Two consecutive microtubule-based epithelial seaming events mediate dorsal closure in the scuttle fly Megaselia abdita
- Centre for Genomic Regulation (CRG), The Barcelona Institute of Science and Technology, Spain
- Universitat Pompeu Fabra, Spain
- Konrad Lorenz Institute for Evolution and Cognition Research (KLI), Austria
Figures
Figure 1 with 4 supplements
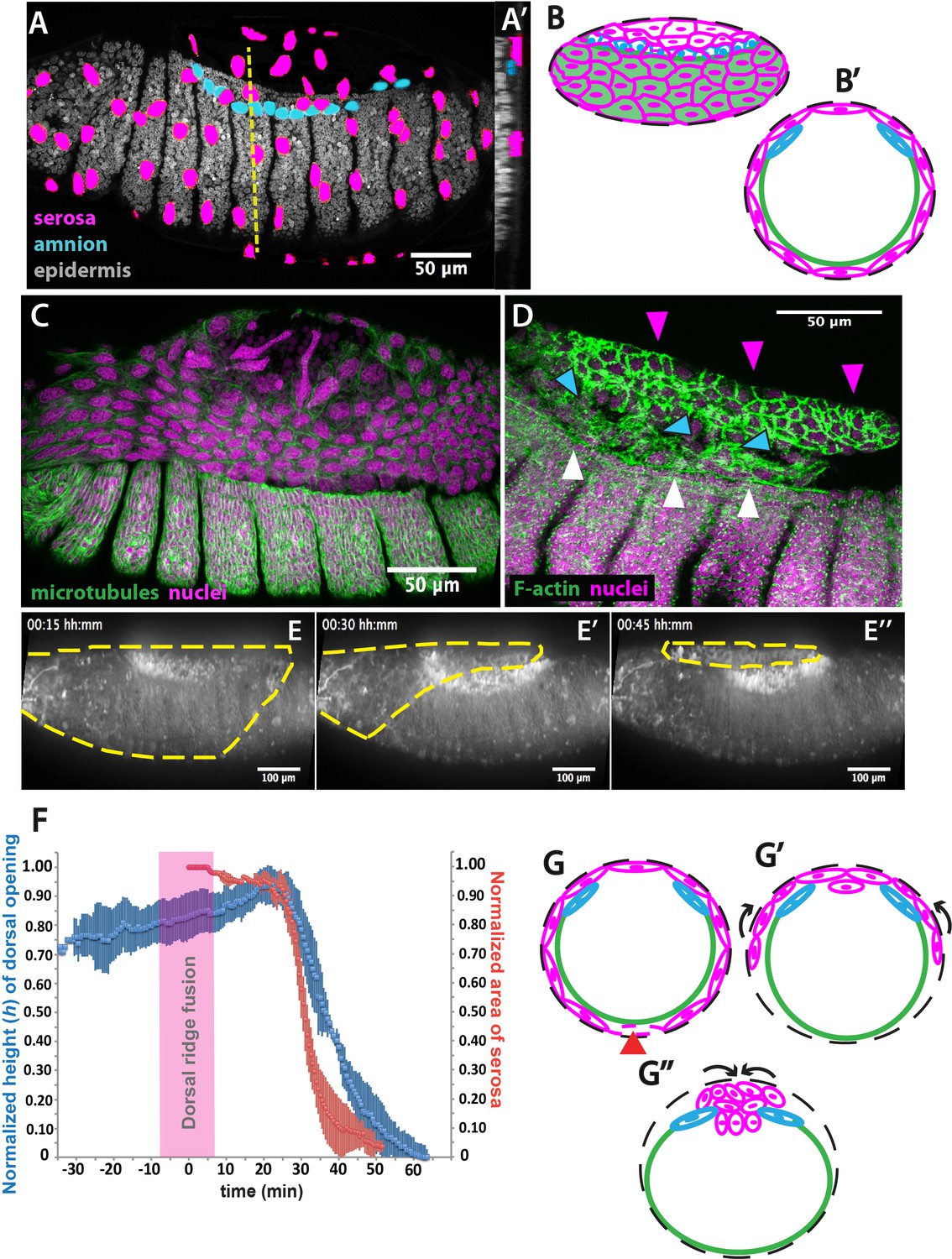
The extraembryonic serosa ruptures and accumulates dorsally previous to epidermal seaming in Megaselia abdita.
(A) Nuclear staining of M. abdita embryos prior to dorsal closure reveals three types of tissues: the extraembryonic serosa (magenta), the extraembryonic amnion (blue), and the embryonic epidermis (gray). (A’) An orthogonal re-sliced stack along the dashed yellow line in A shows the position of the embryonic (gray), amniotic (blue), and serosal (magenta) tissues in transverse view. (B) Schematics depicting the organization of the serosa cells (magenta), amnion cells (blue), and embryonic epidermis (green) in lateral and (B’) transverse view. The black dashed line represents the vitelline envelope (C) M. abdita embryo undergoing rupture and retraction of serosal tissue along the ventral side. Staining against β-tubulin in green, and DAPI nuclear counterstain in magenta. (D) Serosal cells (magenta arrowheads) accumulate on the dorsal side of the embryo after rupture. The serosa remains apposed to the amnion (blue arrowheads), which is in turn apposed to the embryonic epidermis (white arrowheads). Phalloidin stain in green and DAPI nuclear counterstain in magenta. (E) Images from a time-lapse sequence of serosa retraction in a M. abdita embryo injected with the fluorescent lipophilic dye FM 4–64 (from Video 1). Yellow dashed line shows the contour of the serosa covering the embryo during retraction. (F) Relative changes in area of the serosa during retraction (red, n = 15 embryos), and relative changes in height (h) of the dorsal opening (blue, n = 15 embryos) during dorsal closure in M. abdita. Vertical bars represent standard deviation (SD). Time range of dorsal ridge fusion is represented by pink area (n = 15 embryos) as a landmark for the initiation of dorsal closure. The origin of the time axis (T = 0 min) is set at the point of serosa rupture. (G) Schematics depict transverse views of embryos during serosa rupture, retraction, and dorsal accumulation. Serosal cells rupture along the ventral end of the embryo (red arrowhead). (G’) The remaining lateral serosa cells retract towards the dorsal side (black arrows). (G’’) Serosa cells continue retracting until they completely accumulate onto the dorsal opening (black arrows). Color scheme as in B. In all embryos and schematics, dorsal is to the top. Embryos in A, C, D and E show lateral views where anterior is to the left.
Figure 1—figure supplement 1
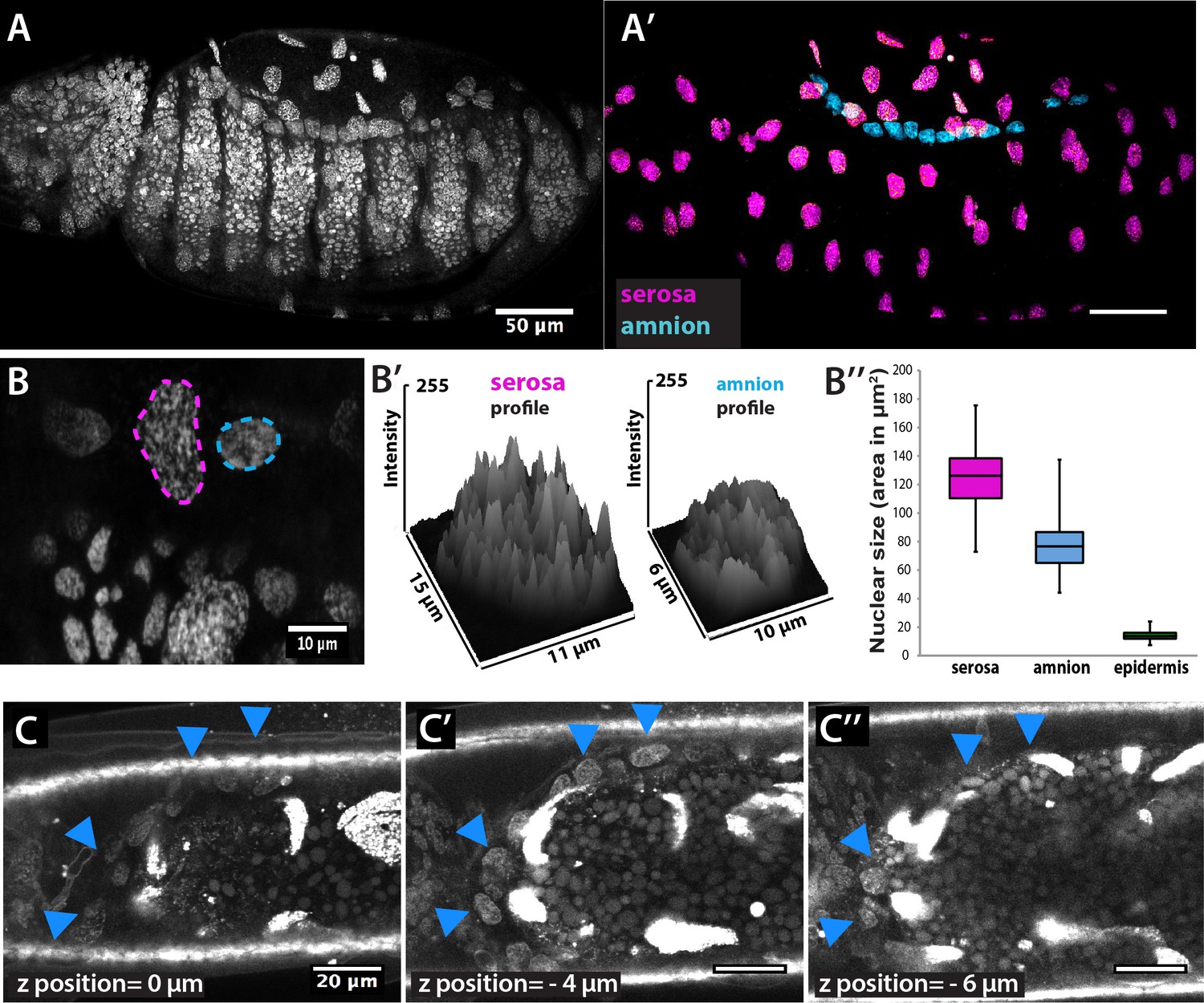
Identification of extraembryonic tissues in Megaselia abdita.
(A) Confocal projection of nuclear staining in an intact M. abdita embryo stained with DAPI. (A’) Confocal projection and image processing of nuclear staining from A to separate the extraembryonic serosa (magenta) and extraembryonic amnion (blue) from the embryonic epidermis. (B) Confocal projection to observe the discontinuous DAPI staining in serosa cells (magenta dashed line) and continuous DAPI staining in amnion cells (blue dashed line). (B’) Plotted intensity profiles of the serosa and amnion showing the discontinuous and continuous DAPI stainings, respectively. (B’’) Nuclear size (measured as area in µm2) of serosa cells (average size 125± 21 µm2, SD, n = 150 cells), amnion cells (average size 77 ± 16 µm2, SD, n = 150 cells) and epidermal cells (average size 14 ± 3 µm2, SD, n = 150 cells). (C–C’’) Dorsal view of three sequential z-stack confocal images (0,–4 and −6 µm) in a live M. abdita embryo injected with DAPI and showing the localization of amnion cells on yolk granules and adjacent to the epidermis (blue arrowheads). In all embryos, anterior is to the left.
