TLR5 participates in the TLR4 receptor complex and promotes MyD88-dependent signaling in environmental lung injury
- National Institute of Environmental Health Sciences, United States
- West Virginia University, United States
- Baylor College of Medicine, United States
- North Carolina State University, United States
- First Affiliated Hospital of Harbin Medical University, China
- Lerner Research Institute, Cleveland Clinic Foundation, United States
- Duke University Medical Center, United States
- Jagiellonian University, Poland
- East Carolina University Brody School of Medicine, United States
Figures
Figure 1 with 2 supplements
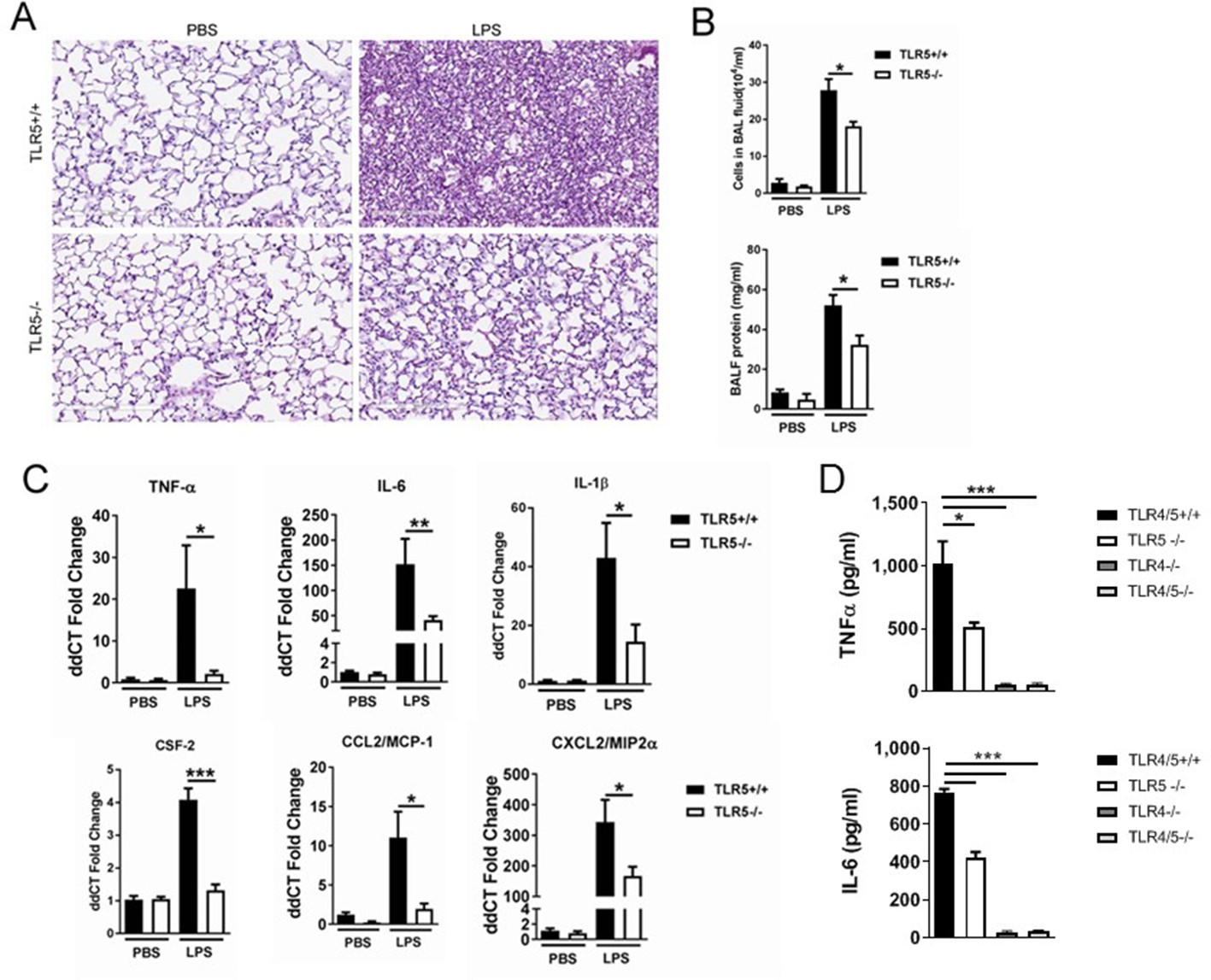
TLR5 deficiency ameliorates the inflammatory lung response to systemic LPS at 24 hr after exposure.
(A) Hematoxylin-Eosin staining of lung sections demonstrates LPS-induced lung injury is ameliorated in Tlr5-deficient (Tlr5-/-) mice. (B) Cellular lung inflammation and lung lavage protein levels are decreased in Tlr5-deficient (Tlr5-/-) mice. (C) Real time quantitative PCR analysis of inflammatory cytokines shows a significant decrease in Tlr5-deficient (Tlr5-/-) mice. N = 5–8 mice per group, experiment repeated twice. (D) Lung lavage cytokine analysis confirms a significant decrease in inflammatory cytokines after ultrapure LPS exposure in Tlr5-deficient (Tlr5-/-) mice. Note absent response in Tlr4-/- and Tlr4/5-/-. N = 4–7 mice per group. Data are represented as mean ± s.e.m. and were analyzed by unpaired t test with Welch's correction *p<0.05 ** p<0.01 and ***p<0.001.
Figure 1—figure supplement 1
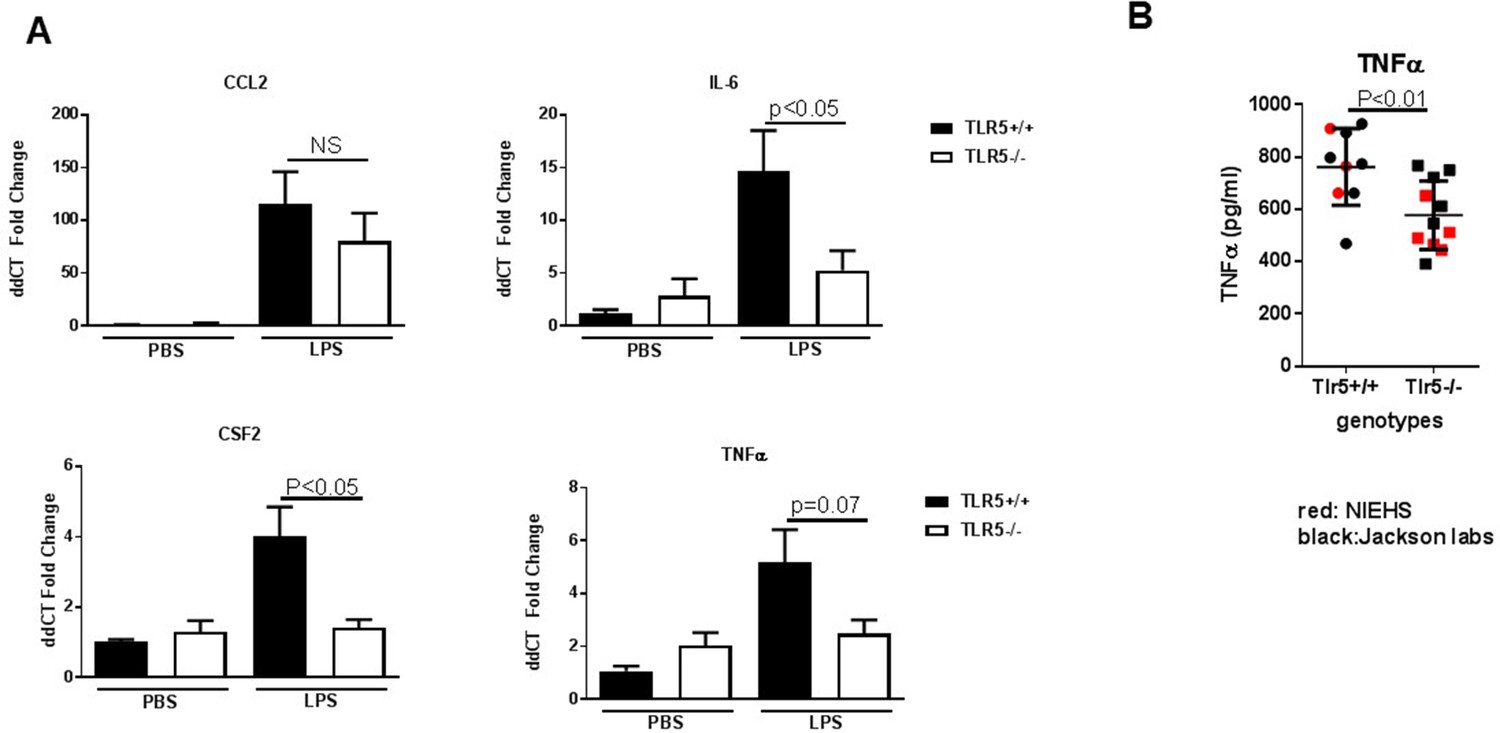
TLR5 deficiency ameliorates the inflammatory response to LPS exposure.
