Yap-lin28a axis targets let7-Wnt pathway to restore progenitors for initiating regeneration
- Guangdong Provincial Key Laboratory of Malignant Tumor Epigenetics and Gene Regulation, Sun Yat-Sen Memorial Hospital, Sun Yat-Sen University, China
- Department of Otolaryngology, Sun Yat-Sen Memorial Hospital, Sun Yat-Sen University, China
- Key Laboratory of Stem Cells and Tissue Engineering, Zhongshan School of Medicine, Sun Yat-Sen University, Ministry of Education, China
Figures
Figure 1 with 2 supplements
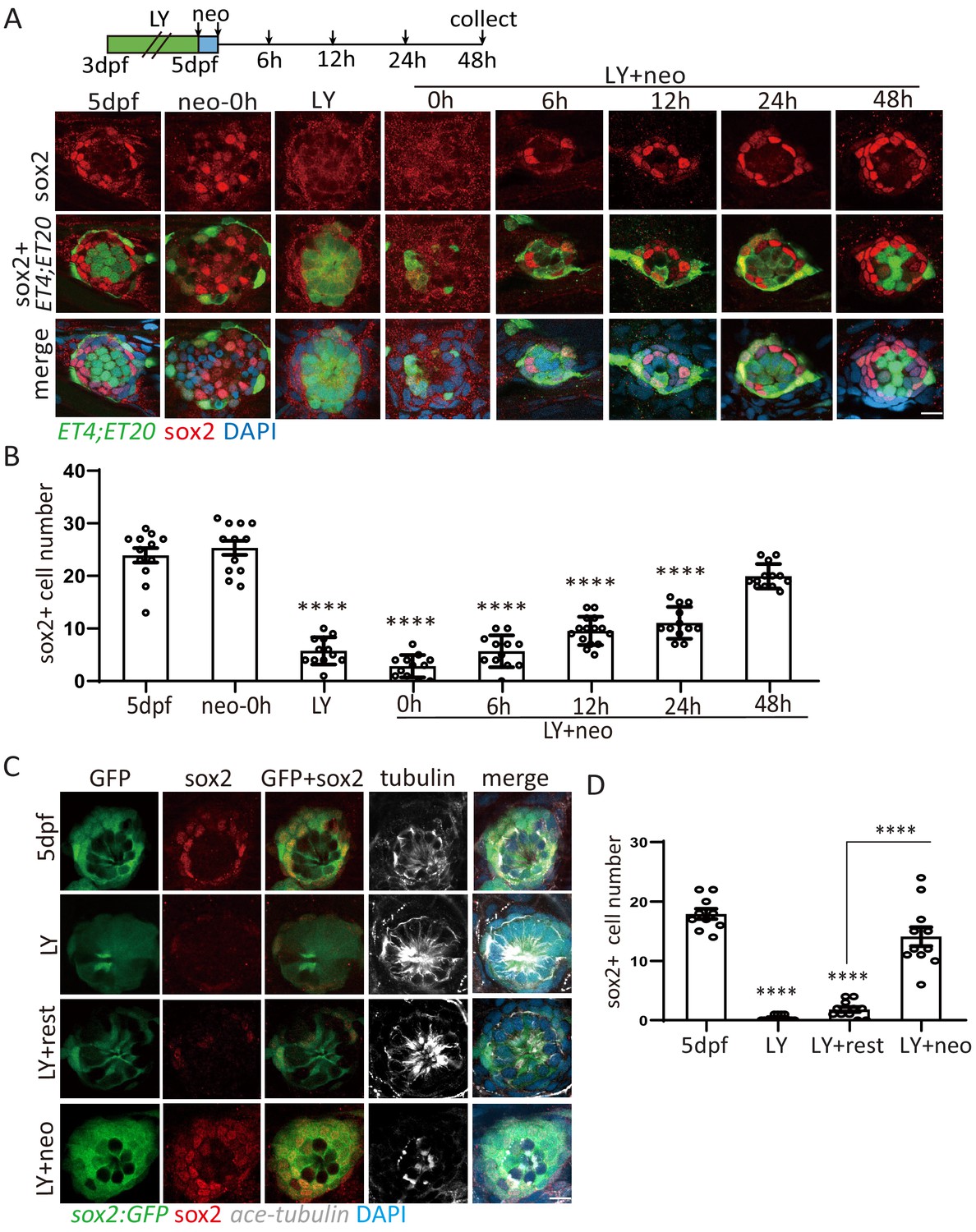
Exhausted sox2+ progenitors were able to restore quickly post severe injury.
(A, B) ET4;ET20 larvae were treated with neomycin, LY411575 (3dpf-5dpf), or neomycin following LY (LY+neo), and collected at indicated time points post neomycin treatment for sox2 immunostaining. The number of sox2+ progenitors was not affected post neo, while it was significantly decreased in LY and LY+neo-0h. The sox2+ progenitors were regenerated post LY+neo and recovered to normal level at 48 hr post LY+neo. (C, D) The sox2:GFP reporter was treated with LY from 3dpf to 5dpf to exhaust GFP+ progenitors. GFP+ progenitors cannot be regenerated when resting in normal medium for 2 days post LY treatment (LY+rest). In contrast, sox2+ progenitors were quickly recovered to normal level at 2-day post LY+neo. Scale bar equals 10 μm. All groups are compared with 5dpf unless indicated.
Figure 1—figure supplement 1

Severe injury causes damage to HCs, SCs and MCs.
ET4;ET20 larvae were treated with 2 μM LY from 3dpf to 5dpf followed by neomycin, and the number of HCs, SCs and MCs were counted before and right after neo treatment. Results showed that HCs were increased while SCs were decreased post LY, indicating that LY induced differentiation of SCs into HCs. Numbers of HCs, SCs and MCs were all significantly decreased post LY+neo compared with 5dpf normal larvae. Scale bar equals 10 μm.
Figure 1—figure supplement 2
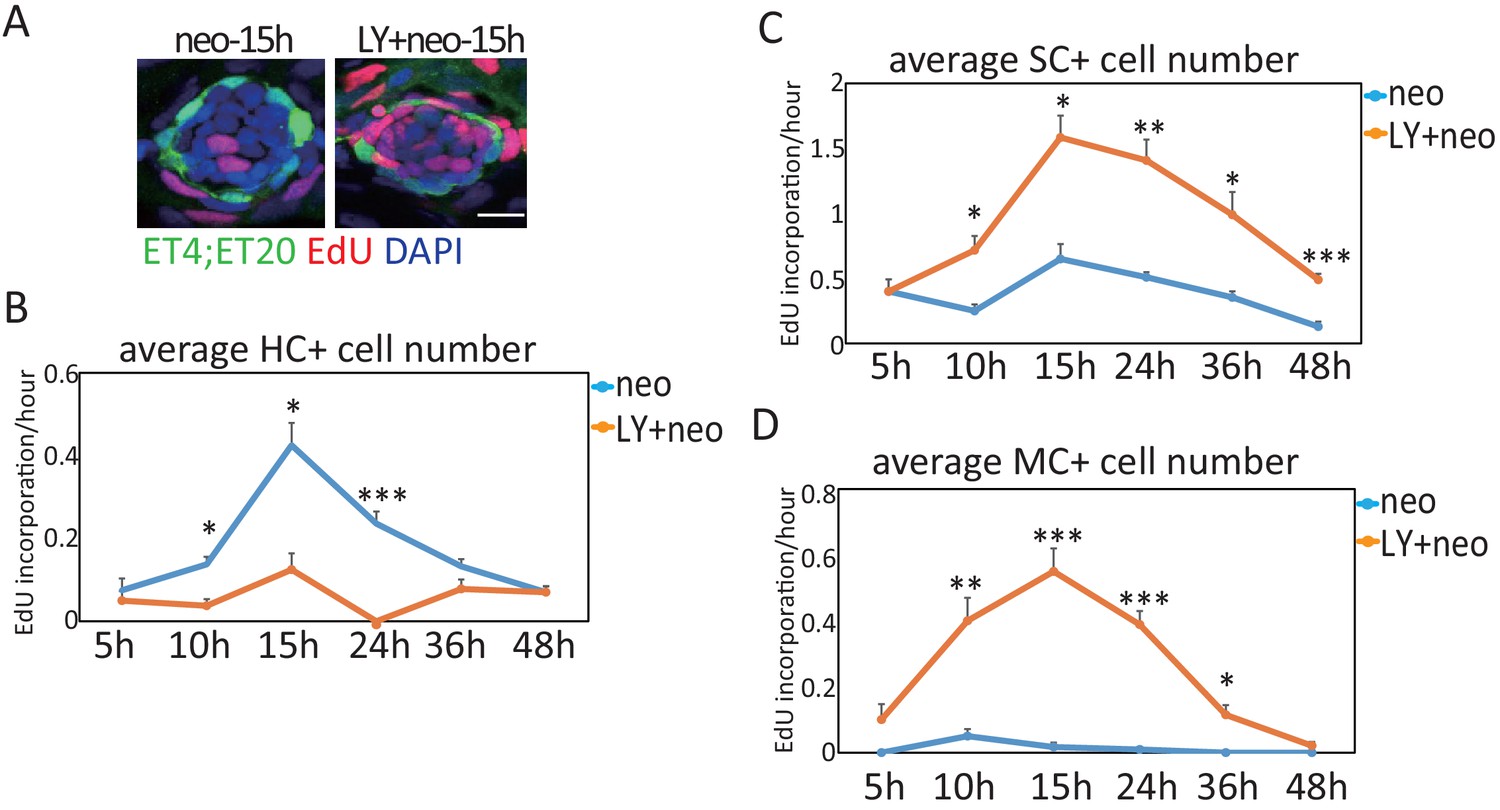
More proliferative SCs and MCs were induced post severe injury compared with normal injury.