Figure 1—figure supplement 2

Description of the three-tissue system anatomy in Megaselia abdita prior to dorsal closure.
(A) Nuclear staining (DAPI) of serosa cells (magenta) attached to the vitelline membrane after devitellinization. (B) Phalloidin staining (green) of a devitellinized embryo (dorsal view) showing a gap on the dorsal side of the embryo left by serosa removal with intact embryonic epidermis and adjacent amnion cells (blue arrowheads). (C) Lateral view of a devitellinized embryo where a couple of serosa cells (white arrowhead) and amnion cells (blue arrowhead) remained intact after removal of the vitelline membrane. This embryo corresponds to an early stage of dorsal closure prior to serosa rupture (T≈0 min) as indicated by large intact serosa cells, the fused dorsal ridge at the anterior end of the dorsal opening, a straightened epidermal leading edge and the large height (h) of the dorsal opening. (C’) Higher magnification of the embryo in D at the site where extraembryonic serosa (white arrowhead) and amnion (blue arrowhead) remained intact upon devitellinization. (C’’) Orthogonal stack re-slicing along the white dashed line in D showing a cross-section of the three-tissue anatomy of M. abdita dorsal closure where amnion cells (blue arrowheads) and serosa cells (white arrowheads) are apposed (yellow arrowhead). In embryos from A to C’ anterior is to the left. In embryos from C to C’’, dorsal is to the top.
Figure 1—figure supplement 3
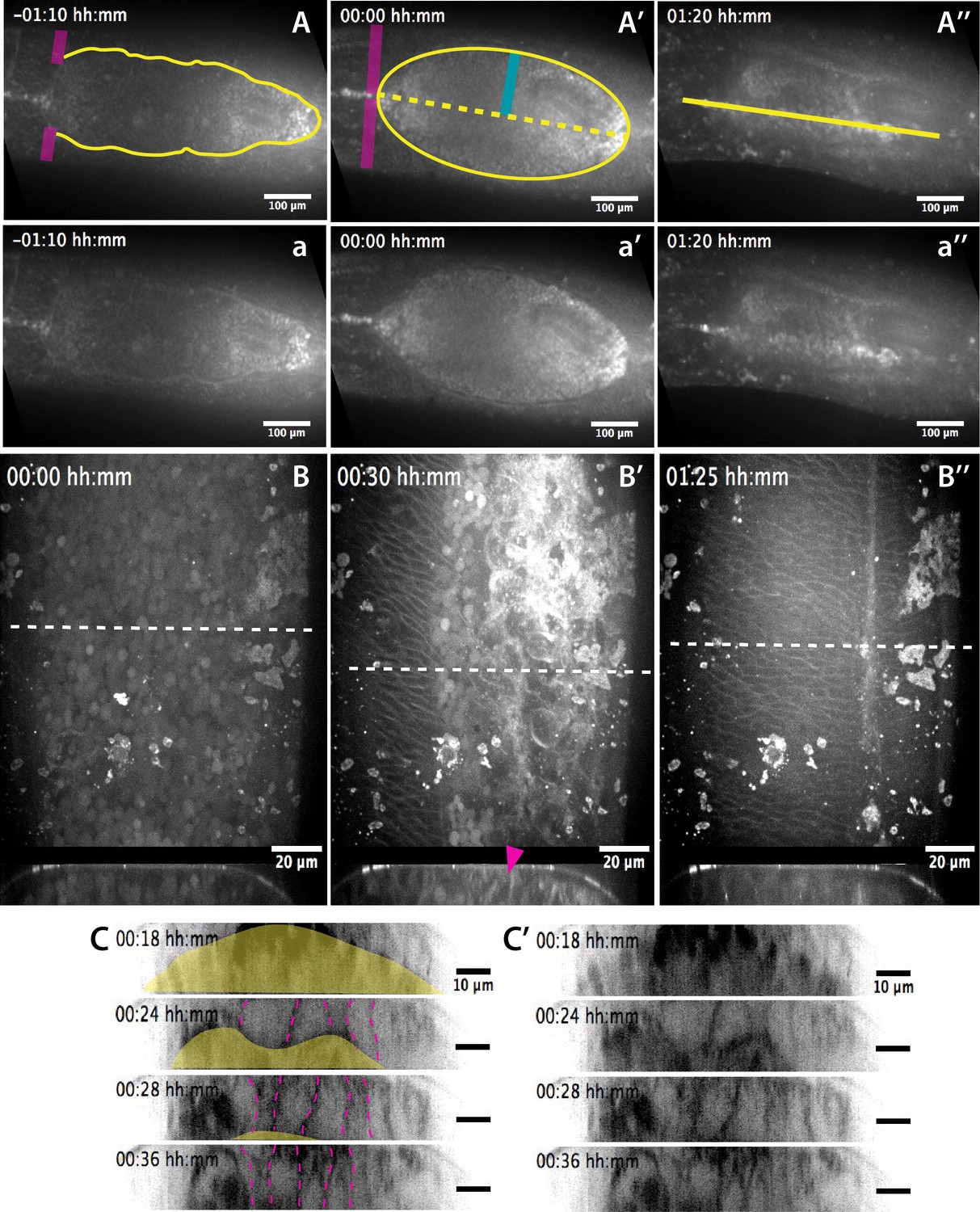
Anatomical landmarks during dorsal closure progression and serosa internalization.
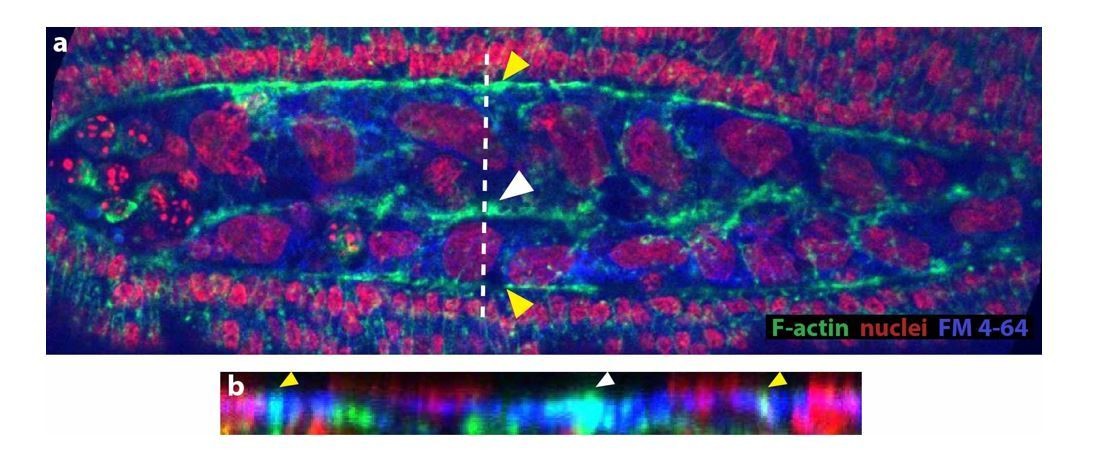
(A–A’’) Dorsal views from a time-lapse sequence of dorsal closure in a M. abdita embryo labeled with the fluorescent dye FM 4–64. Magenta bands depict the position and fusion of the dorsal ridge during the early and mid phase of dorsal closure. Yellow solid lines show the scalloped perimeter of the dorsal opening during early phase (A), the straightened perimeter during mid phase (A’), and the seamed epidermis during late phase (A’’) of dorsal closure. Yellow dotted line shows the dorsal midline and the blue solid line depicts the measured height (h) of the dorsal opening during dorsal closure. (a–a’’) Raw images from A-A’’ without superimposed labels. In all embryos, anterior is to the left. (B–B’') Dorsal (top) and orthogonal (bottom) views from a time-lapse sequence of a FM 4–64-labeled M. abdita embryo during dorsal closure at mid (B) and late (B’–B’’) phase of dorsal closure (from Video 2). The magenta arrow in bottom panel indicates the extraembryonic serosa cells undergoing apicobasal elongation towards the yolk and undergoing internalization. Anterior is to the top in dorsal view and dorsal is to the top in orthogonal view. (C–C’) Time-lapse images (from top panel of Video 7) showing the apicobasal elongation of the serosa during its retraction and internalization. In (C), the yellow area indicates the yolk and the magenta dashed lines highlight serosal apico-basal junctions. (C’) represents the same sequence without labels.
Figure 1—figure supplement 4
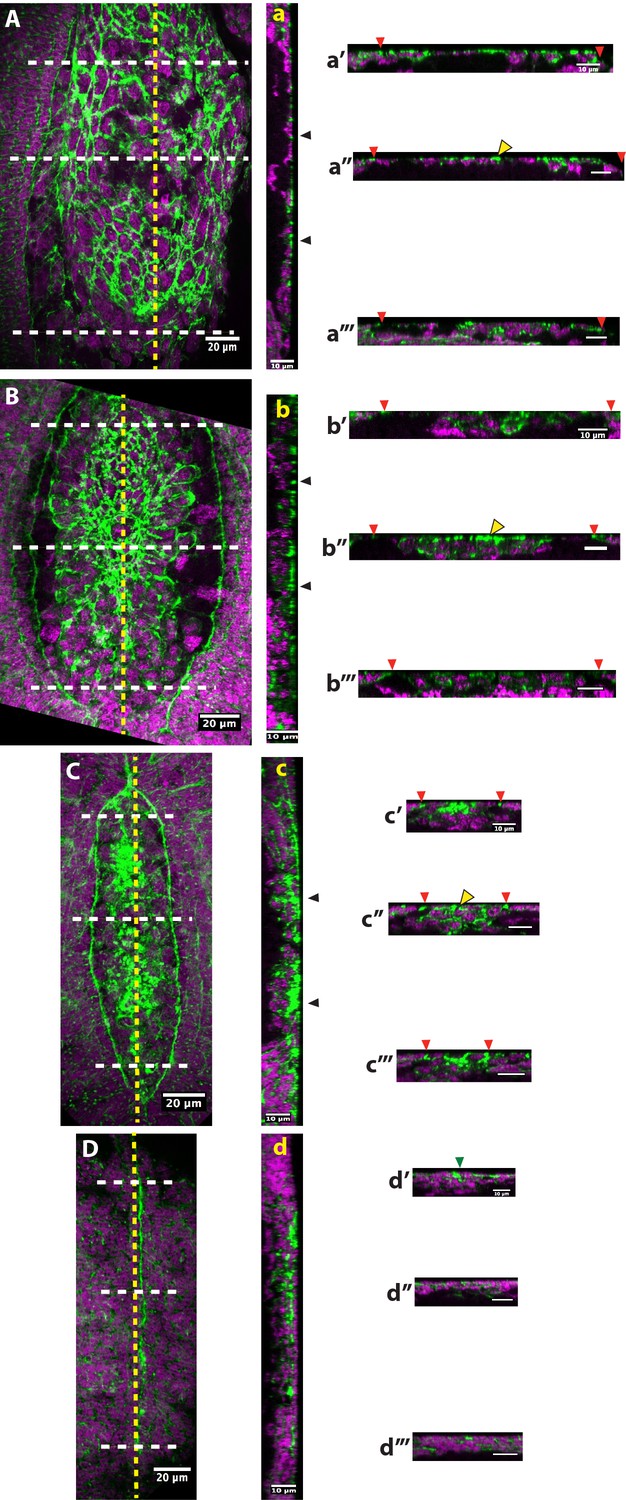
Tissue topology at anterio-posterior positions of different stages of dorsal closure in fixed embryos of Megaselia abdita.
