Rad52-Rad51 association is essential to protect Rad51 filaments against Srs2, but facultative for filament formation
- CEA, France
- Université Paris-Saclay, France
- Institut Gustave Roussy, CNRS UMR 8126, France
- Université Paris-Sud, France
Figures
Figure 1 with 2 supplements
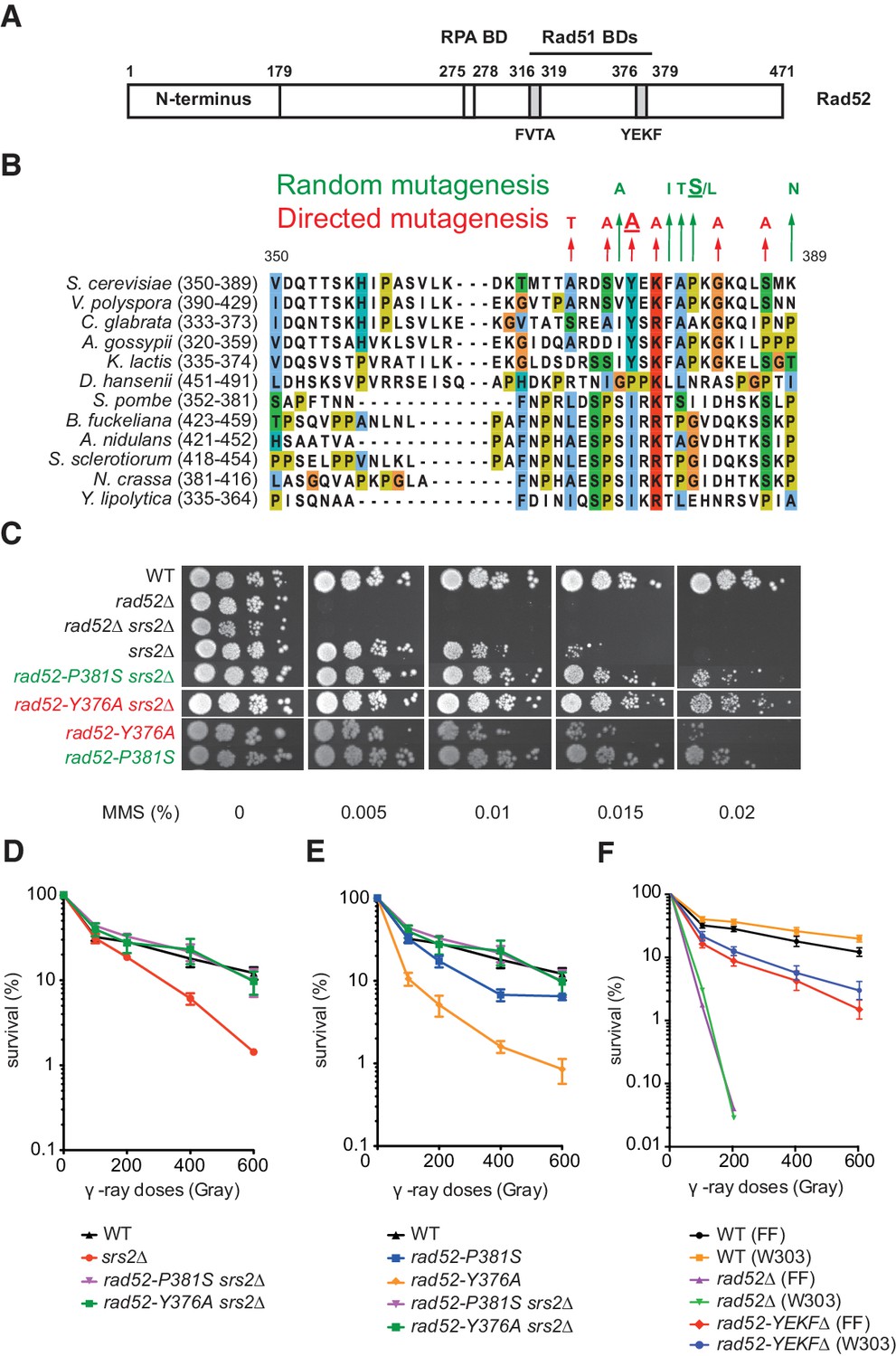
Mutations in the Rad51-binding domain of Rad52 suppress MMS and γ-ray sensitivity of Srs2-deficient cells.
(A) Primary structure of Rad52. The conserved N-terminus as well as the RPA and the Rad51 binding domains (BD) are shown. (B) The mutations selected from random and directed mutagenesis are indicated on the alignment of the region 350-389aa of S. cerevisiae Rad52 with that of Rad52 orthologs in 12 fungal species. The amino acid colors refer to the chemical character of the residues. The rad52-P381S and rad52-Y376A mutations are underlined. (C) Serial 10-fold dilutions of haploid strains with the indicated genotypes were spotted onto rich media (YPD) containing different MMS concentrations. The data presented are a selection from those in Figure 1—figure supplement 1. (D,E,F) Survival curves of haploid cells exposed to γ-ray. rad52-YEKF∆ cells were from two different genetic backgrounds: FF18733 (FF) and W303. Data are presented as the mean ± SEM of at least three independent experiments.
-
Figure 1—source data 1
Survival (%) of haploid cells exposed to γ-ray for Figure 1D,E and F.
