A multi-layered and dynamic apical extracellular matrix shapes the vulva lumen in Caenorhabditis elegans
- Department of Genetics, University of Pennsylvania Perelman School of Medicine, United States
- Department of Neuroscience, Albert Einstein College of Medicine, United States
- Department of Biological Chemistry, David Geffen School of Medicine, University of California, Los Angeles, United States
Figures
Figure 1

Introduction to vulva development.
(A) Cartoon of early L3 larva, indicating the six vulva precursor cells (VPCs) beneath the somatic gonad. (B) Vulva lineages and cell types. An EGF-like signal from the gonadal anchor cell (AC) induces the primary (1°) cell fate in the nearest VPC (black), which then expresses DSL ligands to induce the secondary (2°) cell fate in the adjacent VPCs (gray). The 1° and 2° VPCs divide to generate a total of 22 descendants of 7 different cell types. (C) Cartoon of mid L4 larva, showing the vulva lumen. (D) L4.4 stage vulva cells visualized with the membrane marker MIG-2::GFP (muIs28). The 22 vulval cells are organized into 7 stacked rings (Sharma-Kishore et al., 1999). In the standard lateral or sagittal view, anterior is to the left and ventral is down. An orthogonal XZ view shows the oblong shape of the lumen. (E) 3D rendering of the L4.4 vulva lumen generated with Imaris software (BitPlane, Zurich Switzerland), based on imaging of the matrix factor FBN-1 (see Figure 3). The YZ view at right is comparable to the transverse views of the vulva seen by TEM in Figures 4B and 9C (but note that regions deeper in the Z plane are poorly resolved here). (F) The adult vulva is a slit-like and cuticle-lined passageway through which eggs are laid.
Figure 2
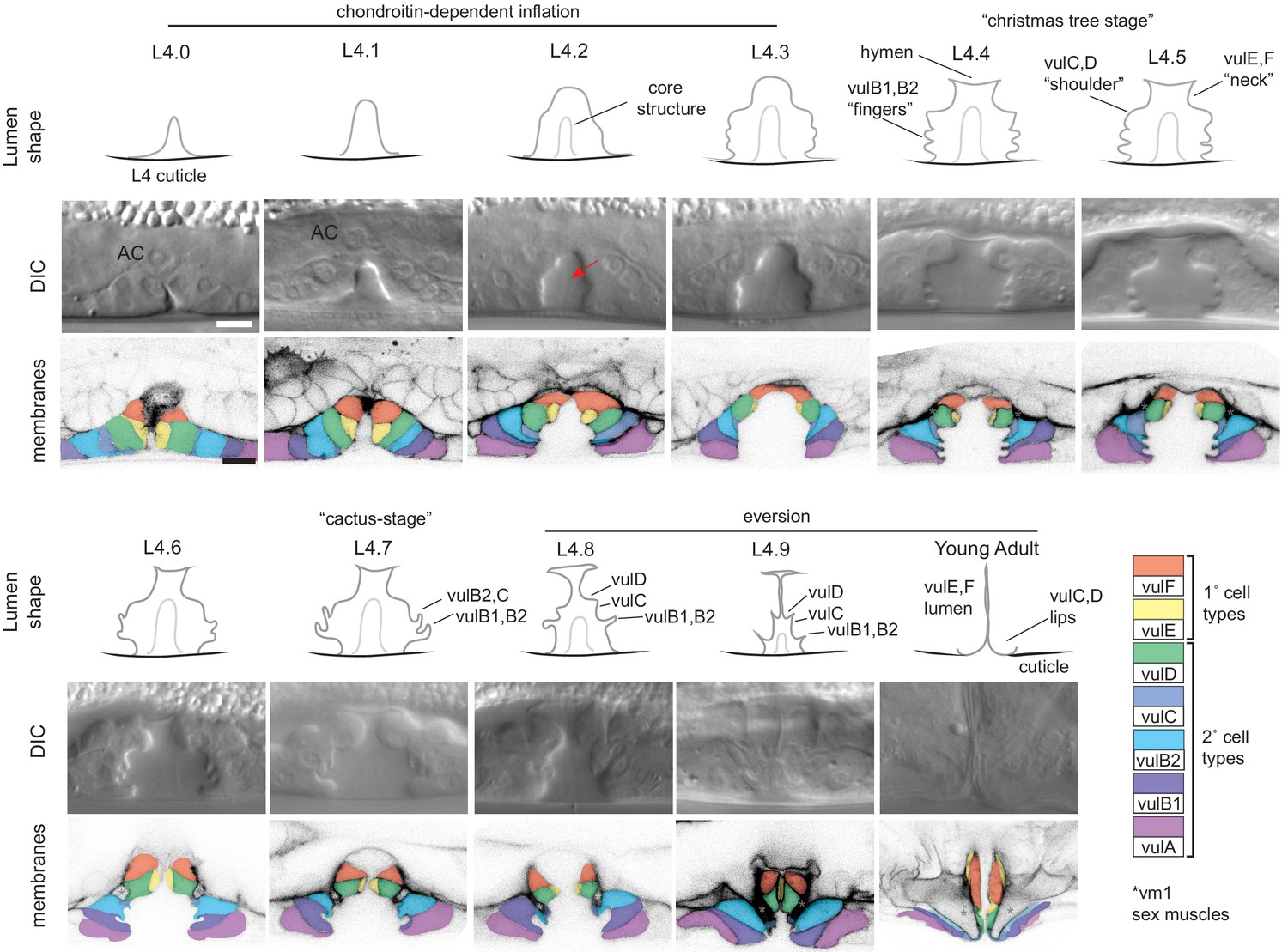
Cell and lumen shape changes during vulva morphogenesis Sagittal views of the central vulva lumen.
Top rows show cartoons of lumen shape for each L4 sub-stage, as defined by Mok et al., 2015. Middle rows show corresponding DIC images. Bottom rows show confocal slices of vulva cell membranes marked by MIG-2::GFP (muIs28); cells are colored according to the key at right. Confocal stacks were collected for at least three animals per stage after L4.3. Luminal core structure is faintly visible beginning at L4.2 (red arrow). At mid-L4 (‘Christmas-tree stage’; Seydoux et al., 1993), the vulF and vulE cells together define the vulva ‘neck’, the vulD and vulC cells define the vulva ‘shoulder’, the vulB1 and vulB2 cells define the vulva ‘fingers’, and the vulA cells make the connection between the vulva cells and the surrounding epidermis. *vm1 sex muscles, which attach to the mature vulva between the vulC and vulD toroids (Sharma-Kishore et al., 1999). Scale bars, five microns.
Figure 3

A dynamic aECM fills the vulva lumen during morphogenesis.
(A) aECM protein schematics. Genbank accession: FBN-1a (AFN70749.1), LET-653b (CAH60755.1), NOAH-1a (CCD66686.2), LPR-3 (CAA92030.1), LET-4 (AEZ55699.1), SYM-1 (CAB43345). Full-length FBN-1::mCherry and some LET-653 fusions were expressed from transgenes; all others were expressed from the endogenous loci tagged by CRISPR-Cas9 genome editing (see Materials and methods). (B) LET-653::SfGFP (csIs64) and FBN-1::mCherry (aaaIs12) show complementary luminal patterns. Medial confocal slices. Arrow, luminal core. Arrowheads, sites of ventro-lateral fibril attachment to vulva cells. Lines, membrane-proximal matrix over 1° cells vulE and vulF. Bracket indicates loss of FBN-1 from the lumen over 1° cells during vulva eversion (n = 3/5 L4.7, 4/4 L4.8). (C) NOAH-1::mCherry (mc68) labels matrix spikes that connect to LET-653-marked luminal fibrils during vulva eversion. Left column shows overlay with cell membrane marker MIG-2::GFP (muIs28) (n = 9 L4.7-L4.9). Right columns show overlay with LET-653::SfGFP (cs262) (n = 12 L4.7-L4.9). (D) Timeline of vulva morphogenesis showing dynamic matrix patterns. Each image is a single confocal slice, inverted for clarity. For each fusion, images were collected for at least three animals per stage per strain; most fusions were imaged in multiple different strains to directly compare the different patterns (as in panels B and C). Solid green underlines indicate stages with consistent and peak localization; dashed green underlines indicate stages with more variable or weak localization. Fusions shown are FBN-1::mCherry (aaaIs12), LET-653::SfGFP (cs262), LET-653(ZP)::SfGFP (csIs66), SfGFP::LPR-3 (cs250), NOAH-1::mCherry (mc68), mCherry::LET-4 (cs265), and SYM-1::GFP (mc85).
Figure 4 with 1 supplement
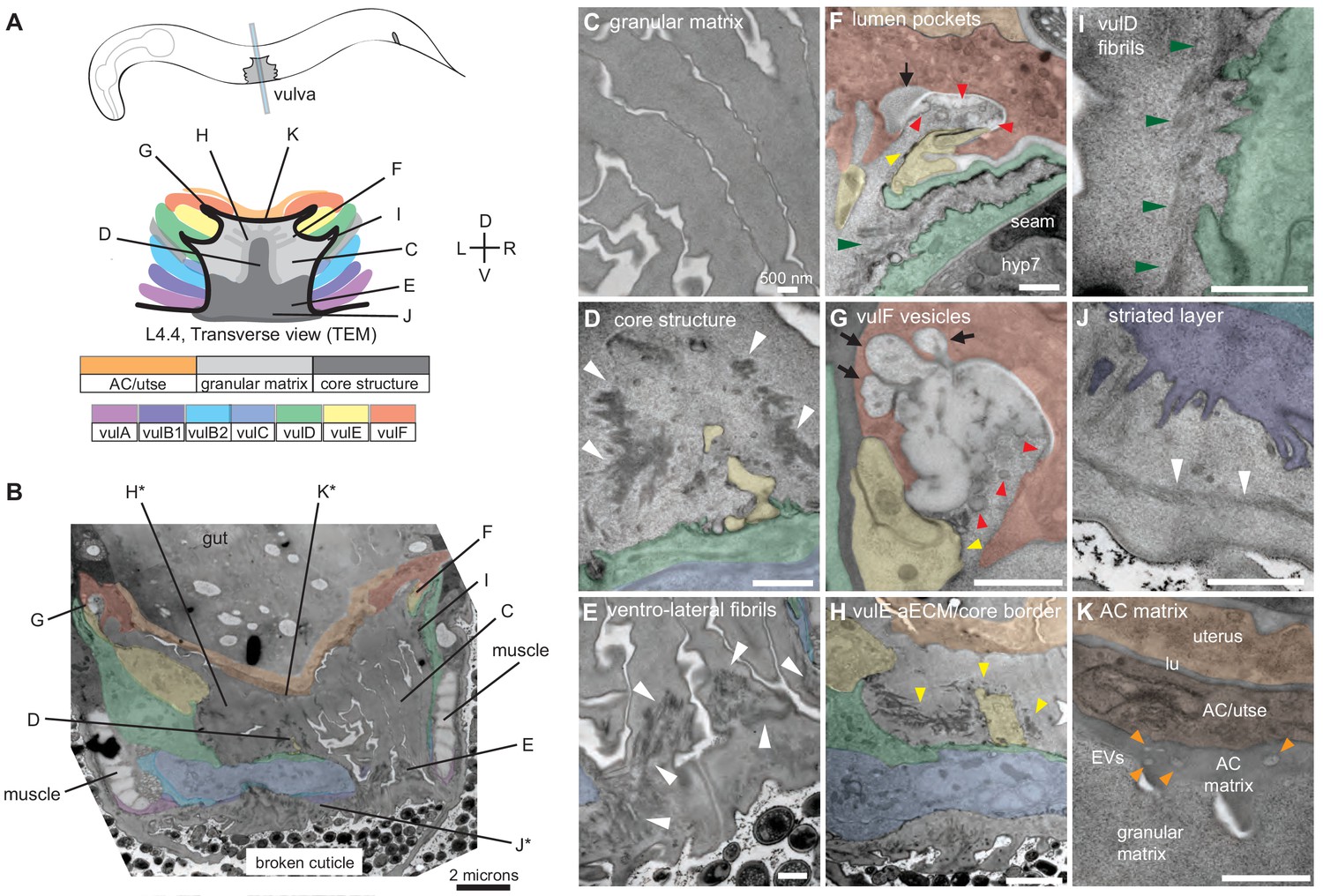
Ultrastructural features of the mid-L4 vulva aECM.