(A) Dorsal view of a M. abdita embryo during serosa dorsal accumulation. (a) A stack reslicing along the anterio-posterior (AP) midline (yellow dashed line) shows a sagittal view of the embryo during serosa dorsal accumulation (black arrowheads). Orthogonal stack reslicing at (a’) anterior, (a’’) mid and (a’’’) posterior positions along the AP midline (white dashed lines) showing optical cross section of the embryo during dorsal accumulation of serosa cells. Red arrowheads show the position of the epidermal leading edge. (B) Dorsal view of a M. abdita embryo during the initial steps of serosa internalization. (b) Sagittal view along AP midline (yellow dashed line) of the embryo during serosa internalization (black arrowheads). Cross-sections at (b’) anterior, (b’’) mid and (b’’’) posterior positions along the AP midline (white dashed lines) showing the internalization of serosa cells. (C) Dorsal view of a M. abdita embryo during amnion seaming at the dorsal midline. (c) Sagittal view along the AP midline (yellow dashed line) of the embryo with serosa cells fully internalized (black arrowheads). Cross sections at (c’) anterior (close to canthus), (c’’) mid and (c’’’) posterior (close to canthus) positions along the AP midline (white dashed lines) showing fully internalized serosa cells underneath the amnion. (D) Dorsal view of a M. abdita embryo at the completion of dorsal closure. (d) Sagittal view along the AP midline (yellow dashed line) of the embryo after epidermal seaming has been completed and extraembryonic serosa and amnion have been internalized. Cross-sections at (d’) anterior, (d’’) mid and (d’’’) posterior positions along the AP midline (white dashed lines) showing a continuous epidermal layer. Green arrowhead shows the position of epidermal fusion in d’. Note that F-actin accumulates preferentially at the apical part of contracting serosa cells (yellow arrowhead) during internalization in a’’, b’’ and c’’. Phalloidin staining in green and DAPI counterstain in magenta. In dorsal and sagittal views, anterior is to the top and posterior to the bottom. In sagittal views, anterior to the top and dorsal to the right. In cross-sections, dorsal is to the top.
Figure 2 with 1 supplement
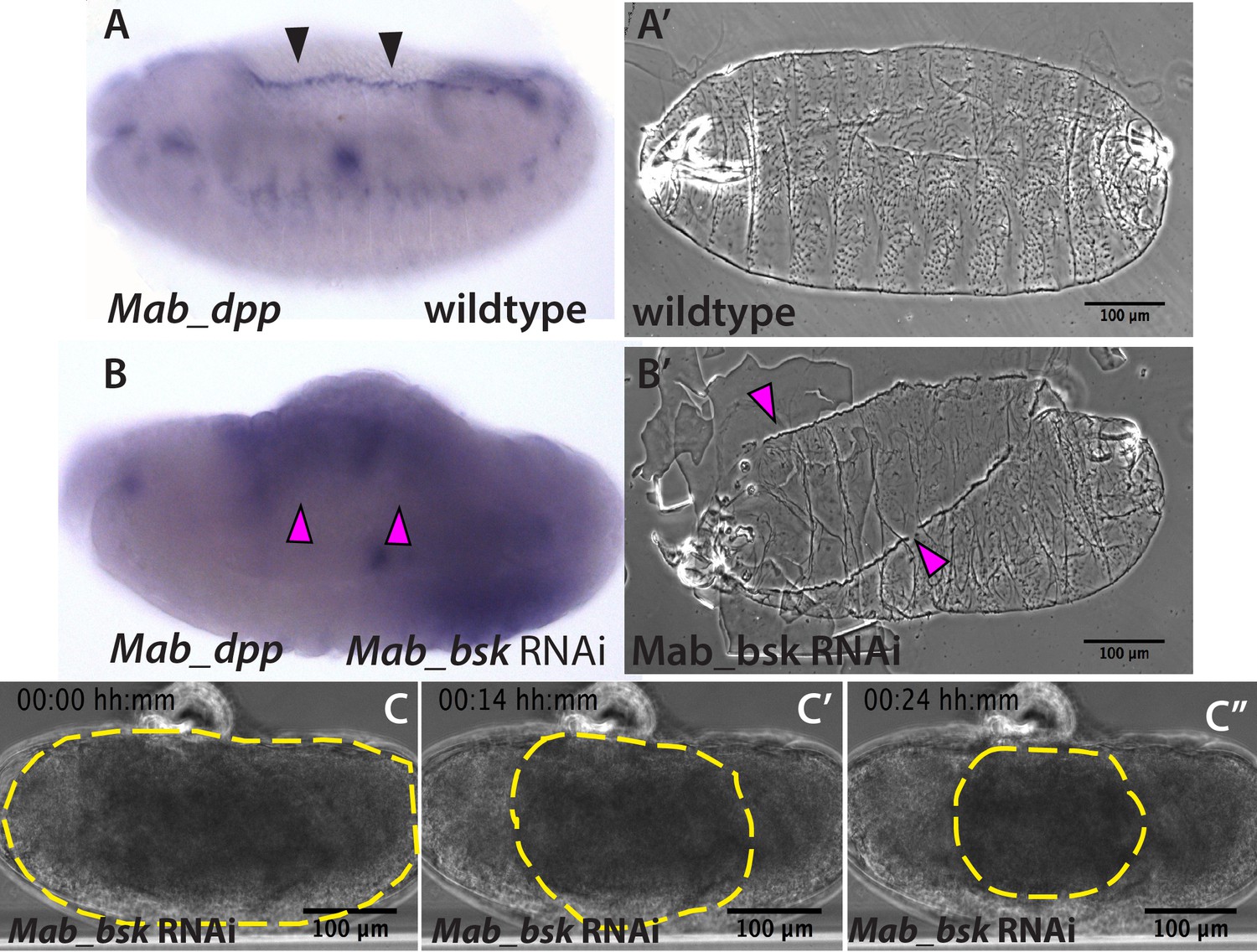
JNK (bsk) is required for dpp expression and the completion of dorsal closure without affecting serosa rupture or retraction in Megaselia abdita.
(A) Wild-type expression of Mab_dpp (purple stain, black arrowheads) along the dorsal leading edge of the epidermis in a M. abdita embryo (lateral view) during dorsal closure. (A’) Cuticle preparation of a wild-type late-stage pre-hatching M. abdita embryo (dorsal view). (B) RNAi knock-down of Mab_bsk abolishes Mab_dpp expression at the leading edge of the epidermis (magenta arrowheads) in a M. abdita embryo (lateral view). (B’) Cuticle preparation of a Mab_bsk RNAi late-stage pre-hatching embryo, showing a dorsal-open phenotype (magenta arrowheads). (C) Images from a bright-field time-lapse sequence of serosa retraction in a M. abdita embryo treated with Mab_bsk RNAi (from Video 3). Yellow dashed lines show the perimeter of the serosa covering the embryo during retraction. In all embryos and cuticles anterior is to the left and dorsal to the top.
Figure 2—figure supplement 1
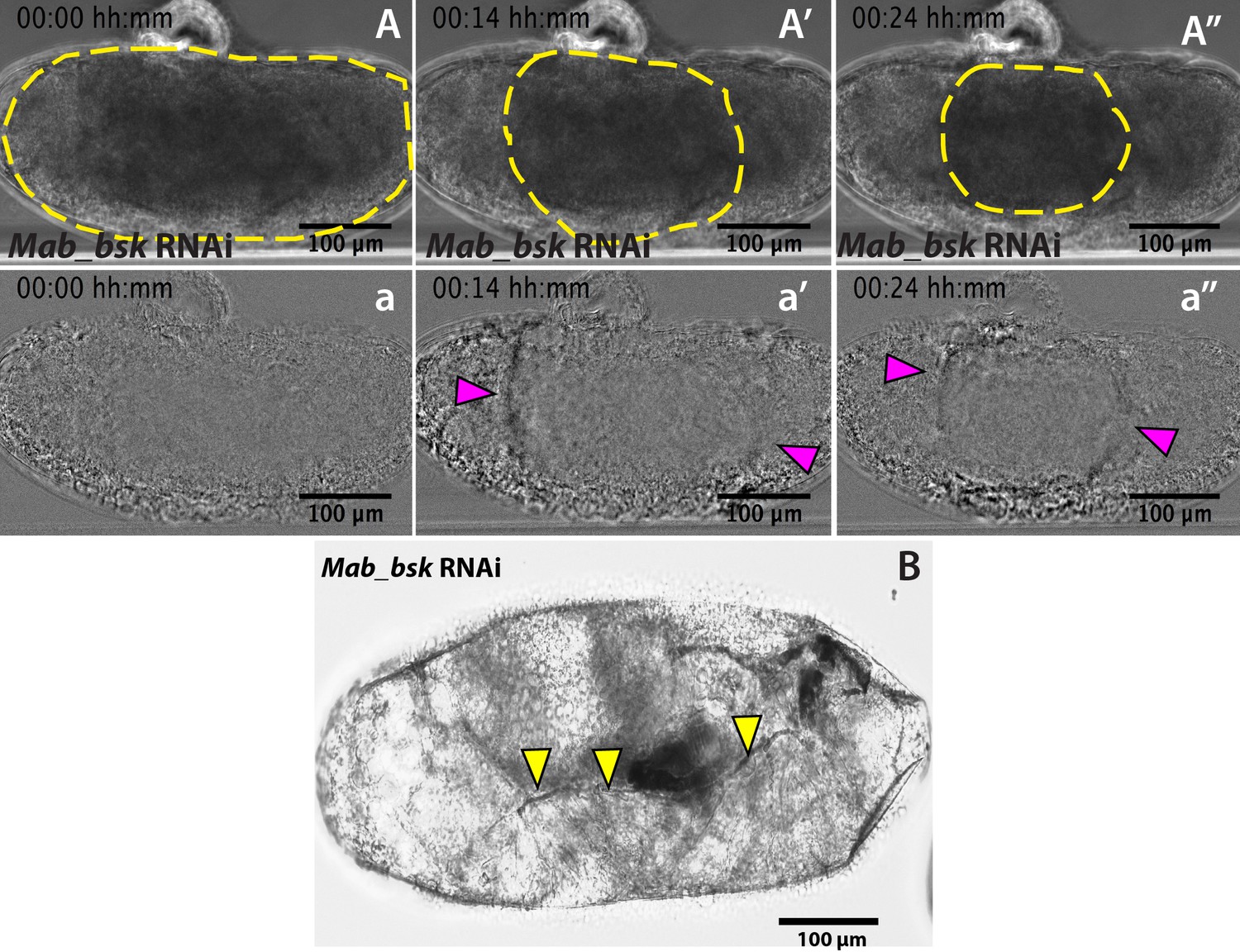
Mab_bsk RNAi results in a lack of dorsal closure progression after serosa rupture.