Larvae treated with neo or LY+neo were incorporated with EdU for different time points and counted for numbers of differentiating cells (EdU+ET4+ cells, HC+), proliferative SCs (EdU+ET4-ET20- cells, SC+), and proliferative MCs (EdU+ET20+ cells, MC+). We calculated EdU incorporation per hour by dividing EdU+ cell number with incorporation time. Results showed that differentiation rate (HC+ in B) in LY+neo was decreased compared with neo, while proliferative rates of SCs (SC+ in C) and MCs (MC+ in D) were significantly increased. Scale bar equals 10 μm.
Figure 2 with 2 supplements
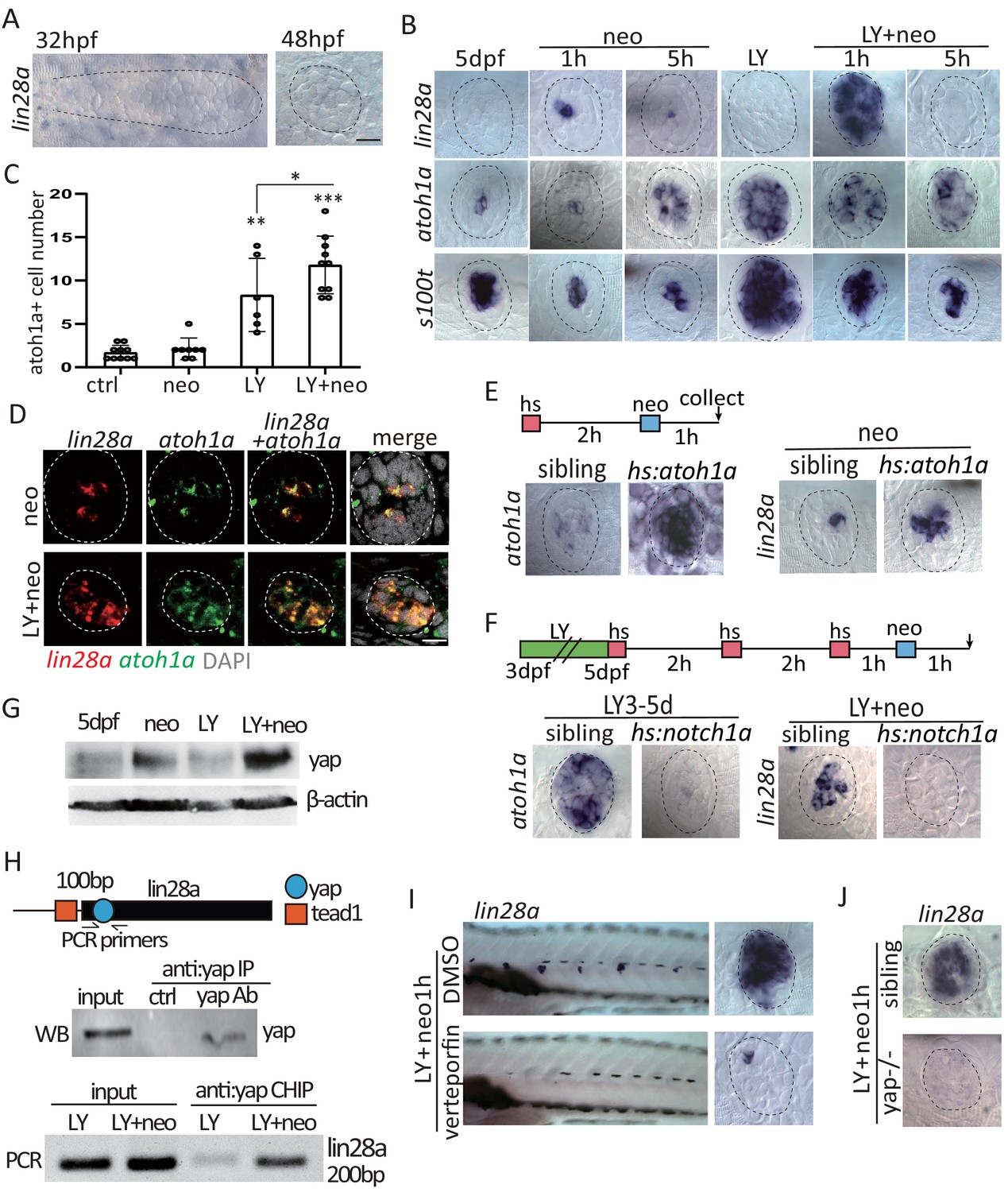
Activated yap upregulate lin28a transcription in atoh1a+ HC precursors post severe injury.
(A) Lin28a was not expressed in the developing lateral line primordium or neuromast. (B) Treatment with Notch inhibitor LY411575 from 3dpf to 5dpf increased expression atoh1a and s100t. More lin28a expression was observed in LY+neo-1h compared with neo-1h. No lin28a was detected at 5 hr post LY+neo or neo. (C) The number of atoh1a-transcribed cells detected by in situ were higher at 1 hr post LY+neo compared with LY. (D) Double fluorescent in situ showed that lin28a was co-expressed with atoh1a post neo or LY+neo. (E) Induction of lin28a post injury was increased when atoh1a was overexpressed in hs:atoh1a. (F) Lin28a expression was completely blocked when atoh1a was inhibited in hs:notch1a. (G) Western blot results showed that LY+neo induced higher yap expression compared with neo alone. (H) Motifs of yap and tead1 (co-transcriptional factor of yap) binding sites were predicted near lin28a transcriptional start site. CHIP-PCR results verified that yap directly binds the predicted motif. (I and J) Inhibition of yap using verteporfin or yap mutant blocked lin28a induction post LY+neo. Scale bar equals 10 μm.
Figure 2—figure supplement 1
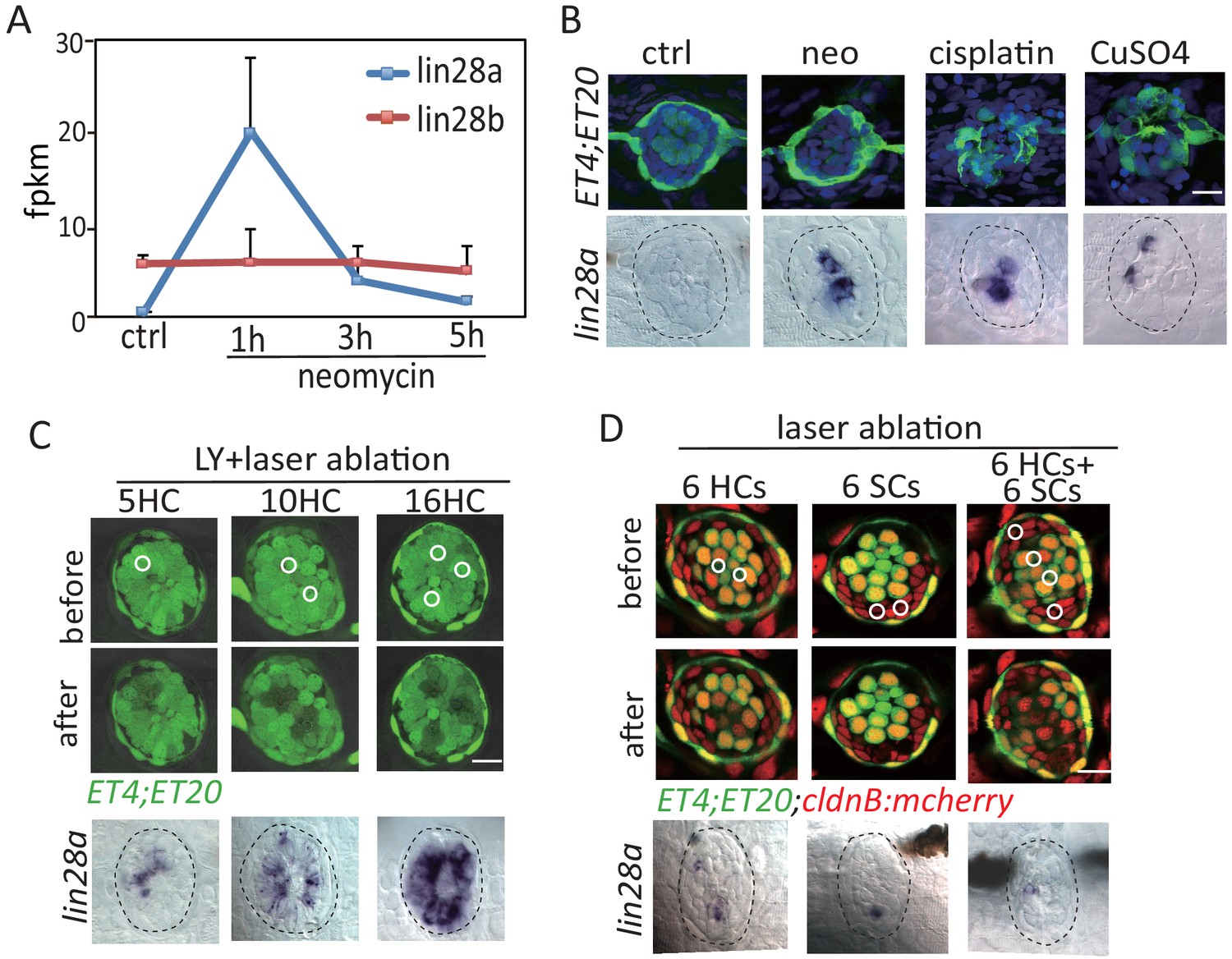
lin28a was induced by various kinds of injury and lin28a expression level was proportional to injury size.