(A) Transverse serial thin sections of an N2 L4.4-L4.5 stage animal were analyzed by TEM. The cartoon depicts the vulva lumen in this orientation (see also Figure 1E), and lines indicate the relative locations of different panel images. (B) Whole vulva view. This thin section captures a portion of the lumen and the cell borders. Vulva cells and AC/utse are pseudo-colored according to the key shown in A. Lines indicate the relative locations of different panel images, and asterisks indicate that the panel shows a region from a different thin section of the same animal. The ventral cuticle has broken during specimen processing and oval objects surrounding the specimen are E. coli bacteria. (C) A rough granular matrix fills the dorsal lumen. (D) Core structure (white arrowheads) rises above vulC and vulD, to the level of vulE. (E) Ventro-lateral fibrils (white arrowheads) and the ventral edge of the luminal matrix, which has pulled away from the broken cuticle. (F) The interface between vulF (red) and vulE (yellow) forms a sequestered lumen pocket where matrix accumulates (red and yellow arrowheads). vulD (green) forms another narrow lumen pocket that is densely populated with fine fibrils (green arrowhead). (G) vulF cells contain large secretory vesicles (black arrows) that are open to the extracellular space and whose contents resemble the membrane-proximal matrices that line vulF and vulE (red and yellow arrowheads, respectively). (H) Lateral view of the matrix lining vulE surfaces. (I, J) Examples of the very protrusive surfaces of 2°-derived cells (vulD and vulB1, respectively) that interface with fibrils. J also shows the ventral-most border of the aECM, which contains a striated layer (white arrowheads) similar to that seen in epidermal cuticle (Page, 2007). (K) A fine-grained aECM separates the AC/utse from the rougher granular matrix of the vulva lumen; this AC matrix contains numerous EVs (orange arrowheads). All scale bars are 500 nm unless otherwise indicated. See Figure 4—figure supplement 1 for uncolored versions of all images.
Figure 4—figure supplement 1
Ultrastructural features of the mid-L4 vulva aECM.
Uncolored images from Figure 4 are shown.
Figure 5 with 1 supplement
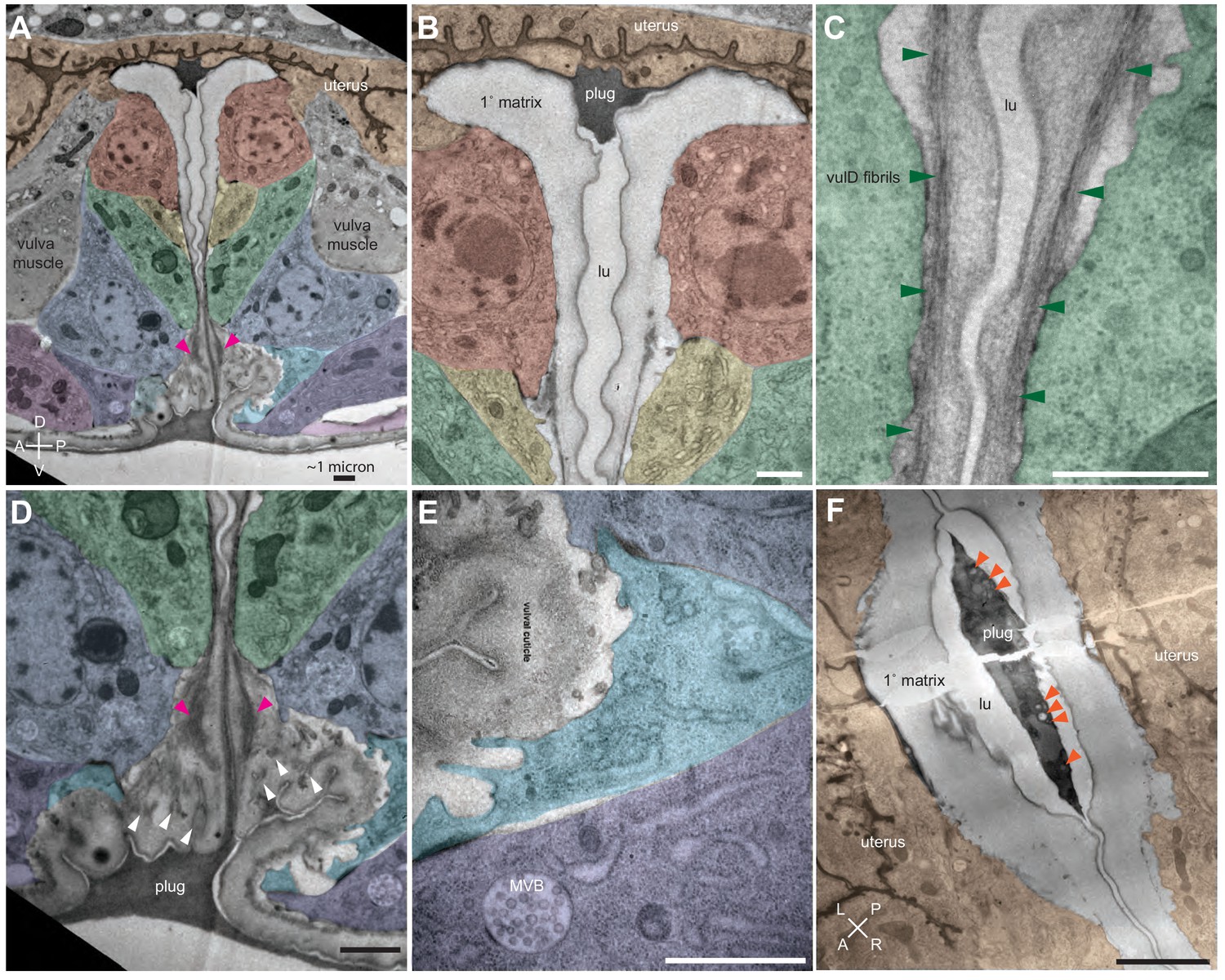
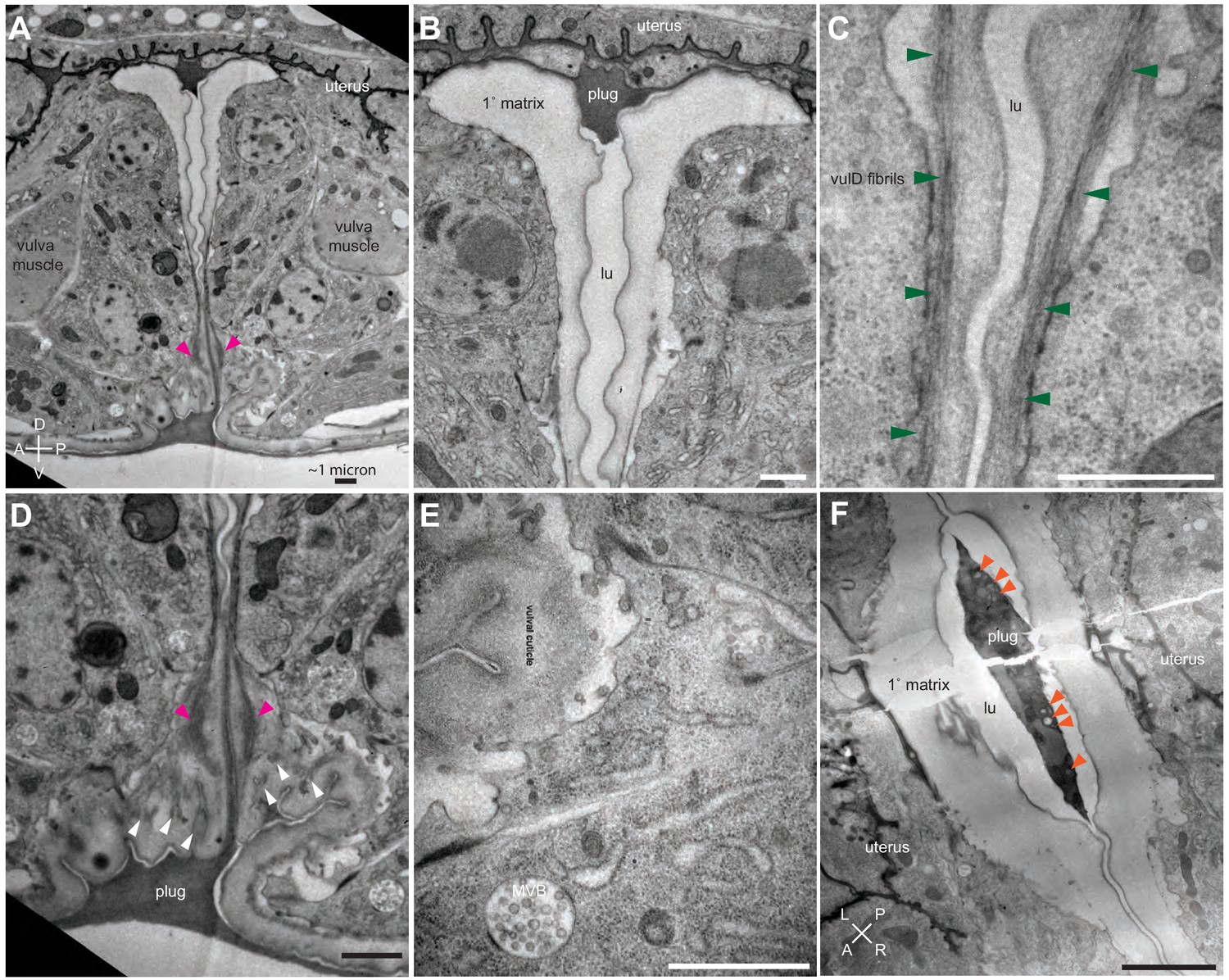
Ultrastructural features of the late L4 vulva aECM.