(A–A’’) Detection of the serosa edge in the bright-field time-lapse sequence of Mab_bsk knock-down embryo from Figure 2C was obtained by image subtraction (a–a’’) to render the retracting serosa visible (magenta arrowheads). (B) Mab_bsk knock-down results in a sclerotized cuticle at the leading edge of the epidermis (yellow arrowheads) in late-stage pre-hatching embryos. This phenotype is due to the lack of dorsal closure progression. Note this embryo is the same as the embryo in the time-lapse sequence in Figure 2C,C’ and C’’ and Video 3, 24 hr after dsRNA injection.
Figure 3 with 2 supplements
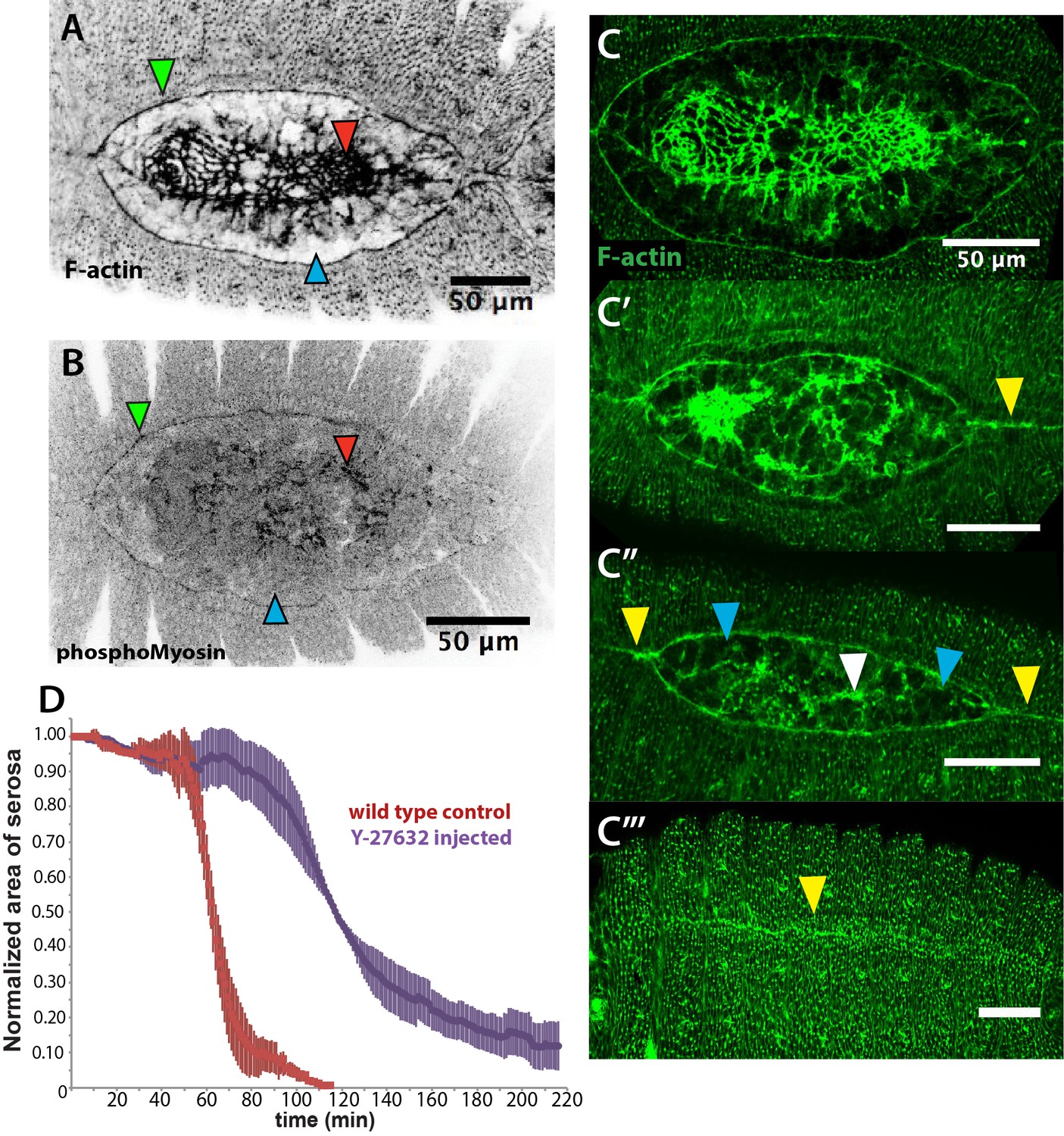
An actomyosin-enriched epidermal cable is necessary for the completion of dorsal closure upon contraction, retraction, and internalization of the serosa in Megaselia abdita.
(A) F-actin enrichment (as observed by inverted intensity of phalloidin staining) and (B) PhosphoMyosin enrichment (inverted intensity of immunostaining) in the epidermal leading edge (green arrowhead) and internalizing serosa (red arrowhead), but not in the amnion (blue arrowhead) of a M. abdita embryo during dorsal closure. (C–C’’’) Time series of different phalloidin-stained M. abdita embryos that show merging of both amniotic and epidermal flanks brought together at the dorsal midline upon serosa internalization. Amniotic merging (blue arrowheads) is visible by a transient accumulation of F-actin (white arrowhead in C’’). Epidermal merging is mediated by the actomyosin cable (yellow arrowheads). (D) Average area of the serosa during retraction in wild-type control (red, n = 15 embryos), and contractility-impaired embryos injected with Y-27632 (purple, n = 15 embryos). The latter show delayed and incomplete serosal retraction and ingression. Vertical bars represent standard deviations (SD). Embryos from A to D are in dorsal view where anterior is to the left. Embryo in D’ is an optical transverse view where dorsal is to the top.
Figure 3—figure supplement 1
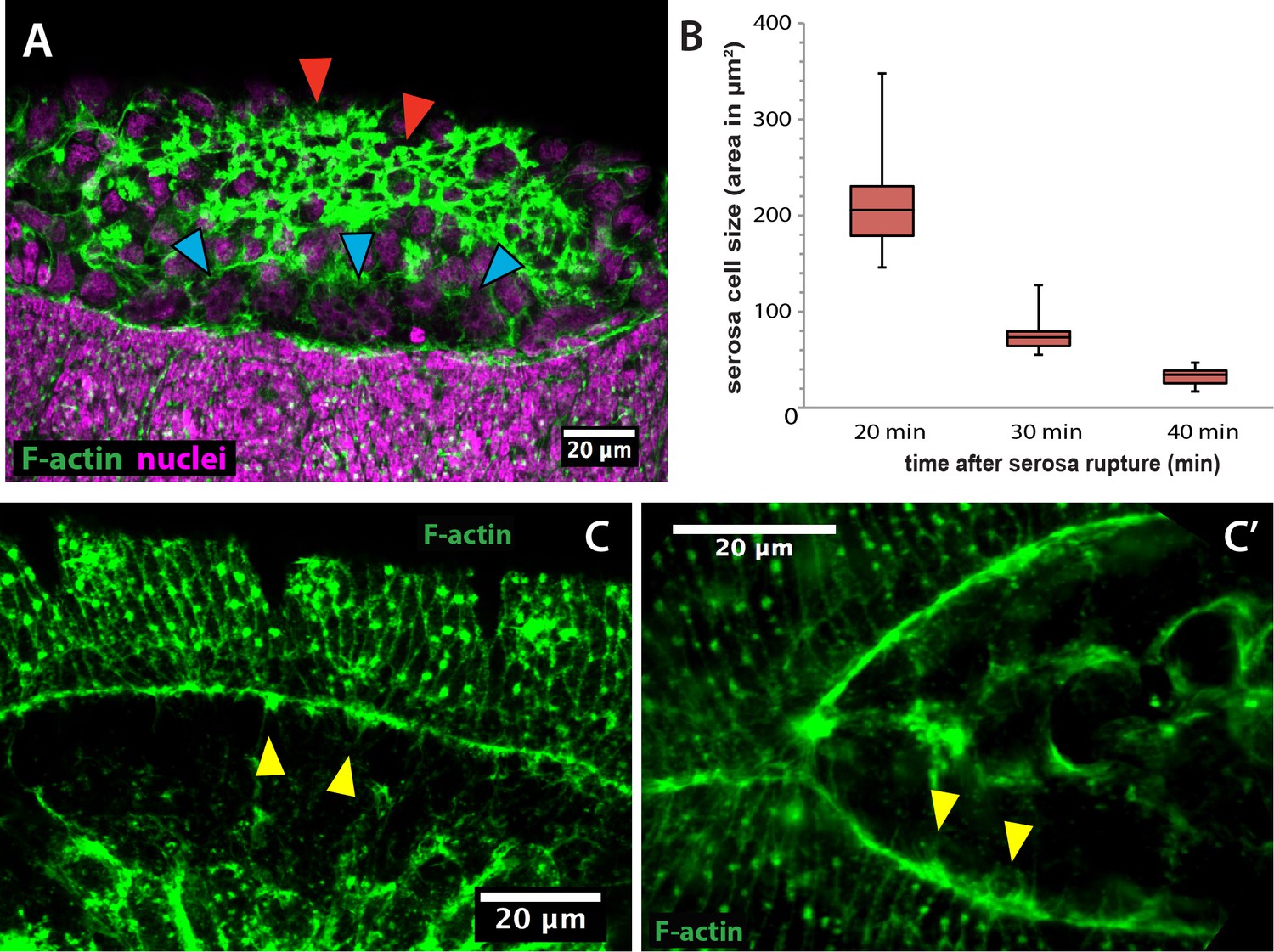
Actin accumulates apically in contracting serosa cells and filopodial protrusions are present in the epidermal actin cable of Megaselia abdita.