(A) RNA-Seq analysis using neomycin-treated neuromast cells revealed that lin28a was highly induced at 1 hr post neo while lin28b expression wa s not changed. (B) lin28a was induced at 1 hr post treatments with neomycin, cisplatin and copper. (C) LY-treated larvae were subjected for laser scan to ablate HCs at the indicated locations (white circles). Expression levels of lin28a was proportional to the injured cell number manipulated by laser ablation. (D) Laser was targeted at the circles for ablating indicated number of HCs or SCs. The corresponding neuromast was performed for lin28a in situ, and the results showed that either HC damage or SC damage could induce lin28a expression. Scale bar equals 10 μm.
Figure 2—figure supplement 2
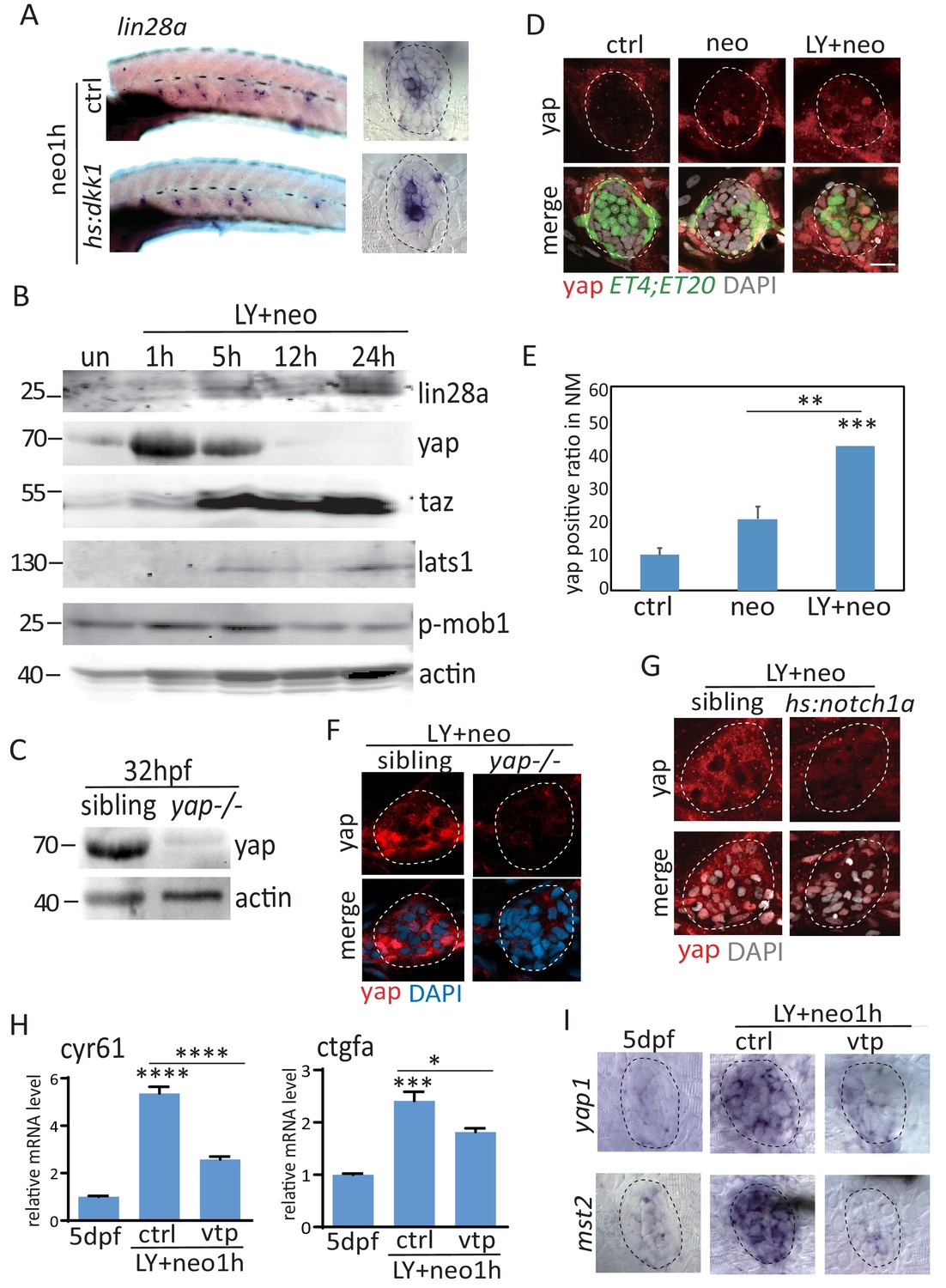
Yap was activated immediately post LY+neo.
(A) Wnt inhibition with hs:dkk1 has no effect on lin28a induction post neo treatment. (B) Expression levels of lin28a, yap, taz and Hippo pathway genes, such as lats1 and mob1, were detected by western blot at various time points post LY+neo. Results showed that yap was increased at 1 hr post LY+neo. Levels of lats1 and phosphorylated mob1 were not changed at 1 hr, suggesting yap is activated independent of canonical Hippo pathway. (C) Specificity of yap antibody was tested with 32hpf sibling and yap mutants. (D–E) Larvae treated with neo or LY+neo were immonstained with yap antibody. Results showed that nuclear localized yap was increased in LY+neo compared with neo. (F) LY+neo induced yap expression in sibling but not yap mutants. (G) Yap activation post LY+neo was inhibited when atoh1a is inhibited with hs:notch1a, suggesting that yap is activated in atoh1a+ HC precursors. (H) Classic yap target genes cyr61 and ctgfa were upregulated post LY+neo, which is partially blocked by verteporfin. (I) Transcripts of yap and mst2 were highly induced post LY+neo, which was blocked by verteporfin. Scale bar equals 10 μm.
Figure 3 with 2 supplements
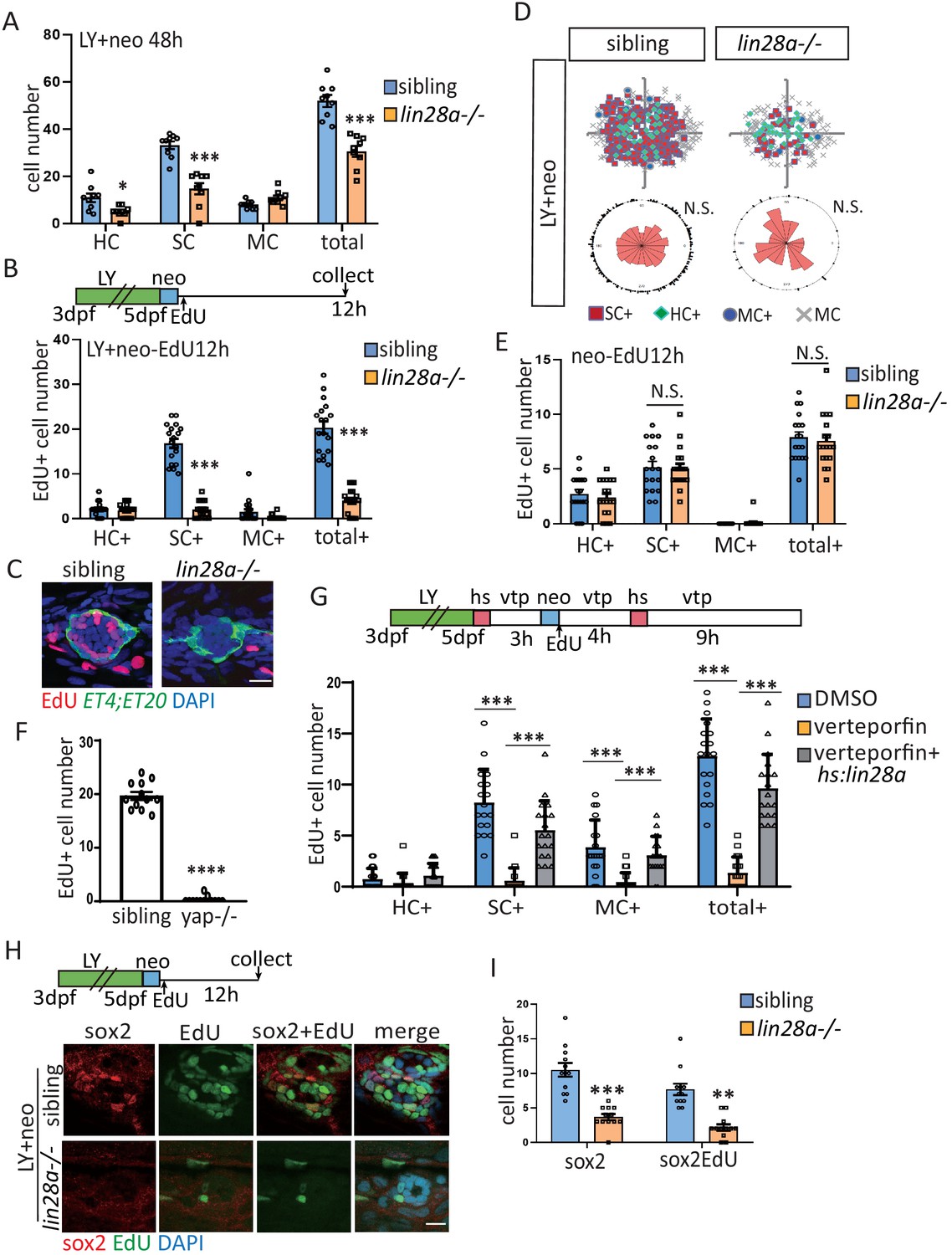
Yap-lin28a pathway is essentially required for progenitor recovery post severe injury.