Figure 1—figure supplement 2
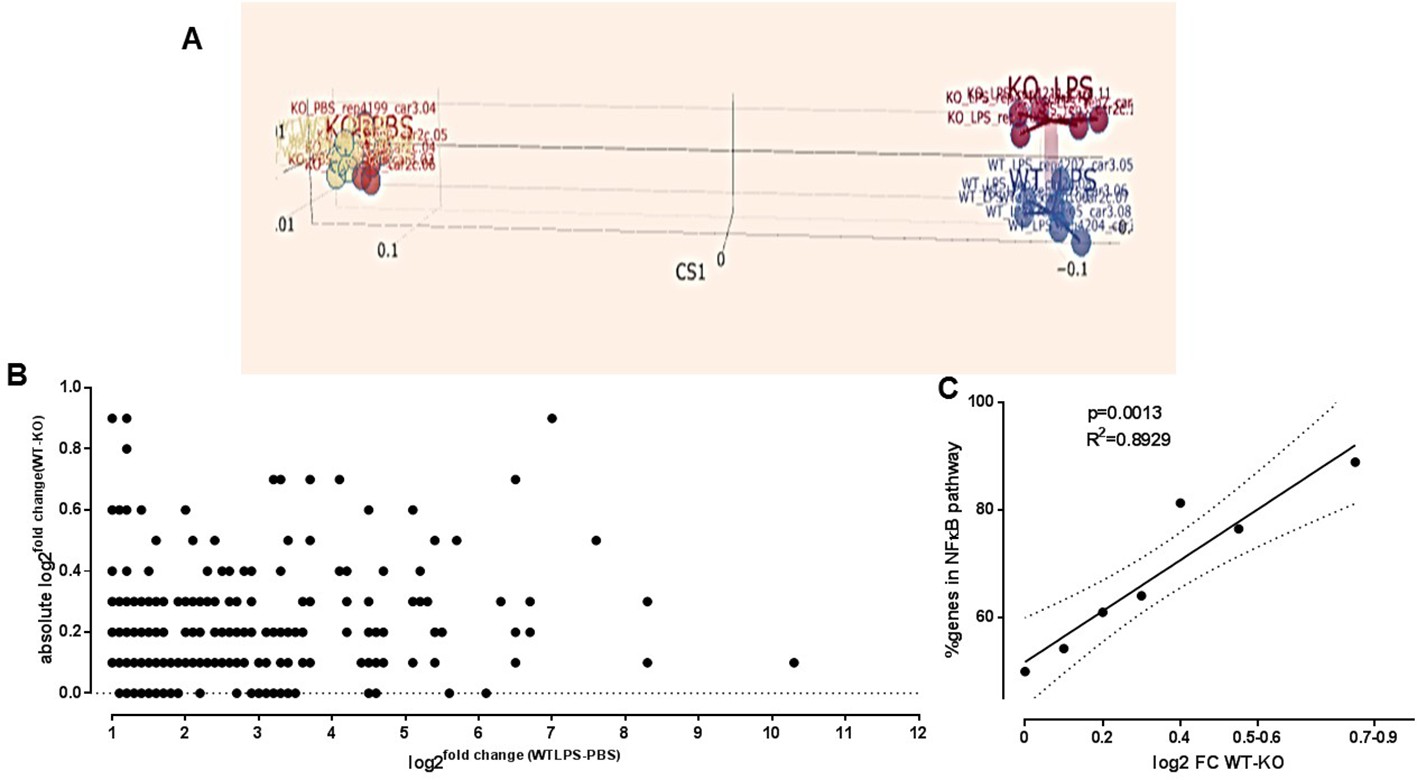
Immune gene expression profiling dependent on TLR5 after ultrapure LPS exposure.
Figure 2
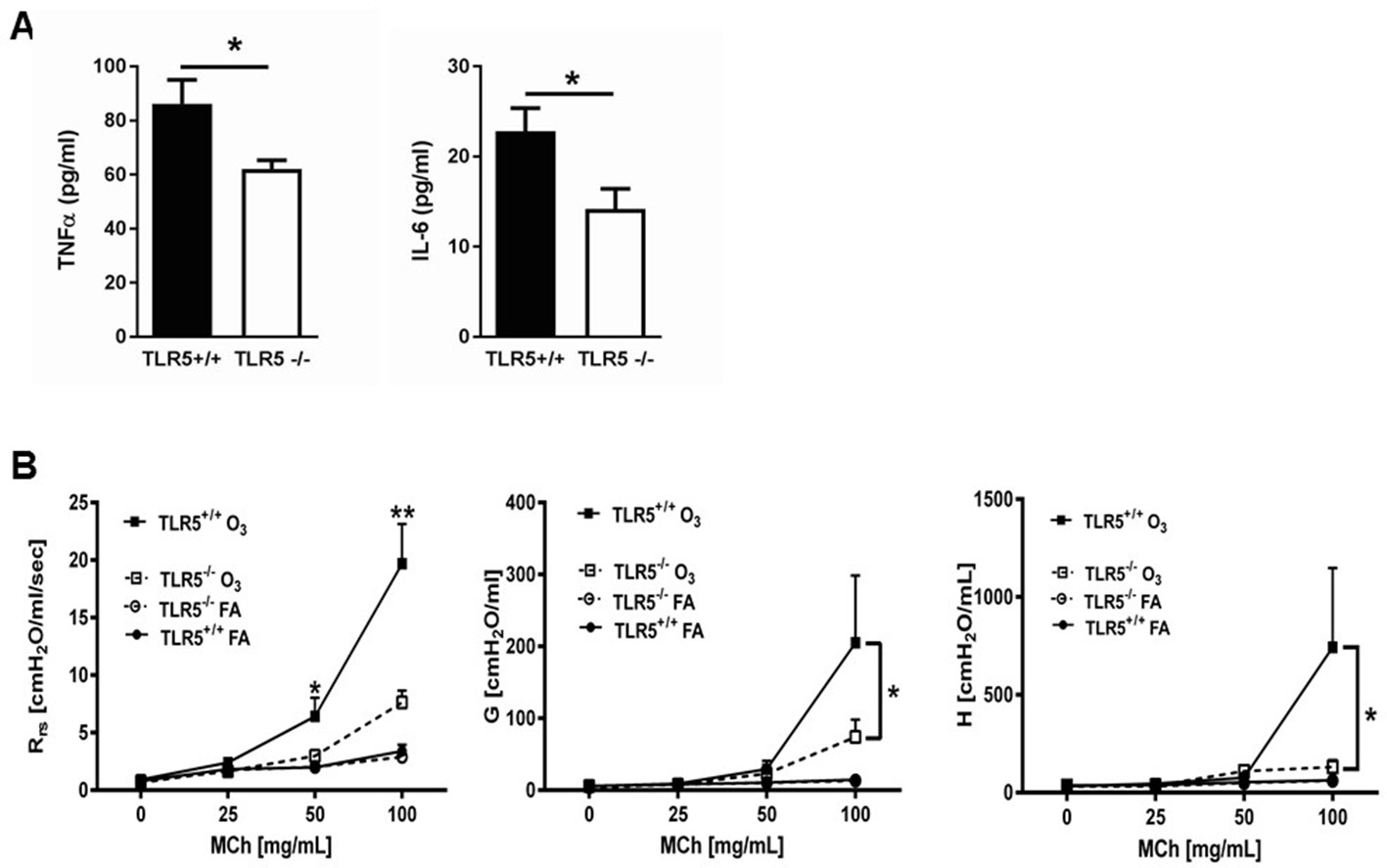
TLR5 deficiency ameliorates the in vivo inflammatory response to inhaled O3.