(A) Lateral view of a M. abdita embryo showing F-actin accumulation at the epidermal leading edge and the internalizing serosa (red arrowheads), but not in the extraembryonic amnion (blue arrowheads). Phalloidin staining in green and DAPI counterstain in magenta. Anterior is to the left and dorsal to the top. (B) Serosa cell surface area reduces over time during contraction from an average of 212 ± 52 µm2 (SD; n = 20 cells) to 76 ± 18 µm2 (SD; n = 20 cells) to 33 ± µm2 (SD; n = 20 cells) at 20, 30 and 40 min after serosa rupture, respectively. (C) and (C’) Filopodia-like structures (yellow arrowheads) extend from the epidermal cable towards the dorsal midline as evidenced in dorsal view of a M. abdita embryo. Phalloidin staining in green. Anterior is to the left.
Figure 3—figure supplement 2
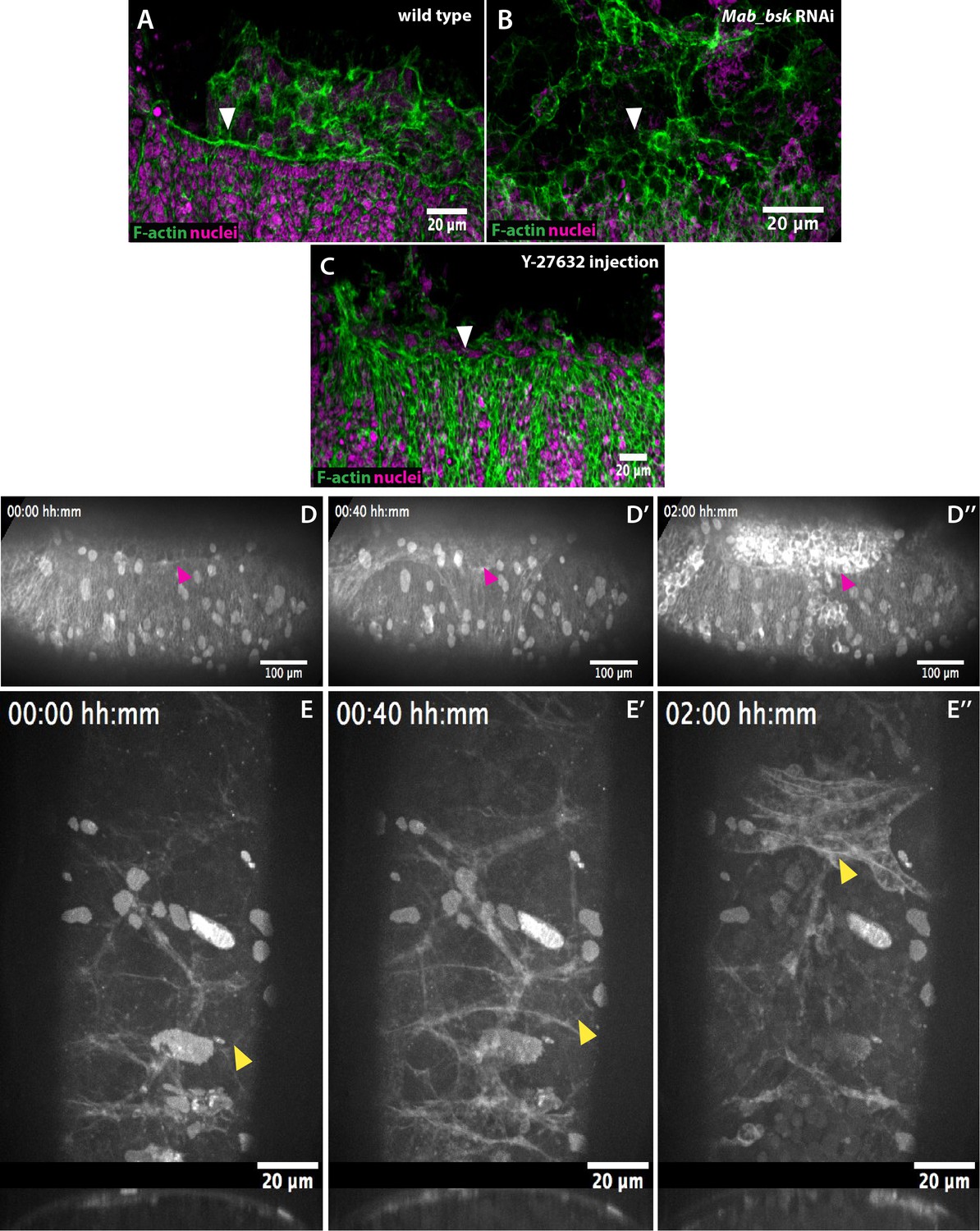
The epidermal actomyosin cable is a contractile structure controlled by JNK (Mab_bsk) expression during dorsal closure.
(A) An actomyosin cable (white arrowhead) at the epidermal leading edge of a wild-type M. abdita embryo. (B) Mab_bsk RNAi results in the absence of an actomyosin cable (white arrowhead) in M. abdita embryos during dorsal closure. (C) Downregulation of the actomyosin cable (white arrowhead) after injection of the Rho kinase (ROCK) inhibitor Y-27632. From A to C, embryos are labeled with phalloidin staining in green and DAPI nuclear counterstain in magenta. Anterior is to the left and dorsal to the top. (D–D’’) Kinetics of serosal retraction in embryo injected with the Rho-kinase inhibitor Y-27632. Drug treatment arrests the progression of the epidermal leading edge (magenta arrowheads) and abolishes the dorsal internalization of the serosa cells (E–E’’) Dorsal and transversal views of the retraction of the serosa in Y-27632 treated embryos. The inward bending and apicobasal cell elongation of the serosa are prevented and disruption of the tissue occurs without apparent contraction. Embryos in D and E are taken from time-lapse sequences of FM 4–64-labeled embryos from Videos 4 and 5, respectively. In D, anterior is to the left and dorsal to the top. In E, the top panel is a dorsal view with anterior to the top and the bottom panel is a transversal view with dorsal to the top.
Figure 4 with 5 supplements
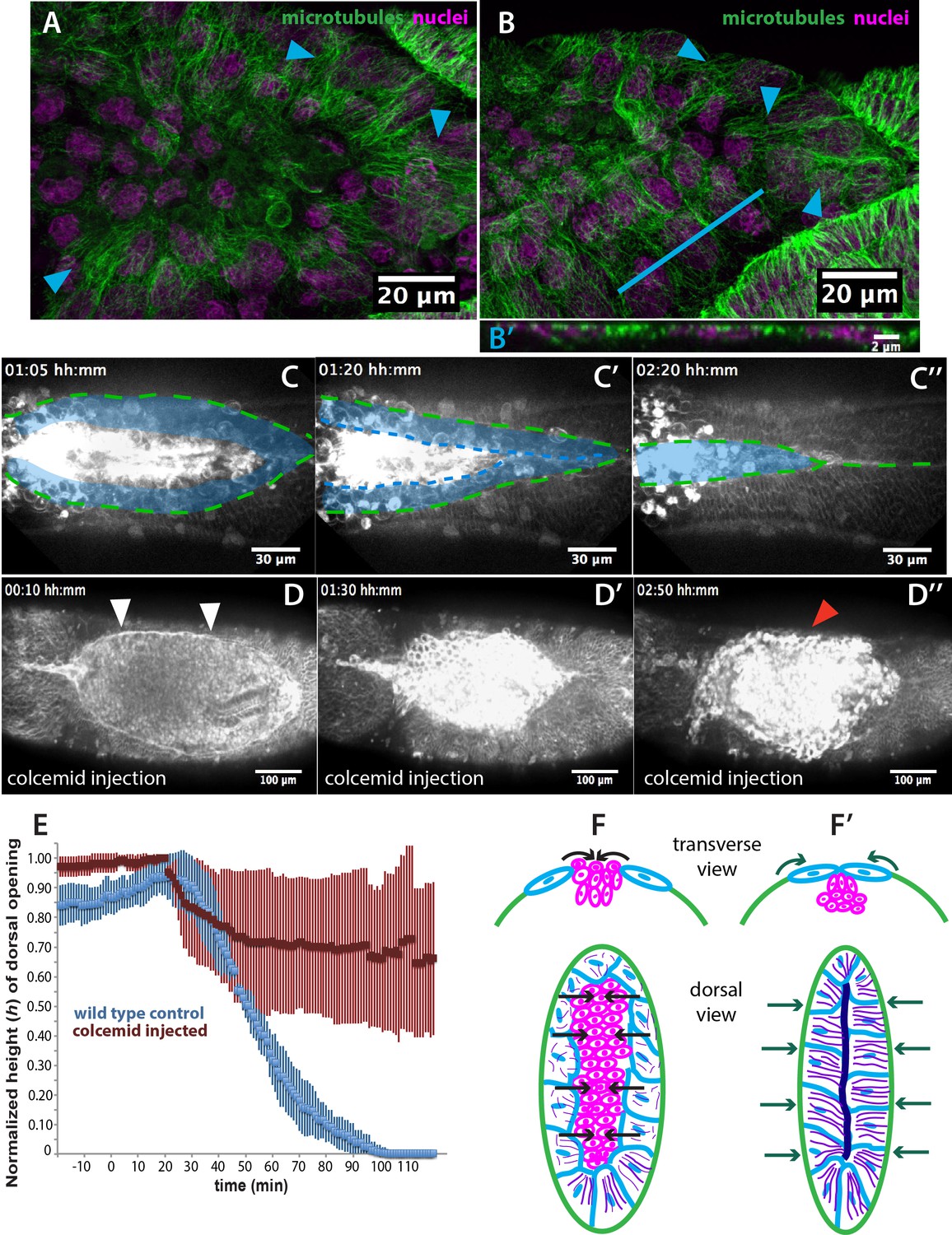
Microtubule-dependent seaming of the extraembryonic amnion is required for subsequent epidermal seaming during dorsal closure in Megaselia abdita.
(A) Confocal projections of microtubules in the extraembryonic amnion orienting toward the internalizing serosa during early amniotic seaming. (B) As amniotic seaming progresses, microtubules maintain alignment, and become localized apically, as revealed by a transverse view (B’), obtained by orthogonal stack re-slicing along the blue line in B. Staining against β-tubulin is shown in green, DAPI nuclear counterstain in magenta. (C) Time-lapse sequence of amniotic seaming (blue dashed line and blue shaded area) followed by epidermal seaming (green dashed line) along the dorsal midline in a M. abdita embryo injected with the fluorescent lipophilic dye FM 4–64 (from Video 6). (D) Time-lapse sequence of dorsal closure in a colcemid-treated embryo to induce microtubule depolymerization. After serosa retraction the process is arrested and closure fails. The embryo is labeled by FM 4–64 (from Video 8). White arrowheads in D show the proper straightening of the epidermal leading edge in colcemid-injected embryos. Red arrowhead in D’’ indicates impaired serosa internalization. (E) Relative changes in height (h) of dorsal opening during in wild-type control (blue, n = 15 embryos) and embryos injected with colcemid (red, n = 15 embryos) to depolymerize microtubules. Vertical bars show standard deviations (SD). These measurements reveal failed epidermal leading edge progression in colcemid-treated M. abdita embryos. (F) Schematics depicting the transverse view (top) and dorsal view (bottom) of embryos during serosa cell internalization, initiation of amnion cell elongation, and microtubule alignment (purple) toward the dorsal midline. Black arrows indicate the direction of amnion progression. (F’) Amnion cells show alignment of apical microtubule bundles toward the dorsal midline, where the two amniotic flanks meet, and amniotic seaming occurs (dark blue line). This is followed by the progression of the epidermal leading edge (dark green arrows), which results in epidermal seaming and completion of dorsal closure (not shown). Serosa in magenta, amnion in blue and embryonic epidermis in green. All embryo images show dorsal views where anterior is to the left.