- https://doi.org/10.7554/eLife.32744.005
Figure 1—figure supplement 1
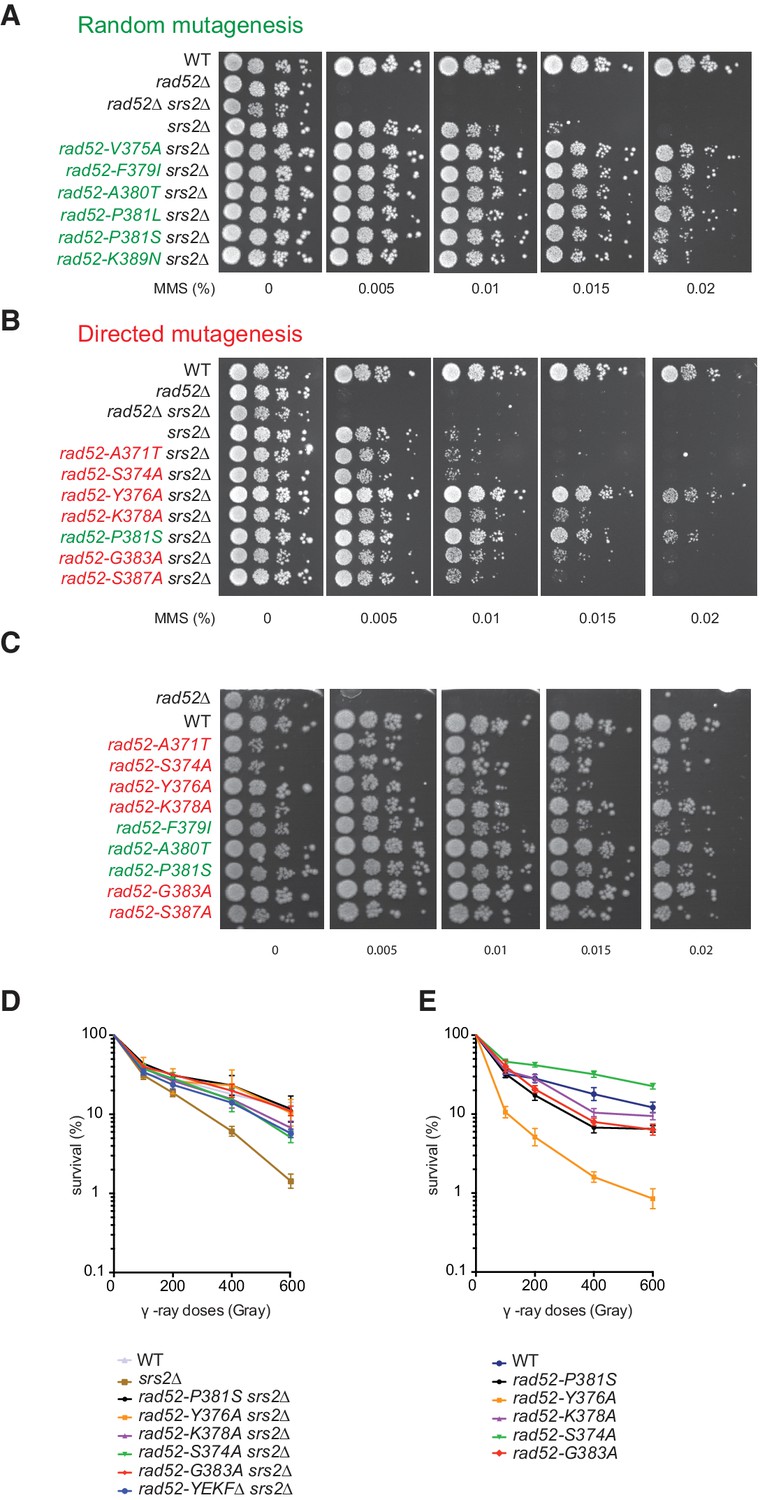
Mutations in the Rad51-binding domain of Rad52 suppress MMS and γ-ray sensitivity of Srs2-deficient cells.
(A,B,C) Serial 10-fold dilutions of haploid strains that were isolated by random mutation screening (green) or that harbor Rad52 mutations created by directed mutagenesis (red). The tested strains are derivative of rad52Δ or rad52Δ srs2Δ null mutants transformed with empty Ycplac111 centromeric plasmid or with wild type or mutated versions of RAD52. (D,E) Survival curves of haploid cells bearing mutations from our collection. Cells were in the log phase of growth when exposed to γ-rays. Data are presented as the mean ± SEM of at least three independent experiments. The rad52-A371T, rad52-S374A and rad52-S387A mutations cannot restore MMS resistance in Srs2-deficient cells, but they can reduce γ-ray sensitivity of Srs2-deficient cells. This difference could be explained by the constant formation of lesions in cells growing in MMS-containing plates, while exposure to γ-rays for a limited period of time creates only a definite number of lesions.
Figure 1—figure supplement 2

The rad52-Y376A, but not the rad52-P381S mutation suppresses the effect of mutations that are synthetically lethal with srs2Δ.
(A) Tetrad analysis of crosses between haploid rad52-P381S srs2Δ strains and haploid mutants that are synthetically lethal with srs2Δ. White diamonds indicate synthetic lethal combinations with or without rad52-P381S. This mutation does not affect the interaction between Rad52 and Rad51 enough to suppress the synthetic lethality. (B) Same crosses as in (A) but with rad52-Y376A. Double mutant spores, which do not contain rad52-Y376A, are indicated by white squares. The white circles mark viable triple mutants.