(A) Longitudinal slice through the vulva of an N2 L4.8-L4.9 stage animal, with orientation similar to that in confocal images. Vulva and uterine cells are pseudo-colored as in Figure 4B. Pink arrowheads indicate matrix spikes as observed with NOAH-1::mCherry (see Figure 3C). (B–E) Higher magnification views of the specimen in panel A. (B) Primary vulva cells are covered in a thick membrane-proximal matrix, and the dorsal-most edge of the lumen is filled with a plug of darkly-staining material. (C) The membrane-proximal matrix continues over vulD and vulC, but becomes filled with dense fibrils (green arrowheads). (D) Matrix spikes (pink arrowheads) extend from vulC/D into a cuticle-like matrix below. Various other fibrils (white arrowheads) are present within this ventral matrix. (E) Multi-layered nascent cuticle over vulB2. Note protrusive surface of vulB2 and multi-vesicular body (MVB) within vulB1. Many MVBs are present in vulva cells at this stage. (F) Longitudinal dorsoventral slice through uterine cells and the primary vulva matrix and plug of a second N2 L4.8-L4.9 stage animal. Note the numerous EVs (orange arrowheads) present within the plug. All scale bars, one micron. See Figure 5—figure supplement 1 for uncolored versions of all images.
Figure 5—figure supplement 1
Ultrastructural features of the late L4 vulva aECM.
Uncolored images from Figure 5 are shown.
Figure 6
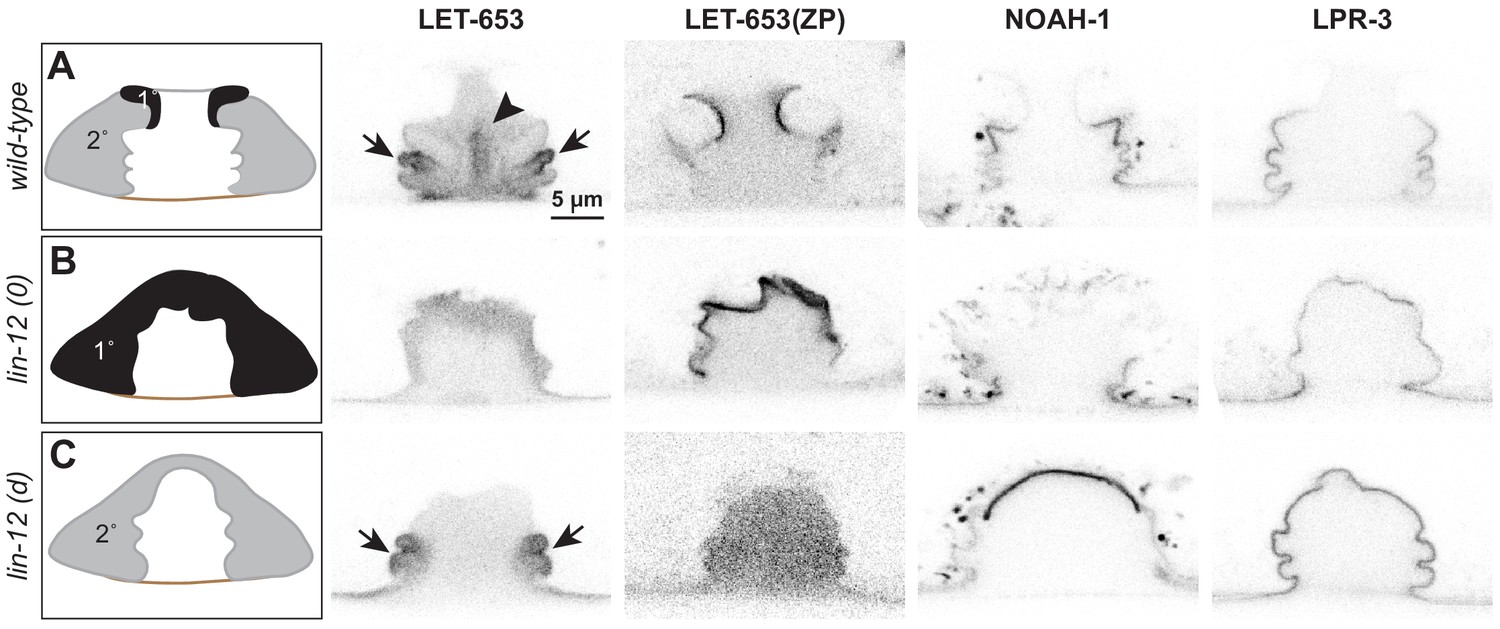
Different vulva cell types produce and assemble different aECMs.
(A–C) Panels in left column show cartoons of vulva cell types and lumen shape at mid-L4. Remaining columns show single confocal slices through the vulva lumen. Fusions used are LET-653(full-length)::SfGFP (cs262), LET-653(ZP)::SfGFP (csIs66), NOAH-1::mCherry (mc68), SfGFP::LPR-3 (cs250). At least n = 8 L4s were imaged for each strain. (A) In wild-type animals, full-length LET-653 predominantly labels the core and ventro-lateral fibrils, LET-653(ZP) labels the membrane-proximal matrix over 1° cells, NOAH-1 labels membrane-proximal matrices over 2° cells (especially vulC and vulD), and LPR-3 transiently labels membrane-proximal matrices over all cells, but then becomes concentrated over 2° cells (see also Figure 3D). (B) Loss of 1° cells in lin-12(0) (null, n137n720) mutants disrupted the luminal core and NOAH-1 localization. Most of the NOAH-1 pattern here is intracellular. (C) Loss of 2° cells in lin-12(d) (hypermorphic, n137) mutants disrupted the luminal core and LET-653(ZP) localization.
Figure 7
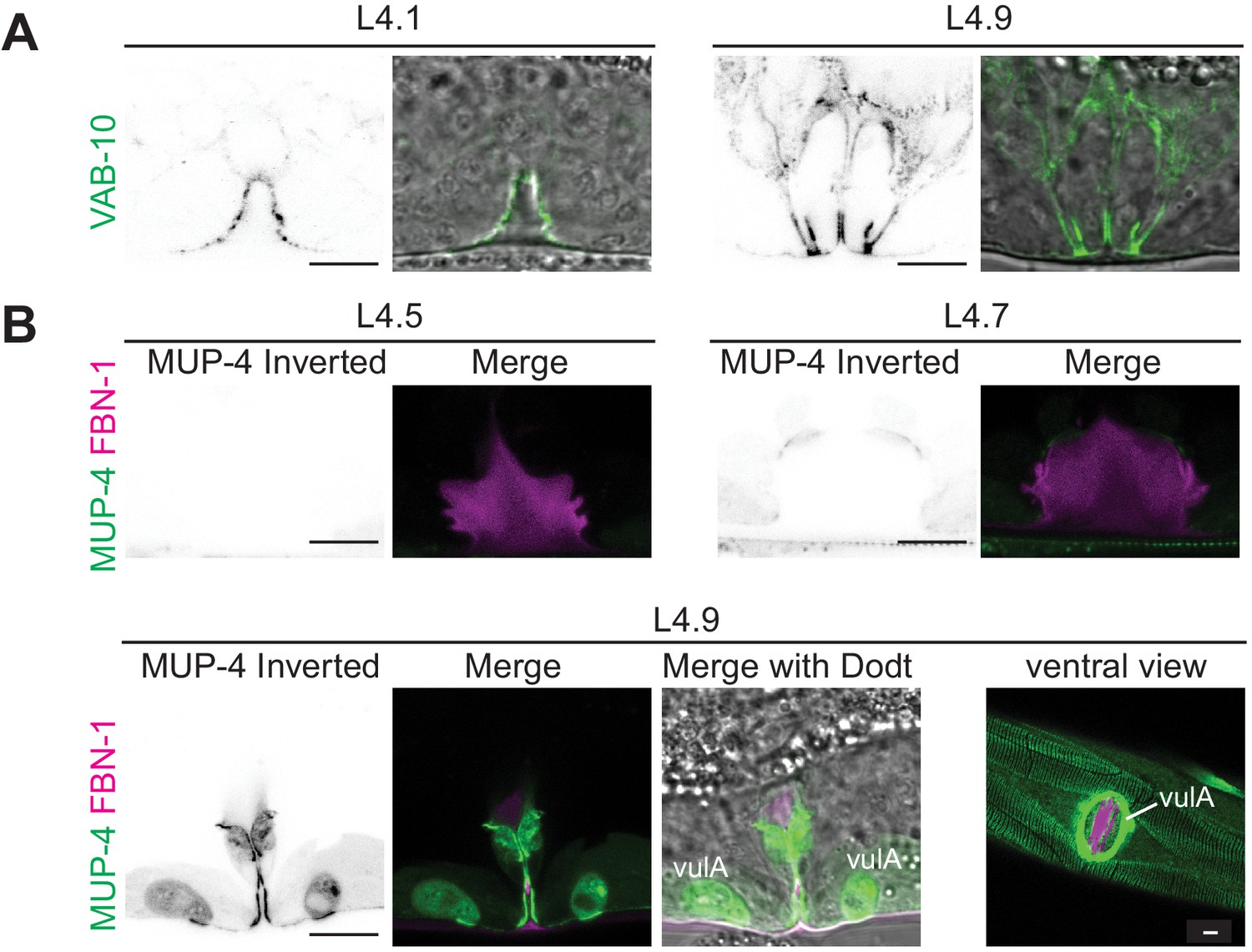
Vulva aECM assembles prior to expression of MUP-4/matrillin.