(A) The number of SCs and total cells were counted at 48 hr post LY+neo and were significantly decreased in lin28a mutant compared with sibling. (B–E) The ET4;ET20 larvae were incorporated with EdU post neo or LY+neo treatment. Three populations labeled with ET4+EdU+ (HC+), ET20+EdU+ (MC+) or ET4-ET20-EdU+ (SC+) were counted and recorded with location information. The proliferative SCs post LY+neo were significantly decreased in lin28a mutant compared with sibling (B and C), while not changed in neo-induced regeneration (E). (D) EdU plots show the positions of EdU+ nuclei of 18 neuromasts superimposed on the same plane, and rose diagrams document the angular positions of SC+. The results show that the proliferative SCs are evenly distributed in each quadrant with no polarization post LY+neo. (F) LY+neo-induced Edu incorporation was significantly reduced in yap mutant. (G) The hs:lin28a larvae were heat-shocked and pre-treated with 10 μM verteporfin before adding neomycin. Samples were collected at 14 hr post LY+neo for EdU+ cell counting. Proliferation (SC+) is significantly decreased post LY+neo in verteporfin, which could be rescued by overexpression of lin28a with hs:lin28a. (H, I) The numbers of regenerated sox2+ progenitors (sox2) and proliferative progenitors (sox2EdU) post LY+neo were both reced in lin28a mutant. Scale bar equals 10 μm.
Figure 3—figure supplement 1
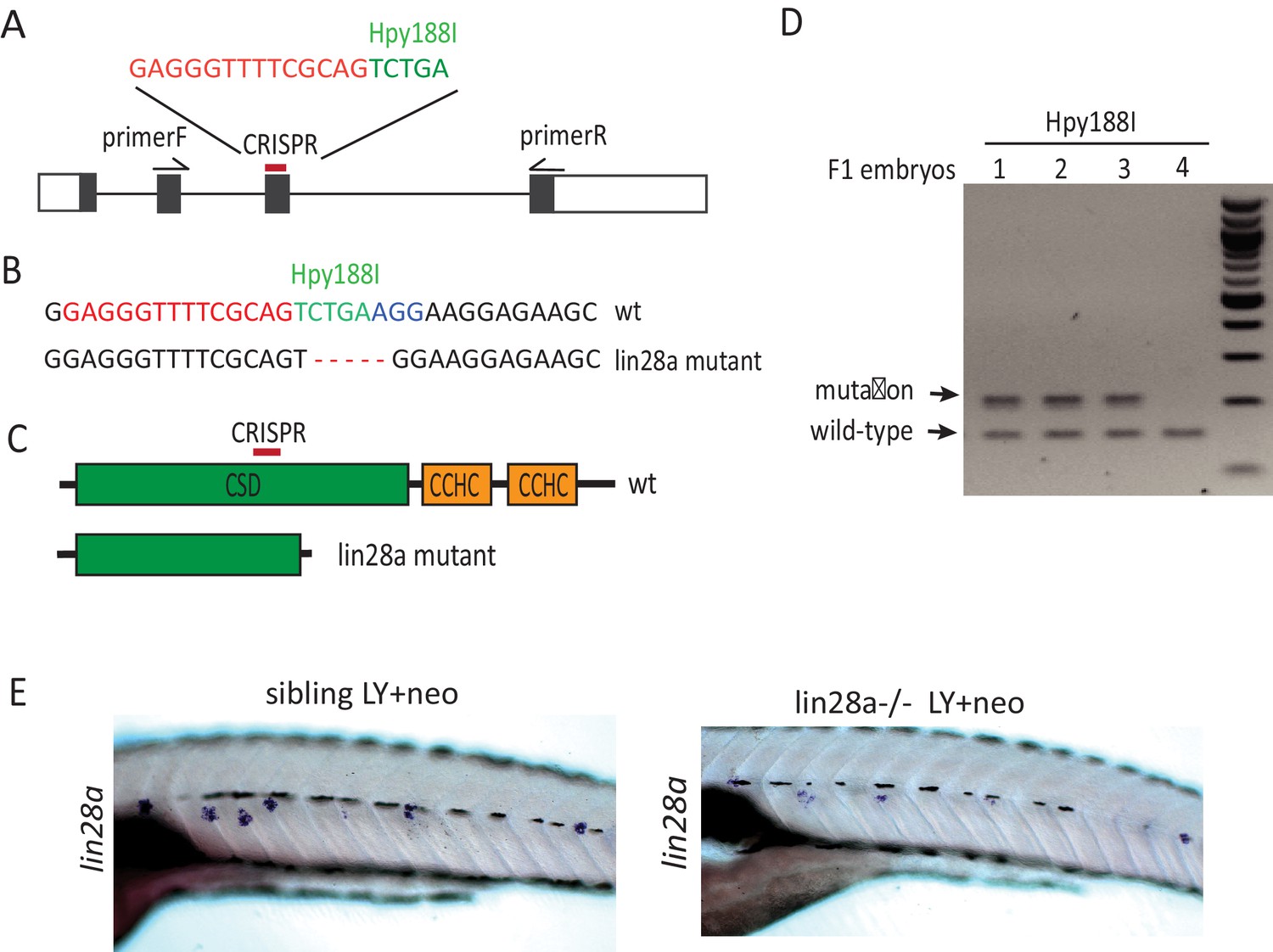
Create lin28a mutant using CRISPR.
(A) Diagram shows four exons of lin28a and the position of CIRPSR that was used for creating lin28a mutant. (B–C) Deletion of five nucleotides in the lin28a mutant allele damages the restriction enzyme site of Hpy188I and leads to a truncated protein by creating a premature stop codon in lin28a mRNA. (D) F1 embryos were genotyped for lin28a mutation by PCR followed by Hyp188I digestion. (E) Transcribed lin28a mRNA level was decreased in lin28a-/- post LY+neo, probably because the mutant form of lin28a mRNA is unstable and degraded.
Figure 3—figure supplement 2

HCs and sox2+ progenitors were not affected by lin28a deficiency in homeostasis, LY and LY+neo.
We compared numbers of ET4+ HCs and sox2+ progenitors in 5dpf sibling and lin28a-/- and found that they were not changed in lin28-/- in homeostasis (A), LY (B) and LY+neo (C). Scale bar equals 10 μm.
Figure 4 with 1 supplement
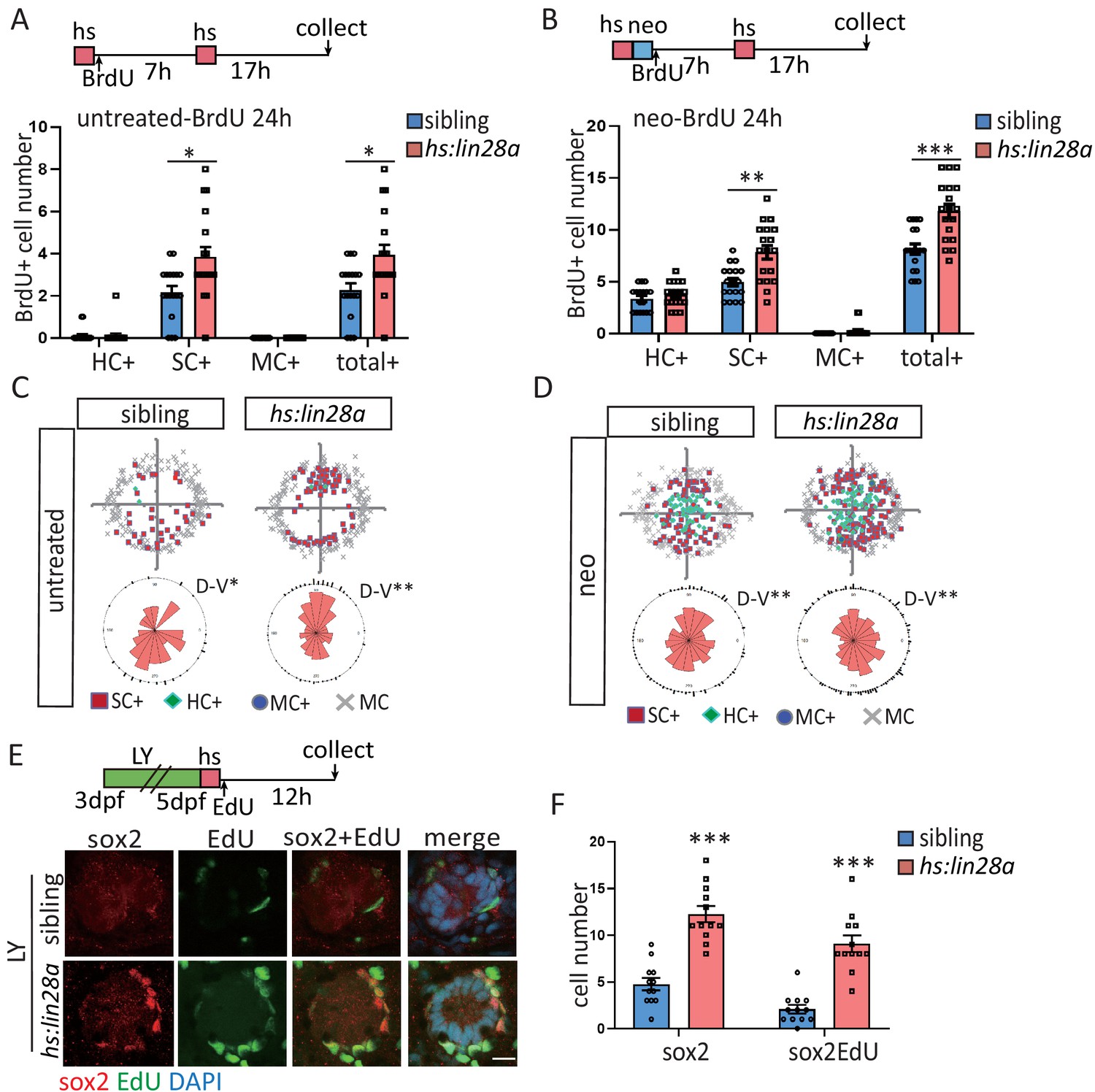
Overexpression of lin28a is sufficient to restore the exhausted progenitors.