(A) TNF-α and IL-6 levels in the lung lavage fluid of Tlr5-deficient (Tlr5-/-) or Tlr5-competent (Tlr5+/+) mice 24 hr after receiving 3 ppm O3 for 3 hr by inhalation. n = 14 mice for Tlr5+/+ and n = 12 mice for Tlr5-/-, experiment repeated twice. (B) Airway physiology measurement (total respiratory resistance Rrs, tissue damping G and tissue elastance H) to indicated doses of methacholine challenge measured with flexiVent in Tlr5-deficient (Tlr5-/-) or Tlr5-competent (Tlr5+/+) mice 24 hr after 2ppm O3 or air (FA) exposure. n = 6 for Tlr5-/-FA and Tlr5-/-O3 and n = 7 for Tlr5+/+FA and TLlr5+/+O3, experiment repeated three times. Data are represented as mean ± s.e.m. and were analyzed by unpaired t test with Welch's correction *p<0.05 and **p<0.01 between Tlr5+/+ and Tl r5-/- mice exposed to O3.
Figure 3
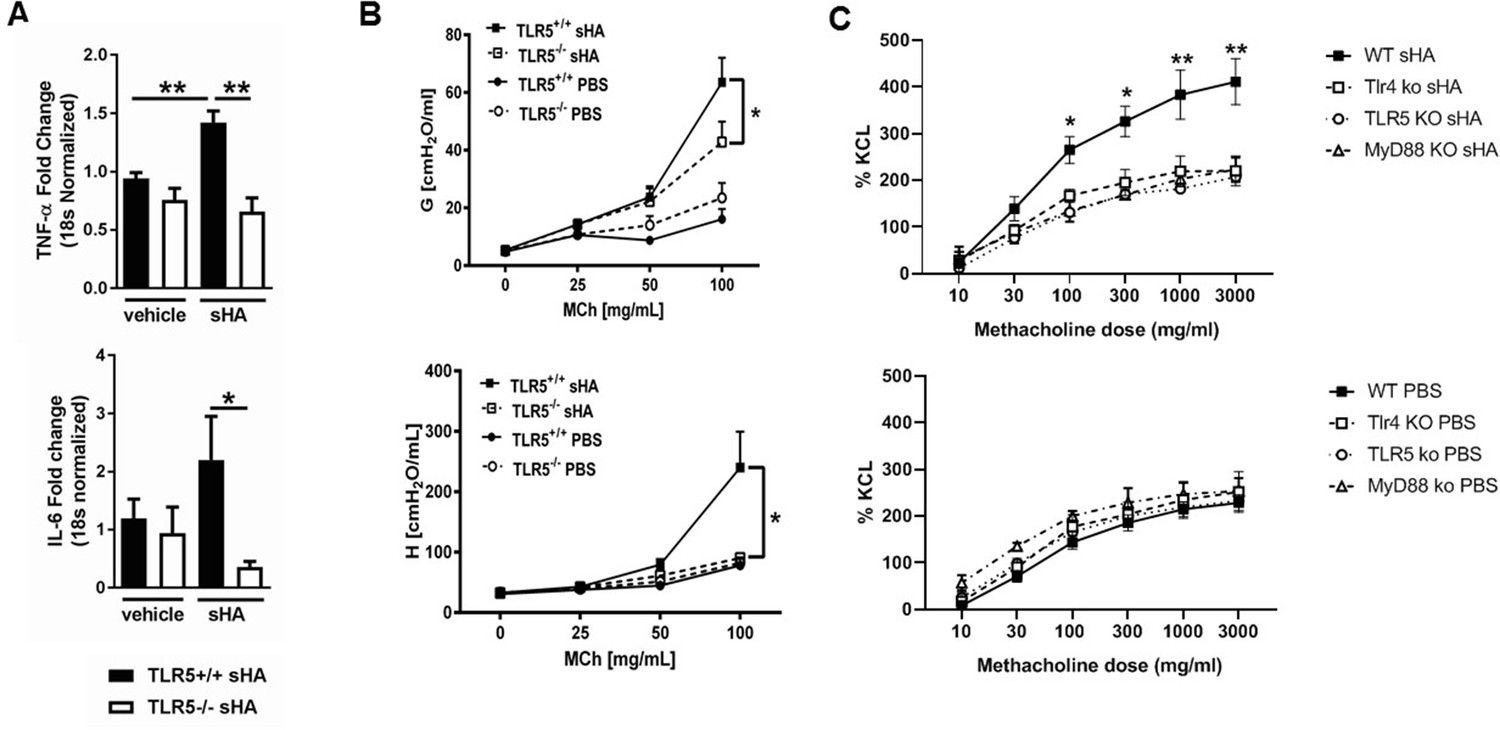
TLR5 deficiency ameliorates the in vivo and in vitro inflammatory and airway constrictive response to ultrapure short-fragment hyaluronan (sHA).
(A) Real time quantitative PCR gene expression of TNF-α and IL-6 in the lung tissues of Tlr5-deficient (TLR5-/-) or Tlr5-competent (Tlr5+/+) mice 6 hr after exposure to vehicle (PBS) or 50 µl of 3 mg/ml short fragment HA. n = 5 for Tlr5-/-PBS and Tlr5-/-sHA and n = 6 for Tlr5+/+PBS and Tlr5+/+sHA. Experiment repeated once. (B) Airway physiology measurement (tissue damping G and tissue elastance H) to indicated doses of methacholine challenge measured with flexiVent 2 hr after exposure to vehicle (PBS) or 2 mg/ml sHA. n = 5 for Tlr5-/-PBS and Tlr5-/-sHA and n = 6 for TlrR5+/+PBS and Tlr5+/+sHA, experiment repeated twice. (C). Constriction measurement of tracheal rings of C57Bl/6J mice (N = 4–6 per condition), incubated in vitro with sHA, which is known to cause hyperresponsiveness to methacholine and then treated with indicated doses of methacholine. Note that deficiency in Tlr4 and Tlr5 abolishes the tracheal ring response to hyaluronan to levels similar with Myd88 deficiency, suggesting that Tlr4 and Tlr5 play non-redundant roles in hyaluronan-Myd88 signaling. Baseline responsiveness to methacholine is not different between these strains (bottom panel C). Data are represented as mean ± s.e.m. and were analyzed by unpaired t test with Welch's correction (panel A) or ANOVA with Tukey’s post-hoc analysis (panels B and C) *p<0.05 and *p<0.01 between groups.