(A) VAB-10::GFP (cas627) marked all apical membranes in the vulva throughout L4 (including 4/4 L4.1/L4.2 stage animals). (B) MUP-4::GFP (upIs1) marked apical membranes beginning in late L4 (0/3 L4.4/L4.5, 3/3 L4.6/L4.7, 2/2 L4.9). FBN-1::mCherry (aaaIs12 or aaaEx78) is also shown. At L4.9, MUP-4 expression is particularly strong in the vulA toroid, which surrounds the vulva opening and connects to the surrounding hyp7 epidermis. The remaining FBN-1 matrix connects to vulA at the left and right sides of the lumen, as seen in the ventral view.
Figure 8 with 2 supplements
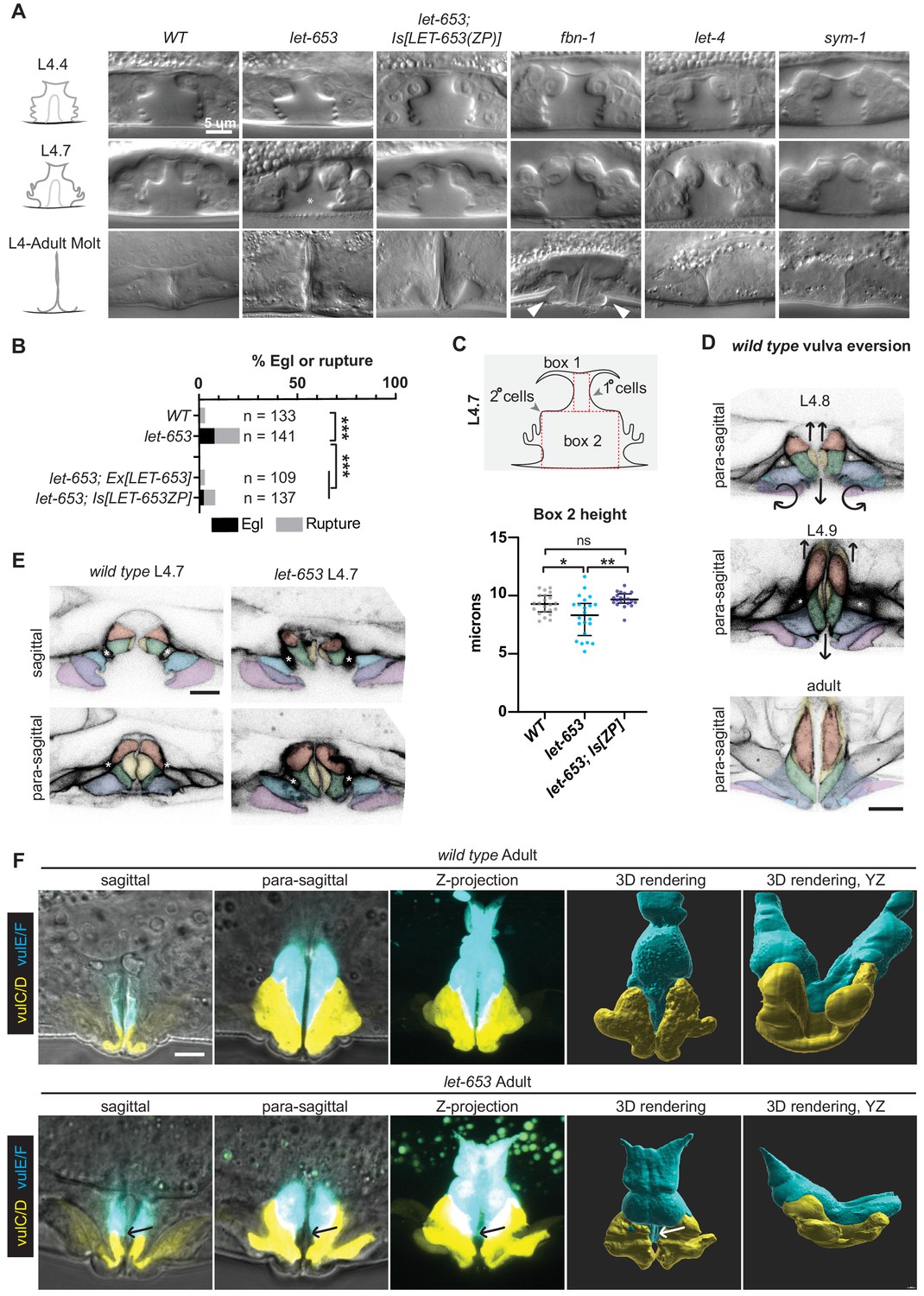
Individual aECM factors play subtle roles in vulva eversion.
(A) DIC images of mutant vulvas at L4.4, L4.7 and L4.9-adult molt. At least 40 L4 animals of each genotype were imaged, including at least five each of the three stages shown. Alleles used: fbn-1(tm290), let-653(cs178) (strains UP3342 and UP3422), let-4(mn105), and sym-1(mn601). Asterisk indicates collapsed lumen morphology in some let-653 mutants (n = 6/22 L4.7, see panel C). Arrowheads indicate abnormal bulges of the vulA and vulB1/B2 cells in fbn-1 mutants (n = 9/13 L4.9). (B) A small proportion of let-653 mutants had progeny that hatched in utero (Egl phenotype) or ruptured at the vulva within eight days of reaching adulthood. These phenotypes were rescued by transgenes expressing full-length LET-653 or just the ZP domain. ***p<0.0001, Fisher’s exact test. (C) Let-653 mutants have reduced lumen dimensions at the L4.7 stage (n = 22). To measure lumen dimensions, the largest box possible was drawn within the 1°-generated (Box 1) and 2°-generated (Box 2) lumen spaces, as visualized by DIC. Dimensions for Box 1 did not differ significantly between genotypes (Figure 8—figure supplement 1), but Box 2 height was somewhat reduced. This phenotype was rescued by a transgene expressing the LET-653 ZP domain. *p=0.031, **p=0.001, WT vs. let-653;Is[ZP] p=0.085, Mann–Whitney U test. (D) WT vulva eversion. Membranes were visualized with MIG-2::GFP. Compare these para-sagittal slices to sagittal slices of the same animals in Figure 2. n = 3 per stage. (E) let-653 mutants have irregular vulva cell shapes at the onset of vulva eversion (L4.7 stage). Membranes were visualized with MIG-2::GFP. Asterisks indicate the vm1 sex muscles. Both sagittal and para-sagittal slices from confocal Z-stacks are shown. In wild-type, vulva cells are symmetrical across the midline, but in let-653 mutants, cell shape and position are mismatched (n = 3/3; WT: n = 0/6). No defects in vulva cell fusion were observed (Figure 8—figure supplement 2). (F) let-653 mutants have subtly irregular vulva cell shapes as older L4s and adults. vulE and vulF were visualized with daf-6pro::CFP, and vulC and vulD were visualized with egl-17pro::CFP (Mok et al., 2015). Both sagittal slices and Z-projections from confocal Z-stacks are shown, along with three- dimensional renderings generated with Imaris (Bitplane) from those Z-stacks. In let-653 mutants, cell shape and position are variably abnormal (n = 4/4, WT: 0/3). In the specimen picture, vulE/F appear less elongated along the dorsal-ventral axis and vulC/D are slightly flattened relative to WT. Arrows indicate the transition zone between vulE/F and vulC/D, where an abnormal gap occurs in the mutant. In the YZ view, WT primary cells form a deep U-shape as they extend toward the seam cells, but let-653 primary cells form a much shallower curve.
-
Figure 8—source data 1
Percentage of WT and mutants C. elegans that fail to lay eggs.
- https://cdn.elifesciences.org/articles/57874/elife-57874-fig8-data1-v2.xlsx
-
Figure 8—source data 2
Measurements of WT and mutant L4.7 stage vulvas.
Measurements are taken from boxes drawn within DIC images of vulvas at L4.7 stage. See Figure 8 and Figure 8—figure supplement 1.
- https://cdn.elifesciences.org/articles/57874/elife-57874-fig8-data2-v2.xlsx
Figure 8—figure supplement 1
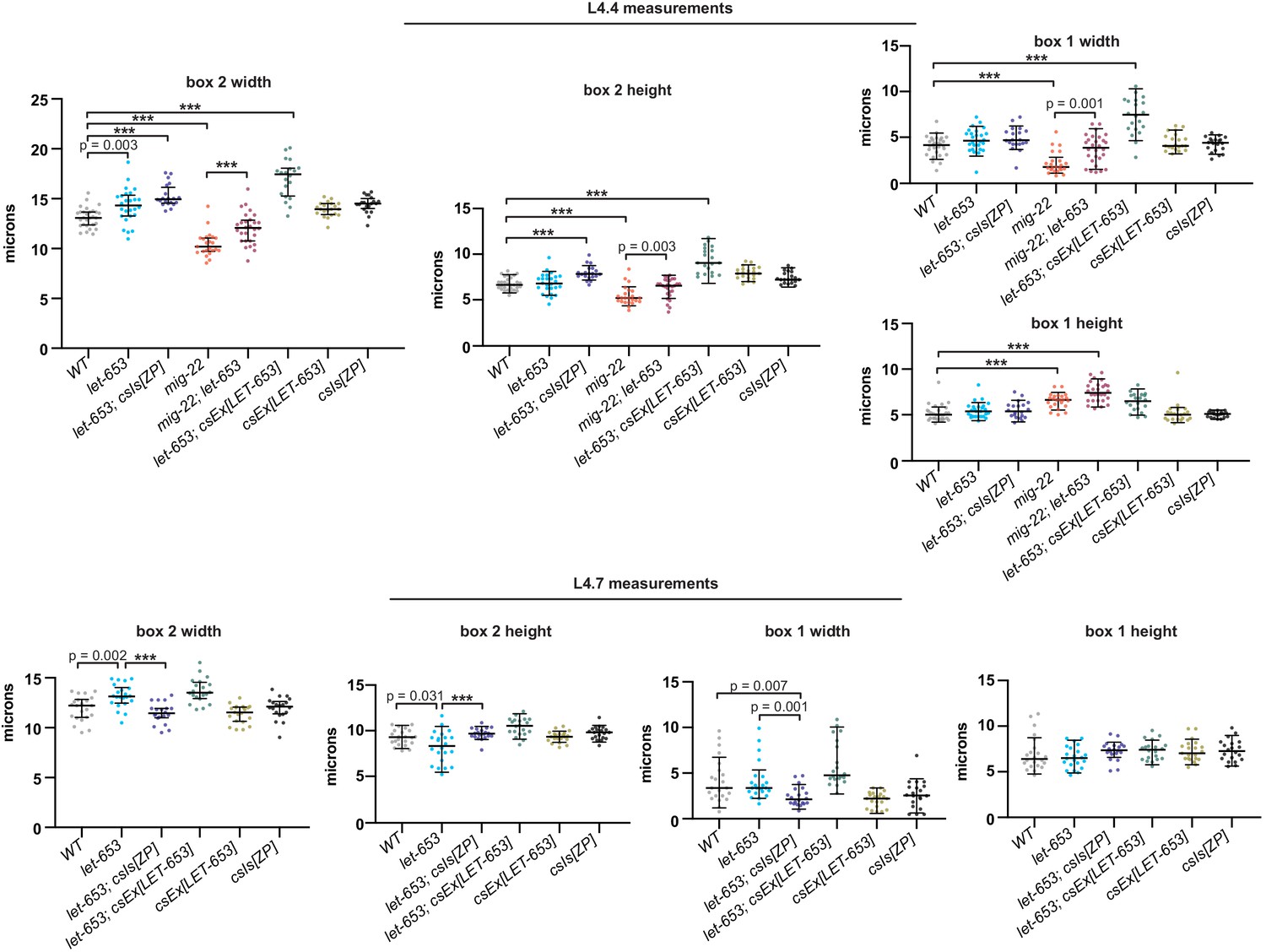
Measurements of let-653, mig-22, and LET-653+ vulvas Lumen dimensions at the L4.4 and L4.7 stages were quantified as in Figures 8 and 11.
