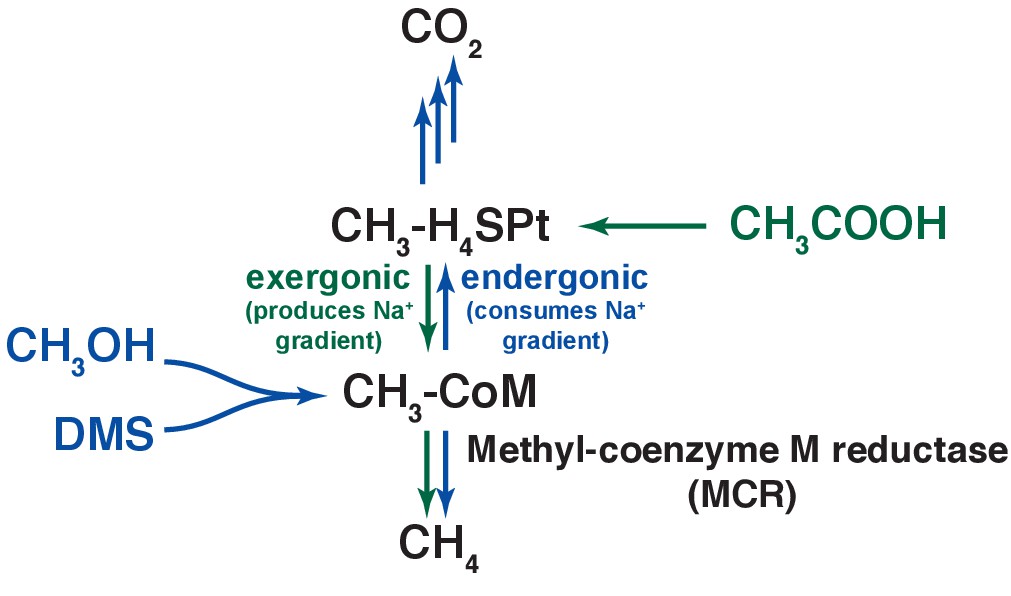
An overview of methanogenic metabolism in M. acetivorans.
Methylotrophic substrates such as methanol (CH3OH) or dimethylsulfide (DMS, CH3-S-CH3) enter the methanogenic pathway via S-methylation of coenzyme M (CoM) and subsequently disproportionate to methane (CH4) and carbon dioxide (CO2; metabolic flux is shown as blue arrows). Notably, the first step in oxidation of CH3-CoM to CO2 is the energy-requiring transfer of the methyl moiety to generate methyl-tetrahydrosarcinapterin (CH3-H4SPt). In contrast, acetic acid (CH3COOH) enters the pathway at the CH3-H4SPt level, followed by reduction to CH4 (green arrows). Thus, the second step of the pathway is exergonic.
Tables
Table 1
Growth phenotypes of M. acetivorans WT and the ∆ycaO-tfuA mutant on different substrates at 36°C.
https://doi.org/10.7554/eLife.29218.017| Substrate* | Strain | Doubling time (h)† | p-value | Maximum OD600† | p-value |
|---|---|---|---|---|---|
| Methanol | WT | 6.94 ± 0.41 | 0.87 | 5.35 ± 0.31 | 0.26 |
| ∆ycaO-tfuA | 6.89 ± 0.15 | 5.06 ± 0.39 | |||
| TMA | WT | 10.92 ± 0.54 | 0.84 | 9.57 ± 0.67 | 0.31 |
| ∆ycaO-tfuA | 10.98 ± 0.43 | 8.91 ± 0.73 | |||
| Acetate | WT | 54.23 ± 4.03 | 0.0002 | 0.44 ± 0.01 | 0.0005 |
| ∆ycaO-tfuA | 124.33 ± 12.52 | 0.35 ± 0.02 | |||
| DMS | WT | 31.27 ± 1.28 | <0.0001 | 0.60 ± 0.01 | <0.0001 |
| ∆ycaO-tfuA | 58.28 ± 1.45 | 0.44 ± 0.01 |
-
*TMA, trimethylamine; DMS, dimethylsulfide.
†The error represents the 95% confidence interval of the mean for three biological replicates and the p-values were calculated using two-tailed unpaired t-tests.
Additional files
-
Supplementary file 1
Lists of plasmids
- https://doi.org/10.7554/eLife.29218.019
-
Supplementary file 2
List of target sequences for Cas9-mediated genome editing
- https://doi.org/10.7554/eLife.29218.020
-
Supplementary file 3
List of strains
- https://doi.org/10.7554/eLife.29218.021
-
Transparent reporting form
- https://doi.org/10.7554/eLife.29218.022
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Post-translational thioamidation of methyl-coenzyme M reductase, a key enzyme in methanogenic and methanotrophic Archaea
eLife 6:e29218.
https://doi.org/10.7554/eLife.29218




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}