Figure 4—figure supplement 1
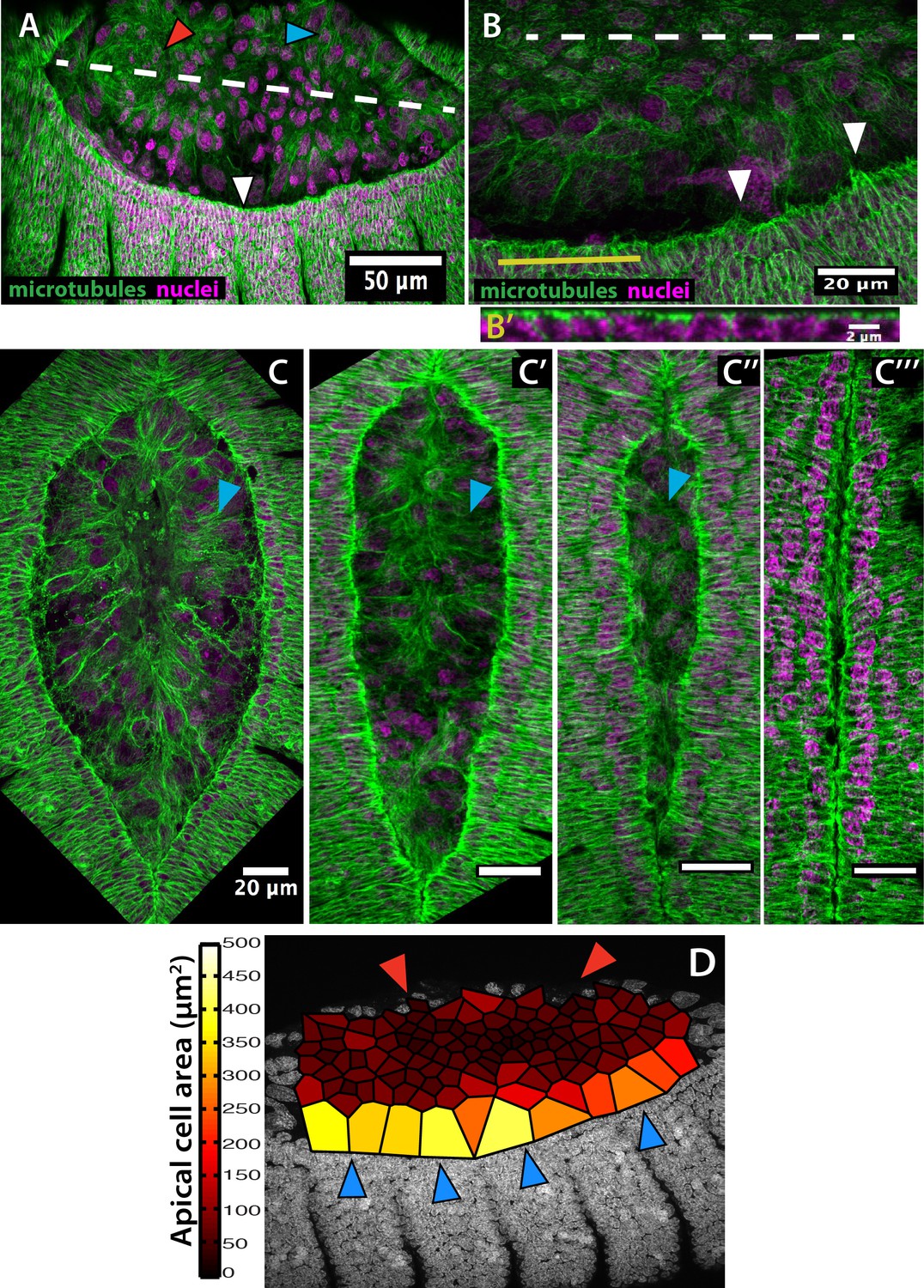
Microtubule alignment is present in both the epidermis and the amnion and correlates with enlarged amniotic cells in Megaselia abdita.
(A) Microtubule bundles orient toward the dorsal midline (white dashed line) in both the epidermis (white arrowhead) and the amnion (blue arrowhead) of M. abdita embryos during dorsal closure. Note that microtubules in the internalizing serosa lack a clear orientation (red arrowhead). (B) At the epidermal leading edge of M. abdita embryos, microtubule bundles orient dorsoventrally (white arrowheads) and show apical localization as revealed by a transverse view (B’) obtained by orthogonal stack re-slicing along the yellow line in B. (C–C’’’) A timed series of fixed embryos shows that microtubule alignment (blue arrowheads) and amnion cell elongation are maintained during serosa internalization (C) and the merging of amnion cells from opposite flanks (C’). Amnion microtubule alignment is lost after the opposite amnion flanks meet at the dorsal midline (C’’). In A-C, β-tubulin staining in green and DAPI nuclear counterstain in magenta. (D) Estimated apical cell surface area by Voronoi tessellation around the nuclei of extraembryonic cells stained with DAPI. Amnion cells (blue arrowheads) present an elongated apical cell surface area compared to serosa cells (red arrowheads). In all embryos, anterior is to the left.
Figure 4—figure supplement 2
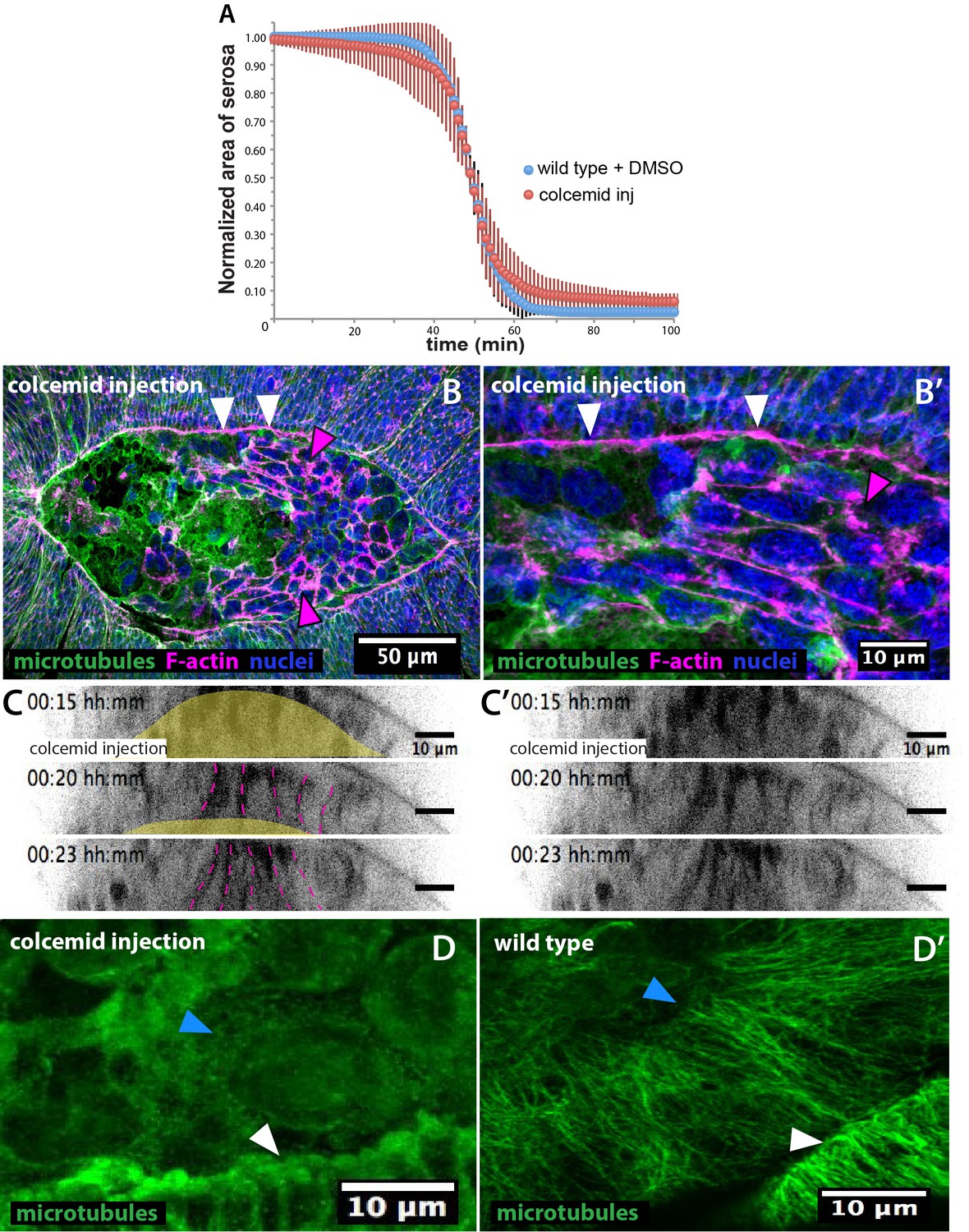
Colcemid treatment prevents microtubule polymerization without affecting serosa retraction or epidermal actomyosin cable in Megaselia abdita.
(A) Average area of serosa during retraction in wild-type control embryos (blue, n = 10 embryos) and colcemid-injected (red, n = 10 embryos). Kinetics of serosa retraction did not show an apparent difference in treated embryos compared to wild-type control. Vertical bars represent standard deviations (SD). (B) Epidermal actomyosin cable (white arrowheads) and contracting serosa (magenta arrowheads) in colcemid-injected embryos do show similar F-actin enrichment than wild type during the initial stages of internalization. The embryo is in dorsal view. Anterior is to the left. (B’) Higher magnification of B showing F-actin accumulation in the epidermal cable (white arrowheads) and the contracting serosa (magenta arrowhead). β-tubulin staining in green, phalloidin counterstain in magenta and DAPI counterstain in blue. (C–C’) Orthogonal views from a time-laspe sequence of colcemid-injected M. abdita embryos labeled with FM 4–64 (from bottom panel of Video 7). Colcemid-treated embryos undergo serosa internalization and apicobasal cell elongation toward the yolk similarly than wild type. The yellow area in C highlights the yolk region and the dashed magenta lines highlight serosa cell apicobasal junctions. C’ shows the sequence in C without annotations. Dorsal is to the top. (D) Colcemid injection during dorsal closure induces microtubules depolymerization in M. abdita embryos as observed in both the leading edge of the epidermis (white arrowhead) and the extraembryonic amnion (blue arrowhead) compared to a wild-type non injected embryo (D’). β-tubulin staining in green. Anterior is to the left.
