A regulatory pathway that selectively up-regulates elongasome function in the absence of class A PBPs
- Department of Microbiology, Cornell University, United States
Figures
Figure 1 with 1 supplement
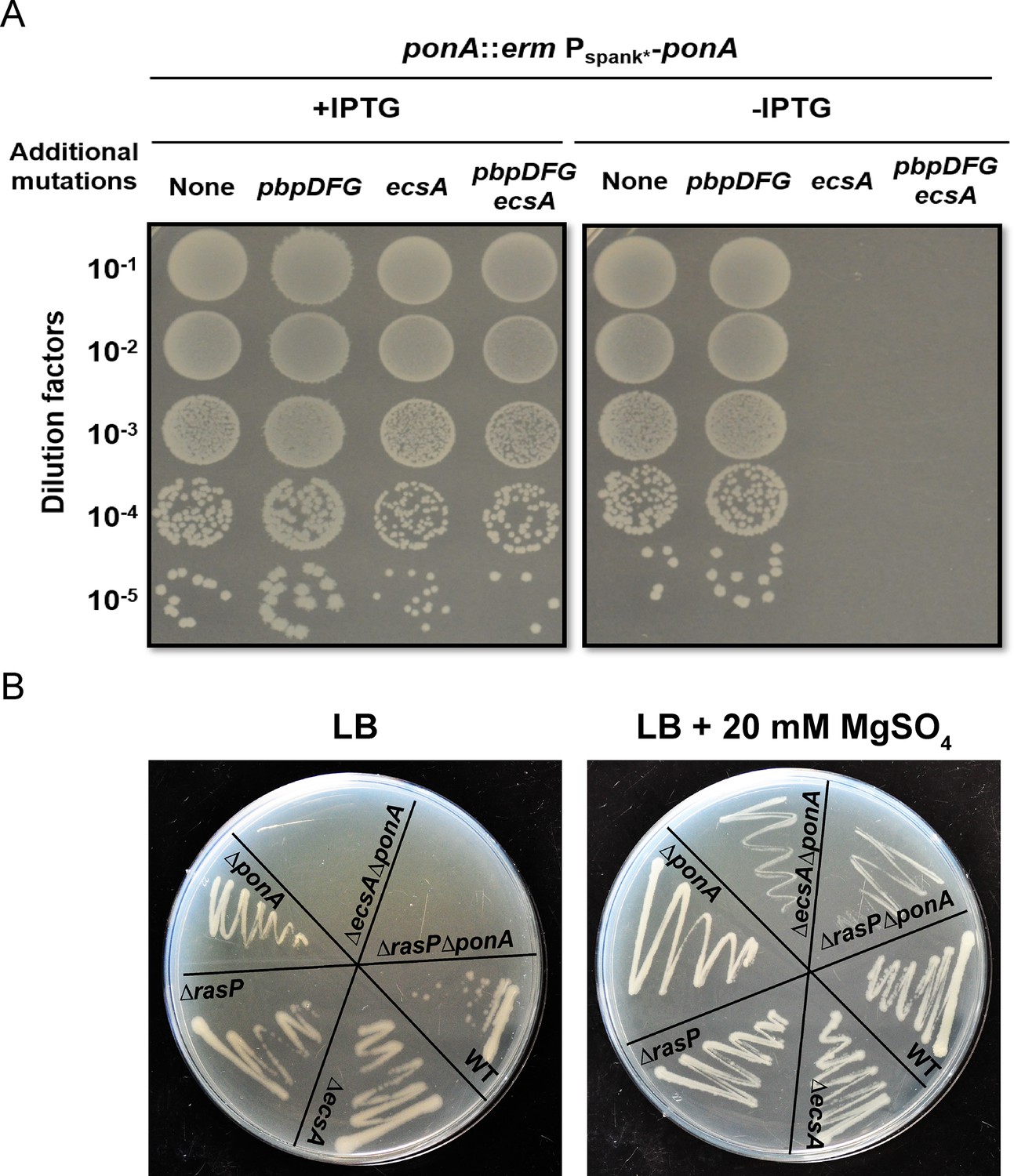
The ecsA and ponA genes are synthetic lethal in LB medium.