***p<0.0001, Mann–Whitney U test. All p values between 0.01 and 0.0001 are reported. p values above 0.01 are not indicated. All measurements were performed by a researcher blinded to genotype. let-653 mutants had approximately wild-type vulvas at L4.4 stage but slightly misshapen vulvas by L4.7 stage. Overexpression of LET-653 caused expanded vulvas in a let-653 mutant background, but not in a WT background, possibly due to lower expression levels.
Figure 8—figure supplement 2
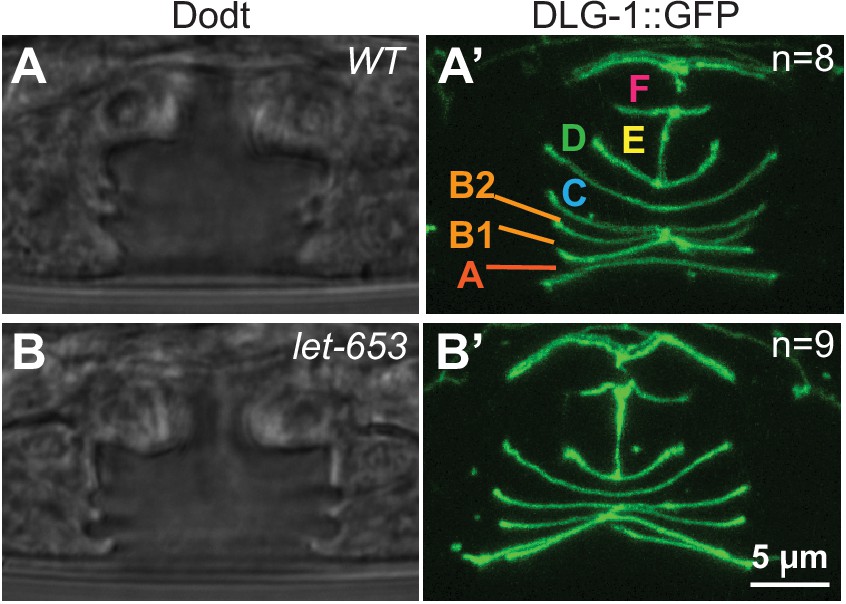
let-653 mutant vulvas have normal cell fusion L4 4 stage WT and let-653(cs178) vulvas with the apical junction marker DLG-1::GFP (mc103).
(A–A’) In WT, vulA, vulC, vulD, and vulF have fused to form toroids by this stage. (B–B’) Let-653 mutants show the same pattern of vulva cell fusion and general junction arrangement as WT. Images are maximum Z-projections of confocal Z-stacks.
Figure 9 with 1 supplement
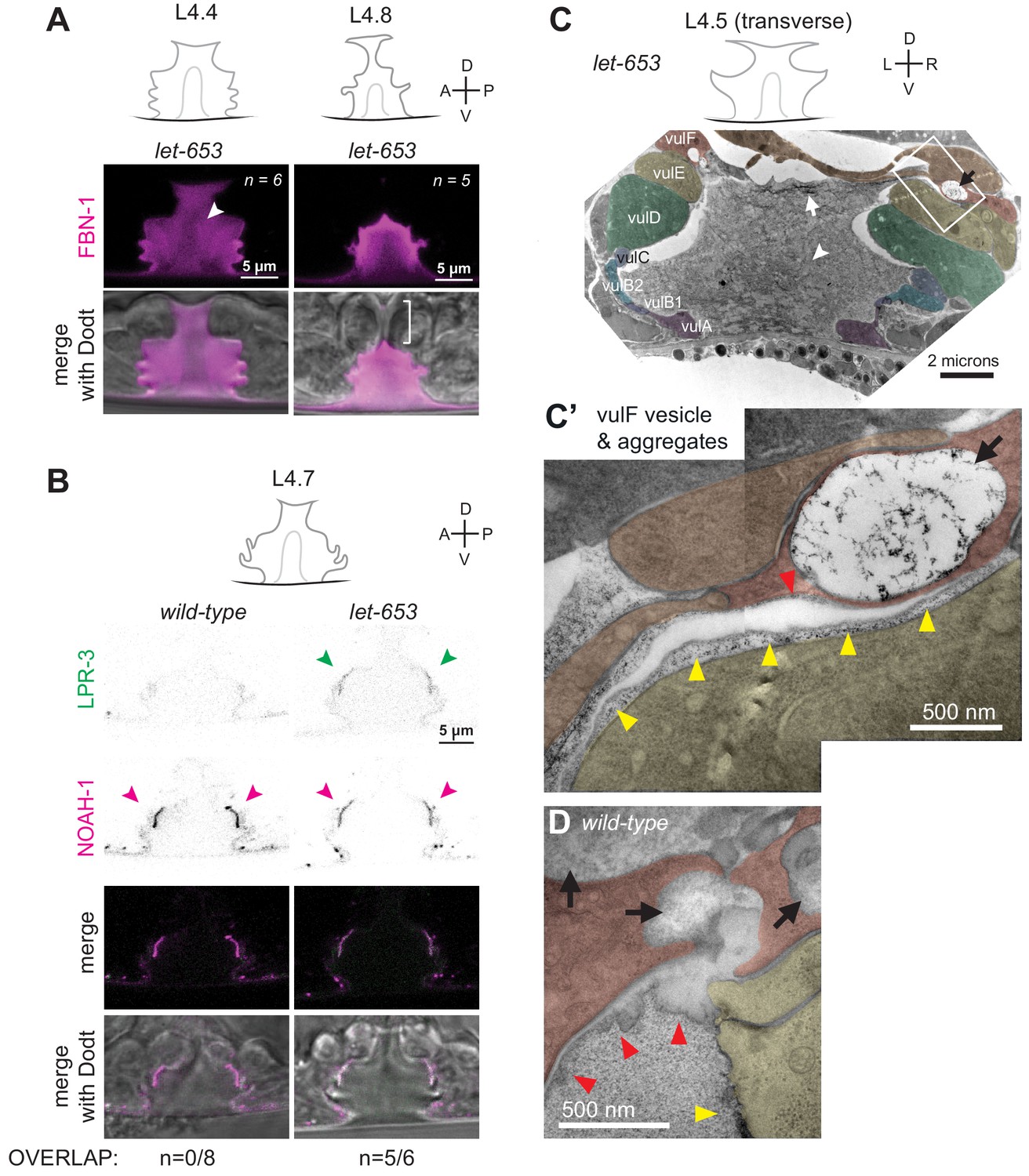
The ZP protein LET-653 is required for proper organization and remodeling of the vulva aECM.
(A) Let-653(cs178) mutants showed relatively normal patterns of FBN-1::mCherry (aaaIs12) localization. Arrowhead indicates exclusion of FBN-1 from the core region. Bracket indicates exclusion of FBN-1 from the 1° lumen during eversion (n = 5/5). Compare to WT in Figure 3B. (B) Let-653(cs178) mutants showed normal recruitment of SfGFP::LPR-3 (cs250) and NOAH-1::mCherry (mc68) to 2° surfaces, but abnormally delayed clearance of SfGFP::LPR-3 from vulC/D. p=0.0445, Fisher’s exact test. (C) Transverse TEM slice of a let-653(cs178) mutant at mid-L4 (~L4.5) stage. Compare to the WT mid-L4 specimen in Figure 4B. Fibrils are present near the AC/utse (arrow) and the core structure (arrowhead) is not well-defined, unlike in WT (see magnified images in Figure 9—figure supplement 1). Box indicates region magnified in C’. (C’) An abnormal secretory vesicle in vulF is filled with dark aggregates that match those present in the membrane-proximal matrix over vulF (red arrowhead) and vulE (yellow arrowheads). A similar matrix continues beneath the AC/utse (see also Figure 9—figure supplement 1). (D) WT vulF vesicles and matrix for comparison. See also Figure 4G.
