Estrogenic-dependent glutamatergic neurotransmission from kisspeptin neurons governs feeding circuits in females
- Oregon Health and Science University, United States
- Howard Hughes Medical Institute, University of Washington, United States
- Oregon National Primate Research Center, Oregon Health and Science University, United States
Figures
Figure 1
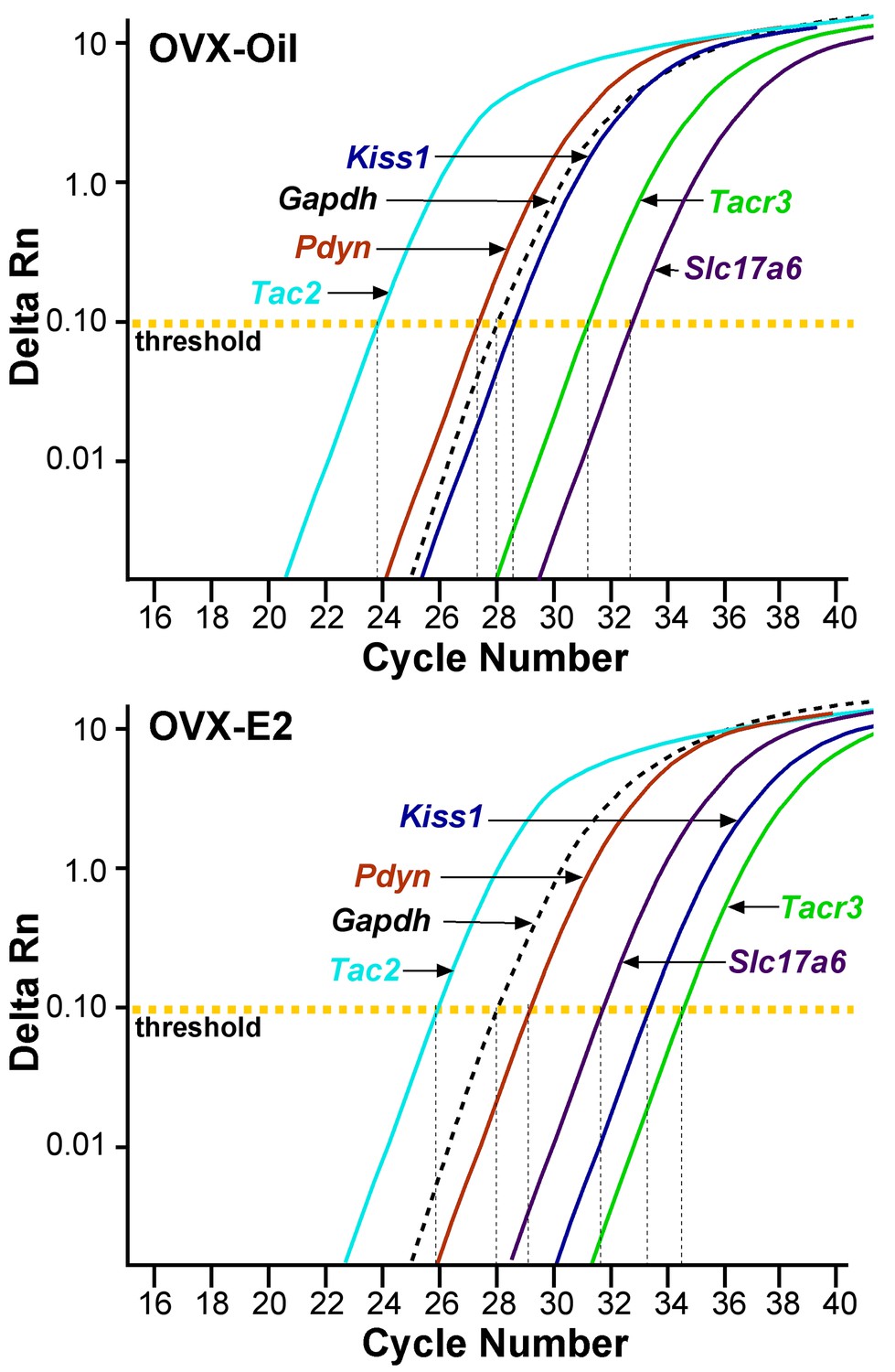
qPCR amplification assay illustrating the cycle threshold (CT) for the different neuropeptides and vGluT2 in Kiss1ARH neurons.
Cycle number is plotted against the normalized fluorescence intensity (∆RN) to visualize the PCR amplification of Tac2, Pdyn, Kiss1, Tacr3, Slc17a6 and the reference gene Gapdh in 5 cell Kiss1ARH pools obtained from oil- and E2-treated, OVX animals. The amplification efficiency for each primer pair is listed in Table 1. These efficiencies allowed us to use the comparative ∆∆CT methods for quantification. The cycle threshold (CT; horizontal dashed line) is the point in the amplification from which sample values were calculated using the 2-∆∆CT equation as described in the Methods.
-
Figure 1—source data 1
OVX + Oil Gene comparisons (fold change generated using OVX + Oil Tacr3 as calibrator) and OVX + E2 Gene comparisons.
- https://doi.org/10.7554/eLife.35656.003
Figure 2
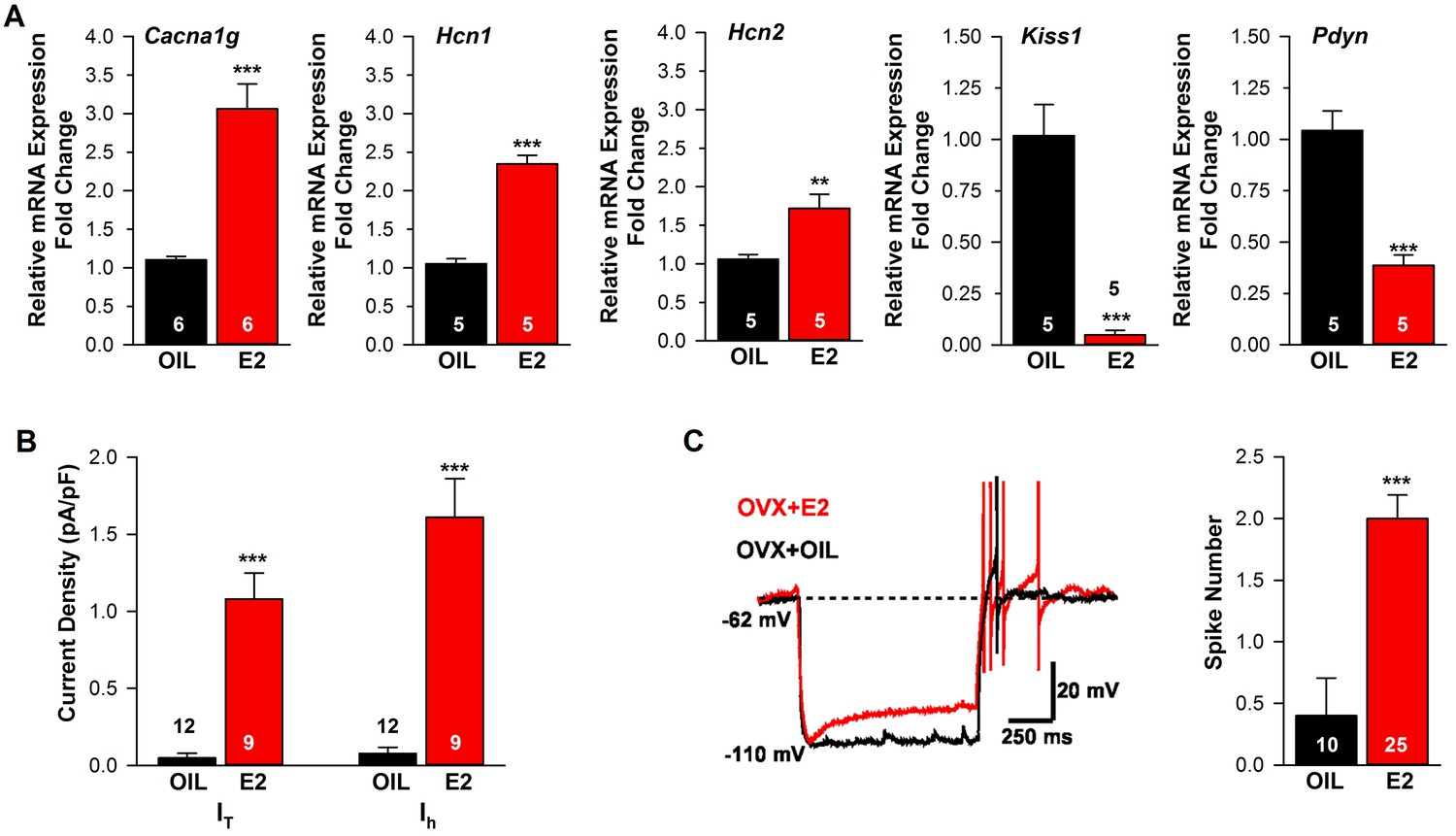
Estradiol regulation of ion channel mRNA expression and excitability of Kiss1ARH Neurons.
(A) Quantitative real-time PCR measurements of Cacna1g (Cav3.1), HCN1, HCN2, Kiss1 and Pdyn mRNAs in Kiss1ARH neuronal pools (3 pools of 5 cells each per animal) from OVX oil- and E2-treated mice (n = 5–6 animals per group). Note that E2 increased the mRNA expression of Cacna1g, Hcn1, Hcn2, but as expected decreased the mRNA expression of Kiss1 and Pdyn in the same Kiss1 neuronal pools (for, Cacna1g, Unpaired t-test t(10) = 6.037, p<0.001; Hcn1, Unpaired t-test, t(8) = 10.13, p<0.0001; Hcn2, Unpaired t-test, t(8) = 3.420, p<0.01; Kiss1, Unpaired t-test, t(8) = 6.348, p<0.001; Pdyn, Unpaired t-test, t(8) = 6.118, p<0.001). (B) T-type calcium current and h-current density (pA/pF) in Kiss1ARH neurons from OVX oil- and E2-treated mice (for T-current, t(19) = 6.956, p<0.0001; for h-current, t(19) = 6.964, p<0.0001; n = 9–12 neurons from 8 animals). Current densities were measured as previously described (Zhang et al., 2013). (C) Example of rebound burst firing in Kiss1ARH neurons (left), which increased fast Na+ spiking with E2, and summary data (right) from oil- versus E2-treated females (n = 10 and 25 neurons, respectively). Rebound firing was measured as previously described (Zhang et al., 2013). Bar graphs represent the mean ±SEM, (Unpaired t-test, t(33) = 4.455, p<0.0001). **p<0.01, ***p<0.001.