Figure 2
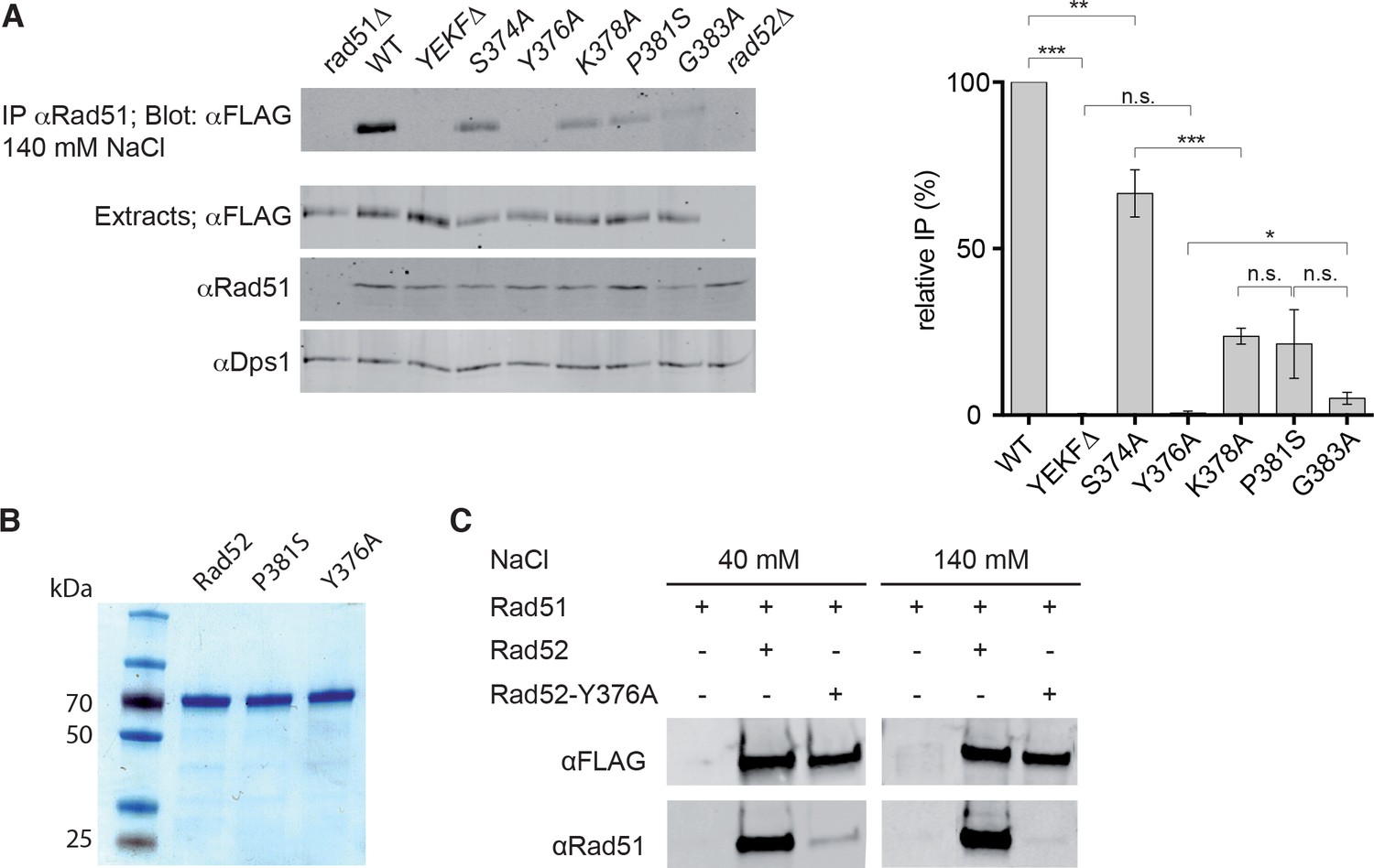
Measurement of Rad52-Rad51 interaction in cells that express various Rad52-FLAG mutants.
(A) Rad51 immunoprecipitation from cells that express different Rad52-FLAG mutants. The anti-Dps1 antibody was used as loading control. The signals corresponding to immunoprecipitated Rad52 mutants (Blot: αFLAG) were quantified relatively to the signal for WT Rad52, corrected to the quantity of Rad52 in each extracts (Extracts: αFLAG) and to the Dps1 signal (αDps1). Data are presented as the mean ± SEM (n = 3). Fisher’s exact test, n.s. p>0.05, *p<0.05, **p<0.01, ***p<0.001. (B) Purification of recombinant Rad52-FLAG, Rad52-P381S-FLAG and Rad52-Y376A-FLAG. (C) Pull-down experiments using Rad52-FLAG WT or Y376A fusion protein in the presence of different salt concentrations.
-
Figure 2—source data 1
Relative IP (%) and Two-tailed p values, unpaired T test for Figure 2A.
- https://doi.org/10.7554/eLife.32744.007
Figure 3

Effect of the rad52-P381S and rad52-Y376A alleles on HR.
(A) Recombination system allowing the formation of a WT ARG4 allele by gene conversion of the arg4-RV or arg4-Bg heteroalleles in diploid cells. (B, C) Survival curves (black) and heteroallelic HR frequencies (red) for the indicated homozygous diploid cells exposed to γ-rays. Data are presented as the mean ± SEM of at least three independent experiments. (D) HO-induced gene conversion between MAT ectopic copies. Cell viability after DSB formation is shown as the mean ± SEM of at least three independent experiments. Fisher’s exact test, n.s. p>0.05, *p<0.05.
-
Figure 3—source data 1
Source data for Figure 3B,C and D.