(A) Plating efficiency of ecsA deletion mutants. Right panel: spot dilutions were used to assess the effect of an ecsA null mutation on growth in a ponA depletion background (-IPTG) with and without additional mutations in pbpD, pbpF, pbpG (to mimic the Δ4 A PBP background). Left panel: ponA was induced (+IPTG) from the Pspank* promoter. (B) Growth of ΔecsA, ΔrasP, ΔponA and the double mutants ΔecsAΔponA and ΔrasPΔponA on LB agar plates with and without supplementation with 20 mM MgSO4.
Figure 1—figure supplement 1

Transposon insertion profile of the ecsAB operon.
Representation of TnSeq insertions in a WT and Δ4 aPBP background. Red bars indicate coverage of transposon insertions in the WT background and green bars indicate the same in Δ4 aPBP background. Shown here is a profile of ecsA and ecsB genes that lack insertions in the Δ4 aPBP strain. In contrast, the genes had insertions at multiple sites in WT strain.
Figure 2 with 2 supplements
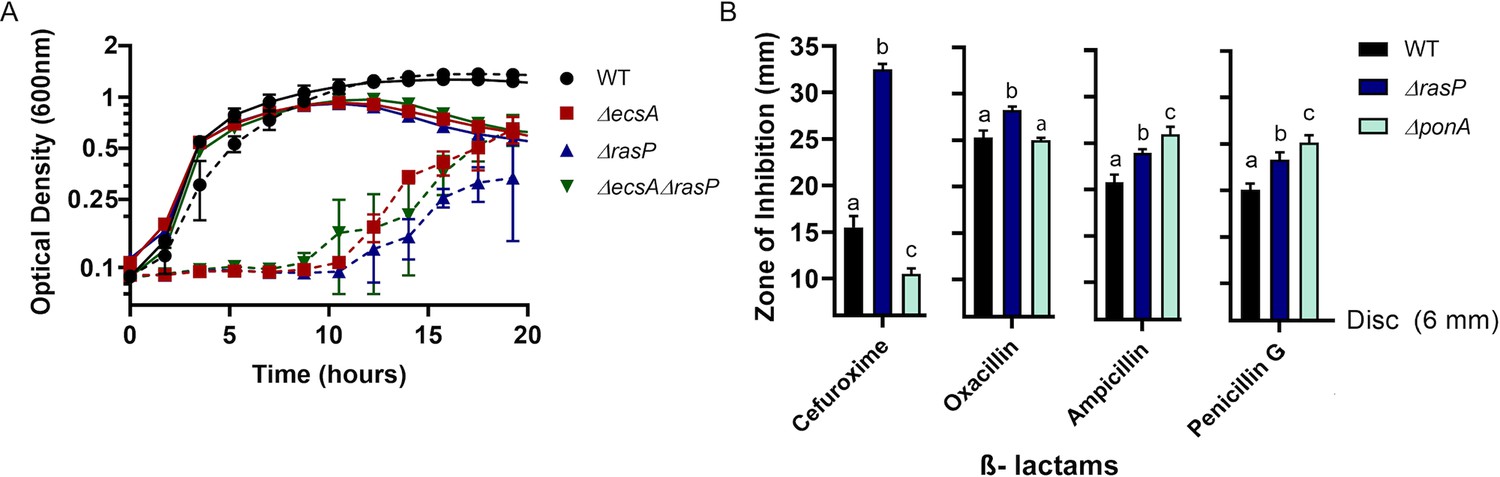
The EcsAB-RasP pathway is important for intrinsic antibiotic resistance.
(A) Growth kinetics of WT, ΔecsA, ΔrasP and the ΔecsAΔrasP double mutant in liquid LB medium with (dotted lines) and without (continuous lines) 0.4 µg/mL moenomycin (MOE). (B) β-lactam sensitivity of ΔrasP and ΔponA strains determined by disc diffusion assay using cefuroxime (CEF) (10 µg), oxacillin (3 µg), ampicillin (15 µg), and penicillin G (20 units). No comparison was done between antibiotic groups. P-value cutoff of <0.001 was used.