Figure 9—figure supplement 1
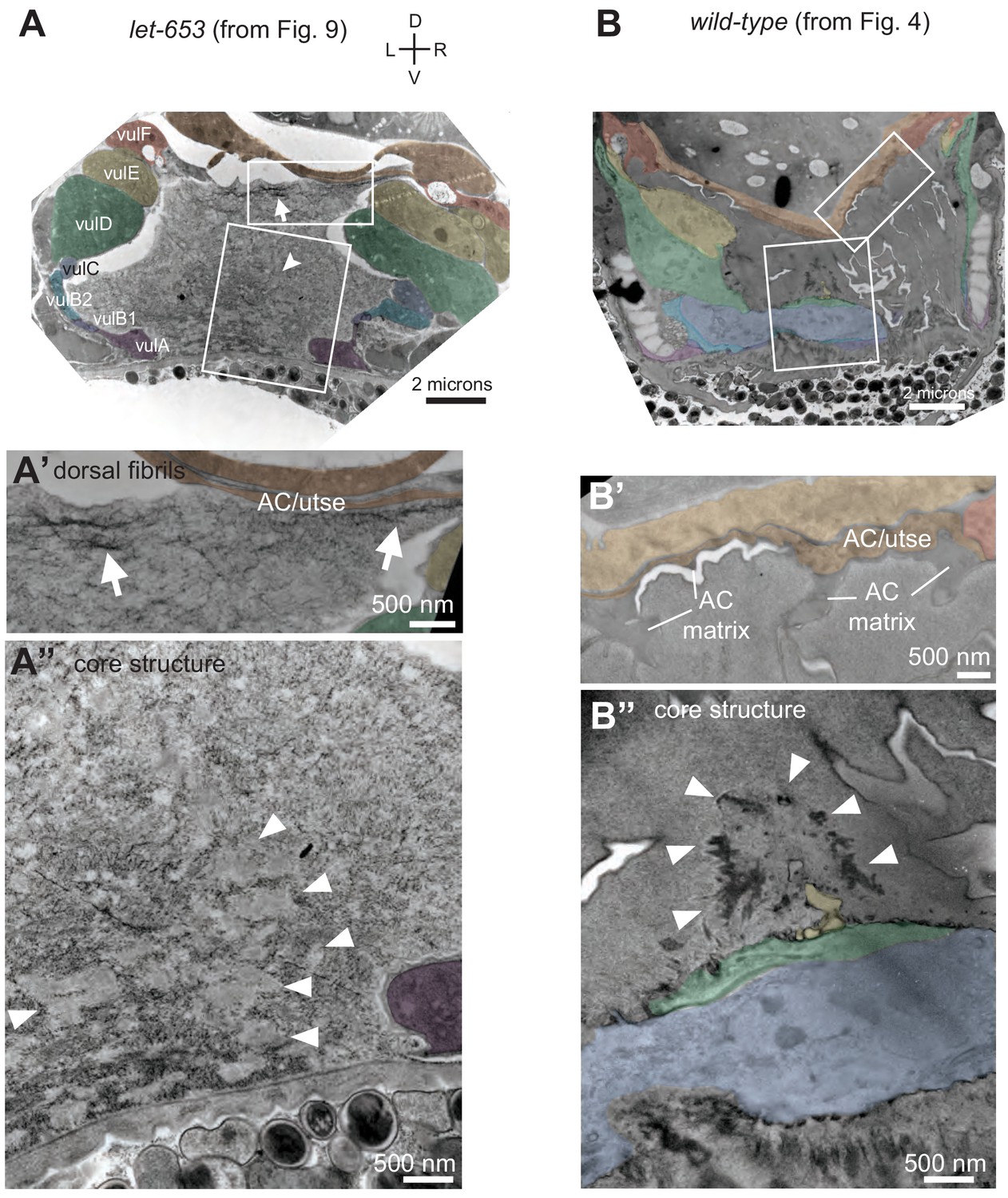
Vulva aECM organization differs between let-653 mutants and WT.
(A) let-653(cs178) mid-L4 TEM specimen from Figure 9C. Boxes indicate regions enlarged in A' and A’. (A') Fibrillar structures (arrows) are present in the dorsal part of the vulva lumen near the AC/utse. (A") The luminal core structure (arrowheads) is ill-defined and interspersed with various aggregates or short fibrils. (B) Wild-type N2 mid-L4 TEM specimen from Figure 4. Boxes indicate regions enlarged in B' and B’. (B') No fibrils are detectable in the dorsal part of the vulva lumen. (B") The luminal core structure is clearly defined and surrounded by a granular matrix.
Figure 10
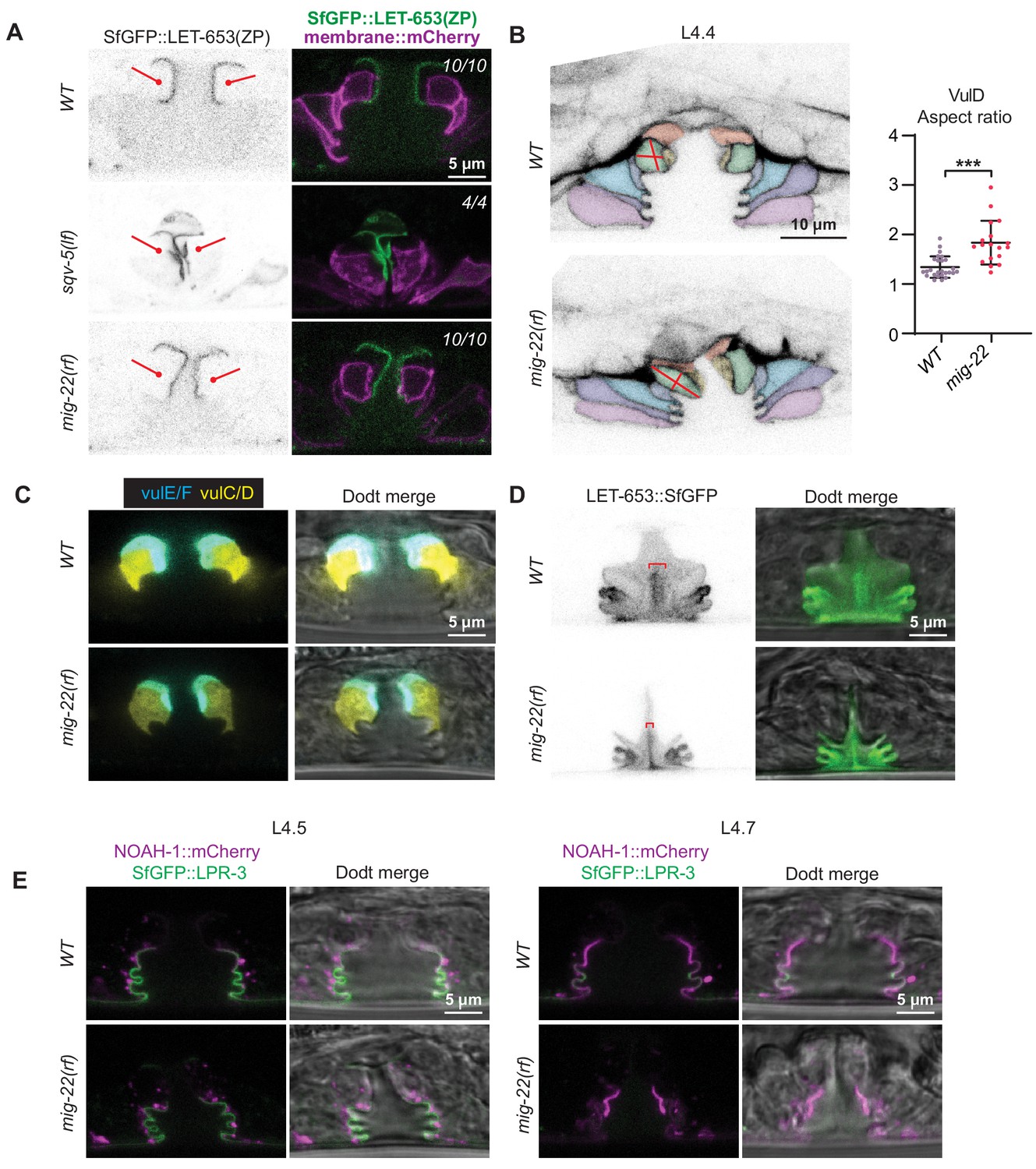
Vulva aECM structures form independently of chondroitin.
(A) Chondroitin mutants showed normal recruitment of LET-653(ZP)::SfGFP (csIs66) to the membrane-proximal matrix over 1° cells. Alleles used were sqv-5(n3611) (n = 4) and mig-22(k141rf) (n = 10). (B) WT vs. mig-22(k141rf) mutants (L4.4 stage) with membrane marker MIG-2::GFP (muIs28). The vulva neck region was taller and narrower in mutants compared to wild- type, while the rest of the lumen was shorter and narrower (n = 15, see quantification in Figure 11B). vulD cells (green) showed the most dramatic shape changes, with an increased aspect ratio (longest axis/shortest axis, p<0.001, Mann Whitney two tailed U test). WT, n = 26. mig-22, n = 18. (C) WT vs. mig-2(k141rf) mutants (L4.5 stage) with vulE/F marker daf-6pro::CFP and vulC/D marker egl-17pro::YFP (n = 3). (D) mig-22(k141rf) mutants assembled a well-organized, but narrow, core structure, as seen with LET-653::SfGFP (cs262) (n = 8). (E) mig-2(k141rf) mutants showed normal localization of LPR-3::SfGFP and NOAH-1::mCherry to apical surfaces (n = 10).
-
Figure 10—source data 1
Shape description of WT and mig-22 vulD cells.
The vulD cells of WT or mig-22 L4.4 stage vulvas were traced from a single confocal slice using signal from MIG-2::GFP. Shape descriptors, including aspect ratio, were calculated from these tracings. See Figure 10.
- https://cdn.elifesciences.org/articles/57874/elife-57874-fig10-data1-v2.xlsx
Figure 11
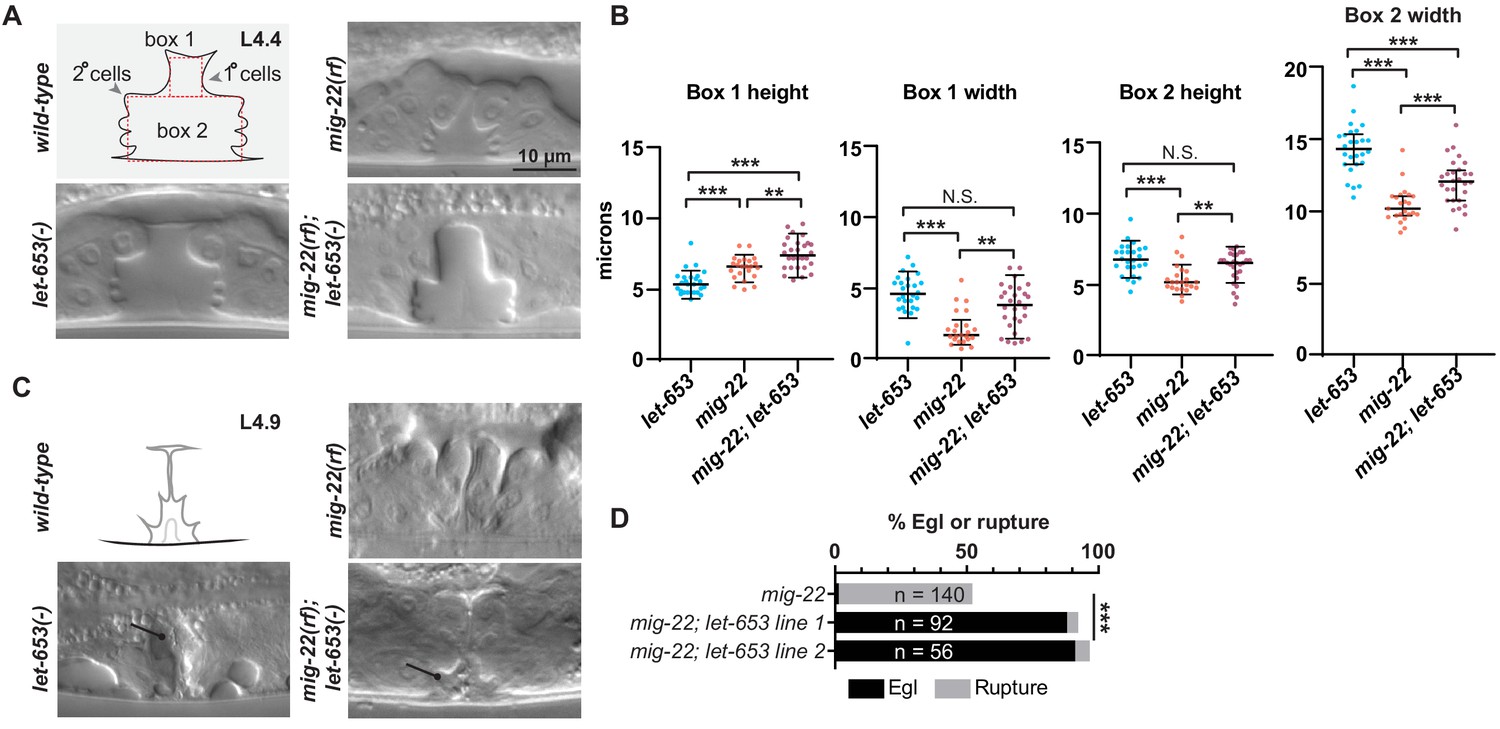
Chondroitin and LET-653 have both lumen-expanding and lumen-constraining roles.