-
Figure 2—source data 1
Cacna1g, Hcn1, Hcn2, Kiss1 and Pdyn expression in Kiss1-ARH neurons (Figure 2A).
- https://doi.org/10.7554/eLife.35656.005
Figure 3

Estradiol regulation of Slc17a6 mRNA expression in Kiss1ARH neurons.
Quantitative real-time PCR measurements of Slc17a6, Tac2 and Tacr3 mRNAs in Kiss1ARH neuronal pools (3–6 pools of 5 cells each per animal) from OVX oil- and E2-treated mice (n = 4–7 animals per group). Note that E2 increased the mRNA expression of Slc17a6, but as expected decreased the mRNA expression of Tac2 and Tacr3 in the same Kiss1 neuronal pools. Bar graphs represent the mean ±SEM (for Slc17a6, Unpaired t-test, t(8) = 4.522, p<0.001; Tac2, Unpaired t-test, t(8) = 6.350, p<0.001; Tacr3, Unpaired t-test, t(6) = 7.161, p<0.001). ***p<0.001.
-
Figure 3—source data 1
Slc17a6, Tac2, Tacr3 expression in Kiss1-ARH neurons.
- https://doi.org/10.7554/eLife.35656.007
Figure 4
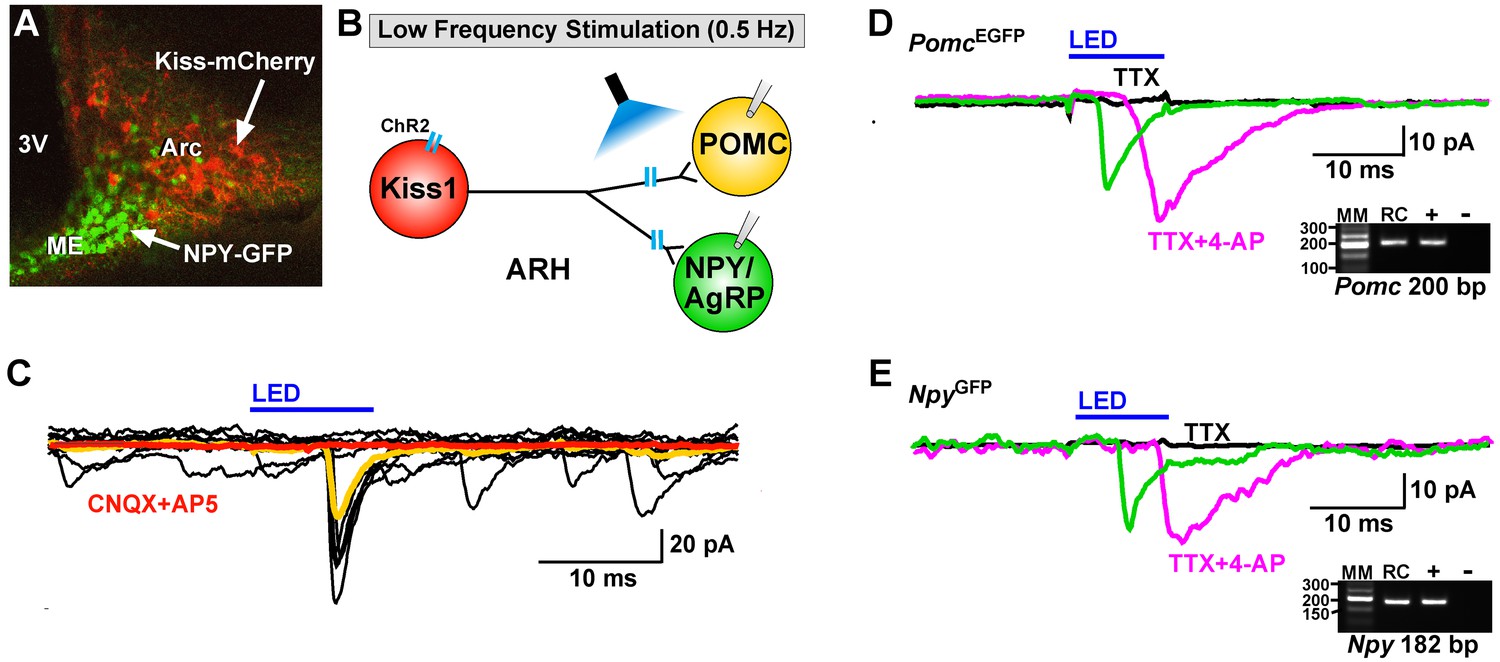
Optogenetic activation of Kiss1-ARH neurons directly excites POMC and NPY/AgRP neurons via glutamate release.
(A) AAV1-DIO-ChR2:mCherry was bilaterally injected into ARH of Kiss1Cre: : NpyGFP mice or Kiss1Cre: : PomcEGFP (not shown). (B), schematic of experimental design; whole-cell, voltage-clamp (Vhold = −60 mV) recordings were made in POMCEGFP or NPYGFP neurons and a single pulse (intensity, 660 μW; 10 ms duration) of blue light (470 nm) was delivered to the ARH. (C), a fast inward current was recorded in both POMC and NPY neurons (yellow trace is average) that was antagonized by CNQX (10 µM) and AP5 (50 µM) (red trace). (D,E), the optogenetic (glutamate) response (green trace) was abrogated in the presence of TTX (1 µM, black trace) but rescued with the addition of the K+ channel blocker 4-AP (100 µM, magenta trace) in both POMC (D) and NPY (E) neurons, n = 3 and 2, respectively. Insets show the scRT-PCR post hoc identification of representative recorded Pomc and Npy neurons. RC, recorded cell; +, positive control (with reverse transcriptase); -, negative control (without reverse transcriptase); MM, molecular marker.
Figure 5
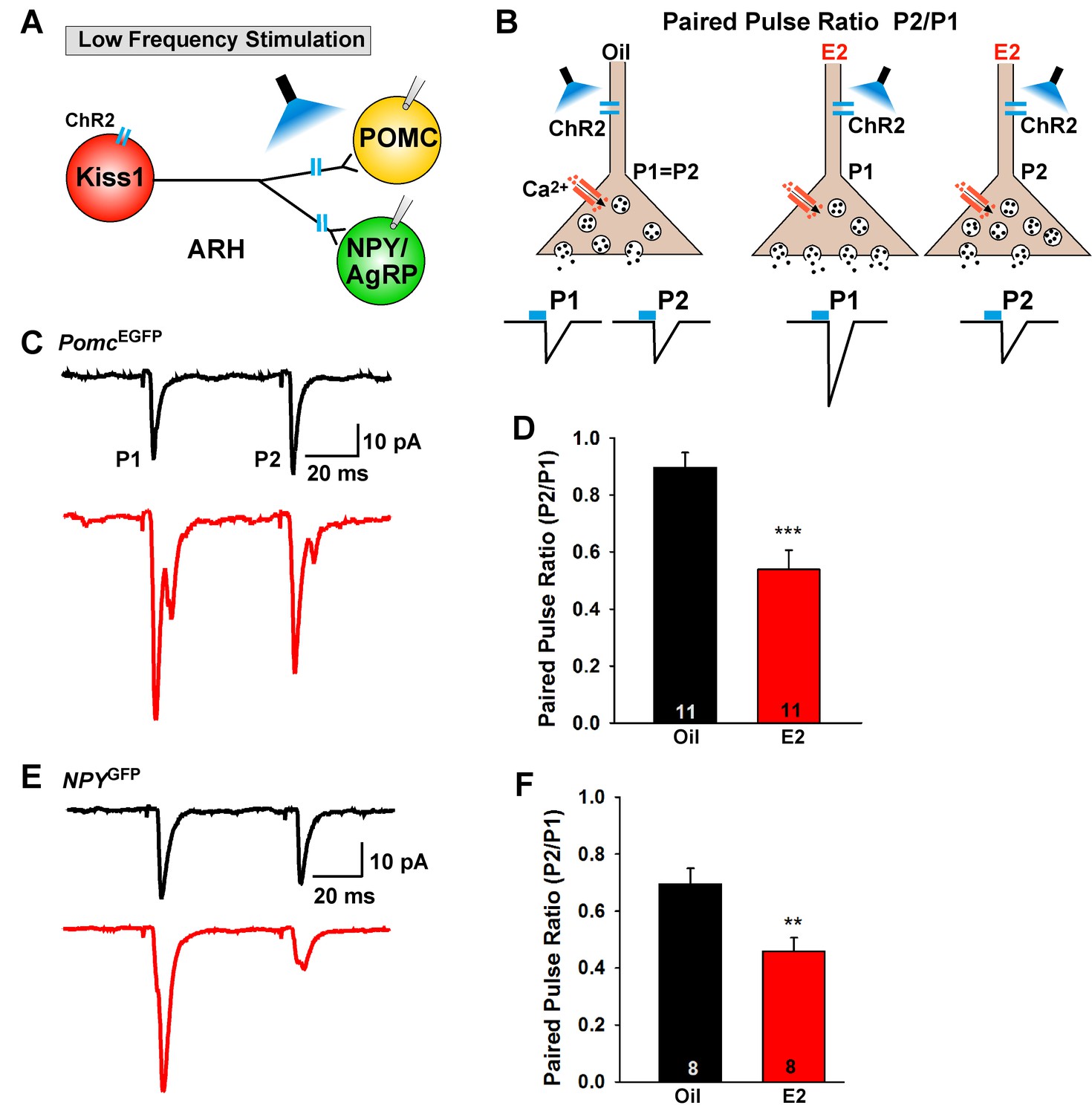
E2-treatment increases glutamate release from Kiss1ARH neurons onto POMC and NPY neurons.
(A) schematic of photostimulation of cells/terminals of Kiss1ARH neurons and recording from POMC or NPY/AgRP neurons. (B), illustration of a paired-pulse regime (two blue light pulses of 5 ms duration separated by 50 ms); fast glutamatergic inward currents (P1 and P2) were recorded to measure the probability of neurotransmitter release in postsynaptic neurons. (C), AAV1-DIO-ChR2:mCherry was bilaterally injected into ARH of Kiss1Cre::NpyGFP mice or Kiss1Cre::PomcEGFP mice. Using a paired-pulse regime, fast glutamatergic inward currents were recorded in POMCEGFP neurons (Vhold = −60 mV) from both oil-treated, OVX (upper trace, black) and E2-treated, OVX (lower trace, red) females. The averaged responses (50 sweeps) are shown. (D) E2-treatment significantly decreased the paired-pulse ratio (P2/P1; indicating that there was a higher probability of glutamate release from Kiss1Cre:GFP-ChR2 neurons (Unpaired t-test, t(20) = 4.184, p<0.001). (E) similarly using a paired-pulse regime, fast glutamatergic inward currents were recorded in NPYGFP neurons (Vhold = −60 mV) from both oil-treated, OVX (upper trace, black) and E2-treated, OVX (lower trace, red) females. The averaged responses (50 sweeps) are shown. (F) E2-treatment significantly decreased the paired-pulse ratio (P2/P1) indicating that there was a higher probability of glutamate release from Kiss1Cre:GFP-ChR2 neurons (Unpaired t-test, t(14) = 3.255, p<0.01). **p<0.01, ***p<0.001.