- https://doi.org/10.7554/eLife.32744.009
Figure 4 with 1 supplement
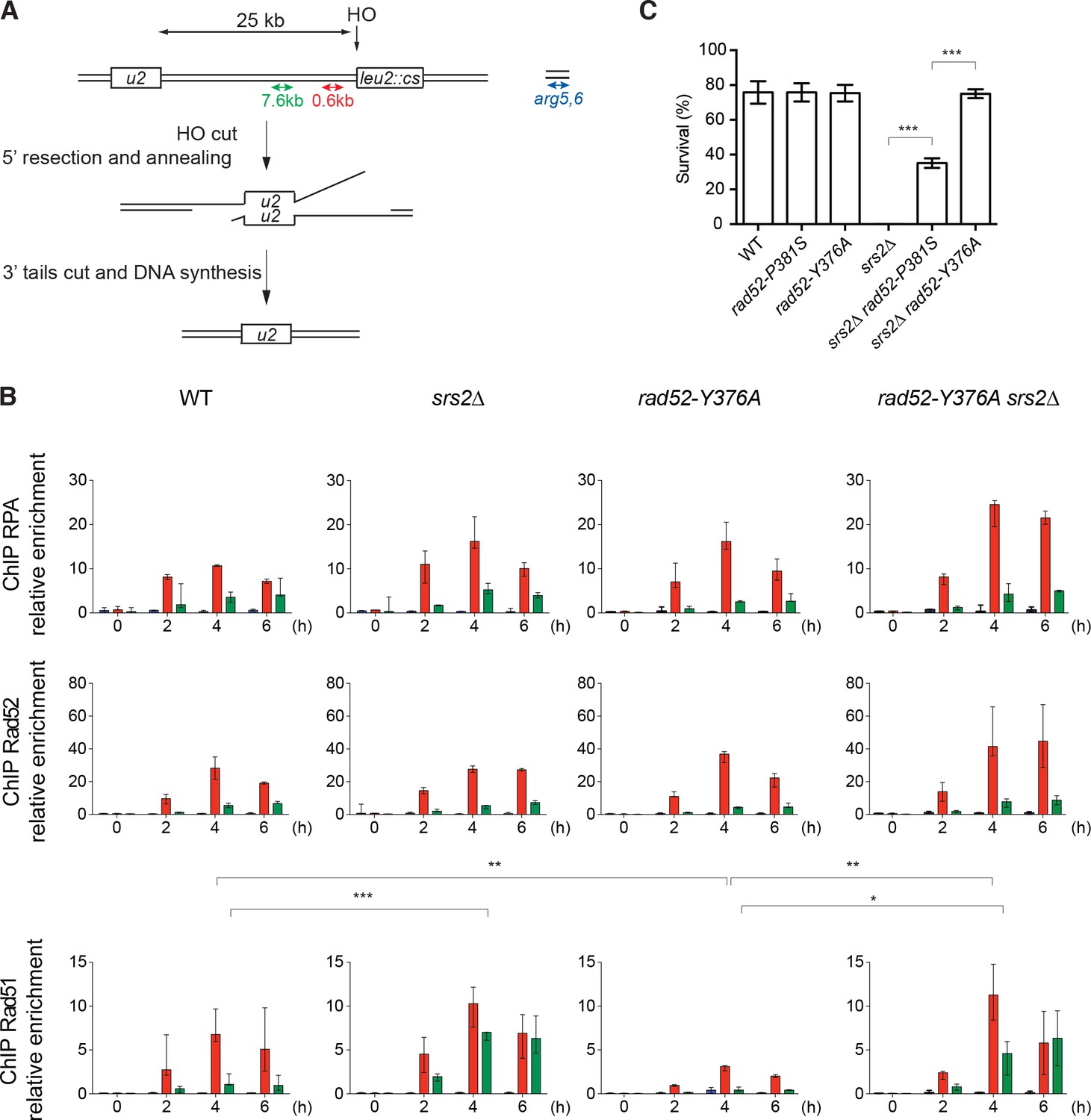
ChIP analysis of Rad51 filament formation at a DSB created by the HO endonuclease.
(A) Schematic of the HO-induced SSA repair system. (B) ChIP was used to assess RPA, Rad52 and Rad51 relative enrichment at 0.6 or 7.6 kb from the DSB site and at the uncut ARG5,6 locus at the indicated time points (hours) after HO induction. The median value (n ≥ 2) is shown and error bars represent the upper and lower values measured. Fisher’s exact test, *p<0.05, **p<0.01, ***p<0.001. Statistical analysis was performed 4 hr after HO-induction; only significant differences are shown. (C) Cell survival after HO-induced DSB formation. Cell viability after DSB formation is shown as the mean ± SEM of at least three independent experiments. Fisher’s exact test, ***p<0.001.
-
Figure 4—source data 1
Source data for Figure 4A.
- https://doi.org/10.7554/eLife.32744.013
Figure 4—figure supplement 1

ChIP analysis of Rad51 filament formation in rad52-P381S mutant cells that lack or not Srs2.
(A) Schematic representation of the HO-induced DSB repair system used. Breakage by HO can be repaired by single strand annealing (SSA) of two directed repeats located 25 kb apart. (B) ChIP analysis after HO induction in wild type (WT) or Srs2-deficient cells that express WT or Rad52-P381S-FLAG proteins. The median value of at least three experiments is shown and error bars represent the upper and lower values. Fisher’s exact test, *p<0.05, **p<0.01, ***p<0.001. Statistical analysis was performed at the 4 hr time point; only significant differences are shown. The rad52-P381S mutation did not significantly change Rad51 or Rad52 enrichment at 7.6 kb (green) and 0.6 kb (red) from the DSB, but a 1.5-fold significant increase in RPA recruitment was observed compared with WT Rad52. This increase could be related to an enrichment of ssDNA coated with both RPA and Rad51. This higher RPA enrichment is dependent on Srs2 presence because RPA loading is reduced by 1.8-fold in rad52-P381S srs2Δ cells compared with rad52-P381S cells. This is associated with a 2.5-fold increase in Rad51 loading at 0.6 kb from the DSB site in rad52-P381S srs2Δ cells compared with rad52-P381S cells. This suggests that Srs2 removes significantly more Rad51 in rad52-P381S cells. Similarly, Rad51 loading at the 7.6 kb site was increased by 3.4-fold in rad52-P381S srs2Δ cells compared with rad52-P381S cells. Therefore, Rad51 filaments assembled by Rad52-P381S are sensitive to Srs2 activity regardless of the distance from the break, confirming that Rad52 association with Rad51 protects Rad51 filaments from Srs2. Interestingly, the recruitment of Rad51 is increased specifically in rad52-P381S srs2∆ cells compared with rad52-P381S cells and srs2∆ cells for a reason we do not understand.
-
Figure 4—figure supplement 1—source data 1
Source data for Figure 4—figure supplement 1.
- https://doi.org/10.7554/eLife.32744.012
Figure 5 with 3 supplements

Biochemical analysis of Rad52-P381S and Rad52-Y376A mutant proteins.