Figure 4—figure supplement 3
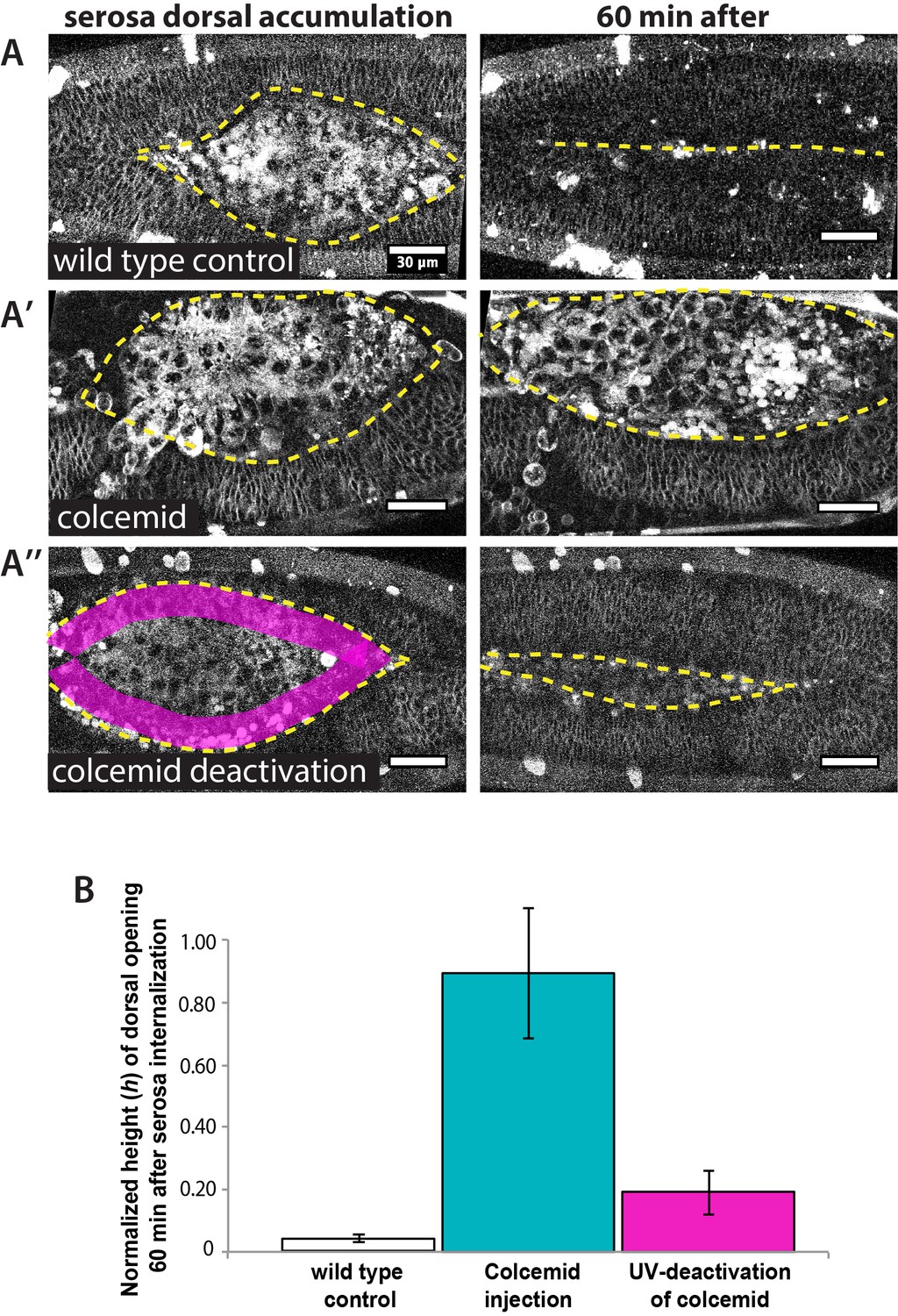
UV-deactivation of colcemid in the amnion region allows dorsal closure progression in treated Megaselia abdita embryos.
(A) (Left panel) Dorsal view of a M. abdita wild-type control embryo injected with FM 4–64 during serosa accumulation at the dorsal opening. Yellow dashed line represents the edge of the epidermal flanks. (Right panel) Dorsal closure is completed 60 min after the initiation of serosa internalization. (A’) (Left panel) Dorsal view of a colcemid injected embryos labeled with FM 4–64 during serosa accumulation at the dorsal opening. (Right panel) 60 min after, dorsal closure does not progress and the epidermal flanks remain open. (A’’) (Left panel) Dorsal view of a colcemid injected embryos labeled with FM 4–64 in which UV light is irradiated in a region between the epidermal flanks and the internalizing serosa (magenta shaded area, corresponding to the amnion) of colcemid-injected embryos. (Right panel) Dorsal closure progressed further in UV-irradiated embryos, although the epidermis did not close completely after 60 min of serosa internalization. Yellow dashed lines indicate the epidermal leading edge (B) Relative changes in height (h) of dorsal opening in wild-type control embryos (white bar; 4 ± 1%, n = 10 embryos), colcemid-injected embryos (blue bar; 90 ± 21%, n = 10 embryos) and colcemid-treated/UV-irradiated embryos (magenta bar; 19 ± 7%, n = 10 embryos). Error bars are standard deviations (SD). These measurements indicate a rescue of dorsal closure progression after UV-deactivation of colcemid in M. abdita injected embryos.
Figure 4—figure supplement 4
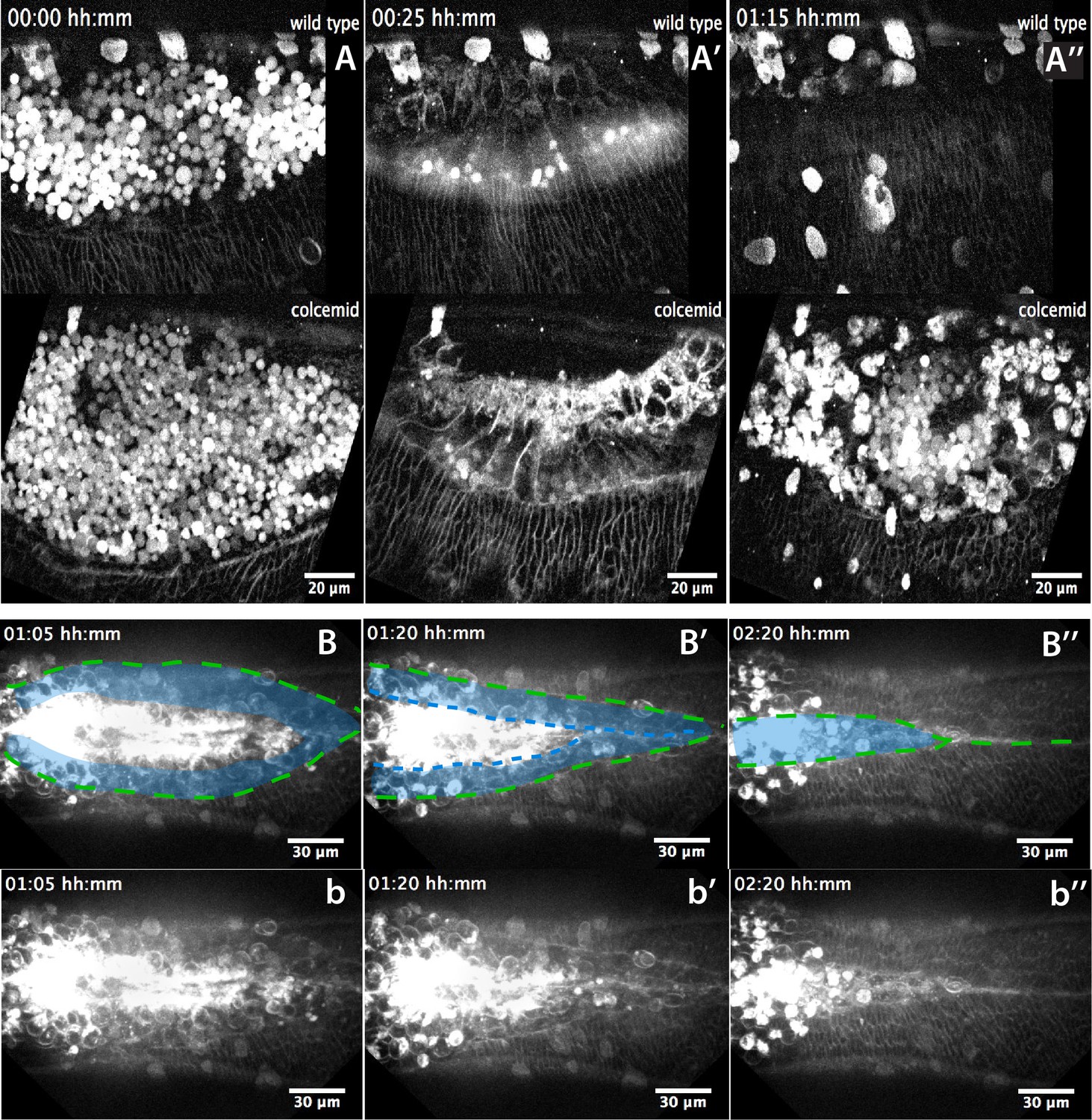
Landmarks of amniotic seaming during dorsal closure progression in Megaselia abdita.
(A–A’’) Lateral view from two time-lapse sequences of dorsal closure progression in M. abdita embryos labeled with FM 4–64 (from Video 9). Top panels show a wild-type embryo undergoing serosa accumulation (A), amnion cell elongation (A’) and epidermal edge progression towards the dorsal midline (A’’). Bottom panels show a M. abdita embryo treated with colcemid undergoing serosa accumulation, amnion cell elongation and further retraction of the epidermal leading edge from the dorsal midline. In all embryos, dorsal is at the top. (B–B’’) Dorsal view from a time-lapse sequence of amniotic seaming (blue dashed line and blue shaded area) followed by epidermal seaming (green dashed line) along the dorsal midline in a M. abdita embryo injected with FM 4–64 (from Video 6). (b–b’’) Raw images from B-B’’ without superimposed labels. In all embryos, anterior is to the left.