-
Figure 5—source data 1
The paired-pulse ratio (P2/P1) for Figure 5D and F.
- https://doi.org/10.7554/eLife.35656.010
Figure 6
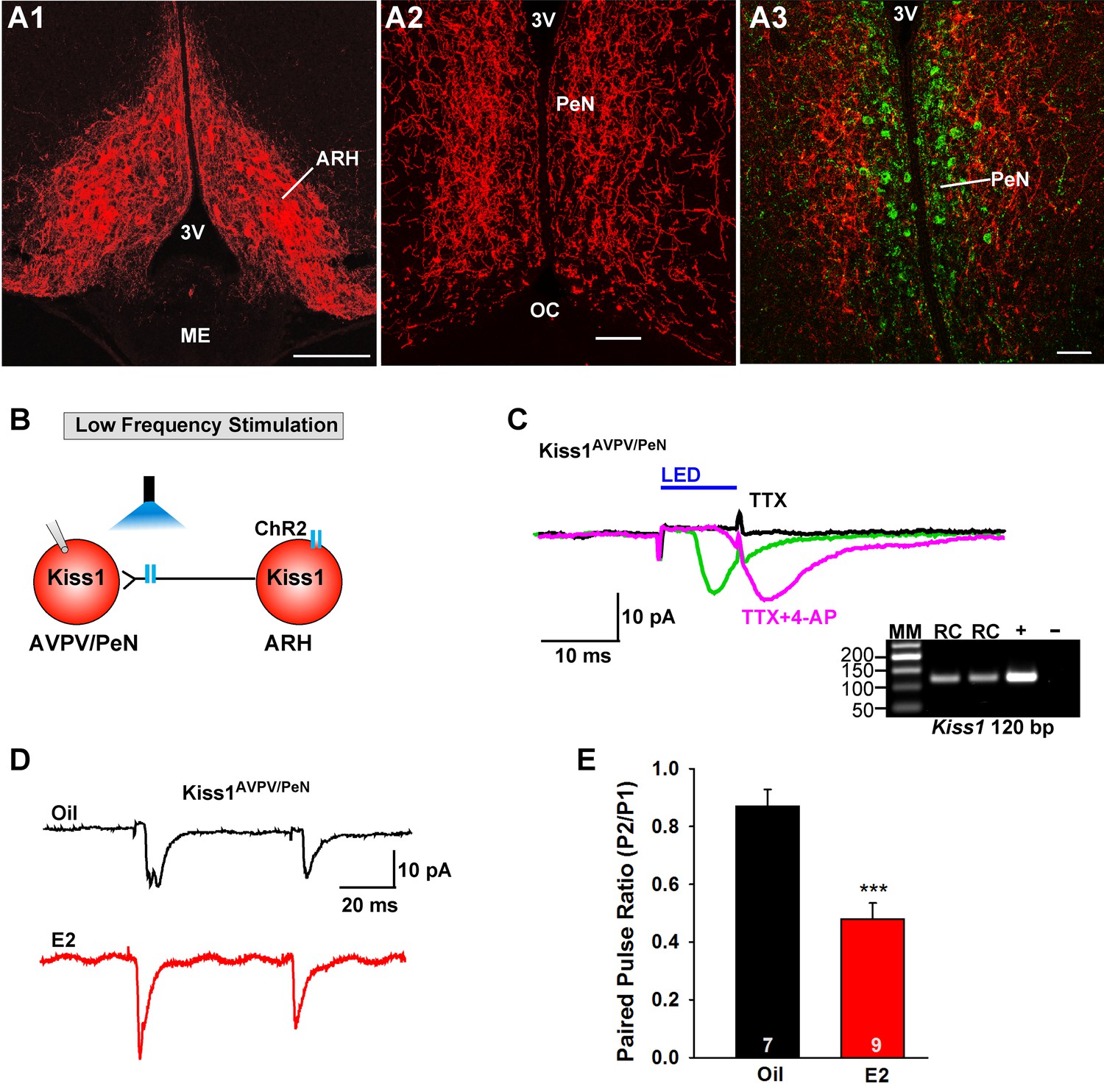
E2 treatment increases the probability of glutamate release from Kiss1ARH neurons onto Kiss1AVPV/PeN neurons.
(A1-A2) Photomicrographs showing the pronounced projections of ChR2:mCherry fibers to the preoptic area including the PeN following bilateral injections of AAV1-DIO-ChR2:mCherry in the ARH of Kiss1CreGFP V2 mice (note that the GFP was not visible in the V2 animals). Therefore, some POA sections from the same animals were stained for kisspeptin using the Caraty 564 antibody and revealed immunoreactive Kiss1 neurons in the PeN (green cells) (A3). Essentially none of the POA somas including the Kiss1 cells expressed ChR2-mCherry. Scale bars = 100 µM (A1,A2); 50 μm (A3). (B) schematic of photostimulation of the terminals of Kiss1ARH neurons and recording of Kiss1AVPV/PeN neurons. (C) following AAV1-DIO-ChR2:YFP (or mCherry) injection into the ARH, a fast inward current was recorded in Kiss1AVPV/PeN neurons following blue light stimulation (green trace). The response was antagonized by CNQX (10 µM) and AP5 (50 µM) (not shown) and was abrogated in the presence of TTX (1 µM, black trace) but rescued with the addition of the K+ channel blocker 4-AP (100 µM, magenta trace; n = 4 neurons). (D) using a paired-pulse regime (two blue light pulses of 5 ms duration separated by 50 ms), fast glutamatergic inward currents were recorded in Kiss1AVPV/PeN neurons (Vhold = −60 mV) from both oil-treated, OVX (upper trace, black) and E2-treated, OVX (lower trace, red) females. The averaged responses (50 sweeps) are shown. (E) E2-treatment significantly decreased the paired-pulse ratio (P2/P1) indicating that there was a higher probability of glutamate release from arcuate Kiss1Cre:ChR2 neurons (Unpaired t-test, t(14) = 4.748, p<0.001). ***p<0.001. Inset shows scRT-PCR post hoc identification of representative recorded Kiss1AVPV/PeN neurons. RC, recorded cell; +, positive control (with reverse transcriptase); -, negative control (without reverse transcriptase); MM, molecular marker.
-
Figure 6—source data 1
The paired-pulse ratio (P2/P1) for Figure 6E.
- https://doi.org/10.7554/eLife.35656.012
Figure 7
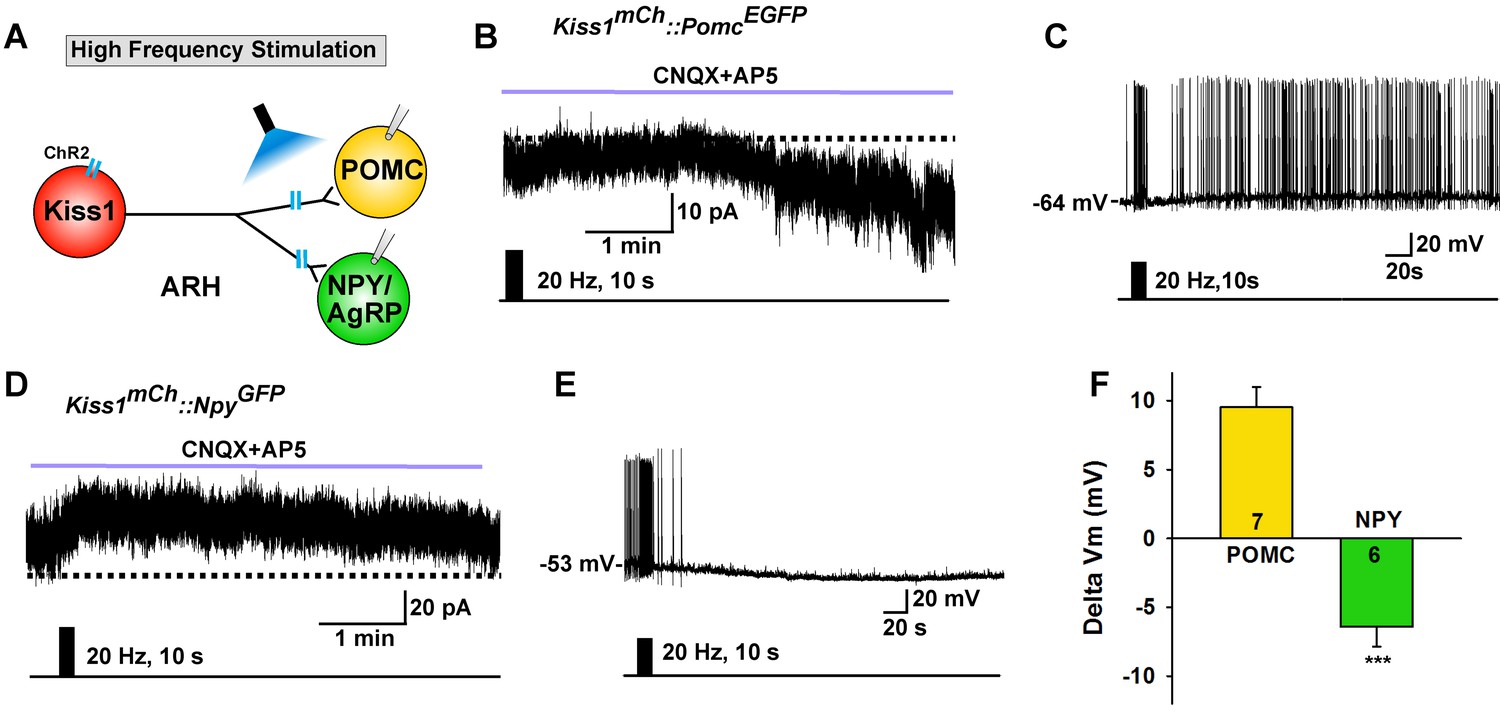
High frequency stimulation of Kiss1Cre:GFP neurons inhibits NPY neurons but excites POMC neurons.