Figure 4 with 1 supplement
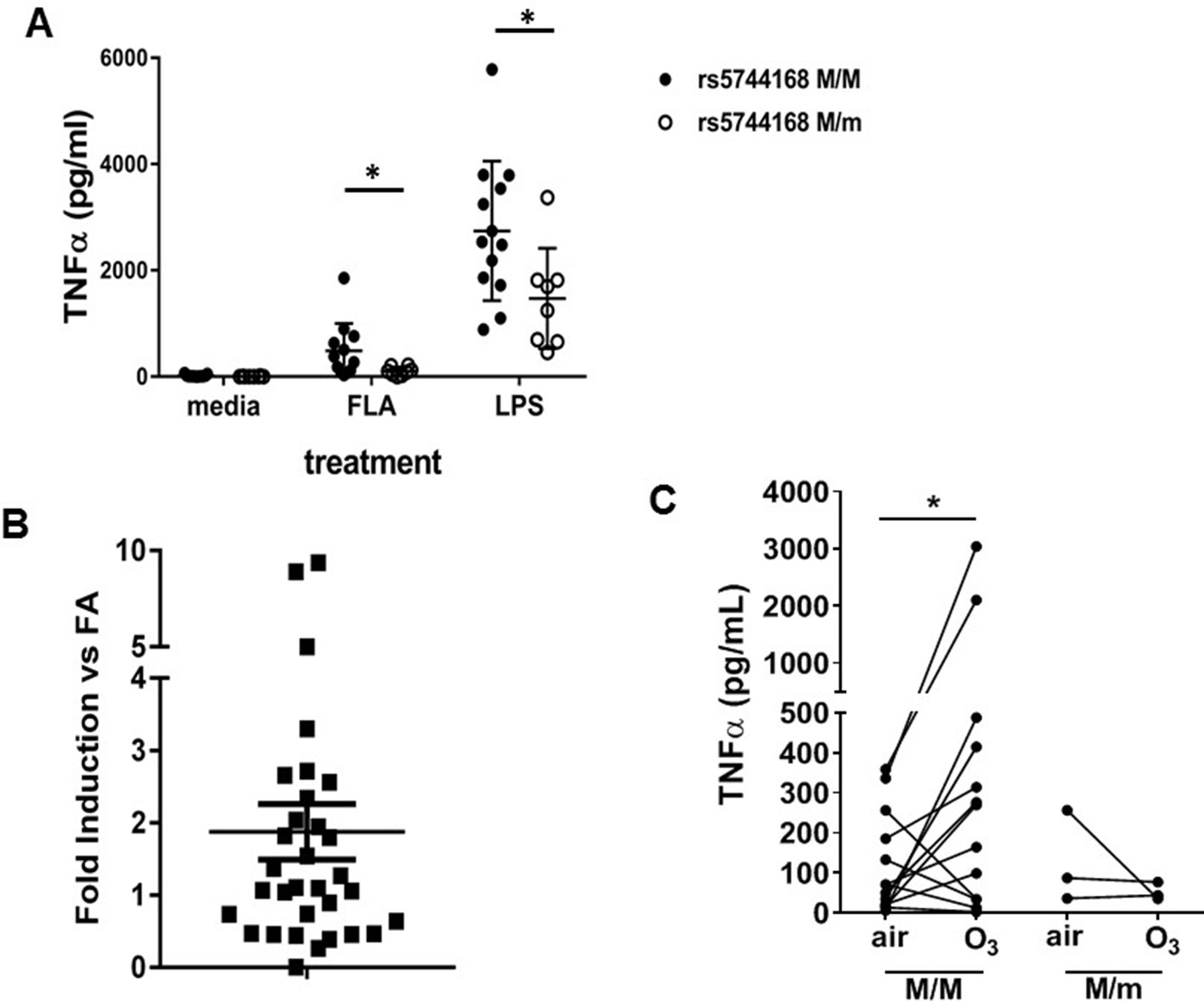
TLR5 participates in TLR4-mediated inflammation in humans.
(A) TNF-α production by peripheral blood monocyte-derived macrophages from human volunteers either homozygous for the major allele (rs5744168 M/M) or carriers of the minor allele (rs5744168 M/m) for the TLR5 single nucleotide polymorphism rs5744168. Cells were exposed to 10 ng/ml ultrapure LPS or 100 ng/ml ultrapure flagellin for 24 hr and TNF-α levels were analyzed by Duoset ELISA kit. Data are represented as mean ± standard deviation and analyzed by unpaired t test with Welch's correction. N = 7–13 individual subjects. (B) TLR5 gene expression in alveolar macrophages from human volunteers exposed to 200 ppb O3 for 135 min, referenced to TLR5 expression in the same individuals after air exposure. N = 32 individual subjects. Data are presented as individual values with mean ± s.e.m. and was analyzed by Wilcoxon pairwise signed rank test. (C) Ex vivo TNF-α production by human alveolar macrophages after exposure to air or O3 n = 3 minor rs5744168 allele carriers and 20 major allele carriers. Data are represented as individual values and trends and analyzed by Wilcoxon matched-pairs signed rank test. *p<0.05.
Figure 4—figure supplement 1
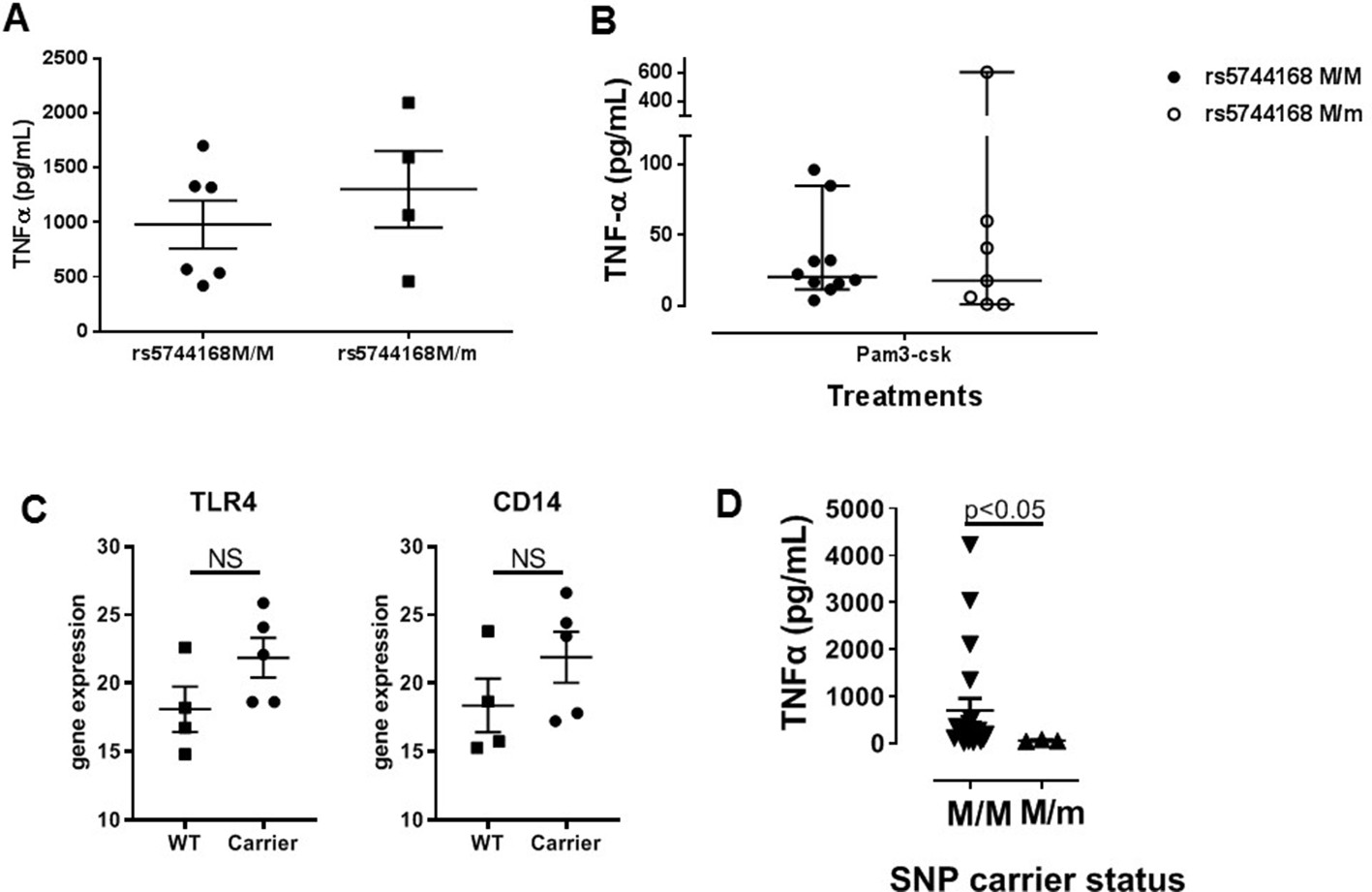
TNFa secretion from monocyte-derived macrophages from human subjects depending on TLR5 status.