All experiments were performed with FLAG-tagged Rad52 proteins. (A) EM analysis of Rad51 filament formation. A representative nucleofilament issued from each reaction is shown. Addition of Rad51 prior to RPA on circular φx174 ssDNA (Rad51 RPA) forms complete Rad51 filaments, whereas addition of RPA before Rad51 (RPA Rad51) results in partial Rad51 filaments. White arrows indicate the presence of RPA on ssDNA. Concomitant addition of Rad52 with Rad51 in this reaction (RPA Rad51 Rad52) leads to complete Rad51 filaments associated with Rad52 (white arrows). The percentage of each molecular species is plotted on the histogram (right panel), showing the weak effect of Rad52 mutants on Rad51 filament formation. Two independent experiments were performed with very similar results. The results of the individual biological replicates were pooled (300 molecules analyzed). (B) Number of Rad52, Rad52-P381S and Rad52-Y376A molecules associated with Rad51 filaments assembled at two NaCl concentrations (n = 40). The median value is represented by the horizontal bar. Fisher’s exact test, ***p<0.001. (C) Effect of Rad52 mutants on strand exchange. The standard reaction (Rad51 RPA) shows that complete Rad51 filaments efficiently catalyze the formation of nicked circular products. Pre-bound RPA inhibits this reaction (RPA Rad51); however, the inhibitory effect of pre-bound RPA is overcome by Rad52 (RPA Rad51 Rad52) and, to a lesser extent, by the Rad52 mutants tested. The percentage of nicked circular product calculated by quantification of the gels is shown. Data are presented as the mean ± SEM of at least three independent experiments. Fisher’s exact test, n.s. p>0.05, *p<0.05, **p<0.01, ***p<0.001. (D) EM analysis of Rad51 filament displacement by Srs2. Srs2 was added for 5 min at the end of the Rad51 filament formation reactions (Rad51 RPA Srs2 and RPA Rad51 Rad52 Srs2). The white arrow shows the appearance of RPA on Rad52-Y376A-assembled Rad51 filaments after Srs2 addition. The percentage of each molecular species is shown in the histogram. Two independent experiments were performed with very similar results. The results of the individual biological replicates were pooled (200 molecules analyzed). (E) Rad52 increases strand exchange efficiency in the presence of Srs2. Strand exchange reactions were performed as in (C) except that increasing concentrations of Srs2 were added with the linear dsDNA for 90 min. Data are shown as the mean ± SEM of at least three independent experiments. (F) Pull-down assay with an anti-FLAG antibody shows the interaction between Rad52 and Srs2.
-
Figure 5—source data 1
Source data for Figure 5E.
- https://doi.org/10.7554/eLife.32744.020
Figure 5—figure supplement 1
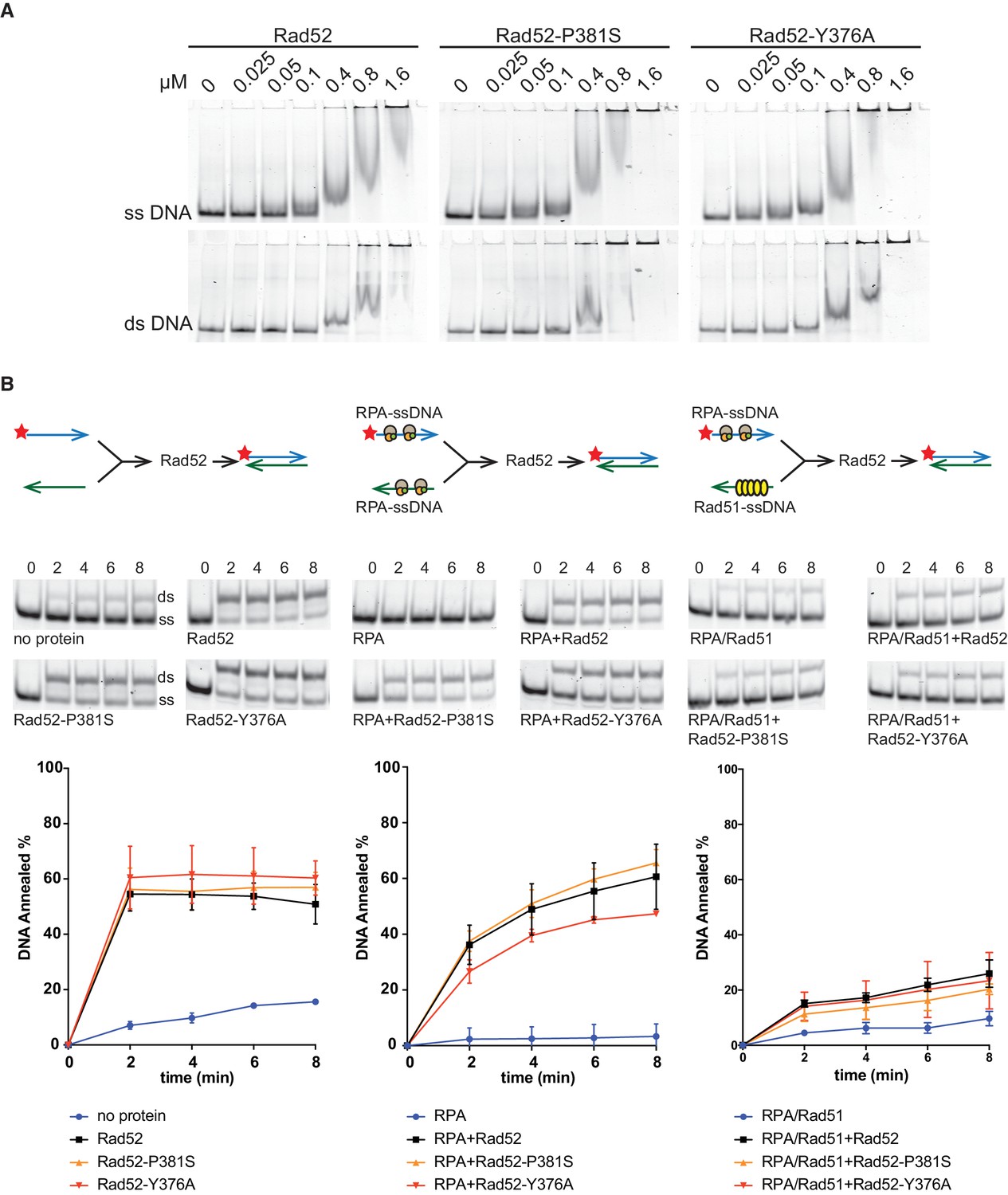
DNA binding and strand annealing activity of Rad52 mutants.