(A) Schematic of photostimulation of the terminals of Kiss1ARH neurons and recording of POMC or NPY/AgRP neurons. (B) high-frequency optogenetic stimulation (20 Hz, 10 s) of Kiss1Cre:GFP neurons/fibers, which were labeled with AAV-DIO-ChR2-mCherry, generated a slow EPSC in POMCEGFP neurons (in the presence of CNQX, 10 μM and AP5, 50 μM). (C) in current clamp the same stimulus depolarized and increased the firing frequency of POMC neurons. (D) in voltage clamp (Vhold = −60 mV) high-frequency stimulation of Kiss1Cre:GFP-ChR2 neurons/fibers generated a slow IPSC in NPYGFP neurons (CNQX, 10 μM; AP5, 50 μM). (E) in current clamp the same stimulus hyperpolarized and inhibited the firing frequency of NPY neurons. (F) summary of the effects of high-frequency stimulation of Kiss1Cre:GFP-ChR2 neurons on POMC neurons (depolarized 9.5 ± 1.5 mV, n = 7) and on NPY neurons (hyperpolarized 6.4 ± 1.4 mV, n = 6). The responses (change in membrane potential, Delta Vm) were significantly different in POMC versus NPY/AgRP neurons (Unpaired t-test, t (11)=7.685, p<0.0001). ***p<0.001.
-
Figure 7—source data 1
Effects of high-frequency stimulation of Kiss1Cre:GFP-ChR2 neurons on POMC and NPY neurons (Figure 7F).
- https://doi.org/10.7554/eLife.35656.014
Figure 8
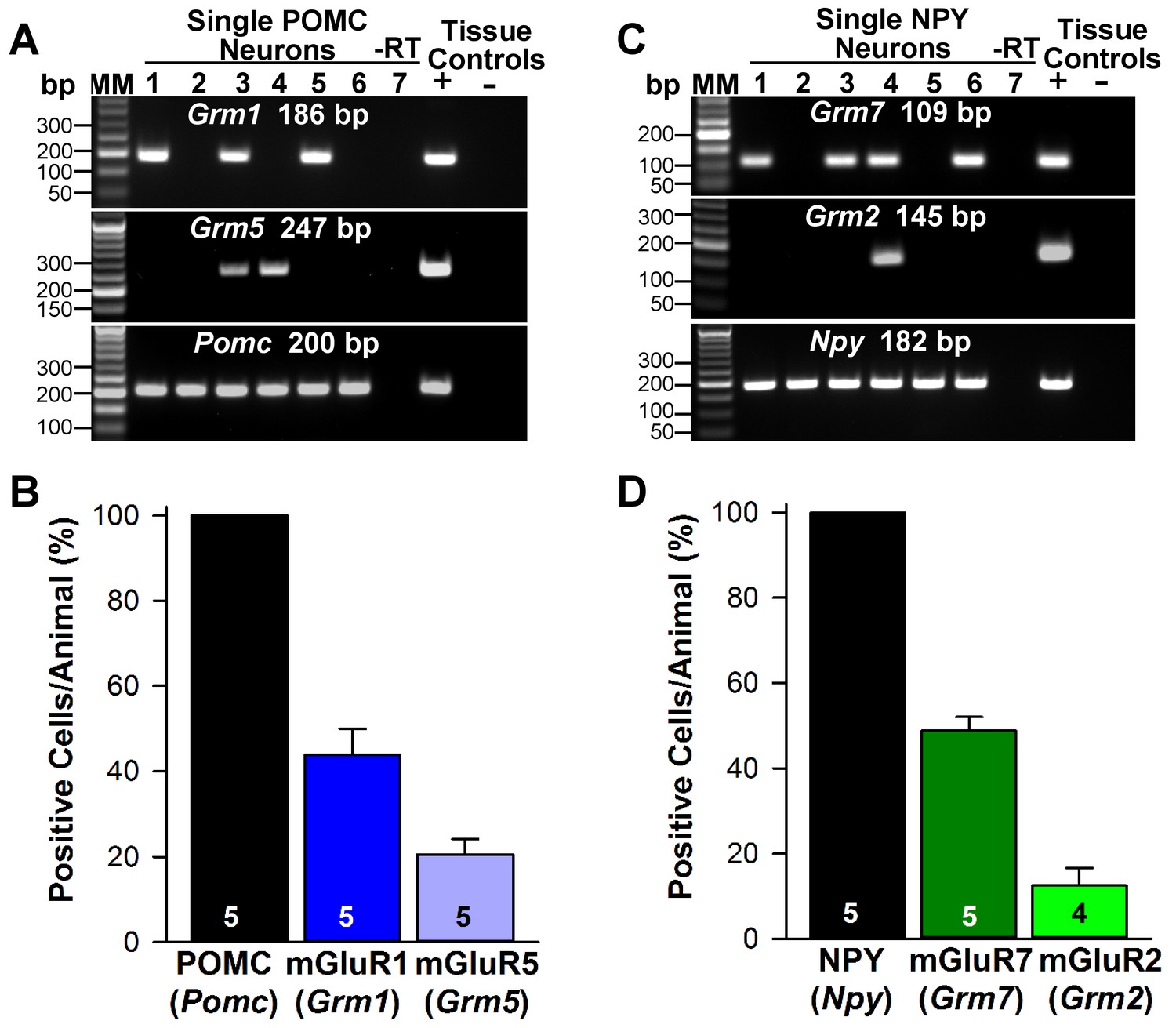
Metabotropic glutamate receptor expression in POMC and NPY neurons.
(A) representative gels illustrating mRNA expression of Pomc, Grm1 (encoding mGluR1) and Grm5 (encoding mGluR5) in individual POMCEGFP neurons. The expected base pair (bp) sizes for Pomc, Grm1 and Grm5 are 200 bp, 186 bp, 247 bp, respectively. (B) bar graphs summarizing the percentage (mean ±SEM) of POMCEGFP cells (24 cells each from 5 animals) that expressed Pomc, Grm1, and Grm5 mRNAs. (C) representative gels illustrating mRNA expression of NPY, Grm2 (encoding mGlur2) and Grm7 (encoding mGlur7) in individual NPYGFP neurons. The expected base pair (bp) sizes for Npy, Grm2 and Grm7 are 182 bp, 145 bp, 109 bp, respectively. (A,C) exclusion of reverse transcriptase (-RT) in a reacted cell was used as negative control. RNA extracted from medial basal hypothalamic tissue was also included as positive (+, with RT) and negative (-, without RT) tissue controls. (D) bar graphs summarizing the percentage (mean ± SEM) of NPYGFP cells (24 cells each from 5 animals) that expressed Npy, Grm2, and Grm7 mRNAs.
-
Figure 8—source data 1
Pomc, Grm1 and Grm5 mRNA expression in POMC neurons (Figure 8B).
Npy Grm2 and Grm7 mRNA expression in NPY neruons (Figure 8D).
- https://doi.org/10.7554/eLife.35656.016
Figure 9
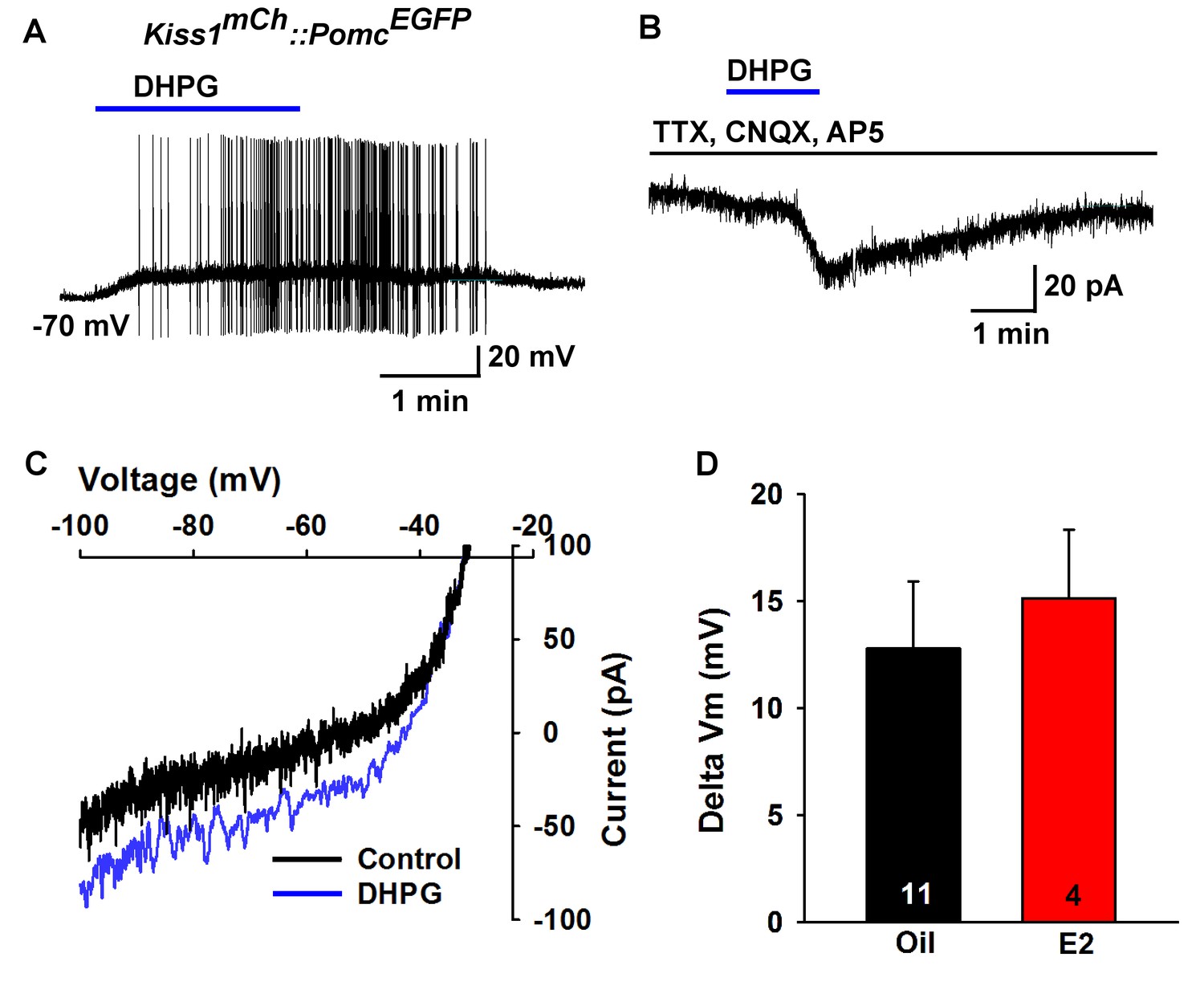
Metabotropic glutamate response is excitatory in POMC neurons.