Figure 5 with 1 supplement
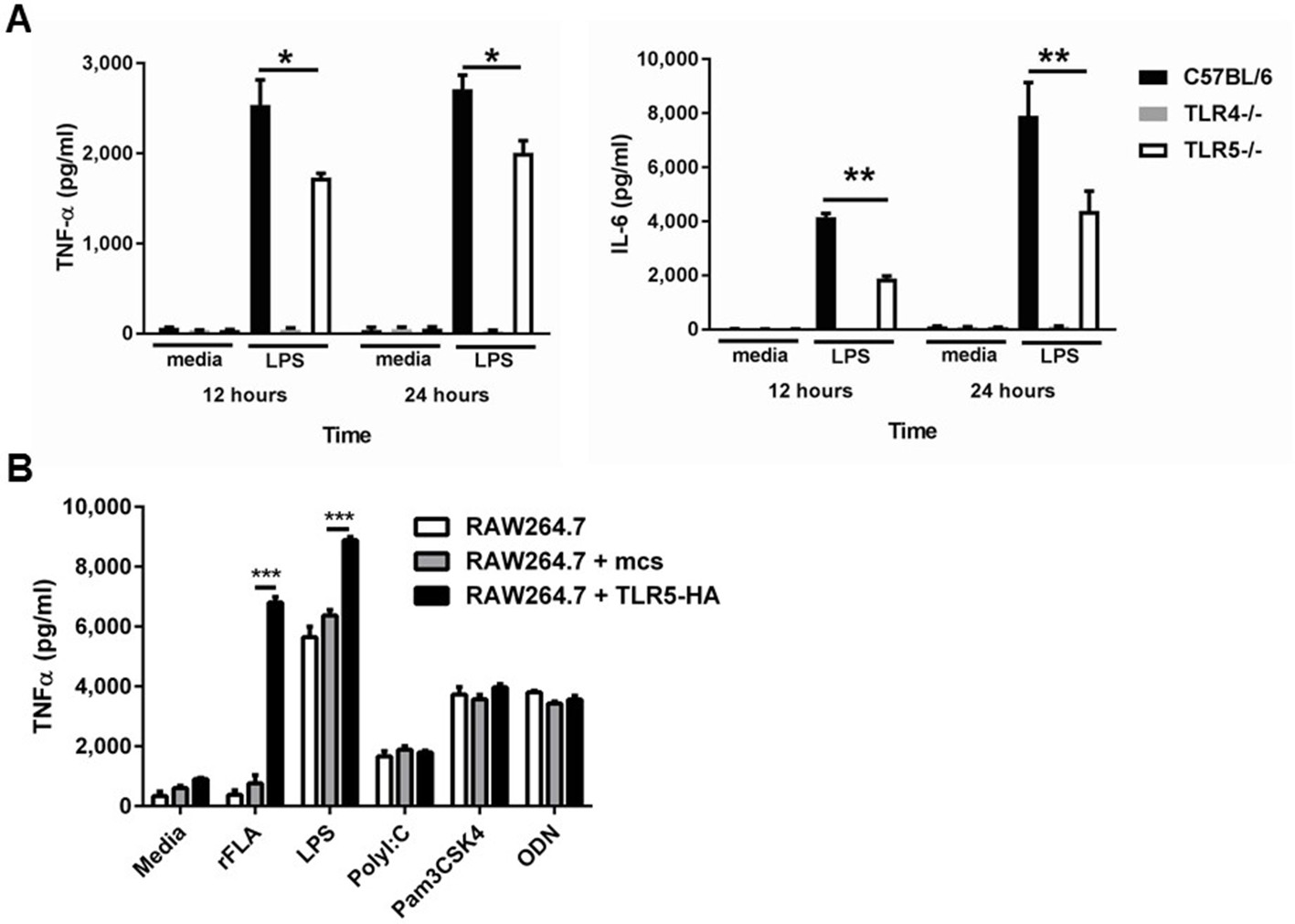
Deficiency of TLR5 reduces ultrapure LPS induced inflammatory cytokine production.
(A) TNFα and IL-6 production by BMDM from Tlr5-deficient (TLR5-/-), Tlr4-deficient (Tlr4-/-) and wild-type (C57BL/6) mice after 10 ng/mL ultrapure LPS or vehicle (media) exposure for 12 or 24 hr. N = 8 per group, experiment repeated at least 5 times. (B) TNF-α secretion by non-transfected (RAW264.7), empty construct transfected (RAW264.7 + mcs) or hemagglutinin-tagged TLR5 construct (TLR5-HA) transfected cells (RAW264.7 + TLR5-HA) after vehicle (media) or TLR5, TLR4, TLR3, TLR2 and TLR9 agonists (100 ng/mL recombinant ultrapure flagellin (rFLA), 10 ng/ml LPS, 10 µg/ml PolyI:C, 1 µg/ml Pam3CSK4, 1 µM ODN). N = 8 per group, experiment repeated twice. Data are represented as mean ± s.e.m. and were analyzed by two-way analysis of variance (ANOVA) followed by Tukey’s post hoc test. *p<0.05, **p<0.01, ***p<0.001.
Figure 5—figure supplement 1
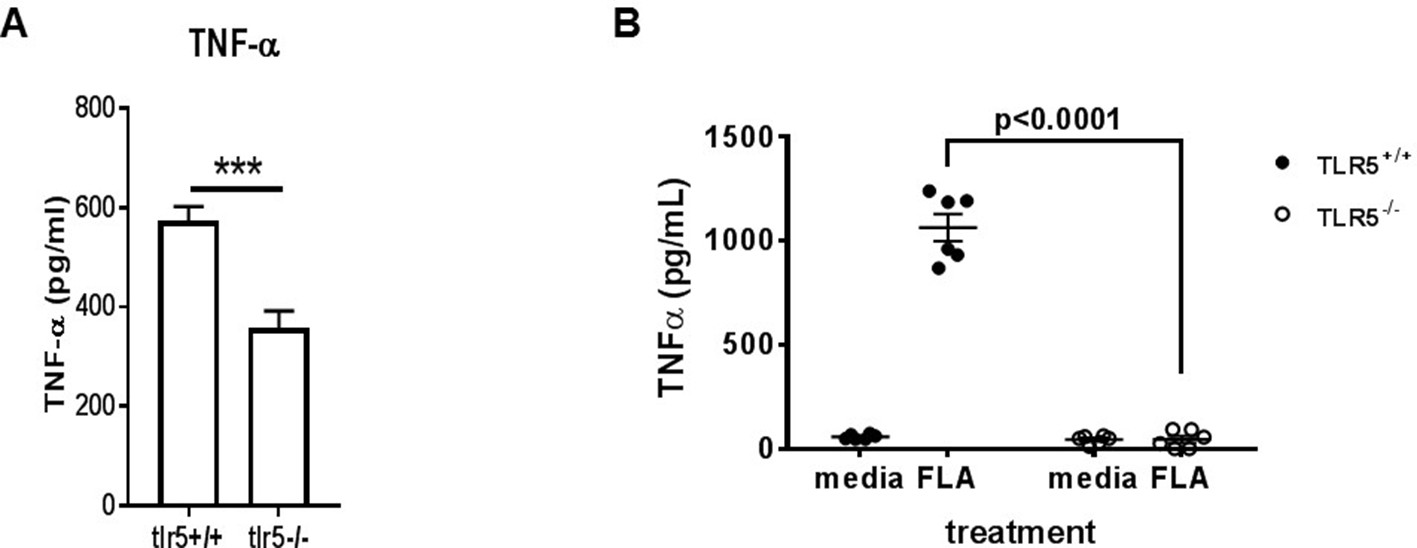
TNFa secretion from murine bone marrow-derived macrophages in response to monophoryl lipid A (A) and to flagellin (B).