(A–D) ET4;ET20;hs:lin28a larvae were incorporated with BrdU for 24 hr following heat-shock and/or neomycin treatment. (A, B) Overexpression of lin28a increased number of proliferative SCs (SC+) in both untreated and neomycin conditions. (C, D) BrdU plots and rose diagrams indicate that locations of SC+ still remain dorsally and ventrally polarized in hs:lin28a. (E, F) OverexpressUion of lin28a is sufficient to partially restore the exhausted sox2+ progenitors post LY. Scale bar equals 10 μm.
Figure 4—figure supplement 1
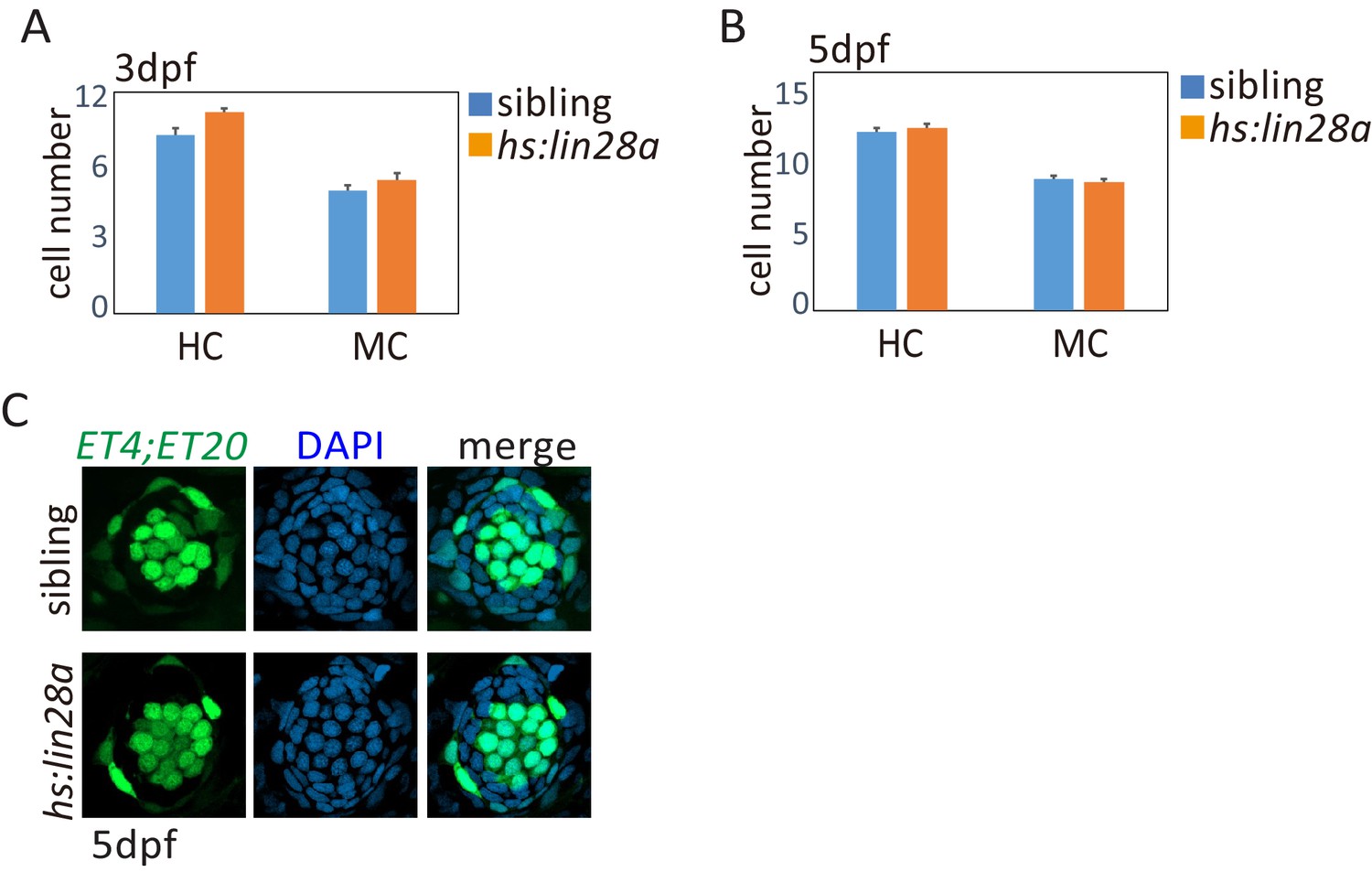
Developing HCs and MCs were not affected in hs:lin28a.
The ET4;ET20;hs:lin28a larvae were heat-shocked at 3dpf or 5dpf and collected after 24 hr for HC and MC counting. Results showed that numbers of HCs and MCs were not significantly changed by overexpression of lin28a at 3dpf (A) and 5dpf (B and C).
Figure 5 with 1 supplement
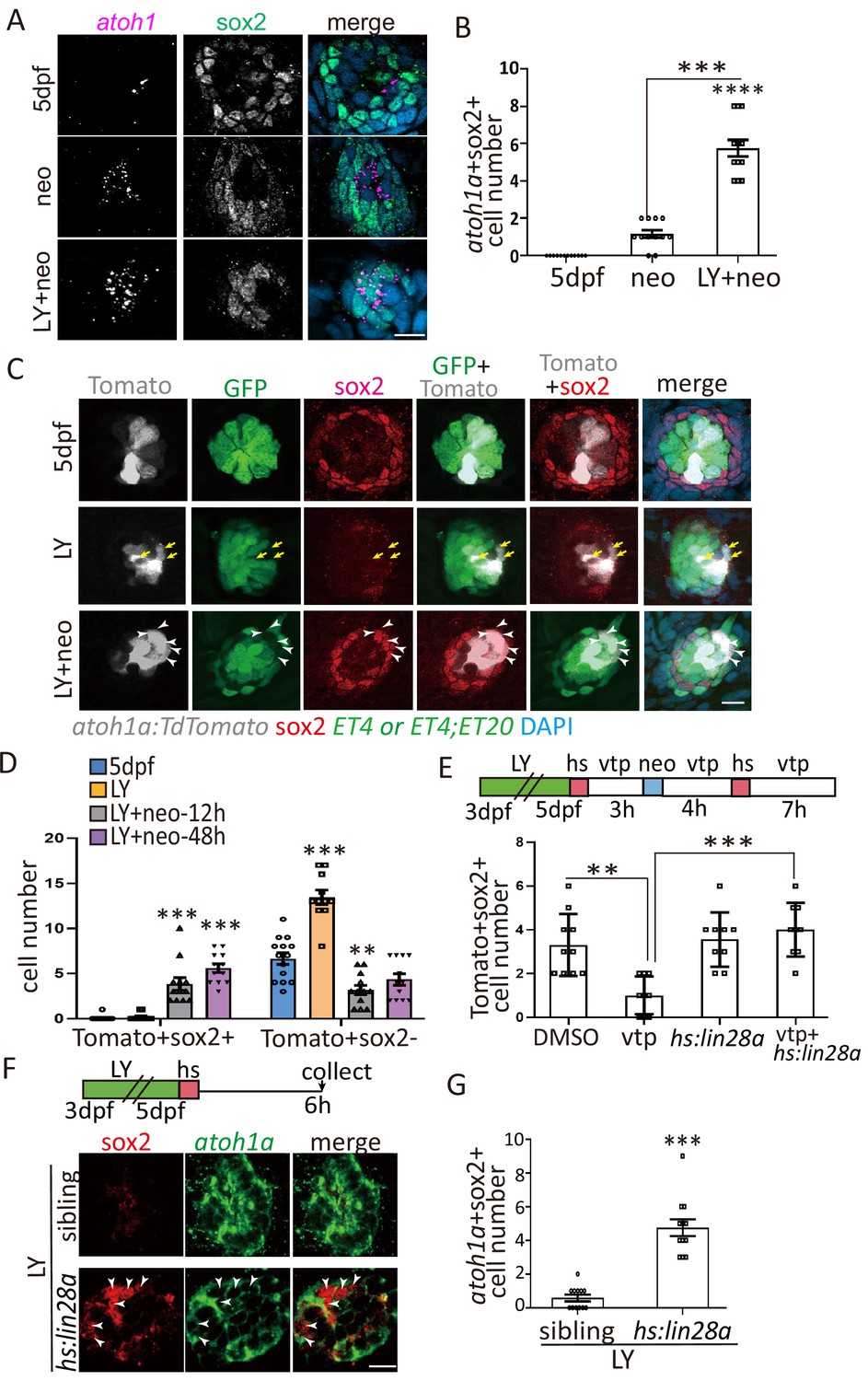
Yap-lin28a pathway promotes sox2 expression in atoh1a+ HC precursors.
(A–B) Larvae were stained with sox2 antibody and atoh1a RNA probe at 6 hr post neo or LY+neo treatment. The number of atoh1a+sox2+ cells was significantly increased in LY+neo group. (C–D) The atoh1a:TdTomato larvae were used to trace atoh1a+ HC precursors in ctrl, LY or LY+neo. Results showed that Tomato+ cells labeled partial ET4+ HCs and ET4-sox2- HC precursors (yellow arrows). However, many atoh1a+ cells start to express sox2 from 12 hr post LY+neo and their numbers were significantly increased compared with normal larvae. The arrowheads in (C) pointed the atoh1a+sox2+ cells at 48 hr post LY+neo. (E) The atoh1a:TdTomato;hs:lin28a larvae was treated with LY+neo and verteporfin and collected for immunostaining with sox2 antibody. The cell number of Tomato+sox2+ is decreased in verteporfin and overexpression of lin28a could rescue the phenotype. (F–G) The LY-treated hs:lin28a larvae were heat-shocked to overexpress lin28a. Samples were collected for staining with sox2 antibody and atoh1a RNA probe. The number of atoh1a+sox2+ cells was significantly increased in hs:lin28a group, indicating that lin28a is sufficient to express sox2 in atoh1a+ HC precursors. Scale bar equals 10 μm.