(A) metabotropic glutamate receptor 1/5 agonist DHPG (50 µM) depolarized and induced firing of a POMC neuron. (B) Rapid bath application of DHPG induced an inward current in the presence of fast sodium channel and ionotropic glutamatergic blockade, Vhold = −60 mV. (C) voltage ramps from 0 to −100 mV were applied (over 2 s) before and during the treatment with DHPG, showed a reversal at −30 mV. (D) summary of the depolarizing effects of DHPG in POMC neurons in oil-treated and E2-treated, OVX females. There was not a significant difference in the response (Unpaired t-test, t(13) = 0.4168, p=0.6831).
-
Figure 9—source data 1
Depolarizing effects of DHPG in POMC neurons in oil-treated and E2-treated, ovariectomized females (Figure 9D).
- https://doi.org/10.7554/eLife.35656.018
Figure 10

Metabotropic glutamate response is inhibitory in NPY neurons and augmented by E2.
(A) the group II mGluR agonist DCG-IV (10 µM) hyperpolarized a NPYGFP neuron. (B) DCG-IV activated an outward current in a NPYGFP neuron in the presence of fast sodium channel (TTX, 0.5 μM), ionotropic glutamatergic (CNQX, 10 μM and AP5, 50 μM) and GABAergic (picrotoxin, 100 μM) blockers (V hold = −60 mV). (C) I-V relationship for DCG-IV- induced current showed a reversal at EK+ (−95 mV). (G) DCG-IV was more efficacious to hyperpolarize NPY neurons in E2-treated versus oil-treated, OVX females (Unpaired t-test, t(32) = 2.261, p=0.031). *p<0.05. (D) the mGluR7 allosteric agonist AMN082 (10 µM) hyperpolarized and inhibited firing of an NPYGFP neuron. (E) AMN082 generated a 25 pA outward current in a NPYGFP neuron in the presence of fast sodium channel and ionotropic glutamatergic blockade (V hold = −60 mV). (F) I-V relationship for AMN082-induced current showed a reversal close to EK+. (H) AMN082 was more efficacious than DCG-IV to hyperpolarize NPY neurons in E2-treated versus oil-treated, OVX females (Unpaired t-test, t(17) = 3.747, p=0.002). (I) Quantitative real-time PCR measurements of Grm7 mRNA in NPYGFP neuronal pools (4 pools of 5 cells each per animal) from oil- and E2-treated, OVX mice (n = 4–5 animals per group). Bar graphs represent the mean ± SEM (Unpaired t-test, t(7)=6.020, p=0.0005). ***p<0.001.
-
Figure 10—source data 1
DCG-IV was more efficacious to hyperpolarize NPY neurons in E2-treated versus oil-treated, OVX females (Figure 10G).
AMN082 was more efficacious than DCG-IV to hyperpolarize NPY neurons in E2-treated versus oil-treated (Figure 10H). Grm7 mRNA expression in NPY neurons (Figure 10I).
- https://doi.org/10.7554/eLife.35656.020
Figure 11
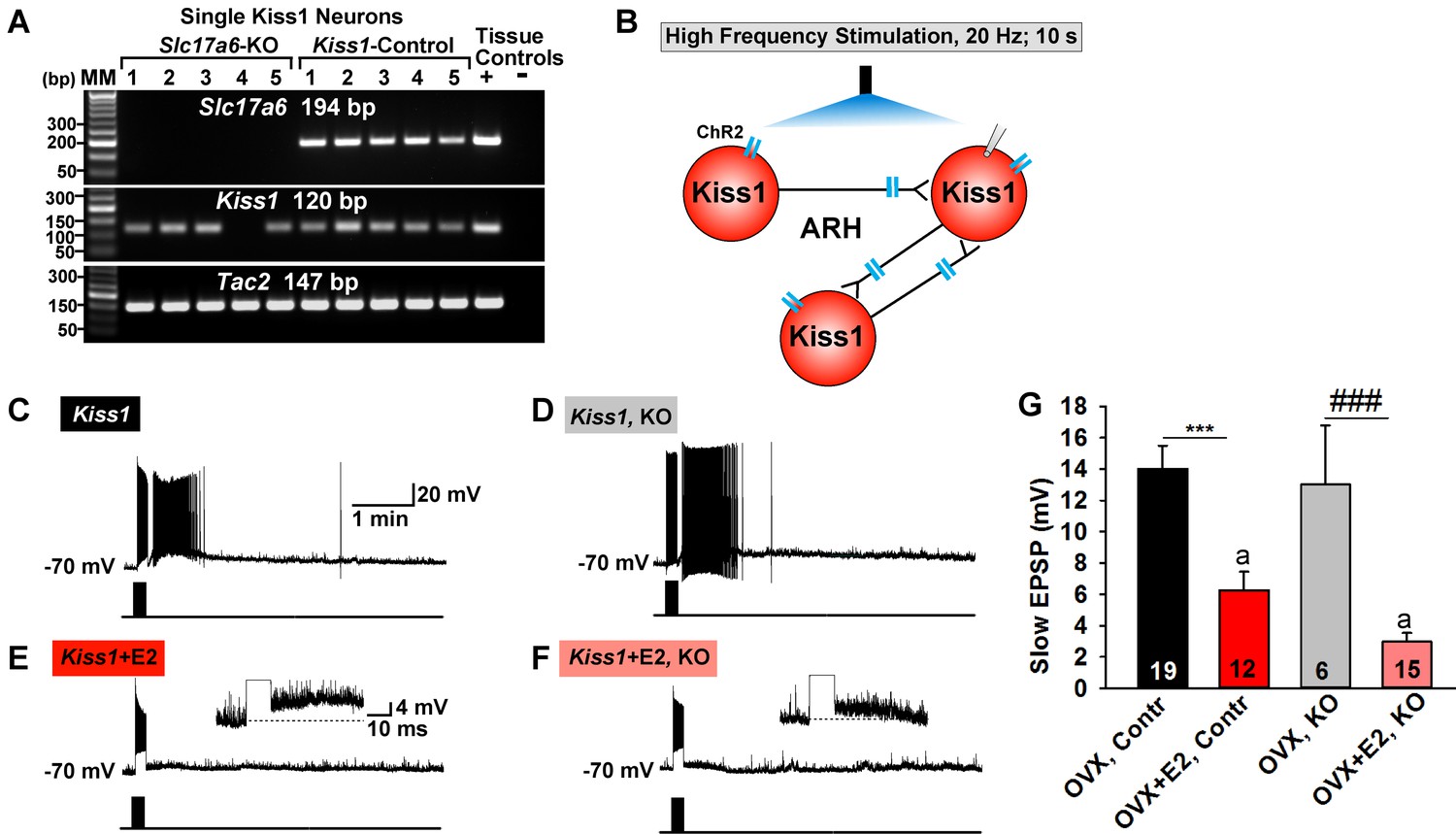
Deletion of Slc17a6 in Kiss1ARH neurons attenuates the slow EPSP in Kiss1ARH neurons from E2-treated mice.
(A) representative gels illustrating mRNA expression of Slc17a6, Kiss1 and Tac2 in Slc17a6 KO Kiss1ARH neurons and in control Kiss1ARH cells. The expected base pair (bp) sizes for Slc17a6, Kiss1 and Tac2 are 194, 120 and 147 bp, respectively. RNA extracted from the medial basal hypothalamic tissue was used as positive (+, with RT) and negative (-, without RT) tissue controls. MM, molecular marker. (B) Experimental protocol: AAV1-DIO-ChR2:mCherry was bilaterally injected into ARH of Kiss1Cre:GFP control and Slc17a6 KO mice, followed by high-frequency photostimulation of Kiss1ARH neurons/terminals and recording of Kiss1ARH neurons. (C) high-frequency optogenetic stimulation (20 Hz, 10 s) generated a slow EPSP in an arcuate Kiss1Cre:GFP neuron from OVX, control Kiss1 mice. (D), high-frequency response (slow EPSP) in arcuate Kiss1Cre:GFP neurons from OVX, Kiss1Cre:GFP::Slc17a6lox/∆ (KO) mice.(E) high-frequency response (slow EPSP) in arcuate Kiss1Cre:GFP neuron from E2-treated, OVX control Kiss1 mice. Inset shows full amplification of sEPSP. (F) high-frequency response in arcuate Kiss1Cre:GFP neuron from E2-treated, OVX Kiss1Cre:GFP::Slc17a6lox/∆ mice. Inset shows full amplification of sEPSP. (G) summary of the effects of vGluT2 deletion on slow EPSP amplitude: (one-way ANOVA, effect of treatment, F(3, 50)=14.13, p<0.0001; Newman-Keuls’ Multiple-comparison test, *** or ###, indicates p<0.005). Although knockout of vGluT2 did not significantly diminish the slow EPSP amplitude in OVX females, it did attenuate the response in E2-treated, OVX females (Unpaired t-test, t (25)=2.735, p=0.0113). a-a, p<0.05.
-
Figure 11—source data 1
Data for Figure 11G.