Figure 6 with 1 supplement
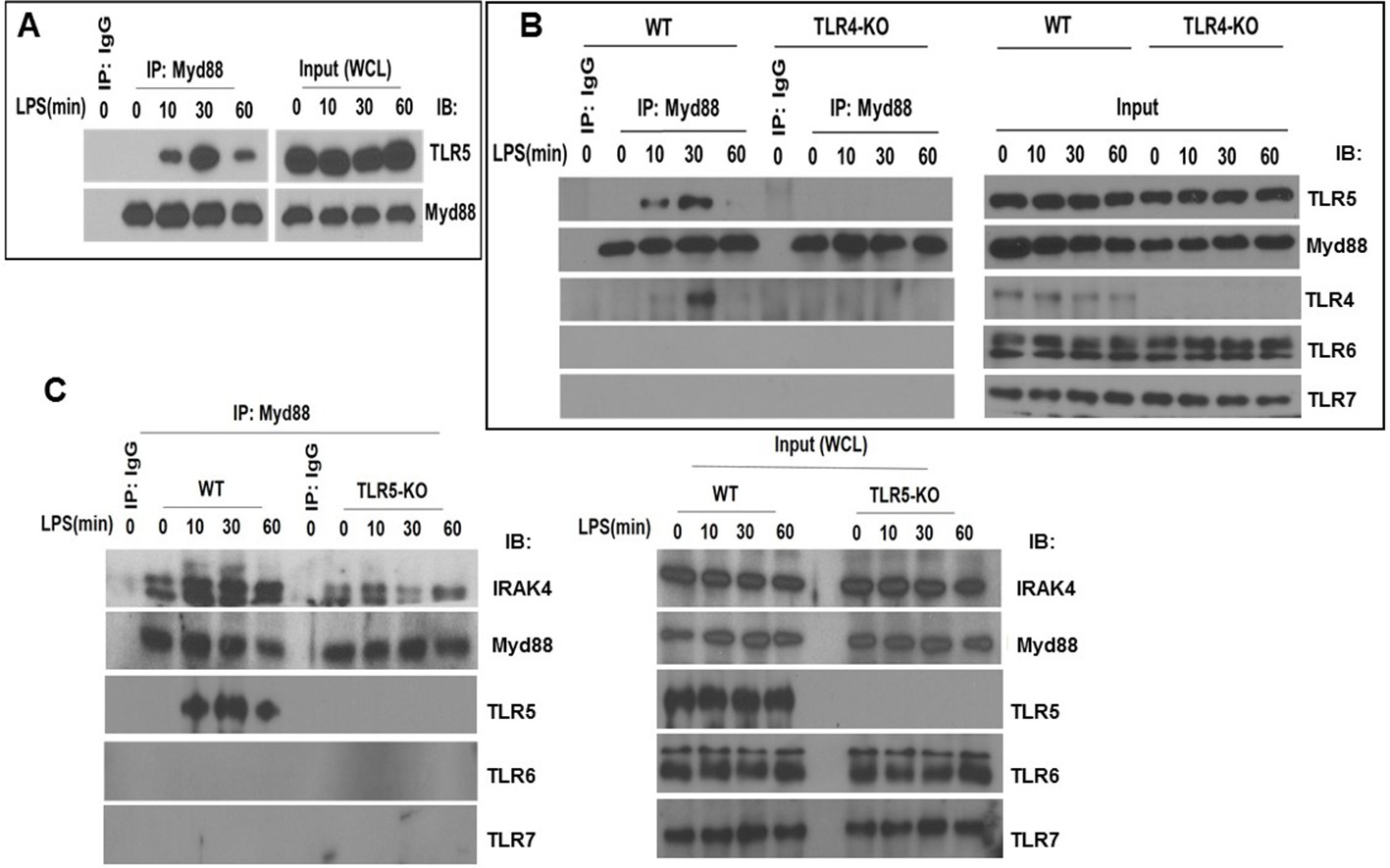
TLR5 engages with MyD88 and promotes Myddosome assembly after TLR4 activation.
(A) Co-immunoprecipitation of TLR5 with Myd88 in BMDM from C57BL/6J mice after 100 ng/mL LPS exposure for indicated time points. n = 7, experiment was repeated twice. (B) Immunoprecipitation of TLR5 with Myd88 in BMDM from Tlr4-deficient (TLR4-KO) or Tlr4-competent (C57BL/6, wildtype WT) mice after 100 ng/mL LPS exposure for indicated time points. Representative of 3 separate experiments. (C) Immunoprecipitation of IRAK4 with Myd88 in BMDM from Tlr5-deficient (TLR5 KO) or Tlr5-competent (C57BL/6, wildtype WT) mice after 100 ng/mL LPS exposure for indicated time points.
Figure 6—figure supplement 1
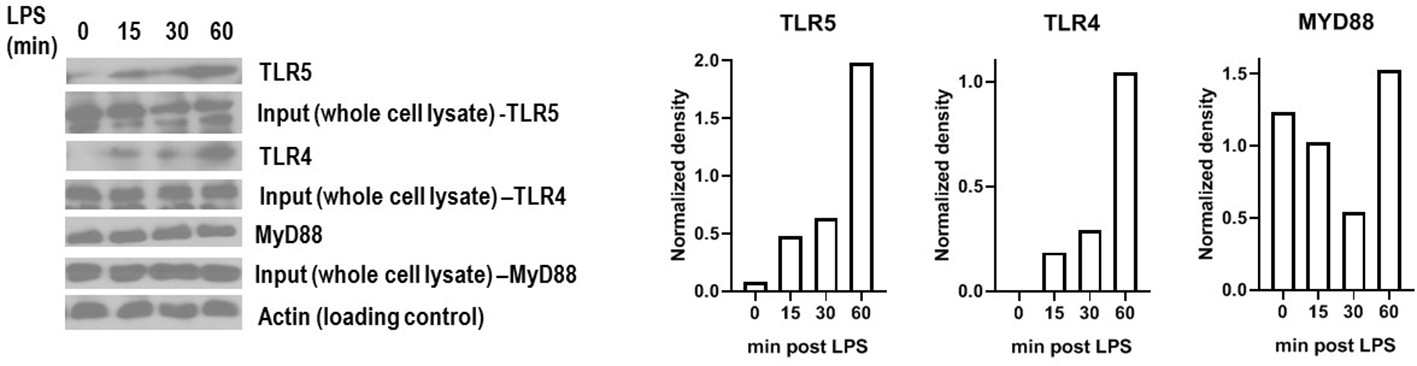
TLR4 and TLR5 co-immunoprecipitate with MYD88 after LPS exposure in primary human monocyte-derived macrophages.
Figure 7 with 1 supplement
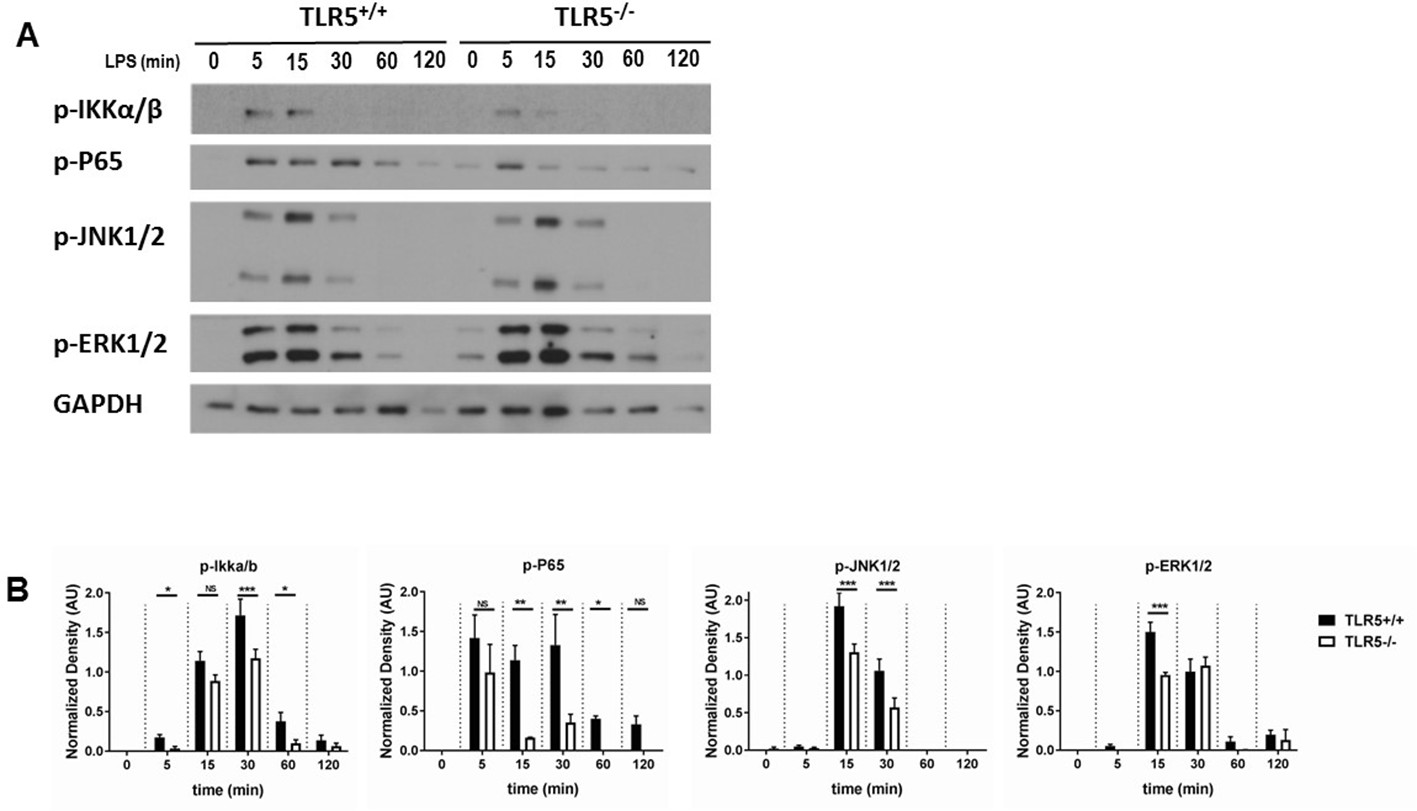
TLR5 promotes MyD88 downstream signaling.