-
Figure 2—source data 1
Data of growth kinetics and zone of inhibition.
- https://cdn.elifesciences.org/articles/57902/elife-57902-fig2-data1-v1.xlsx
Figure 2—figure supplement 1

Antibiotic susceptibility of ΔecsA and ΔrasP mutants.
(A) Growth kinetics of WT, ΔecsA, ΔecsA-Pspac(hy)ecsA, ΔecsA Pspac(hy)ecsAecsB, ΔrasP and ΔrasP-Pspac(hy)rasP in LB medium supplemented with 1 µg/mL MOE and 0.25 mM IPTG for inducing the ectopic copies of ecsA/ecsB and rasP. (B) Disc diffusion assay for screening WT, ΔrasP and ΔponA strains for their sensitivity towards nisin and vancomycin; antibiotics which can affect the activity of both the aPBPs and the elongasome. No comparison was done between antibiotic groups. P-value cutoff of <0.0001 was used.
Figure 2—figure supplement 2
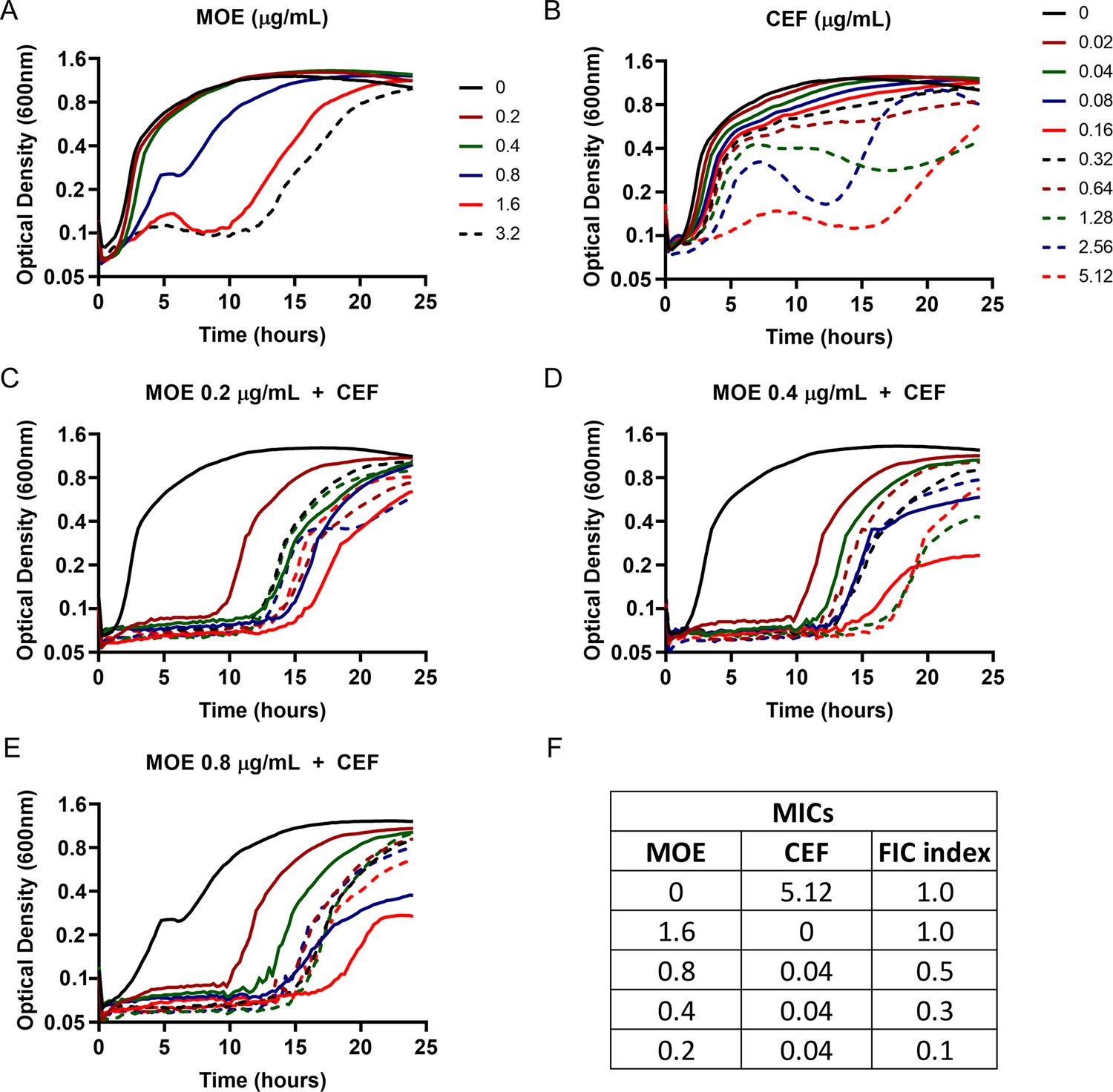
Synergistic interaction of MOE and CEF in B. subtilis.