- https://doi.org/10.7554/eLife.35656.022
Figure 12
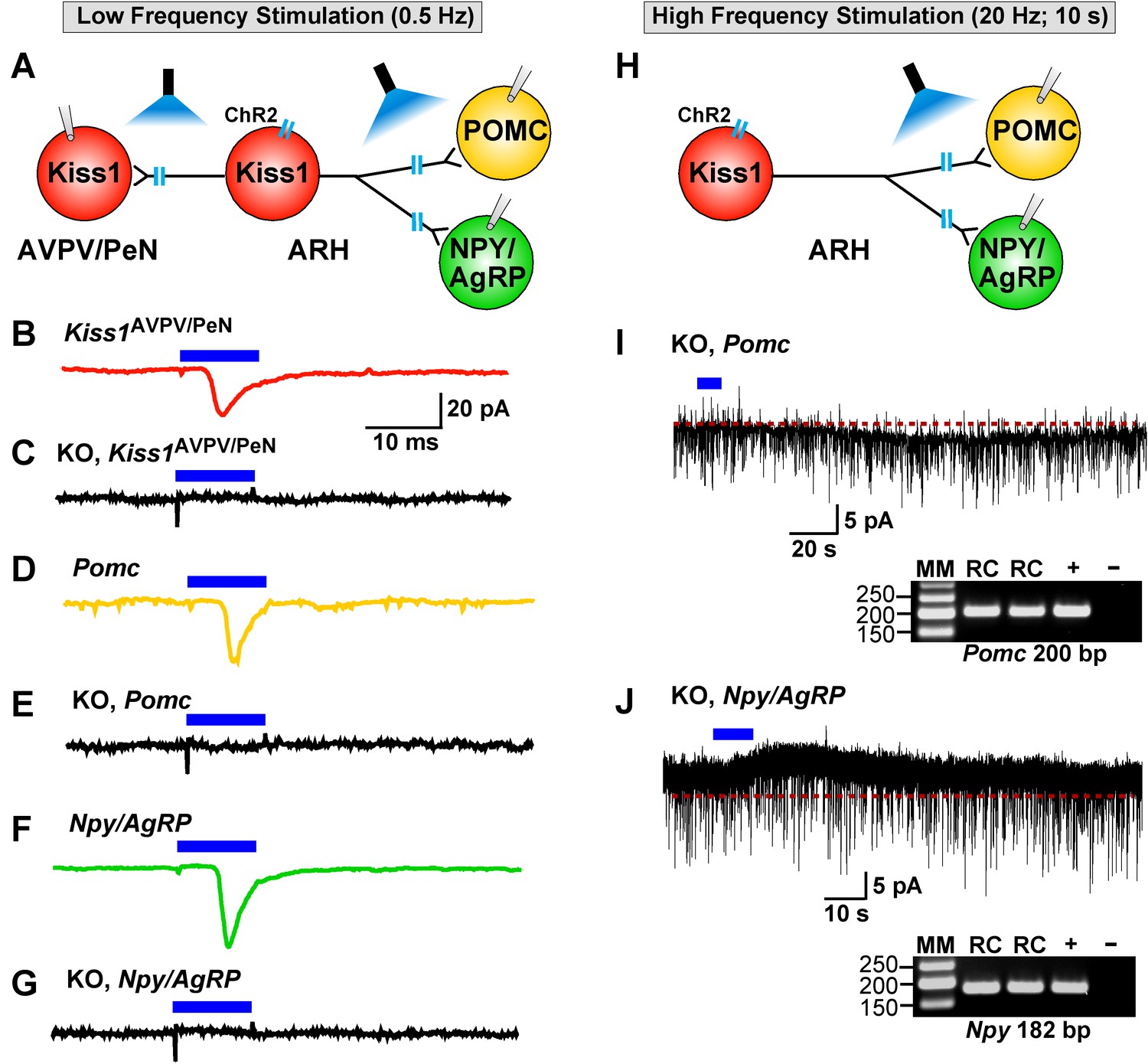
Deletion of Slc17a6 in Kiss1ARH neurons abrogates fast glutamatergic responses in Kiss1AVPV/PeN, POMC and NPY/AgRP neurons.
(A) Experimental protocol: AAV1-DIO-ChR2:mCherry (or YFP) was bilaterally injected into ARH of Kiss1Cre:GFP mice. Thereafter, low-frequency photostimulation of the terminals of Kiss1ARH neurons were done, and postsynaptic responses in Kiss1AVPV/PeN, POMC or NPY/AgRP neurons were recorded. (B,C) whole-cell, voltage clamp (Vhold = −60 mV) recordings in Kiss1AVPV/PeN neurons show that low-frequency optogenetic stimulation (0.5 Hz) evoked fast glutamatergic postsynaptic responses in control female Kiss1Cre mice (B, red trace), but failed in Kiss1AVPV/PeN cells (n = 10) from Kiss1Cre:GFP::Slc17a6lox/∆ mice (C, black trace). (D,E) and F,G) similarly, the response could be induced in POMC neurons (D, yellow trace) or NPY/AgRP neurons (F, green trace) from control Kiss1Cre:GFP mice, but abrogated in POMC neurons (E, black trace) (n = 28) or NPY/AgRP neurons (G, black trace) (n = 30) from Kiss1Cre:GFP::Slc17a6lox/∆ mice. (H) Experimental protocol: high-frequency photostimulation of the terminals of Kiss1ARH neurons and recording of POMC or NPY/AgRP neurons. (I) high-frequency stimulation (20 Hz, 10 s) of arcuate Kiss1 neurons from Kiss1Cre:GFP::Slc17a6lox/∆ mice evoked a small inward current (2.8 ± 0.5 pA, n = 7) in POMC neurons (identified post hoc by scRT-PCR, gel inset). (J) likewise, high-frequency stimulation evoked a small outward current (4.0 ± 1.6 pA, n = 4) in NPY/AgRP neurons (identified post hoc by scRT-PCR, gel inset). Insets show scRT-PCR post hoc identification of representative recorded POMC and NPY neurons. RC, recorded cells; +, positive tissue control reacted with RT; -, negative tissue control reacted without RT; MM, molecular marker.
Figure 13
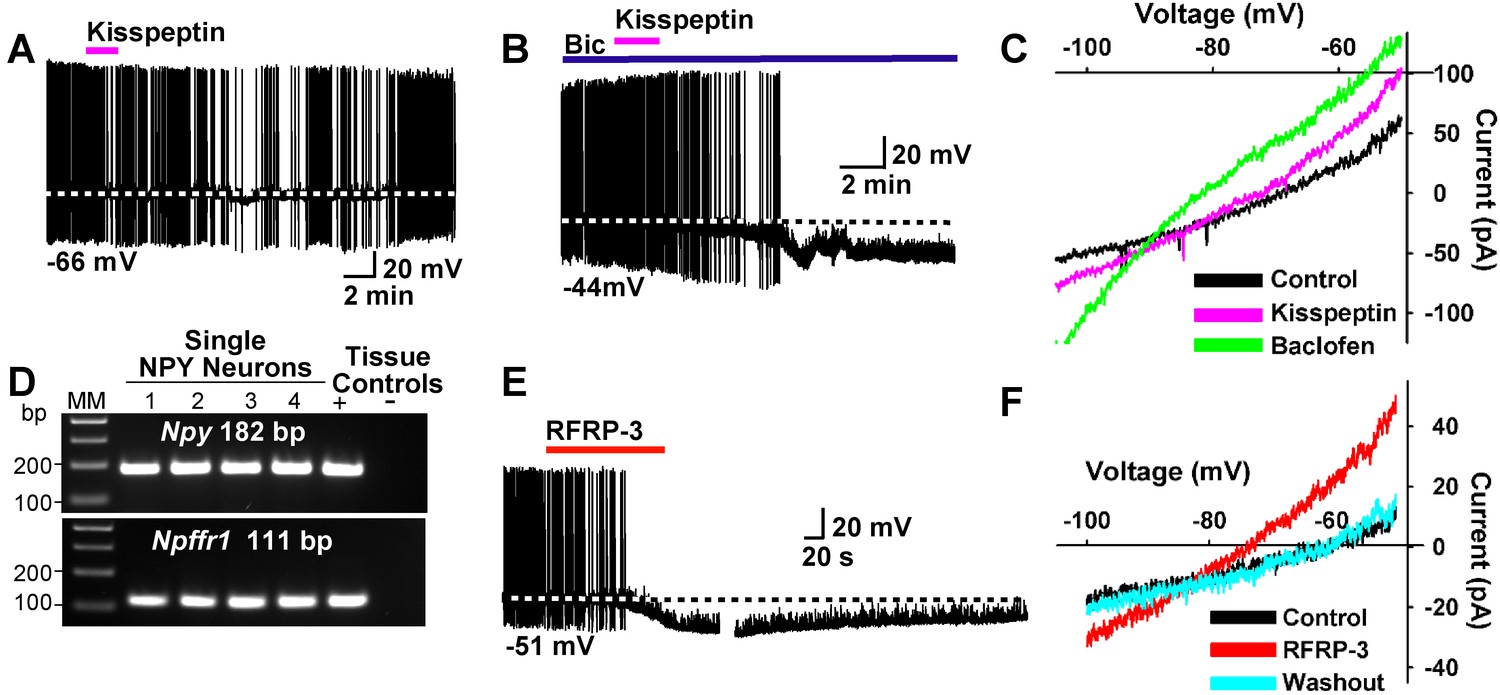
Kisspeptin and RFRP-3 inhibit NPY neurons.
(A,B) kisspeptin (200 nM) inhibited the firing and hyperpolarized NPY neurons even in the presence of GABAA blocker bicuculline (BIC) (10 μM). (C) Similar to GABAB receptor agonist baclofen (10 μM), kisspeptin induced an outward, albeit smaller, current with a reversal potential at EK+ (−90 mV). (D) scRT-PCR expression of Npffr1 in NPY/AgRP neurons. (E) RFRP-3 (10 μM), selective agonist for NPFFR1/NPFFR2, hyperpolarized and inhibited firing in NPY neurons. (F) the I/V plots of the RFRP-3 current showed a reversal potential close to EK+ (−85 mV).
Figure 14

Kisspeptin excites POMC neurons by activating a non-selective cation conductance.
(A) representative gel illustrating the scRT-PCR expression of Kiss1r (GPR54) transcript in POMC neurons. (B) kisspeptin (200 nM) depolarized and increased firing of POMC neurons (n = 8). (C) I/V (digital subtraction of control I/V from I/V with kisspeptin using a Cs+-based internal solution; see Materials and Methods) showed that kisspeptin activated a non-selective cationic channel that reversed at −10 mV.
Figure 15

Protocol for inducing CPP with sucrose.
The protocol for conditioning and preference testing consisted of four phases over the course of 11 days (sucrose habituation, a baseline place preference (BPP) test, sucrose conditioning, and a conditioned place preference (CPP) test). Food-motivated behavior was assessed during the dark cycle using an unbiased procedure. On the day before BPP, Day −1, sucrose habituation occurred where mice received sucrose pellets overnight (O/N) in their home cage to prevent neophobia. The initial BPP (black vs. white chamber) was assessed on Day 1 in a three-chamber place preference apparatus and the chamber pairing was assigned in an unbiased manner. During sucrose conditioning, mice were given access to sucrose-filled (CS+, Days 2, 4, 6, 8) or empty (CS-, Days 3, 5, 7, 9) lids on alternating days. Mice were given access to sucrose-filled lids in one chamber (e.g. white), then on alternating days they were presented with empty lids in the other chamber (e.g. black). Mice were tested for acquisition of a CPP to sucrose on Day 10, which was indicated by increased time spent in the sucrose-conditioned chamber. Animals were fed ad lib standard low-fat chow in their home cage throughout the study. For cyclical estradiol treatment, animals were given a priming (0.25 μg) and a surge (1.5 μg) dose of 17β-estradiol Benzoate (E2) at 9 AM prior to the BPP and prior to the CPP as indicated. During the sucrose-conditioning (phase 3), the animals were treated twice with a 1 μg maintenance dose of E2.