(A) Loss of let-653 suppressed the mig-22(rf) Sqv phenotype and caused over-inflation of the dorsal lumen (n = 28). Alleles used were let-653(cs178) and mig-22(k141rf). (B) Lumen dimensions at the L4.4 stage were quantified as indicated in panel A. let-653 single mutant dimensions appeared similar to wild-type at this stage (n = 26; see Figure 8—figure supplement 1). p values derived from Mann–Whitney U test; **p<0.01, ***p<0.0001. Box 1 height, mig-22 vs. mig-22; let-653 p=0.002. Box 1 width, mig-22 vs. mig-22; let-653 p=0.001. Box 2 height, mig-22 vs. mig-22; let-653 p=0.003. (C) At late L4 stages, some let-653 single mutants (n = 3/16) and mig-22(rf); let-653 double mutants (n = 5/22) had disorganized material within the vulva lumen (line). (D) Two independently generated mig-22(rf); let-653 double mutant strains were analyzed, and both were severely egg-laying defective. ***p<0.0001, Fisher’s exact test.
-
Figure 11—source data 1
Measurements of WT and mutant L4.4 stage vulvas.
Measurements are taken from boxes drawn within DIC images of vulvas at L4.4 stage. See Figure 11 and Figure 8—figure supplement 1.
- https://cdn.elifesciences.org/articles/57874/elife-57874-fig11-data1-v2.xlsx
Figure 12

Model for aECM-dependent shaping of the vulva lumen during morphogenesis.
(A) The vulva lumen is initially expanded by chondroitin proteoglycans (pink arrows). Black; cuticle. Gray; apical membrane. (B) A membrane-proximal aECM appears alongside matrix fibrils and a central core to halt and/or stabilize vulva expansion. vulE/F aECM; red. vulC/D aECM; green. vulA/B aECM; blue. (C) The lumen narrows in the anterior-posterior axis. We propose that the central core and fibrils attach to the aECM and underlying membranes and pull ventrally, anteriorly, and posteriorly to shape the vulva lumen. The aECM changes over time; transient components turn over and the membrane-proximal matrix develops cuticle-like features. (D) By adulthood, the lumen narrows into a slit and is lined by cuticle.
Tables
Table 1
Strains used in this work.
| Strain | Genotype |
|---|---|
| ARF335 | fbn-1(tm290) III; aaaEx78 [fbn-1pro::FBN-1::mCherry; ttx-3pro::GFP] (Katz et al., 2018) |
| ARF379 | aaaIs12 [fbn-1pro::FBN-1::mCherry; ttx-3pro::GFP] V (Katz et al., 2018) |
| ARF359 | upIs1 [MUP-4::GFP; rol-6(su1006)] V; aaaEx78 [fbn-1pro::FBN-1::mCherry; ttx-3pro::GFP] (Hong et al., 2001; Katz et al., 2018) |
| HM24 | upIs1 V; aaaIs12 V (Hong et al., 2001; Katz et al., 2018) |
| GOU2043 | vab-10(cas602 [vab-10a::gfp]) I (Yang et al., 2017b) |
| JU486 | mfIs4 [egl-17pro::YFP; daf-6pro::CFP; unc-119(+)] (Mok et al., 2015) |
| ML2482 | noah-1(mc68 [NOAH-1::mCH(int)]) I (Vuong-Brender et al., 2017) |
| ML2547 | sym-1(mc85 [SYM-1::GFP]) X (Vuong-Brender et al., 2017) |
| ML2615 | dlg-1(mc103 [DLG-1::GFP]) X (Vuong-Brender et al., 2017) |
| N2 | WT |
| NF68 | mig-22(k141) III (Suzuki et al., 2006) |
| SP2163 | sym-1(mn601) X (Niwa et al., 2009) |
| UP2386 | csEx358 [lpr-1pro::LET-653b; unc-119pro::GFP] (Gill et al., 2016) |
| UP3244 | let-653(cs178) IV; csEx358 (Gill et al., 2016) |
| UP3342 | let-653(cs178) IV; csEx766 [lin-48pro::LET-653b::SfGFP; myo-2pro::GFP] (Forman-Rubinsky et al., 2017) |
| UP3349 | aaaIs12 V; csIs64 [let-653pro::LET-653b::SfGFP; rol-6(su1006)] |
| UP3353 | let-653(cs178) IV; aaaIs12 V; csEx766 |
| UP3422 | csIs66 [let-653pro::LET-653(ZP)::SfGFP; let-653pro::PH::mCherry] X |
| UP3444 | sqv-5(n3611)/hT2 [bli-4(e937) let-?(q782) qIs48] I,III; csIs66 X |
| UP3462 | let-653(cs178) IV; csIs66 X |
| UP3666 | lpr-3(cs250 [ssSfGFP::LPR-3]) X |
| UP3693 | noah-1(mc68 [NOAH-1::mCH(int)]) I; lpr-3(cs250 [ssSfGFP::LPR-3]) X |
| UP3746 | let-653(cs262 [LET-653::SfGFP]) IV |
| UP3756 | let-4(cs265 [ssmCherry::LET-4]) X |
| UP3757 | dpy-19(e1259) lin-12(n137)/hT2 [bli-4(e937) let-?(q782) qIs48] I,III; csIs66 X |
| UP3758 | unc-32(e189) lin-12(n137 n720)/hT2 [bli-4(e937) let-?(q782) qIs48] I,III; csIs66 X |
| UP3788 | noah-1(mc68 [NOAH-1::mCH(int)]) I; let-653(cs262 [LET-653::SfGFP]) IV |
| UP3856 | let-653(cs262 [LET-653::SfGFP]) IV; let-4(cs265 [ssmCherry::LET-4]) X |
| UP3861 | muIs27 [MIG-2::GFP; dpy-20+]; let-4(cs265 [ssmCherry::LET-4]) X (Honigberg and Kenyon, 2000) |
| UP3939 | let-4(mn105) X; csEx819 [grl-2pro::LET-4; myo-2pro::mRFP] (Forman-Rubinsky et al., 2017) |
| UP3967 | mig-22(k141) III; let-653(cs178) IV; csEx766 |
| UP3968 | mig-22(k141) III; let-653(cs178) IV; csEx766 |
| UP3970 | mig-22(k141) III; csIs66 X |
| UP3979 | let-653(cs178) IV; dlg-1(mc103 [DLG-1::GFP]) X; csEx766 |
| UP3995 | muIs28 [MIG-2::GFP; unc-31+] (Honigberg and Kenyon, 2000) |
| UP3966 | noah-1(mc68 [NOAH-1::mCH(int)]) I; let-653(cs178) IV; lpr-3(cs250 [ssSfGFP::LPR-3]) X; csEx766 |
| UP4004 | mig-22(k141) III; muIs28 |
| UP4005 | let-653(cs178) IV; muIs28; csEx766 |
| UP4014 | noah-1(mc68 [NOAH-1::mCH(int)]) I; mig-22(k141) III; lpr-3(cs250 [ssSfGFP::LPR-3]) X |
| UP4025 | noah-1(mc68 [NOAH-1::mCH(int)])/hT2 [bli-4(e937) let-?(q782) qIs48] I; unc-32(e189) lin-12(n137 n720)/hT2 [bli-4(e937) let-?(q782) qIs48] III |
| UP4027 | mig-22(k141) III; let-653(cs262 [LET-653::SfGFP]) IV |
| UP4038 | unc-32(e189) lin-12(n137 n720)/hT2 [bli-4(e937) let-?(q782) qIs48] I,III; lpr-3(cs250 [ssSfGFP::LPR-3]) X |
| UP4039 | mig-22(k141) III; mfIs4 |
| UP4040 | let-653(cs178) IV; mfIs4; csEx766 |
| UP4042 | unc-32(e189) lin-12(n137 n720)/hT2 [bli-4(e937) let-?(q782) qIs48] I,III; let-653(cs262 [LET-653::SfGFP]) IV |
| UP4043 | dpy-19(e1259) lin-12(n137)/hT2 [bli-4(e937) let-?(q782) qIs48] I,III; lpr-3(cs250 [ssSfGFP::LPR-3]) X |
| UP4044 | dpy-19(e1259) lin-12(n137)/hT2 [bli-4(e937) let-?(q782) qIs48] I,III; let-653(cs262 [LET-653::SfGFP]) IV |
| UP4045 | noah-1(mc68 [NOAH-1::mCH(int)]) I; dpy-19(e1259) lin-12(n137) III |
| UP4047 | noah-1(mc68 [NOAH-1::mCH(int)]) I; muIs28 |
Appendix 1—key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Gene (Caenorhabditis elegans) | fbn-1 III | Wormbase WS277 | fbn-1; ZK783.1 | FiBrilliN homolog |
| Gene (C. elegans) | let-653 IV | Wormbase WS277 | let-653; C29E6.1 | LEThal; ZP domain |
| Gene (C. elegans) | let-4 X | Wormbase WS277 | let-4; C44H4.2 | LEThal; eLRRon |
| Gene (C. elegans) | lin-12 III | Wormbase WS277 | lin-12; R107.8 | abnormal cell LINeage; Notch-related |
| Gene (C. elegans) | mig-22 III | Wormbase WS277 | mig-22; PAR2.4 | abnormal cell MIGration; chondroitin polymerizing factor |
| Gene (C. elegans) | mup-4 III | Wormbase WS277 | mup-4; K07D8.1 | MUscle Positioning; matrilin-related |
| Gene (C. elegans) | noah-1 I | Wormbase WS277 | noah-1; C34G6.6 | NOmpA homolog; ZP domain |
| Gene (C. elegans) | sqv-5 I | Wormbase WS277 | sqv-5; T24D1.1 | SQuashed Vulva; chondroitin sulfate synthase |
| Gene (C. elegans) | sym-1 X | Wormbase WS277 | sym-1; C44H4.3 | SYnthetic lethal with Mec; eLRRon |
| Gene (C. elegans) | vab-10 I | Wormbase WS277 | vab-10; ZK1151.1 | VAriaBle abnormal morphology; dystonin ortholog |
| Genetic reagent (C. elegans) | fbn-1 (tm290) III | Kelley et al., 2015 | strain ARF335 | 604 bp deletion/frameshift; presumed null |
| Genetic reagent (C. elegans) | let-4 (mn105) X | Mancuso et al., 2012 | strain UP3939 | Q305ochre; presumed null |