Figure 5—figure supplement 1
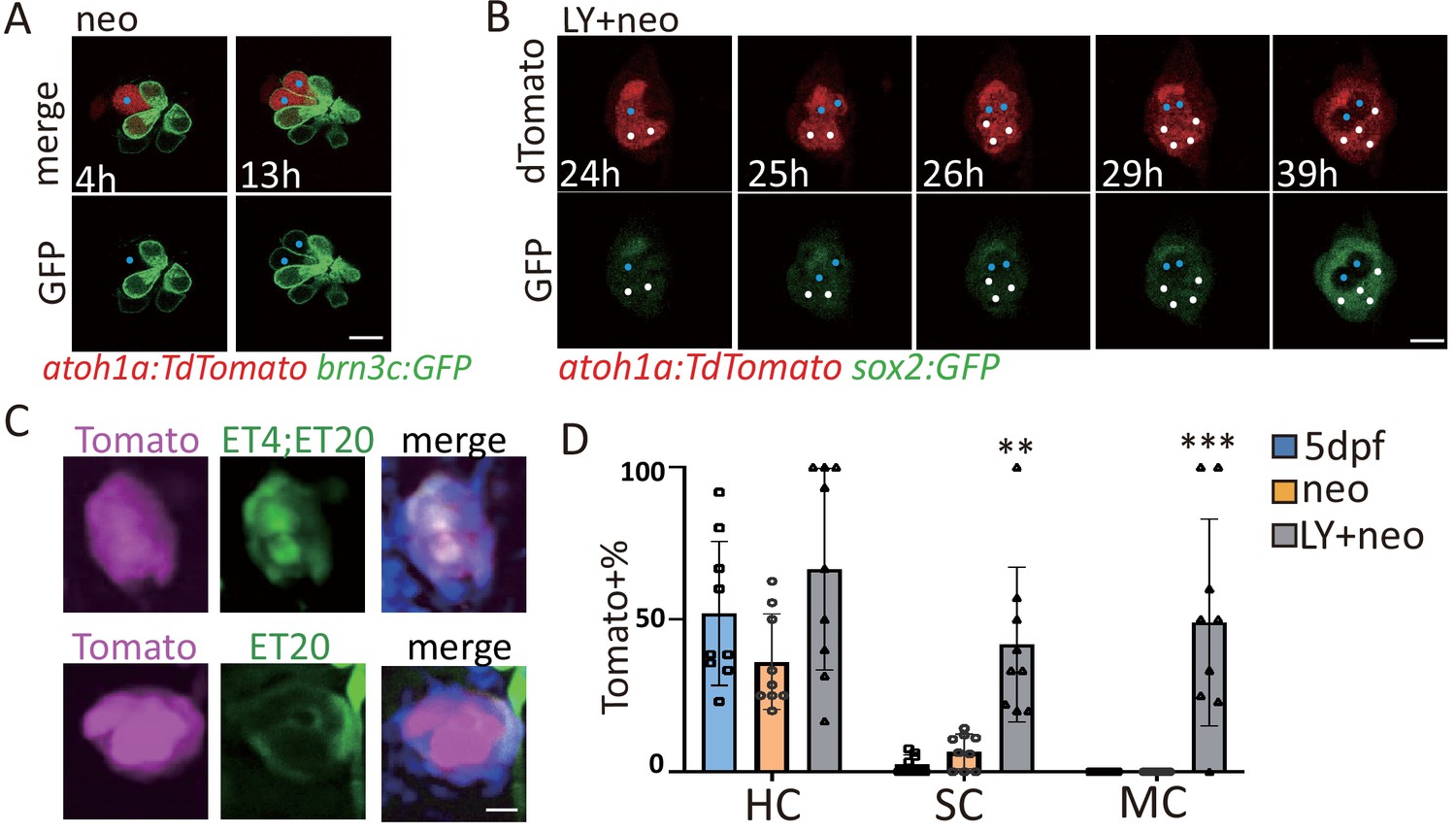
Atoh1a+ cells became SCs and MCs post severe injury.
(A) The Neo-treated atoh1a:TdTomato;brn3c:GFP larvae was imaged for time lapse. Results showed that one Tomato+ cell turned into two HCs with membrane-bound GFP post neo (blue dots). (B) The LY+neo-treated atoh1a:TdTomato;sox2:GFP larvae was imaged for time lapse. The blue dots indicate that one Tomato+ cell divided and turned into two sox2:GFP-negative HCs post LY+neo. The white dots indicate that two Tomato+ cells divided and became four sox2:GFP-positive cells post LY+neo. (C and D) The atoh1a:TdTomato;ET4;ET20 larvae were collected at 48 hr post neo or LY+neo. Tomato expressing HCs, SCs and MCs were separately divided by total numbers of each to calculate Tomato+ ratios. The results showed that significantly more Tomato+ SCs were detected post LY+neo compared with neo. In addition, many Tomato+ MCs were detected post LY+neo, but none was detected in 5dpf and neo.
Figure 6 with 1 supplement
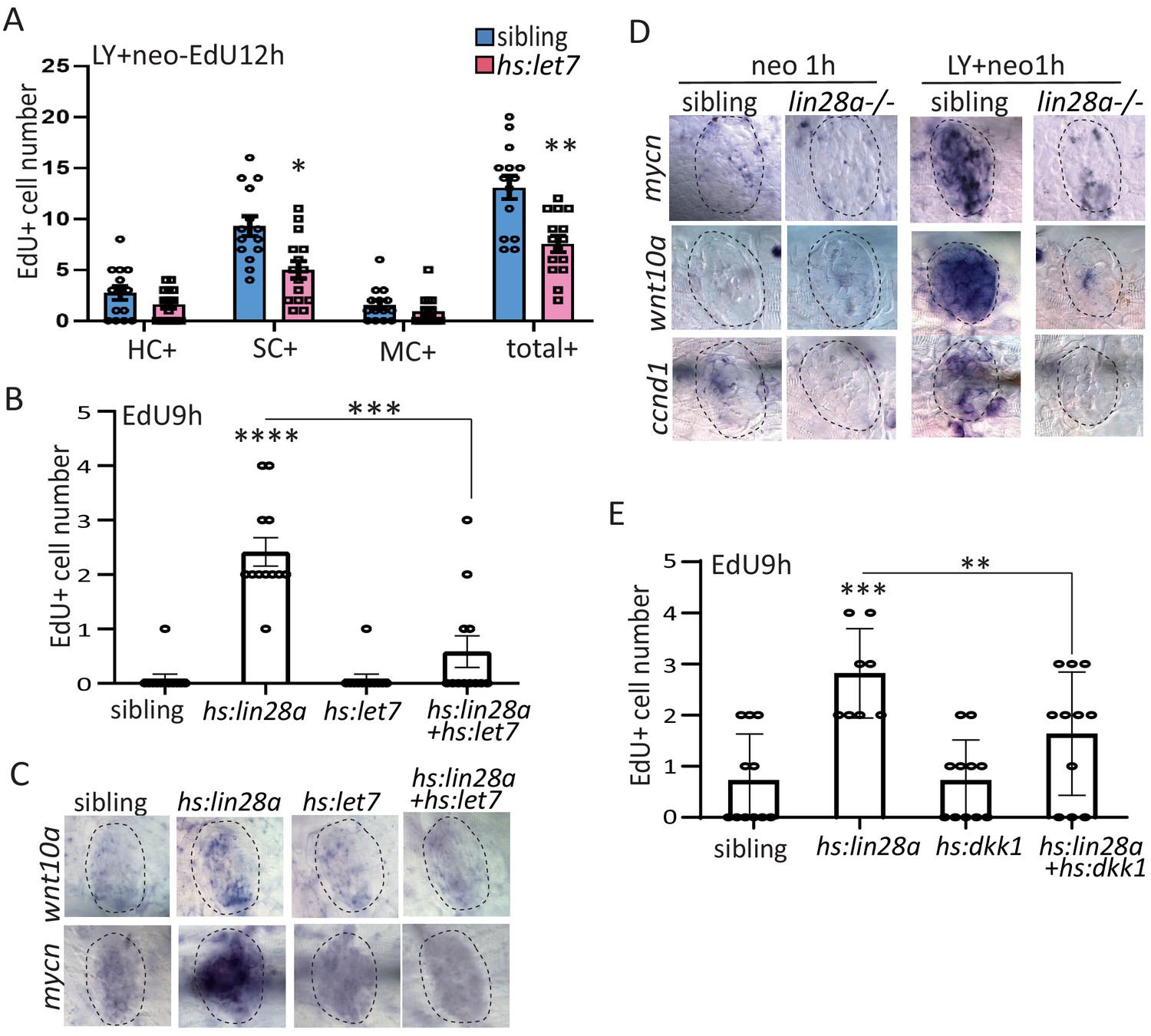
MicroRNA let7 acts downstream of lin28a to activate Wnt pathway for promoting progenitor regeneration.