Figure 16 with 1 supplement
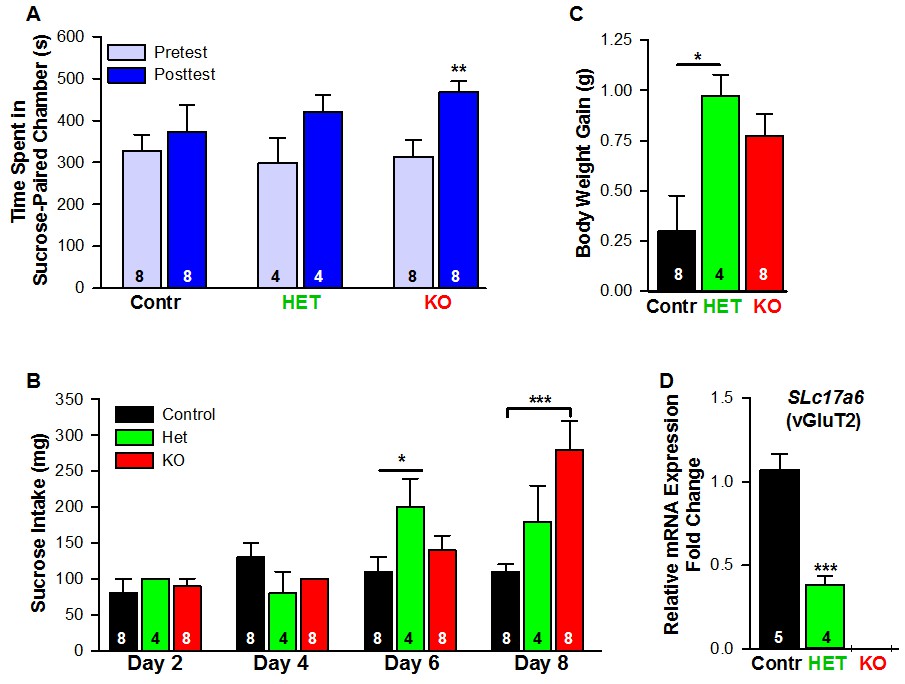
Female mice lacking Slc17a6 in Kiss1ARH neurons develop a conditioned place preference for sucrose.
(A) Time spent in sucrose-paired chamber by control Kiss1 female mice (n = 8), Slc17a6 Het (n = 4) and Slc17a6 KO Kiss1 females (n = 8) was measured during the Pretest (Day 1, Baseline Place Preference) and the Posttest (Day 10, Conditioned Place Preference). All animals were OVX and E2-treated, and had free access to standard mouse chow in their home cage throughout the study. After sucrose conditioning, Slc17a6 KO mice developed a preference for the sucrose-paired chamber (Bonferroni post hoc test, p=0.001). Slc17a6 Het mice displayed a trend to develop a preference for the sucrose-paired chamber (Bonferroni post hoc test, p=0.086). Control Kiss1Cre female mice, however, failed to develop a preference (Bonferroni post hoc test, p=0.619). [Also, see Figure 16—figure supplement 1A for comparison between E2-treated, OVX Kiss1 female and intact Kiss1 male mice]. Two-way ANOVA: main effect of experimental group (F(2,17) = 0.298, p=0.746), main effect of protocol day (F(1,17) = 20.34, p=0.0003), and interaction (F(2,17) = 2.33, p=0.128); **p<0.01.(B) Sucrose consumption during the CPP. Sucrose intake (mg) was measured during the four sucrose-paired days (Days 2, 4, 6, and 8). Slc17a6 KO mice slightly increased their sucrose intake on Day 6 and this was significantly increased by Day 8 (Bonferroni post hoc test, p<0.0001). Slc17a6 Het mice displayed a smaller, but significant increase in sucrose intake on Day 6 (Bonferroni post hoc test, p=0.0464). [Also, see Figure 16—figure supplement 1B for comparison between E2-treated, OVX Kiss1 females and intact Kiss1 males]. Two-way ANOVA: main effect of experimental group (F(2,17) = 3.788, p=0.0436), main effect of protocol day (F(3,51) = 12.75, p<0.0001), and interaction (F(6,51) = 5.763, p<0.0001). *p<0.05, Het mice versus Kiss1 control; ***p<0.001, Slc17a6 KO mice versus Kiss1 control. (C) Body weight-gain during the ten-day CPP period. Despite that both the Slc17a6 KO and Het mice gained weight in comparison to control Kiss1 mice, only Slc17a6 Het mice were significantly different (Bonferroni post hoc test, p=0.0312, Slc17a6 Het vs Kiss1 control; p=0.066, Slc17a6 KO vs Kiss1 control;). One-way ANOVA: main effect of experimental group (F(2,17) = 5.232, p=0.017). *p<0.05, Het mice versus Kiss1 control. (D) Quantitative real time PCR measurement of Slc17a6 in Kiss1ARH neuronal pools from control Kiss1Cre:GFP mice (5 Kiss1 neurons in each pool and 5 pools from each of 5 animals) and Slc17a6 Het Kiss1 mice (5 Kiss1 neurons in each pool and 5 pools from each of 4 animals). Slc17a6 KO Kiss1 mice did not express Slc17a6 in Kiss1ARH neurons. (Unpaired t-test, t(7) = 5.791, p=0.0007). ***p<0.001, Het mice versus Kiss1 control.
-
Figure 16—source data 1
CPP Time Spent: Ovx Kiss2 Female Mice (n = 8); Ovx HET Female Mice (n = 4); Ovx KO Female Mice (n = 8) for Figure 16A.
CPP Sucrose intake: Ovx Kiss2 Control Female Mice (n = 8); Ovx HET Female Mice (n = 4); Ovx KO Female Mice (n = 8) for Figure 16B. CPP Weight Gain (Day 10 Day 1): Ovx Kiss2 Control Female Mice (n = 8); Ovx HET Female Mice (n = 4); Ovx KO Female Mice (n = 8) for Figure 16C. Slc17a6 mRNA expression in Kiss1-ARH neurons for Figure 16D.
- https://doi.org/10.7554/eLife.35656.030
Figure 16—figure supplement 1
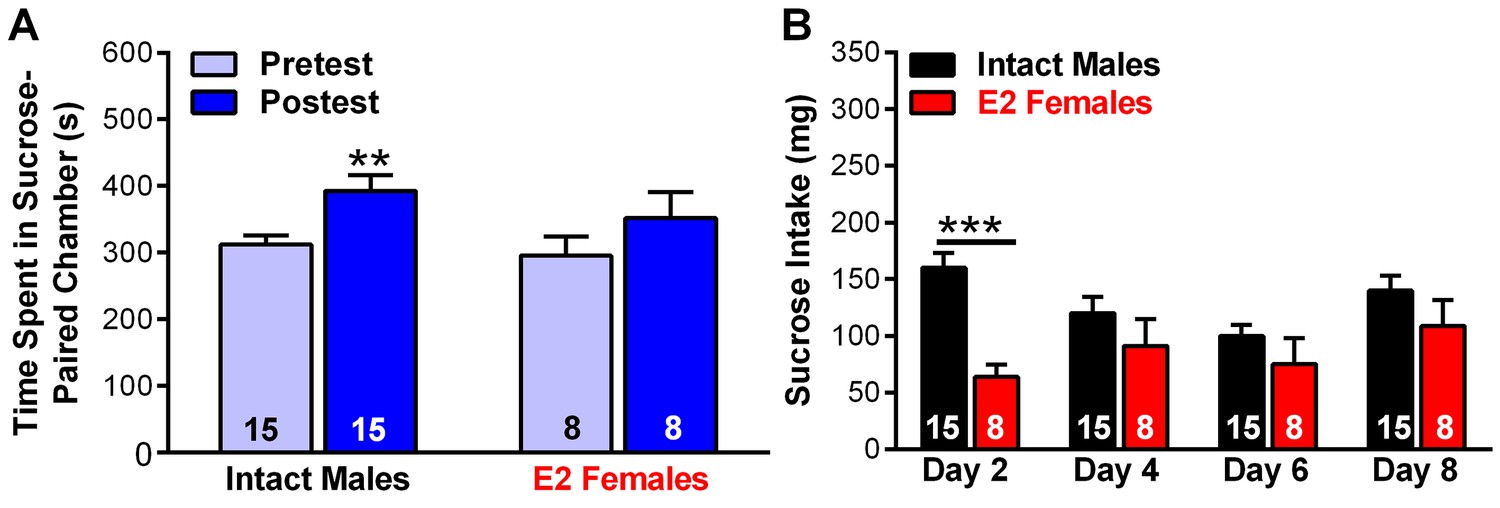
Studies documenting that intact male mice develop a conditioned place preference for sucrose.
(A) Time spent in sucrose-paired chamber by intact Kiss1Cre male mice (n = 15) and E2-treated, OVX Kiss1Cre female mice (n = 8) was measured during the Pretest (Day 1, Baseline Place Preference; BPP) and the Posttest (Day 10, Conditioned Place Preference; CPP). Male mice did not receive any treatment, whereas female mice were OVX and E2-treated. All animals were handled daily prior to the BPP/CPP tests. After sucrose conditioning intact Kiss1 males, which expressed lower levels of vGluT2 in Kiss1ARH neurons in comparison to E2-treated, OVX females, developed a preference for the sucrose-paired chamber (Bonferroni post hoc test, p<0.01). E2-treated, OVX Kiss1 female mice failed to develop a preference for the sucrose-paired chamber (Bonferroni post hoc test, p=0.124). Two-way ANOVA: main effect of experimental group (F(1, 21)=0.823, p=0.375), main effect of protocol day (F(1, 21)=14.89, p<0.001), and interaction (F(1, 21)=0.457, p=0.506); **p<0.01. (B) Sucrose consumption (mg) during the CPP was measured during the four sucrose-paired days (Days 2, 4, 6, and 8). The intact males consumed significant more sucrose on Day 2 of the test as compared to E2-treated, OVX females. Two-way ANOVA: main effect of experimental group (F(1, 21)=8.373, p<0.01), main effect of protocol day (F(3,63) = 2.49, p=0.068), and interaction (F(3,63) = 3.062, p<0.05); ***p<0.001. During this time, the animals had free access to standard low-fat mouse chow in their home cage.