(A) Binding of purified Rad52-FLAG, Rad52-P381S-FLAG and Rad52-Y376A-FLAG to ssDNA and dsDNA (protein/base ratio: 1/10, 1/5, 1/2.5, 1/1.25, 1/0.6, and 1/0.15). (B) Rad52-FLAG, Rad52-P381S-FLAG and Rad52-Y376A-FLAG annealing activities are similarly inhibited by RPA and Rad51. Experiments are represented schematically at the top of the panels. Reactions were performed with labeled 48bp-long complementary primers. Each oligonucleotide was incubated without proteins, with 30 nM RPA or with 134nM Rad51 before addition of 40 nM of wild type (WT) Rad52 or mutant proteins. Samples were separated on 8% native TBE polyacrylamide gels. The ratio between annealed DNA and the sum of ssDNA and dsDNA was plotted. Data are presented as the mean ± SEM of at least three independent experiments.
-
Figure 5—figure supplement 1—source data 1
DNA annealed (%) for Figure 5—figure supplement 1B.
- https://doi.org/10.7554/eLife.32744.016
Figure 5—figure supplement 2
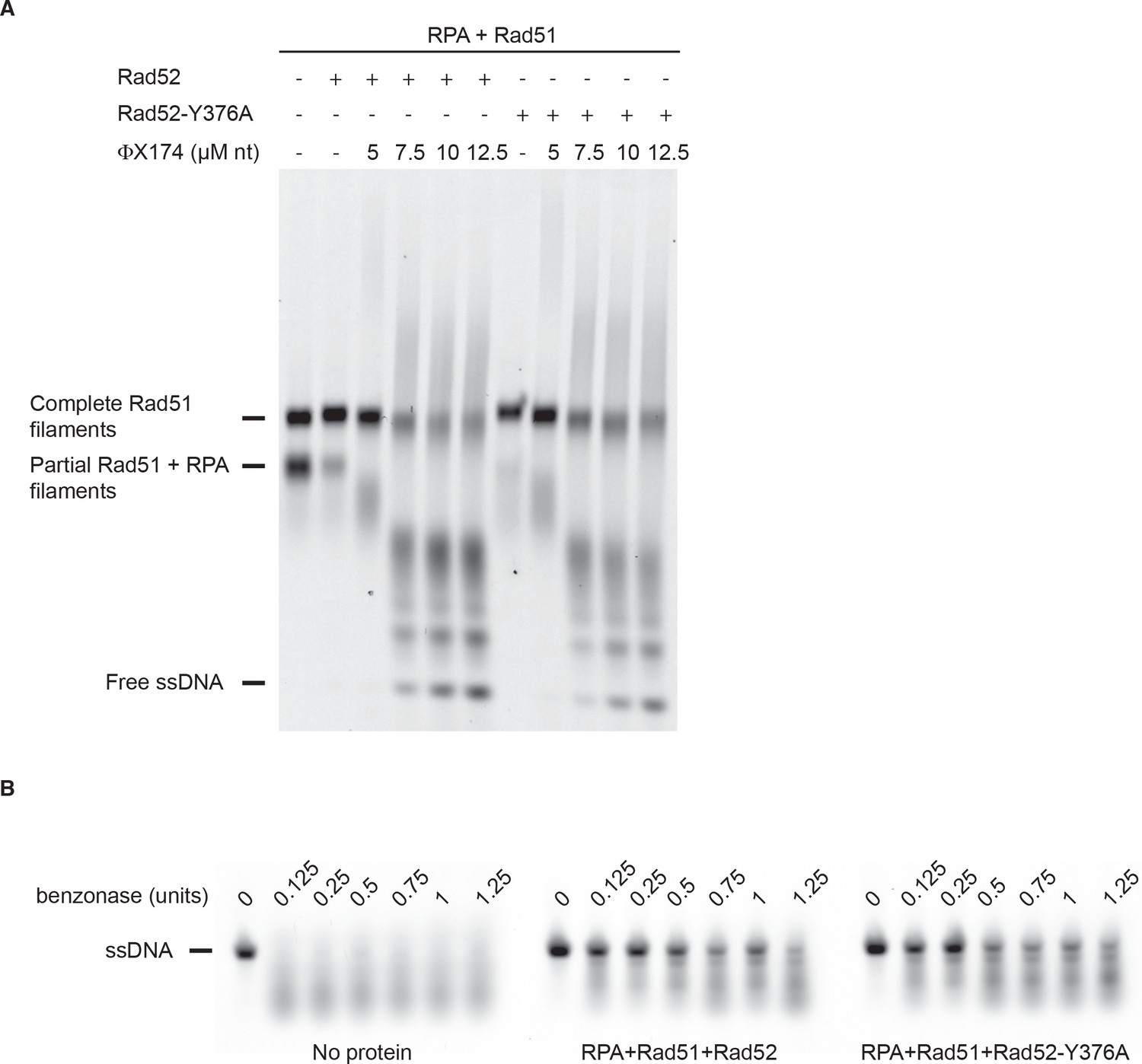
Rad51 filament stabilization by Rad52.
All experiments were performed with FLAG-tagged Rad52 proteins. (A) Rad51 filament stability was challenged with excess ssDNA. The addition of Rad52 or Rad52-Y376A together with Rad51 to RPA-coated 400-nucleotide-long ssDNA strongly catalyzes the formation of complete Rad51 filaments. Addition of increasing amounts of competitor PhiX174 ssDNA after Rad51 filament formation leads to their destabilization. (B) Benzonase assay. Rad51 was assembled as in (A) with Rad52 or Rad52-Y376A and increasing amount of benzonase was added to the reaction. ssDNA was loaded on agarose gel after deproteinization. As a control, naked ssDNA was also incubated with benzonase.