(A) We created hs:let7 transgenic line and found that it recapitulates the phenotype of lin28a-/- by decreasing proliferative SCs post LY+neo. (B) The induction of EdU+ proliferative cells was blocked by hs:let7, indicating that let7 microRNA acts downstream of lin28a to induce proliferation. (C) In situ hybridization results showed that expression of Wnt pathway genes, such as wnt10a and mycn, were increased in hs:lin28a. The activation of Wnt pathway genes in hs:lin28a were blocked when let7 was overexpressed, indicating that let7 acts downstream of lin28a to inhibit Wnt pathway. (D) Expressions of Wnt pathway genes, such as mycn, wnt10a and ccnd1 (cyclind1), were highly induced at 1 hr post LY+neo in sibling, but were not detected in lin28a-/-. (E) Inhibition of Wnt pathway with hs:dkk1 decreased lin28a-induced EdU+ proliferative cells, indicating that Wnt activation acts downstream of lin28a to induce regeneration.
Figure 6—figure supplement 1
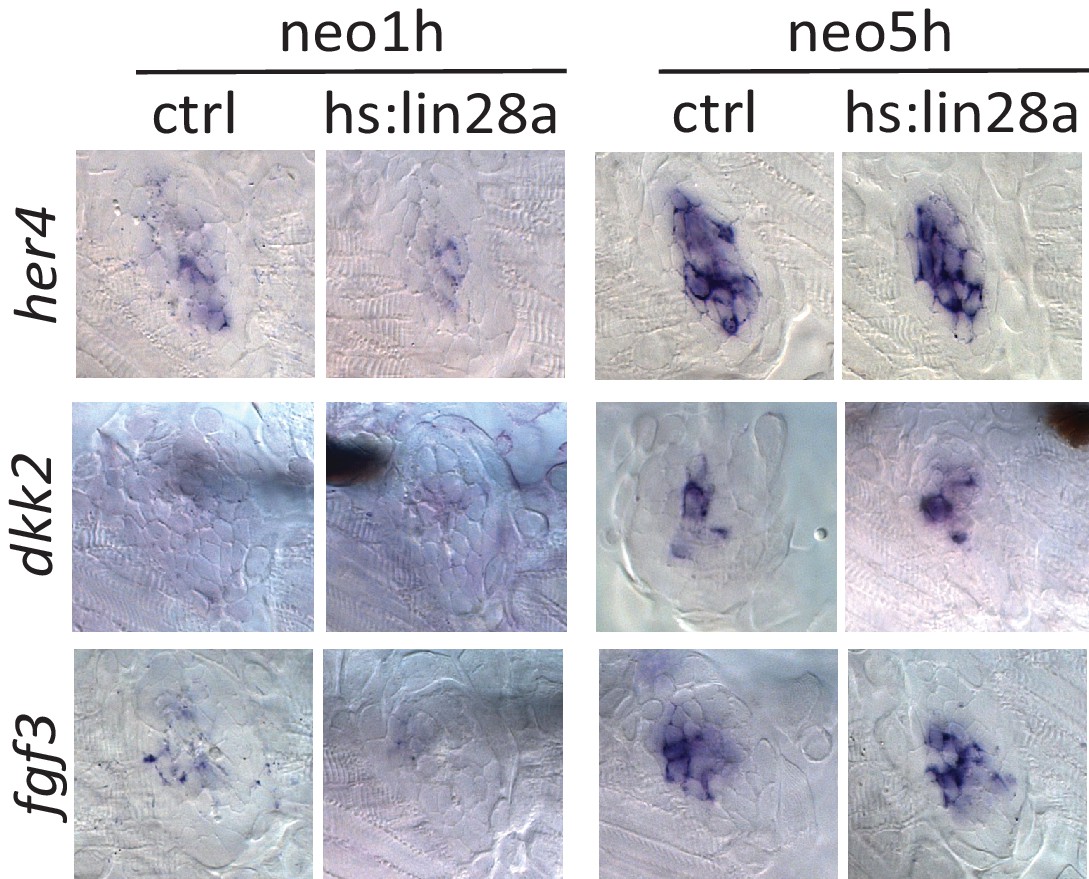
Notch and Fgf pathways were not affected by lin28a overexpression.
In situ results show that expression of her4 and dkk2, Notch pathway genes, and fgf3, a Fgf pathway gene, were not affected in hs:lin28a post neo.
Figure 7
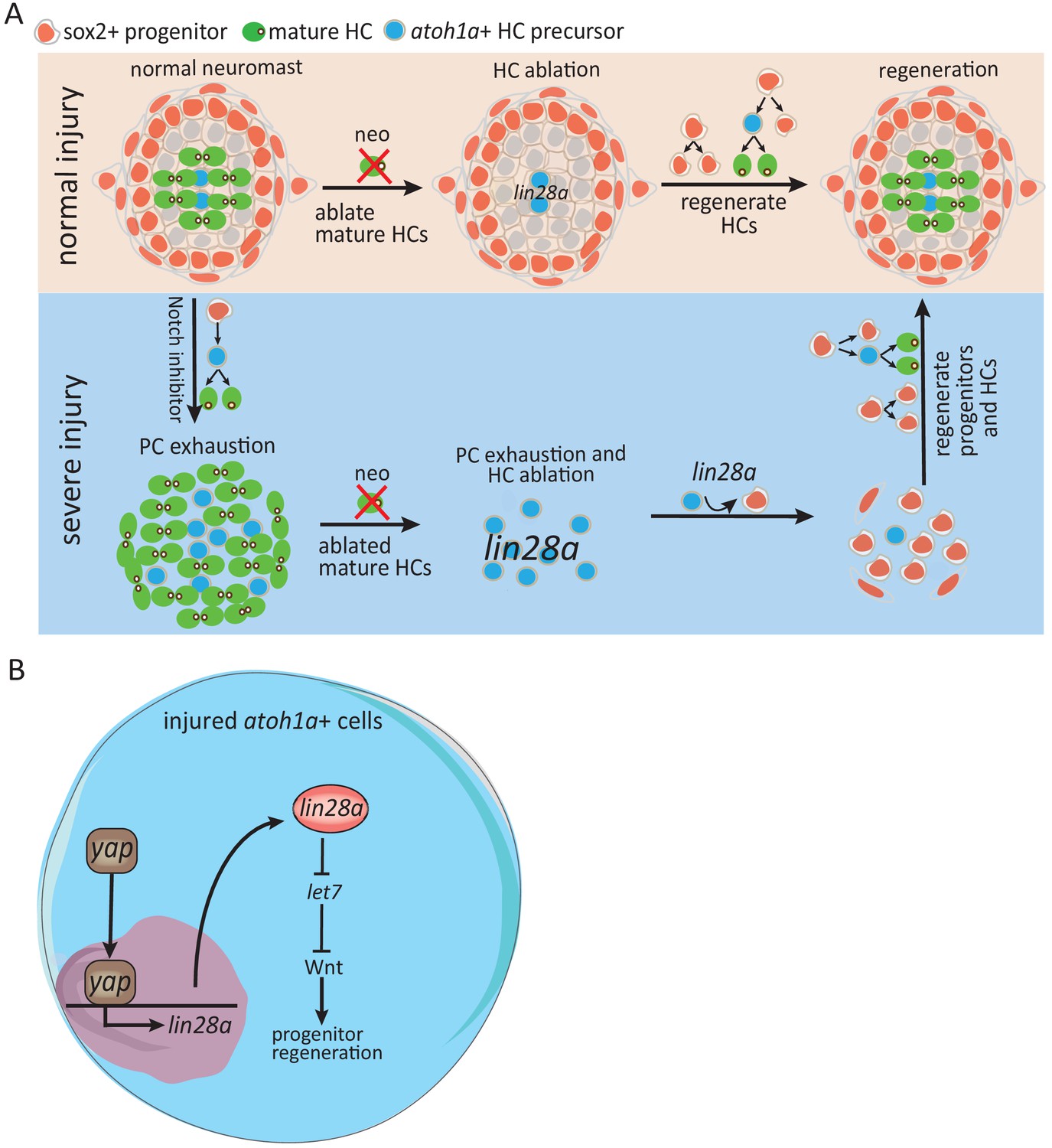
Schematic model illustrates how Yap-lin28a pathway regulates progenitor regeneration post severe injury.
(A) It’s known that sox2+ progenitors that are preserved post neo-induced HC injury divide and differentiate to regenerate HCs. To simulate the situation in mammalian inner ear where the progenitors are absent, we created a severe injury model by exhausting sox2+ progenitors with Notch inhibitor and ablating mature HCs with neomycin. We found that the exhausted sox2+ progenitors in severe injury have high potential to restore themselves within 48 hr, with HCs being regenerated afterwards. The atoh1a+ HC precursors, the main population that survived post severe injury, were converted into proliferative progenitors to initiate regeneration through yap-lin28a pathway. (B) Yap is activated in atoh1a+ HC precursors post severe injury and binds directly to lin28a promoter for initiating its transcription. Lin28a activates Wnt pathway through microRNA let7 to promote regenerative proliferation.