-
Figure 16—figure supplement 1—source data 1
Intact Kiss2 Male Mice (n = 15) and Kisspeptin E2 Female Mice (n = 8) for panels A and B.
- https://doi.org/10.7554/eLife.35656.029
Figure 17
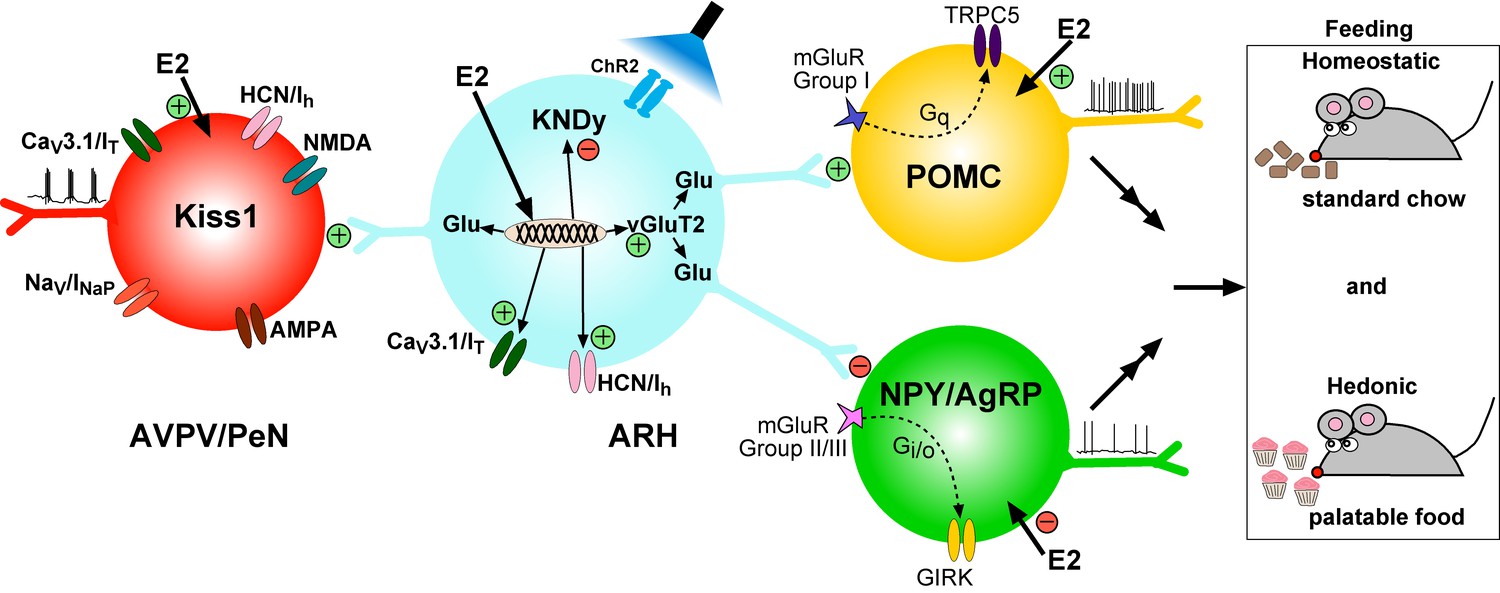
Working Model. KNDy (Kisspeptin, NKB, Dynorphin) neurons in the ARH express CaV3 (IT) and HCN (Ih) channels (currents) that are upregulated by E2 and contribute to increased excitability of Kiss1ARH neurons.
Kiss1AVPV/PeN neurons also express CaV3 (IT), HCN (Ih) and Nav (INaP) channels that are highly up-regulated by E2 along with Kiss1 mRNA expression. Notably, E2 induces spontaneous, repetitive burst firing activity in Kiss1AVPV/PeN neurons necessary for the release of GnRH (Wang et al., 2016; Zhang et al., 2015). E2 also directly excites POMC neurons via inhibition of GIRK current, but inhibit NPY/AgRP neurons via activation of GIRK current (Kelly and Rønnekleiv, 2015). These congruent actions of E2 on POMC and NPY/AgRP neurons contribute to the control of homeostatic feeding. High frequency photo-stimulation (focal light stimulation of channel rhodopsin, ChR2) in Kiss1ARH neurons releases glutamate to further excite POMC neurons via mGluRs group I and inhibit NPY/AgRP neurons via mGluRs group II/III; and excite Kiss1AVPV/PeN neurons via NMDA/AMPA receptors. Ablating Slc17a6 from Kiss1ARH neurons, results in the abrogation of glutamate release onto POA and ARH neurons. The lack of glutamate release from Kiss1ARH neurons appears to have little or no effect on estrous cyclicity, an indication that the direct effects of E2 to increase the excitability of Kiss1AVPV/PeN neurons is sufficient to drive the reproductive cycle. However, E2-treated Slc17a6 KO Kiss1 mice develop a condition place preference for sucrose indicative of positive motivational effect of sucrose in these females.
Tables
Key resources table
| Reagent type (species)or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Strain, strain background (M.Musculus) | C57BL/6J | The Jackson laboratory | RRID :IMSR_JAX:000664 | |
| Genetic reagent (M.Musculus) | Kiss1Cre:GFP | Dr. Robert A Steiner; University of Washington; PMID:21933870 | RRID:IMSR_JAX:017701 | Full nomenclature: Kiss1tm1.1(cre/EGFP)Stei |
| Genetic reagent (M.Musculus) | Kiss1Cre:GFP version 2 (V2) | Dr. Richard D. Palmiter; University of Washington; PMID: 29336844 | ||
| Genetic reagent (M.Musculus) | PomcEGFP | Dr. Malcolm J. Low; University of Michigan; PMID:11373681 | RRID:IMSR_JAX:009593 | Full nomenclature: Tg(Pomc-EGFP) 1Low |
| Genetic reagent (M.Musculus) | NpyGFP | Dr. Brad Lowell; Harvard University; PMID:19357287 | JAX stock #006417 | Full nomenclature: Tg(Npy-hrGFP) 1Lowl |
| Genetic reagent (Adeno- associated virus) | AAV1-Ef1α-DIO- ChR2:YFP | Dr. Stephanie L. Padilla; University of Washington; PMID: 25429312 | ||
| Genetic reagent (Adeno- associated virus) | AAV1-Ef1α-DIO- ChR2: mCherry | Dr. Stephanie L. Padilla; University of Washington; PMID: 25429312 | ||
| Antibody | Anti-mCherry (rabbit polyclonal) | Abcam | Abcam: ab167453 RRID:AB_2571870 | (1:10,000) |
| Antibody | Anti-kisspeptin (rabbit polyclonal) | Dr. Alain Caraty Universite Francois -Rabelais Tours; PMID:16621281 | No. 564 AB_2622231 | (1:2500) |
| Antibody | Goat anti-rabbit conjugated to Alexa 594 | Life Technologies (ThermoFisher) | Cat. No.: # A-11037 RRID: AB_2534095 | (1:500) |
| Antibody | Goat anti-rabbit conjugated to Alexa 488 | Life Technologies (ThermoFisher) | Cat. No.: # A-11034 RRID: AB_2576217 | (1:500) |
Table 1
Primer Table
https://doi.org/10.7554/eLife.35656.032| Gene name (encodes for) | Accession Number | Primer Location (nt) | Product Length (bp) | Annealing Temp (°C) | Efficiency Slope | Efficiency r2 | Efficiency % |
|---|---|---|---|---|---|---|---|
| Kiss1 (Kiss1)a,b | NM_178260 | 64–80 167–183 | 120 | 57a, 60b | −3.410 | 0.989 | 97 |
| Pomc (POMC)a | NM_008895 | 145–164 327–344 | 200 | 60.5 | |||
| Npy (NPY)a | NM_023456 | 106–125 268–287 | 182 | 60 | |||
| Grm1 (mGluR1)a | NM_001114333 | 2044–2063 2210–2229 | 186 | 59 | |||
| Grm2 (mGluR2)a | NM_001160353 | 2448–2466 2574–2592 | 145 | 59 | |||
| Grm5 (mGluR5)a | NM_01143834 | 1436–1453 1663–1682 | 247 | 59 | |||
| Grm7 (mGluR7)a,b | NM_001346640 | 1354–1373 1445–1462 | 109 | 55a, 60b | −3.306 | 0.985 | 100 |
| Npffr1 (Npffr1)a | NM_001177511 | 360–378 450–470 | 111 | 55 | |||
| Kiss1r (GPR54)a | NM_053244 | 1900–1917 2125–2144 | 245 | 60 | |||
| Slc17a6 (vGluT2)c | NM_080853 | 1038–1056 1213–1231 | 194 | 57 | |||
| Slc17a6 (vGluT2)b | NM_080853 | 872–889 967–984 | 113 | 60 | −3.293 | 0.920 | 100 |
| Slc32a1 (vGAT)b | NM_009508 | 813–834 928–949 | 137 | 60 | −3.290 | 0.906 | 100 |
| Pdyn (Dyn)b | NM_018863 | 210–228 345–363 | 154 | 60 | −3.516 | 0.990 | 93 |
| Tac2 (NKB)a,b | NM_009312 | 79–97 207–225 | 147 | 60 | −3.324 | 0.992 | 100 |
| Tacr3 (Tacr3)b | NM_021382 | 764–783 864–883 | 120 | 60 | −3.504 | 0.911 | 93 |
| Cacna1g (Cav 3.1)b | NM_009783 | 5004–5025 5060–5083 | 80 | 60 | −3.372 | 0.968 | 98 |
| Hcn1 (HCN1)b | NM_010408 | 1527–1546 1641–1662 | 136 | 60 | −3.253 | 0.958 | 100 |
| Hcn2 (HCN2)b | NM_008226 | 1122–1143 1199–1218 | 97 | 60 | −3.279 | 0.969 | 100 |
| Gapdh (GAPDH)b | NM_008084 | 689–706 764–781 | 93 | 60 | −3.352 | 0.998 | 99 |
| Actb (β-actin)b | NM_007393 | 446–465 535–555 | 110 | 60 | −3.465 | 0.996 | 95 |
-
aprimers used for scRT-PCR.
bprimers used for qPCR.
-
cprimers used to confirm the vGluT2-KO using scRT-PCR.
Additional files
-
Transparent reporting form
- https://doi.org/10.7554/eLife.35656.033
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Estrogenic-dependent glutamatergic neurotransmission from kisspeptin neurons governs feeding circuits in females
eLife 7:e35656.
https://doi.org/10.7554/eLife.35656




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}