Growth kinetics of WT treated with (A) MOE (0.2–3.2 µg/mL) (B) CEF (0.02–5.12 µg/mL) and (C–E) combination of MOE at 0.2 µg/mL, 0.4 µg/mL and 0.8 µg/mL with a range of CEF concentration (0.02–5.12 µg/mL) (F) A table for the Fractional Inhibitory Concentration (FIC) index for the combinatorial treatment of MOE and CEF. FIC index was calculated using the formula mentioned in Hall et al., 1983. A FIC index value of ≤0.5 is considered as a synergistic interaction (Odds, 2003). MIC of each drug individually or in combination was defined based on significant growth inhibition up to at least 10 hr of treatment.
Figure 3 with 2 supplements
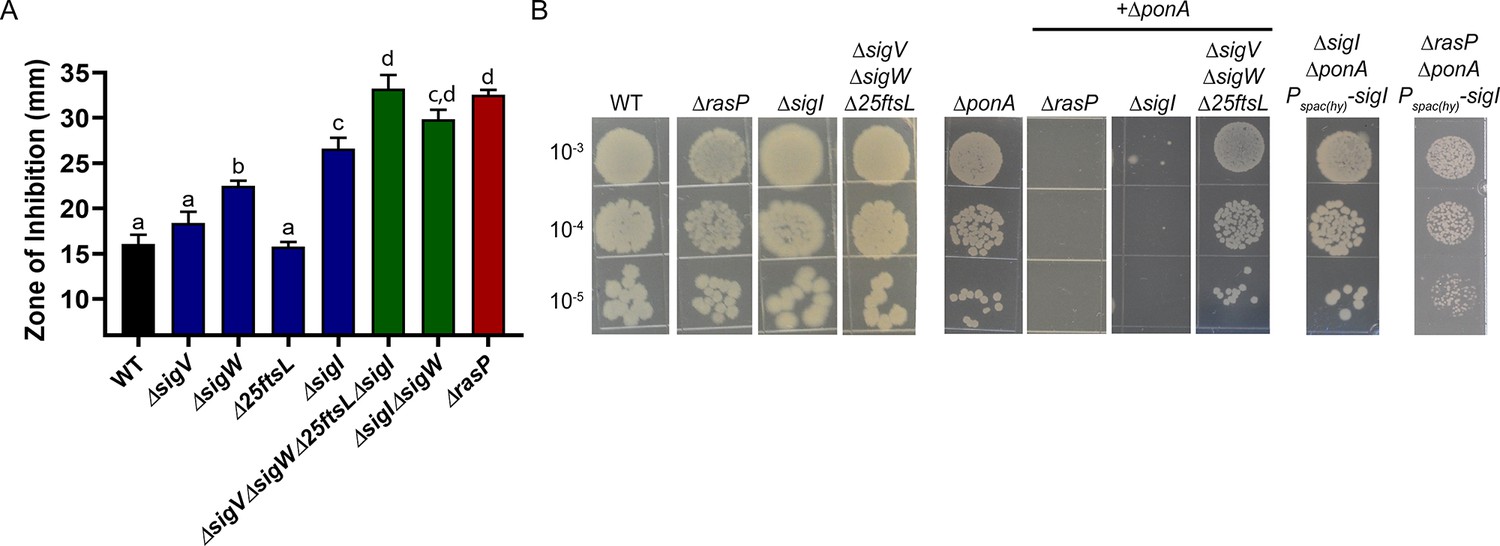
The EcsAB-RasP pathway functions largely through sigI.