Figure 5—figure supplement 3
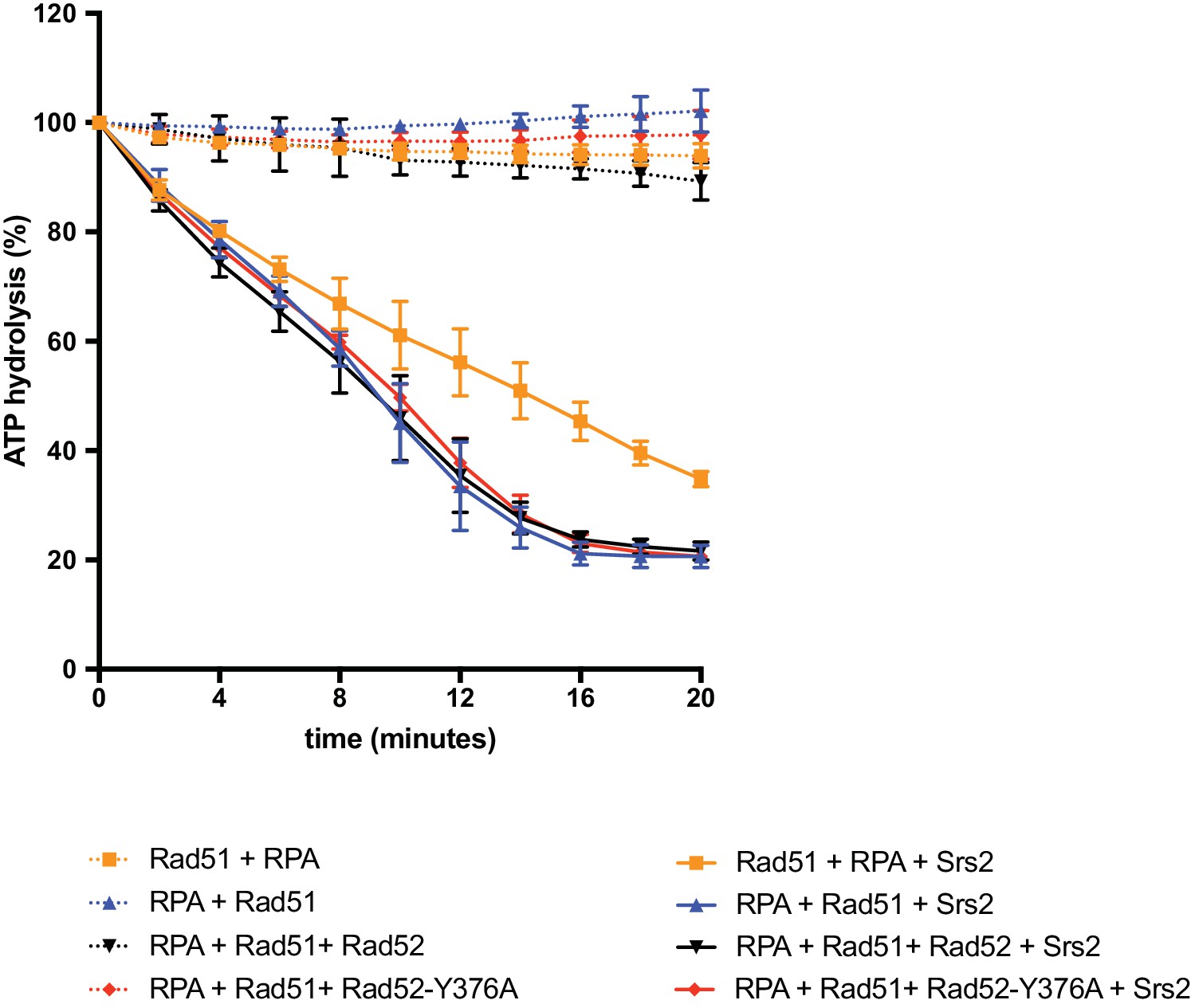
Measurement of Srs2 ATPase activity.
Rad51 filament formation reactions were performed as described in the Materials and methods section and the main text. ATP hydrolysis was measured as a function of time after Rad51 filament formation and addition (or not) of 50 nM Srs2. Data are presented as the mean ± SEM of at least two independent experiments.
-
Figure 5—figure supplement 3—source data 1
ATP hydrolysis (%) for Figure 5—figure supplement 3.
- https://doi.org/10.7554/eLife.32744.019
Figure 6
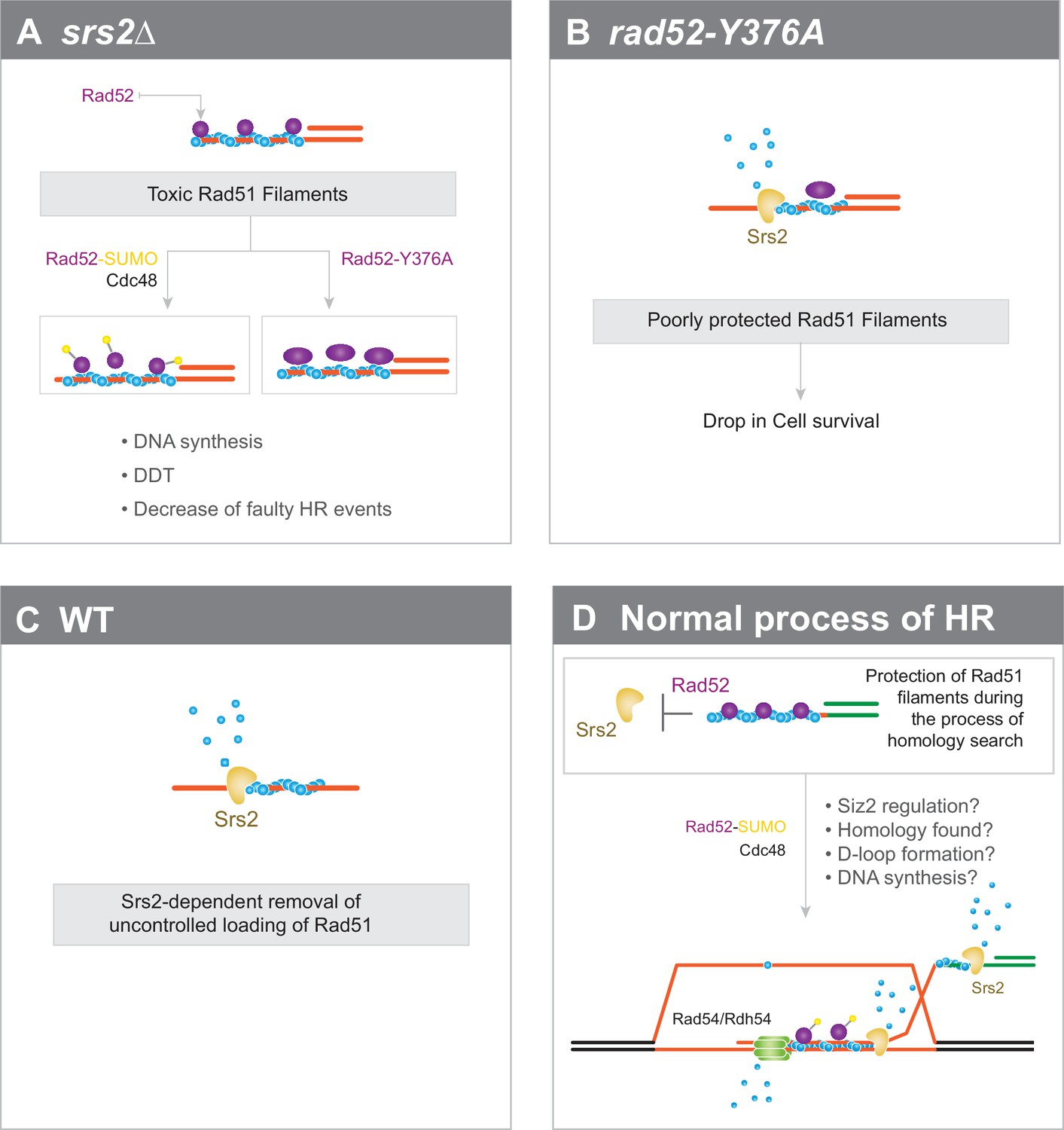
Interplay between Rad52 sumoylation and Srs2 in the control of Rad51 filament formation.