(A) Western blot analysis of p-P65, p-IKKα/β, p-JNK1/2 and p-ERK1/2 after exposure to 100 ng/mL ultrapure LPS exposure in BMDMs from Tlr5-competent (TLR5+/+) and Tlr5-deficient (TLR5-/-) mice. (B) Quantification of densitometric analysis of 3 separate blots similar to (7A). Data are represented as mean ± s.e.m. and were analyzed by repeated unpaired t test with Holm-Sidak correction. NS = not significant, *p<0.05, **p<0.01, ***p<0.001.
Figure 7—figure supplement 1

Analysis of downsteam signaling after TLR4 activation, dependent on TLR5 status.
Figure 8 with 1 supplement
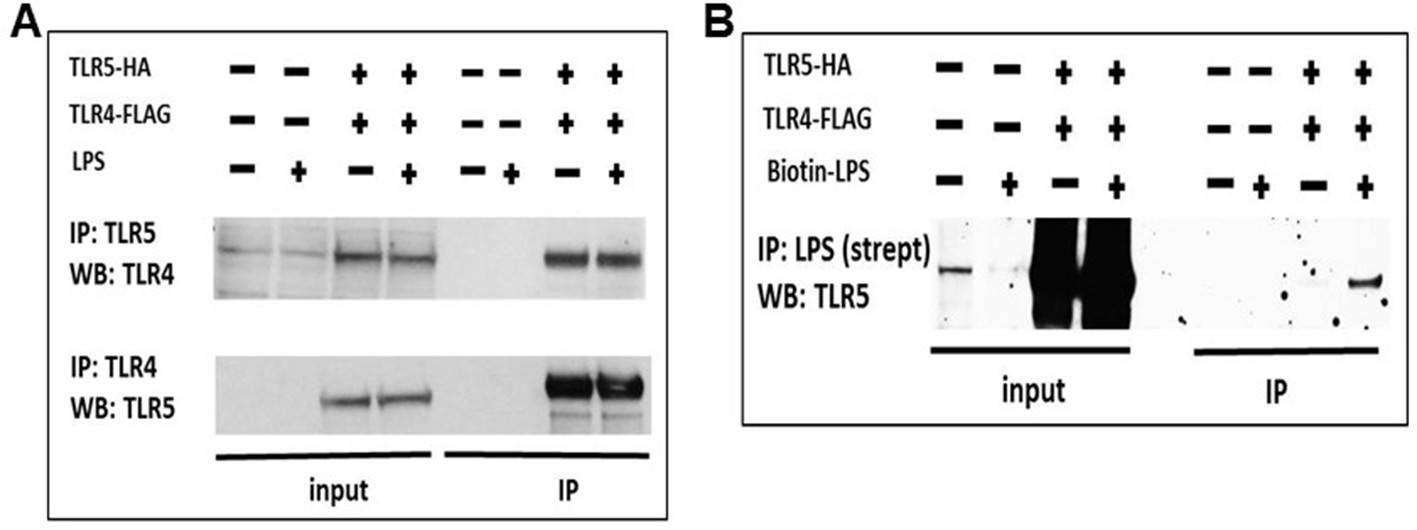
TLR5 participates in TLR4 signaling complex.
(A) Co-immunoprecipitation of hemagglutinin-tagged TLR5 (TLR5-HA) and FLAG-tagged TLR4 (TLR4-FLAG) in HEK293 cells after 100 ng/mL ultrapure LPS exposure. (B) Immunoprecipitation of TLR5 with biotinylated ultrapure LPS (Biotin-LPS) in TLR5-HA and TLR4-FLAG transfected HEK293 cells after 100 ng/mL Biotin-LPS exposure for 15 min. Representative of 2 separate experiments.
Figure 8—figure supplement 1
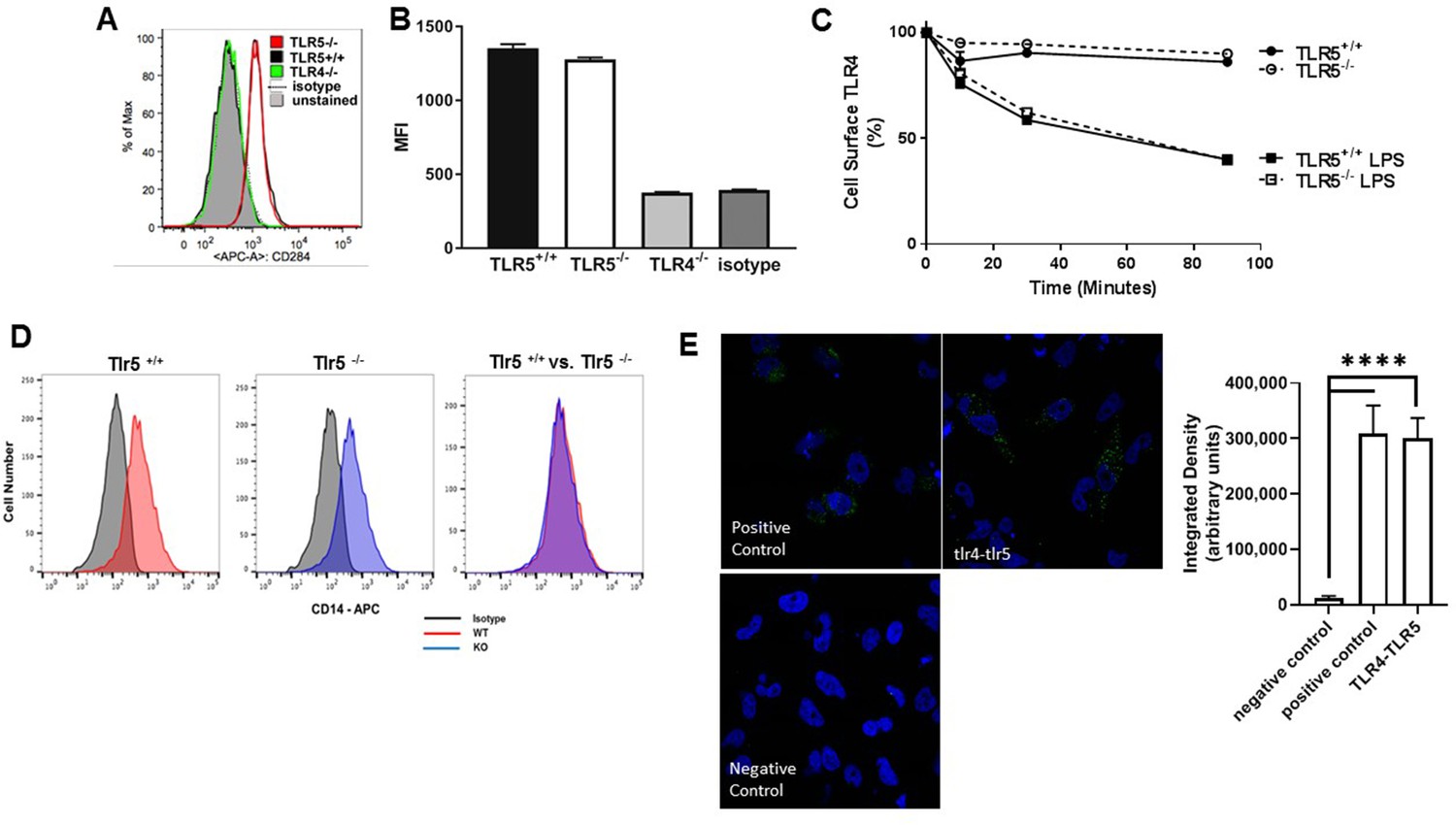
Cell surface expression of TLR4 and its co-localization with TLR5.