Author response image 1
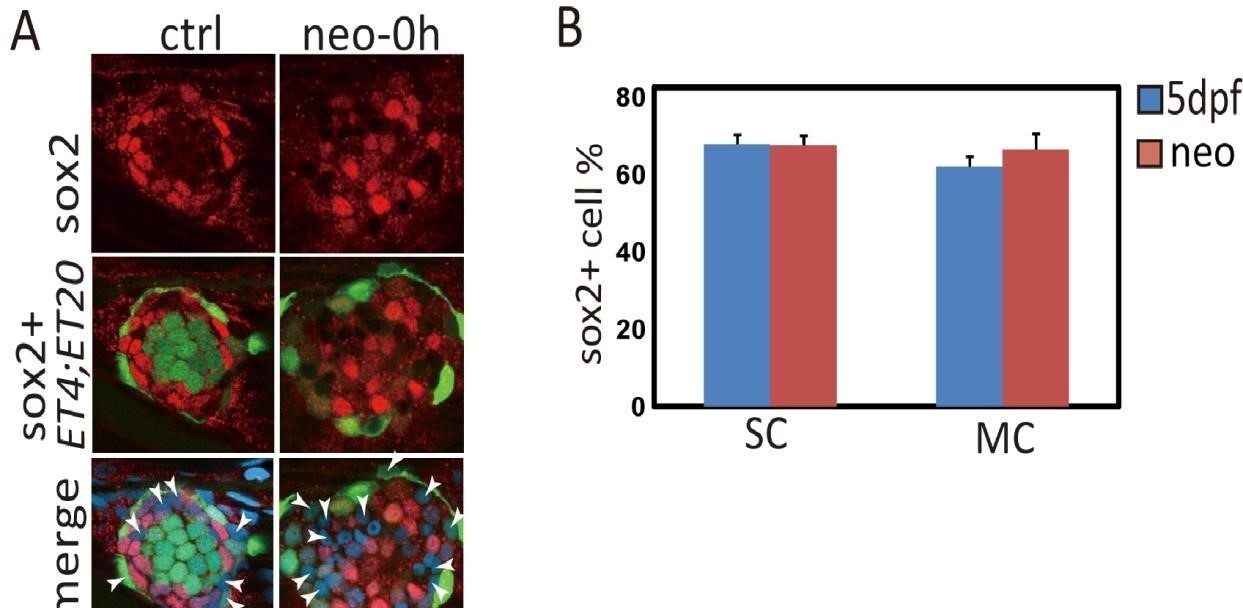
Some SCs and MCs in neuromast are negative with sox2 expression.
(A)ET4;ET20 Larvae were treated with neo and immediately collected for sox2 immunostaining. The white arrowheads pointed out SCs and MCs that were sox2 negative. (B) Total number of SCs or MCs and sox2+ ones were counted, and sox2+ ratios were calculated. About 60% of SCs or MCs express sox2.
Author response image 2
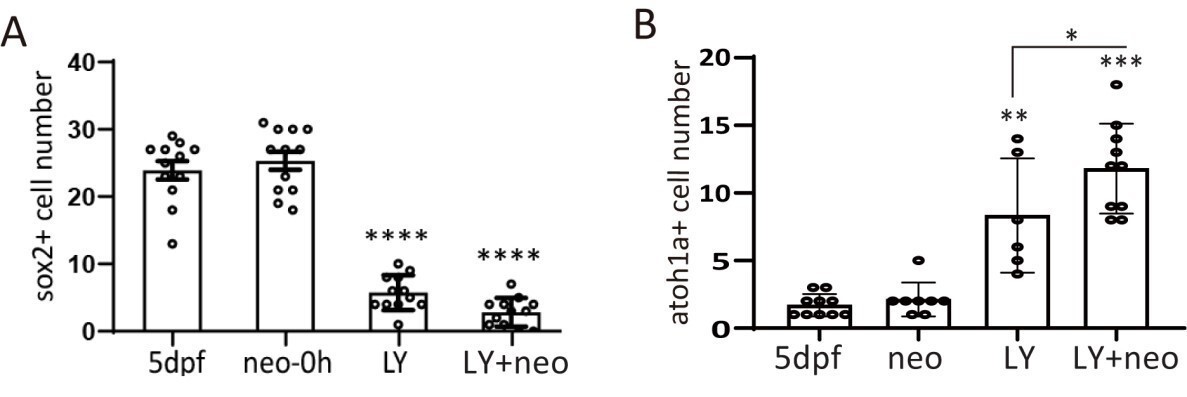
Sox2+ progenitors were eliminated and atoh1a+ HC precursors dominated post severe injury.
(A) Larvae were treated with LY, neo or LY+neo and collected immediately after neo treatment for sox2 immunostaining. Sox2+ cell numbers were counted, and the results showed that very few sox2+ cells survived post LY+neo. (B) Larvae were treated with LY, neo or LY+neo and collected immediately after neo treatment for atoh1a RNA probe hybridization. Results showed that significantly higher number of atoh1a+ cells survived post LY+neo.
Videos
Video 1
ET4;ET20;cldnB:H2Amcherry larvae treated with LY+neo were processed for time lapse.
Results showed the intensive cell divisions (CDs) during severe-injury-induced regeneration. Scale bar equals 10 μm.
Video 2
The neuromast of atoh1a:TdTomato;brn3c:GFP larvae treated with neo was imaged for time lapse.
Result showed that one Tomato+ cell divided and turned into two GFP+hair cells post neo. Scale bar equals 10 μm.
Video 3
The neuromast of atoh1a:TdTomato;sox2:GFP larvae treated with LY+neo was imaged for time lapse.
One Tomato+ cell (blue dot) divided and turned into two HCs that are GFP negative in the center. The other two Tomato+ cells (white dots) divided and converted into four sox2+ progenitors. Scale bar equals 10 μm.
Tables
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Genetic reagent (Danio rerio) | Et(krt4:EGFP)sqet20ET | Parinov et al., 2004 | sqET20; RRID:ZFIN_ZDB-ALT-070628-20 | |
| Genetic reagent (Danio rerio) | Et(krt4:EGFP)sqet4ET | Parinov et al., 2004 | sgET4; RRID:ZFIN_ZDB-GENO-110323-1 | |
| Genetic reagent (Danio rerio) | Tg(atoh1a: TdTomato)nns8 | Wada et al., 2010 | nns8; RRID:ZFIN_ZDB-GENO-120530-1 | |
| Genetic reagent (Danio rerio) | sox2-2a-sfGFPstl84 | Shin et al., 2014 | stl84; RRID:ZFIN_ZDB-GENO-150721-11 | |
| Genetic reagent (Danio rerio) | Tg(brn3c:GAP43-GFP) s356t | Xiao et al., 2005 | S356t; RRID:ZFIN_ZDB-GENO-200218-3 | |
| Genetic reagent (Danio rerio) | Tg(hsp70l:dkk1-GFP)w32 | Stoick-Cooper et al. (2007) | W32; RRID:ZFIN_ZDB-ALT-070403-1 | |
| Genetic reagent (Danio rerio) | Tg(hsp70l:myc-notch1a;cryaa:Cerulean)fb12 | Zhao et al. (2014) | fb12; RRID:ZFIN_ZDB-ALT-140522-5 | |
| Genetic reagent (Danio rerio) | Tg(hsp70:atoh1a)x20 | Millimaki et al. (2010) | X20; RRID:ZFIN_ZDB-GENO-110315-10 | |
| Genetic reagent (Danio rerio) | yapmw48 | Miesfeld et al., 2015 | Ms48; RRID:ZFIN_ZDB-ALT-160122-5 | |
| Genetic reagent (Danio rerio) | Tg(hsp70:lin28a-P2Amcherry;cmlc:GFP)psi30 | this paper | Details in Fish strain information | |
| Genetic reagent (Danio rerio) | Tg(hsp70:let7-P2AGFP;cryaa:Venus) | this paper | Details in Fish strain information | |
| Genetic reagent (Danio rerio) | lin28apsi37 | this paper | Details in Fish strain information | |
| Antibody | anti-sox2 (Rabbit, polyclonal) | Abcam | Cat# Ab97959; RRID:AB_2341193 | IF(1:200) |
| Antibody | anti-yap (Rabbit, polyclonal) | CST | Cat# 4912, RRID:AB_2218911 | IF(1:200) |
| Antibody | anti-taz (Rabbit, polyclonal) | Abcam | Cat# Ab84927; RRID:AB_1925489 | IF (1:200), WB (1:500) |
| Antibody | anti-GFP (Mouse, monoclonal) | Invitrogen | Cat# A11120; RRID:AB_221568 | IF(1:500) |
| Antibody | Anti-yap (Mouse, monoclonal) | Santa Cruz | Cat# sc-271134; RRID:AB_10612397 | WB (1:1000) |
| Antibody | Anti-lin28a (Rabbit, polyclonal) | CST | Cat# 3978; RRID:AB_2297060 | WB (1:500) |
| Antibody | Anti-lats (Rabbit, monoclonal) | CST | Cat# 3477; RRID:AB_2133513 | WB (1:500) |
| Antibody | Anti-p-mob1 (Rabbit, monoclonal) | CST | Cat# 8699; RRID:AB_11139998 | WB (1:500) |
| Antibody | Anti-β-actin (Mouse, monoclonal) | Sigma | Cat# A1978; RRID:AB_476692 | WB (1:2000) |
| Antibody | Anti-digoxingenin POD (sheep, polyclonal) | Roche | 11207733910; RRID:AB_514500 | 1:2000 |
| Antibody | Anti-fluorescein POD(sheep, polyclonal) | Roche | 11426346910; RRID:AB_840257 | 1:2000 |
| Chemical compound, drug | EdU | Carbosynth | NE08701 | 3.3 mM |
| Chemical compound, drug | LY411575 | Santa Cruz | sc-364529 | 2 μM |
| Chemical compound, drug | Neomycin sulfate | Sigma | N6386 | 300 μM |
| Chemical compound, drug | Alexa Fluor-594 Azide | Thermo Fisher Scientific | N6386 | |
| Chemical compound, drug | Verteporfin | Selleckchem | S1786 | 5 μM |
| Chemical compound, drug | Copper sulfate | Sigma | 451657 | 50 μM |
| Chemical compound, drug | cisplatin | Sigma | 33342 | 500 μM |
| Commercial assay or kit | TSA-Cyanine 3 Reagent | PerkinElmer | SAT704A001EA | |
| Commercial assay or kit | TSA-FITC Reagent | PerkinElmer | SAT704A001EA | |
| Commercial assay or kit | dynabeads | Invitrogen | 10015D |
Additional files
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Yap-lin28a axis targets let7-Wnt pathway to restore progenitors for initiating regeneration
eLife 9:e55771.
https://doi.org/10.7554/eLife.55771




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}