Figure 4—figure supplement 5
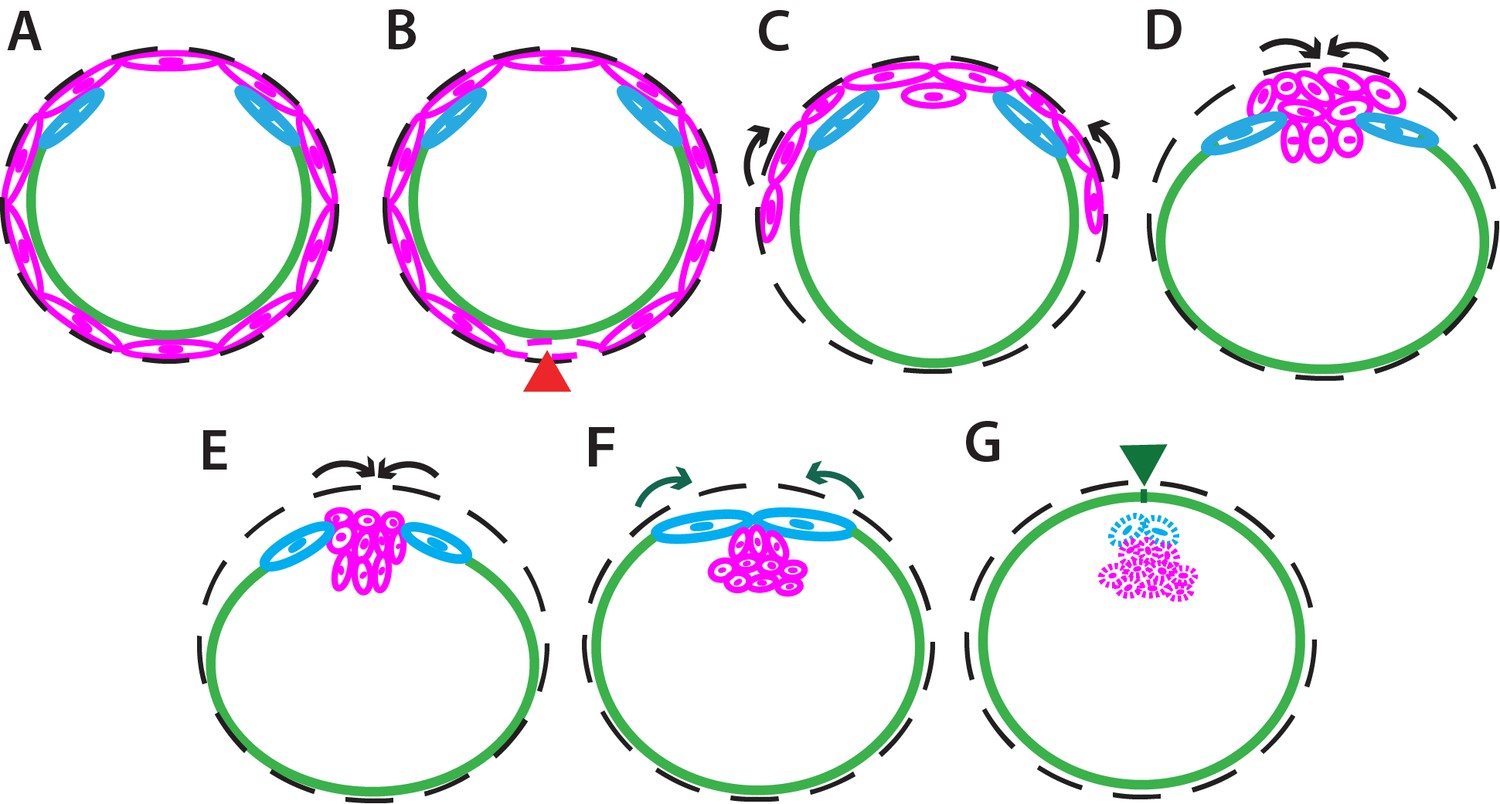
Schematics of dorsal closure in Megaselia abdita.
(A) Schematics depicting in transverse view the organization of the serosa cells (magenta), amnion cells (blue), and embryonic epidermis (green) prior to dorsal closure. The black dashed line represents the vitelline envelope. (B) Serosa rupture propagates along the ventral midline of the embryo (red arrowhead) and the remaining lateral serosa cells retract toward the dorsal opening (C). Serosa cells accumulate on the dorsal side of the embryo (D) and internalize through an actomyosin-dependent contraction (E). Black arrows depict the direction of serosa retraction and internalization. (F) Upon serosa internalization, amnion cells elongate toward the dorsal midline where the two amniotic flanks meet and fuse through a microtubule-dependent mechanism. Amniotic seaming is followed by the progression of the epidermal leading edge (green arrows). (G) The fused amnion also internalizes following the serosa, while the epidermal flanks meet at the dorsal midline and fuse through a second microtubule-dependent seaming event (green arrowhead). Extraembryonic tissue internalization (serosa and amnion) and epidermal seaming lead to the completion of dorsal closure, resulting in a continuous epidermis surrounding the embryo. In all schematics dorsal is to the top and ventral to the bottom.
Author response image 1
Author response image 2
Author response image 3

Author response image 4
Author response image 5
Videos
Video 1
Dorsal closure in Megaselia abdita involves the rupture and retraction of the serosa and advancement of the epidermal flanks to the dorsal midline.
Time-lapse sequence of dorsal closure in two M. abdita embryos injected with the fluorescent label FM 4–64. Top: serosa rupture and retraction in lateral view. Rupture initiates at a ventral-posterior location and spreads anteriorly along the ventral side of the embryo. The ruptured serosa accumulates on the dorsal side, where epidermal seaming occurs at the end of dorsal closure. Bottom: dorsal view of the initiation of dorsal closure, marked by the fusion of the dorsal ridge and straightening of the epidermal leading edge (see Figure 1—figure supplement 3A). Epidermal flanks are brought together to the dorsal midline where epidermal seaming occurs. In both embryos, anterior is to the left.
Video 2
The extraembryonic serosa of Megaselia abdita embryos internalizes into the yolk prior to epidermal seaming.
Time-lapse sequence of a dorsal (top) and orthogonal view (bottom) of an FM 4–64-labeled embryo. After serosa rupture and accumulation at the dorsal opening, the extraembryonic tissue internalizes into the yolk, as observed by an inward bending of the tissue and apicobasal cell elongation. Upon internalization of the serosa, the epidermal flanks advance and fuse at the dorsal midline. In the dorsal view, anterior is to the top. In the orthogonal view, dorsal is to the top.
Video 3
Knock-down of Mab_bsk by RNAi does not prevent serosa rupture and retraction despite preventing epidermal flanks from closing in Megaselia abdita embryos.
Bright-field time-lapse sequence of serosa retraction in a M. abdita embryo injected with Mab_bsk dsRNA at early stages of development. Embryo in lateral view. Anterior is to the left, dorsal to the top.
Video 4
Injection of the Rho kinase inhibitor Y-27632 prevents dorsal closure and slows down serosa retraction in Megaselia abdita.
Time-lapse sequence of dorsal closure in a FM 4–64-labeled M. abdita embryo injected with the Rho kinase (ROCK) inhibitor Y-27632, which downregulates actomyosin-based contractility. Note the slow rupture and retraction of the serosa, and the failure of the epidermal flanks to advance and close. Embryo in lateral view where anterior is to the left, dorsal to the top. Note that the static stains observed are stains on the vitelline envelope arising from embryo treatment prior to imaging.
Video 5
Injection of the Rho kinase inhibitor Y-27632 prevents internalization of the serosa in Megaselia abdita embryos.
Time-lapse sequence of a dorsal (top) and orthogonal view (bottom) of serosa rupture in an FM 4–64-labeled M. abdita embryo. Injection of the Rho kinase (ROCK) inhibitor Y-27632 (to downregulate actomyosin-based contractility) prevents internalization of the serosa. The inward bending and apicobasal cell elongation of the extraembryonic tissue into the yolk is not observed in orthogonal view. In the dorsal view, anterior is to the top. In the orthogonal view, dorsal is to the top. Note that the static stains observed are stains on the vitelline envelope arising from embryo treatment prior to imaging.
Video 6
Amniotic seaming followed by epidermal seaming during the late stage of dorsal closure in Megaselia abdita.
Time-lapse sequence of the final stage of dorsal closure in a M. abdita embryo fluorescently labeled with FM 4–64. The amnion flanks are brought together and seamed at the dorsal midline upon serosa ingression. This process is followed by seaming of the epidermal flanks at the dorsal midline. Both seaming processes initiate at the posterior end of the embryo. Embryo in dorsal view. Anterior is to the left. Note that the static stains observed are stains on the vitelline envelope arising from embryo treatment prior to imaging.
Video 7
The initial stages of serosa internalization in colcemid-injected embryos of Megaselia abdita occur as in wild-type embryos.
Time-lapse sequence in orthogonal view of dorsal closure during serosa internalization in FM 4–64-labeled M. abdita embryos. The inward bending of the extraembryonic tissue and apicobasal cell elongation into the yolk is observed in wild-type control (top) and colcemid-injected embryos (bottom). Injection of colcemid induces microtubule depolymerization and impairs the late stages of dorsal closure (see Video 8). Dorsal is to the top.
Video 8
Colcemid injection prevents dorsal closure in Megaselia abdita.
Time-lapse sequence of impaired dorsal closure in an FM 4–64-labeled M. abdita embryo after injection of colcemid to induce microtubule depolymerization. The initial stages of dorsal closure (straightening of the epidermal leading edge and serosa rupture and retraction) are not affected. The process of dorsal closure is aborted during the late stages (amniotic seaming and epidermal seaming). Embryo in dorsal view. Anterior is to the left.
Video 9
Colcemid injection impairs amniotic flank seaming during dosal closure in Megaselia abdita.
Time-lapse sequence in lateral view of the latest stages of dorsal closure in FM 4–64-labeled M. abdita embryos. Amnion cell elongation can be observed in a wild-type embryo (top) resulting in the seaming of the amniotic flanks followed by seaming of the epidermal flanks. In colcemid-injected embryos (bottom), amnion cells initially elongate but fail to maintain elongation and retract from the amniotic merging at the dorsal midline. Note that the static stains observed are stains on the vitelline envelope arising from embryo treatment prior to imaging.
Video 10
UV-deactivation of colcemid in the amnion region of treated Megaselia abdita embryos rescues amnion seaming.
Time-lapse sequence in dorsal view of dorsal closure in FM 4–64-labeled M. abdita embryos during serosa accumulation and internalization. Colcemid-injected embryos do not complete dorsal closure (top). UV-deactivation of colcemid in a region of interest (ROI) between the epidermal flanks and the internalizing serosa (bottom) rescues serosa internalization.
Additional files
-
Transparent reporting form
- https://doi.org/10.7554/eLife.33807.029
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Two consecutive microtubule-based epithelial seaming events mediate dorsal closure in the scuttle fly Megaselia abdita
eLife 7:e33807.
https://doi.org/10.7554/eLife.33807




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}