(A) In Srs2-deficient cells, the association of Rad52 with Rad51 filaments is responsible for their potential toxicity. Massive sumoylation of Rad52 could trigger the Cdc48-dependent release of Rad52, as do mutations that affect the interaction between Rad52 and Rad51. Excessive stabilization of Rad51 filaments by Rad52 could block DNA synthesis or DNA damage tolerance (DTT) events and promote faulty HR events. (B) Impaired interaction between Rad52 and Rad51 would sensitize Rad51 filaments to Srs2 activity. (C) In WT cells, uncontrolled loading of Rad51 might lead to the production of toxic Rad51 filaments that are efficiently removed by Srs2. (D) Global view of Rad51 filament regulation during HR (see text for more details).
Tables
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Gene (Saccharomyces cerevisiae) | RAD52 | NA | SGD: YML032C | |
| Gene (S. cerevisiae) | RAD51 | NA | SGD: YER095W | |
| Gene (S. cerevisiae) | Srs2 | NA | SGD: YJL092W | |
| Genetic reagent (Bacteriophage φ-X174) | viral (+) strand of φX174 DNA | |||
| Strain, strain background (S. cerevisiae) | FF18733 | Cell line maintained in E. Coïc's lab | ||
| Strain, strain background (S. cerevisiae) | YFP17; JKM146 | Cell line maintained in J. Haber's lab | ||
| Antibody | anti-Rad51 (rabbit polyclonal) | Abcam | ab63798 | |
| Antibody | anti-FLAG (Mouse polyclonal) | Sigma | F3165 | |
| Antibody | anti-DPS1 (Rabbit polyclonal) | S. Marcand's lab | ||
| Antibody | anti-RPA (Rabbit polyclonal) | V. Géli's lab | ||
| Antibody | anti-Mouse IR800 Goat monoclonal) | Advansta | R-05061–250 | |
| Antibody | anti-Rabbit IR700 Goat monoclonal) | Advansta | R-05054–250 | |
| Antibody | anti-Mouse IR700 Goat monoclonal) | Advansta | R-05055–250 | |
| Antibody | anti-Srs2 (Goat polyclonal) | Santa-Cruz | sc-11991 | |
| Recombinant DNA reagent | YCplac111-Rad52-6His-3FLAG | This study | progenitors: YCplac111 (GenBank: X75457, L26350) | |
| Recombinant DNA reagent | Yiplac211-rad52-P381S; Yiplac211-rad52-Y376A | This study | progenitors: Yiplac211 (GenBank: X75462, L26358) | |
| Recombinant DNA reagent | pET15b-Rad52-FLAG | Progenitor: pET15b (addgene: 69661-3) | ||
| Peptide, recombinant protein | Rad52-FLAG | This study | ||
| Peptide, recombinant protein | RPA | This study | ||
| Peptide, recombinant protein | Rad51 | This study | ||
| Peptide, recombinant protein | Srs2 | This study | ||
| Peptide, recombinant protein | 3XFLAG | Sigma | F4799 | |
| Commercial assay or kit | GeneMorph II EZClone Domain Mutagenesis | Agilent Technologies | 200552–5 | |
| Commercial assay or kit | Dynabeads coupled to Protein A | Invitrogen | 11041 | |
| Commercial assay or kit | Dynabeads coupled to PanMouse | Invitrogen | 10002D | |
| Commercial assay or kit | Platinum SYBR Green qPCR SuperMix-UDG | Invitrogen | 11733–046 | |
| Software, algorithm | PSI-Blast | Altschul et al. (1997) | ||
| Software, algorithm | Muscle | Edgar (2004) | ||
| Software, algorithm | Jalview | Waterhouse et al. (2009) | ||
| Software, algorithm | iTEM software | Olympus, Soft Imaging Solutions | ||
| Software, algorithm | ImageQuant TL software | GE Healthcare Life Sciences |
Additional files
-
Supplementary file 1
RAD52 mutations selected by random and directed mutagenesis.
- https://doi.org/10.7554/eLife.32744.022
-
Supplementary file 2
S. cerevisiae strains.
- https://doi.org/10.7554/eLife.32744.023
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Rad52-Rad51 association is essential to protect Rad51 filaments against Srs2, but facultative for filament formation
eLife 7:e32744.
https://doi.org/10.7554/eLife.32744




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}