| Genetic reagent (C. elegans) | let-4(cs265 [mCherry:: LET-4]) X | This study | strain UP3756 | mCherry fused to endogenous LET-4 near its N-terminus by Cas9-triggered homologous recombination |
| Genetic reagent (C. elegans) | let-653 (cs178) IV | Gill et al., 2016 | strain UP3244 | C54ochre; presumed null |
| Genetic reagent (C. elegans) | let-653(cs262 [LET-653:: SfGFP]) IV | This study | strain UP3746 | SfGFP fused to endogenous LET-653 at its C-terminus by Cas9-triggered homologous recombination |
| Genetic reagent (C. elegans) | lin-12 (n137n720) III | (Sternberg and Horvitz, 1989). Caenorhabditis Genetics Center (CGC). | strain MT2343 | presumed null |
| Genetic reagent (C. elegans) | lin-12 (n137) III | (Greenwald et al., 1983). CGC. | strain MT2343 | S872F; hypermorph |
| Genetic reagent (C. elegans) | lpr-3(cs250 [SfGFP:: LPR-3]) X | This study | strain UP3666 | SfGFP fused to endogenous LPR-3 near its N-terminus by Cas9-triggered homologous recombination |
| Genetic reagent (C. elegans) | mig-22(k141) III | (Suzuki et al., 2006). CGC. | strain NF68 | G227E; hypomorph |
| Genetic reagent (C. elegans) | noah-1(mc68 [NOAH-1:: mCherry]) I | (Vuong-Brender et al., 2017). Michel Labouesse lab. | strain ML2482 | mCherry fused to endogenous NOAH-1 at internal site by Cas9-triggered homologous recombination |
| Genetic reagent (C. elegans) | sqv-5(n3611) I | (Hwang et al., 2003b). CGC. | strain MT10996 | deletion; presumed null |
| Genetic reagent (C. elegans) | sym-1 (mn601) X | (Davies et al., 1999). CGC. | strain SP2163 | Q275ochre; presumed null |
| Genetic reagent (C. elegans) | sym-1(mc85 [SYM-1:: GFP]) X | (Vuong-Brender et al., 2017). Michel Labouesse lab. | strain ML2547 | GFP fused to endogenous SYM-1 at its C-terminus by Cas9-triggered homologous recombination |
| Genetic reagent (C. elegans) | vab-10 (cas602 [VAB-10a:: GFP]) | (Yang et al., 2017b). CGC. | strain GOU2043 | GFP fused to endogenous VAB-10a at its C-terminus by Cas9-triggered homologous recombination |
| Genetic reagent - Transgene (C. elegans) | aaaIs12 [fbn-1pro::FBN-1::mCherry; ttx-3pro::GFP] | This study | aaaIs12, strain ARF379 | Transgene expressing full-length FBN-1 tagged internally with mCherry inserted just prior to the ZP domain |
| Genetic reagent - Transgene (C. elegans) | csEx766 [lin-48pro:: LET-653:: SfGFP; myo-2pro:: GFP] | Forman-Rubinsky et al., 2017 | csEx766 strain UP3342 | Duct-specific rescue transgene |
| Genetic reagent - Transgene (C. elegans) | csEx819 [grl-2pro:: LET-4; myo-2p:: mRFP] | Forman-Rubinsky et al., 2017 | csEx819 strain UP3939 | Duct/pore-specific rescue transgene |
| Genetic reagent - Transgene (C. elegans) | csIs64 [let-653pro:: LET-653:: SfGFP; lin-48pro:: mRFP] | Gill et al., 2016 | csIs64 strain UP3070 | Transgene expressing SfGFP-tagged LET-653 |
| Genetic reagent - Transgene (C. elegans) | csIs66 [let-653pro:: SfGFP:: LET-653(ZP); let-653pro:: PH::mCherry; lin-48pro:: mRFP] | Cohen et al., 2019 | csIs66 strain UP3422 | Transgene expressing SfGFP-tagged LET-653(ZP) domain |
| Genetic reagent - Transgene (C. elegans) | mfIs4[egl-17pro::YFP; daf-6pro::CFP; unc-119+] | (Félix, 2007). CGC. | mfIs4 strain JU486 | Transgene expressing CFP in primary vulva descendants and YFP in secondary vulva descendants. |
| Genetic reagent - Transgene (C. elegans) | muIs28 [mig-2pro:: MIG-2::GFP; unc-31+] | (Honigberg and Kenyon, 2000). CGC. | muIs28 strain CF693 | Transgene expressing GFP-tagged MIG-2 (membrane marker) |
| Genetic reagent - Transgene (C. elegans) | upIs1 [mup-4::GFP + rol-6(su1006)] | (Hong et al., 2001). CGC. | upIs1; strain EE86 | Transgene expressing GFP-tagged MUP-4 |
| recombinant DNA reagent | pDD282 (plasmid) | Dickinson et al., 2013 | Addgene plasmid #66823 | GFP^SEC^3xFlag vector with ccdB markers for cloning homology arms |
| recombinant DNA reagent | pDD162 (plasmid) | Dickinson et al., 2013 | Addgene plasmid #47549 | eft-3p::Cas9 + empty sgRNA plasmid |
| recombinant DNA reagent | pJC39 | This study | pDD282 with GFP replaced by SfGFP | |
| recombinant DNA reagent | pJC49 | This study | let-653 homology repair template generated by PCR with OJC201+OJC202 and oJC203+oJC204 and cloned into pJC39 | |
| recombinant DNA reagent | pJC50 | This study | let-653 sgRNA sequence (5’-TTGAGATATTACGTTCGAAC-3’) cloned into pDD162 | |
| recombinant DNA reagent | pJC67 | This study | let-4 homology repair template generated by PCR with oJC269+oJC270 and oJC271+oJC282 and cloned into pMLS291 | |
| recombinant DNA reagent | pJC68 | This study | let-4 sgRNA sequence (5’- GACTCCAGGACAAGCATTTG −3’) cloned into pDD162 | |
| recombinant DNA reagent | pMLS291 (plasmid) | Schwartz and Jorgensen, 2016 | Addgene plasmid #73724 | SapTrap vector with mCherry |
| recombinant DNA reagent | pMLS328 (plasmid) | Schwartz and Jorgensen, 2016 | Addgene plasmid #73717 | eft-3p::2xNLS-Cre, unc-119+, for SEC excision |
| recombinant DNA reagent | pRFR60 | This study | lpr-3 sgRNA sequence (5’-TTTGGCTACGACGTTAGCTG −3’) cloned into pDD162 | |
| recombinant DNA reagent | pRFR70 | This study | lpr-3 homology repair template generated by PCR with oRFR69+oRFR90 and oRFR71+oRFR72 and cloned into pJC39 | |
| sequence-based reagent | oJC201 | This study | PCR primer | 5'_ACGTTGTAAAACGACGGCCAGTCGCCGGCA-CAAAAATCAGTCTATCATTCC_3' |
| sequence-based reagent | oJC202 | This study | PCR primer | 5'_TCCAGTGAAAAGTTCTTCTCCTTTGCTGAT-GATGTTTgCAGTTCGAACG_3' |
| sequence-based reagent | oJC203 | This study | PCR primer | 5'_CGTGATTACAAGGATGACGATGACAAGAGA-TGAAAATACACACAAAAAATG_3' |
| sequence-based reagent | oJC204 | This study | PCR primer | 5'_TCACACAGGAAACAGCTATGACCATGTTAT-GTCTGGTAGCTGCTTTGATGATGG_3' |
| sequence-based reagent | oJC269 | This study | PCR primer | 5'_gtgGCTCTTCgTGGgtttaaacacgtatttcacacatttttcag_3' |
| sequence-based reagent | oJC270 | This study | PCR primer | 5'_gtgGCTCTTCgCATTCCAGGACAAGCATTTGTCGAG_3' |
| sequence-based reagent | oJC271 | This study | PCR primer | 5'_gtgGCTCTTCgGGTGTCATTACTCAAGCGTGCTTC_3' |
| sequence-based reagent | oJC282 | This study | PCR primer | 5'_GTGGCTCTTCgTACGATGGCACTGCAGTCATATTG_3' |
| sequence-based reagent | oRFR69 | This study | PCR primer | 5’_ACGTTGTAAAACGACGGCCAGTCGCCGGCAcatataataaagcattttgtctg_3' |
| sequence-based reagent | oRFR90 | This study | PCR primer | 5’_TCCAGTGAAAAGTTCTTCTCCTTTGCTgATGCCTAGTGCgACAGCTAAC_3' |
| sequence-based reagent | oRFR71 | This study | PCR primer | 5'_CGTGATTACAAGGATGACGATGACAAGAGAGCTATTAGCGAAGCAGACGTACC_3' |
| sequence-based reagent | oRFR72 | This study | PCR primer | 5’_TCACACAGGAAACAGCTATGACCATGTTATCGGTAACGGTCTTGACTCCGGC_3' |
| software, algorithm | ImageJ | NIH | ||
| software, algorithm | Adobe Illustrator | Adobe | ||
| software, algorithm | Imaris | Bitplane | ||
| software, algorithm | Prism | Graphpad |
Additional files
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
A multi-layered and dynamic apical extracellular matrix shapes the vulva lumen in Caenorhabditis elegans
eLife 9:e57874.
https://doi.org/10.7554/eLife.57874




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}