Figure 9
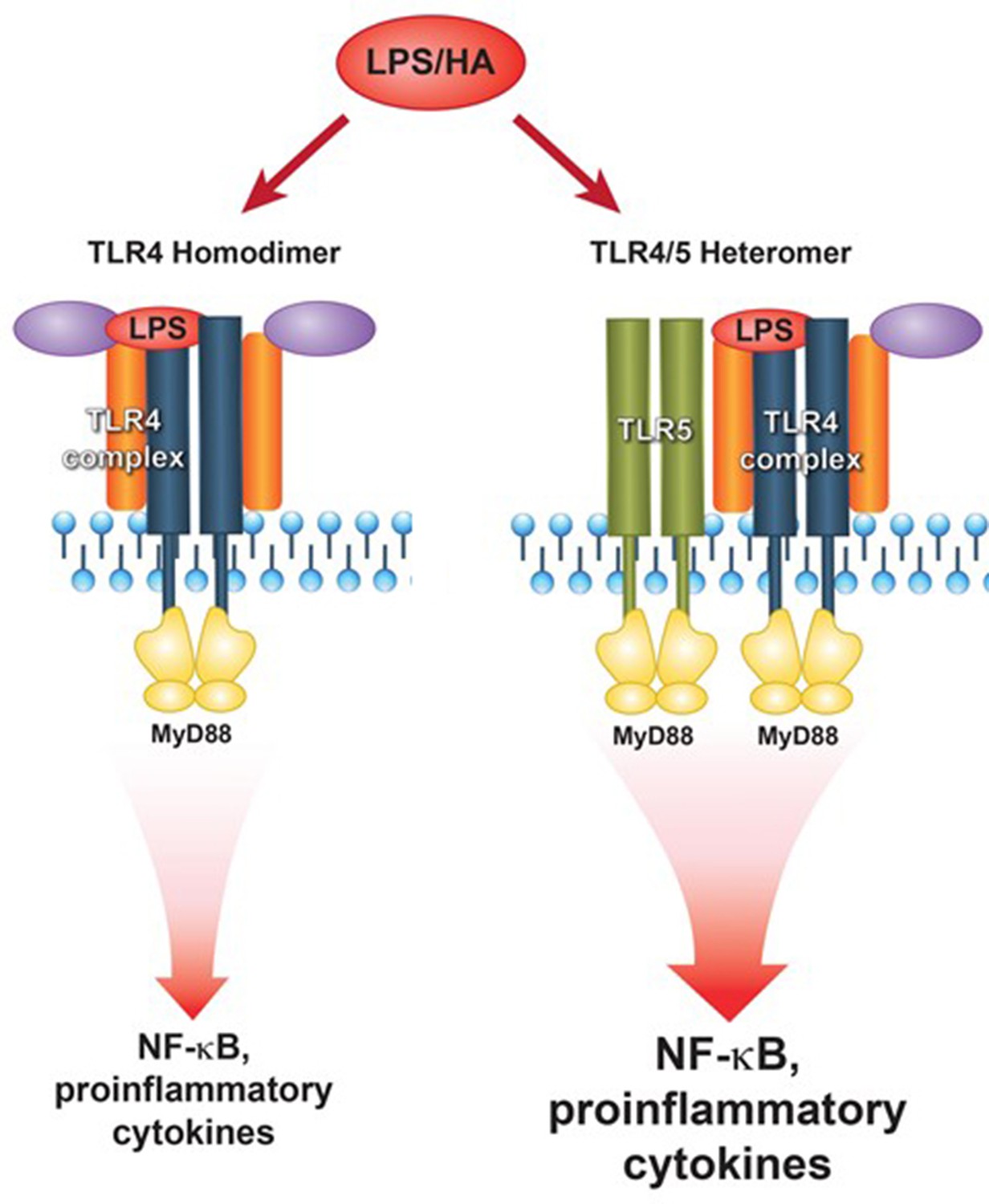
Proposed model of TLR5-TLR4 interaction during environmental lung injury.
The current model of canonical TLR4 activation rather applies to the TLR5 deficient status (left panel). In the presence of TLR5 (right panel), TLR5 participates in the TLR4 signaling complex, and promotes signaling downstream of the MyD88 pathway.
Author response image 1
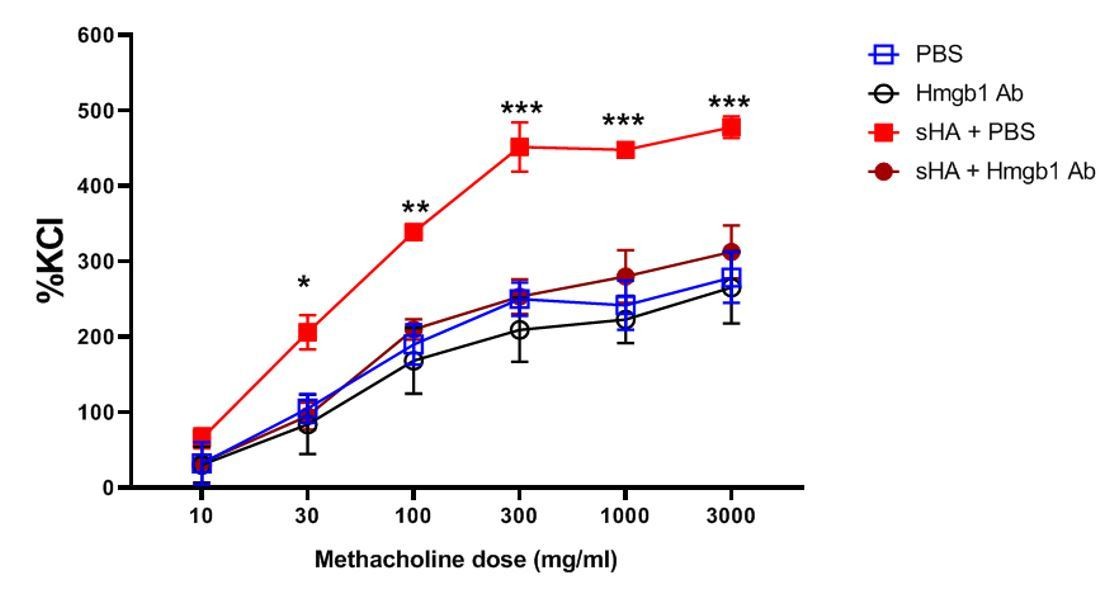
Neutralization of HMGB1 inhibits hyaluronan-induced airway hyperresponsiveness.
Murine tracheal rings were exposed to short-fragment hyaluronan which is known to elicit hyperresponsiveness of smooth muscle to methacholine. The rings were incubated with increasing doses of methacholine (an airway constrictor) and the muscle force generation was assayed, normalized to the standard response to potassium chloride (KCl), a nonspecific constrictor of the muscle. Treatment with the HMGB1-neutralizing antibody reduced the responsiveness to methacholine after hyaluronan exposure to baseline (PBS exposure) levels.
Additional files
-
Supplementary file 1
List of genes.
- https://cdn.elifesciences.org/articles/50458/elife-50458-supp1-v2.xls
-
Transparent reporting form
- https://cdn.elifesciences.org/articles/50458/elife-50458-transrepform-v2.docx
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
TLR5 participates in the TLR4 receptor complex and promotes MyD88-dependent signaling in environmental lung injury
eLife 9:e50458.
https://doi.org/10.7554/eLife.50458




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}