(A) CEF (10 µg) sensitivity (disc diffusion assay) for WT, ΔrasP, ΔsigV, ΔsigW, Δ25ftsL, ΔsigI, ΔsigWΔsigI and ΔsigVΔsigWΔ25ftsLΔsigI strains. P-value cut-off of <0.0001 was used. (B) Plating efficiency of ΔrasP, ΔsigI and ΔsigVΔsigWΔ25ftsL strains in WT and ΔponA deletion background. This assay was done by plating 10 µL of mid-log phase cultures (grown in LB with 20 mM MgSO4) on LB agar plates (no Mg supplementation). The plating efficiency of ΔsigIΔponA double mutant was also evaluated after ectopic expression of sigI from the leaky promoter Pspac(hy).
-
Figure 3—source data 1
Data of zone of inhibition.
- https://cdn.elifesciences.org/articles/57902/elife-57902-fig3-data1-v1.xlsx
Figure 3—figure supplement 1
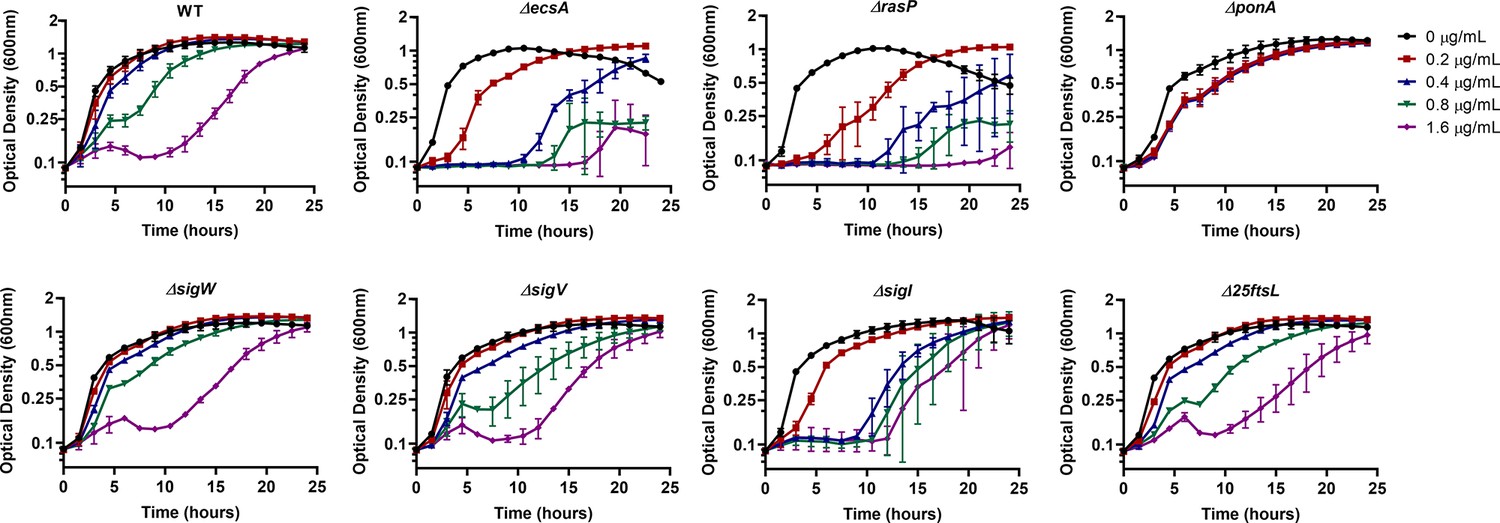
σI and RasP have similar MIC against MOE.
Growth kinetics of WT, ΔecsA, ΔrasP, ΔponA, ΔsigW, ΔsigV, ΔsigI, Δ25ftsL in the presence of 0, 0.2, 0.4, 0.8, 1.6 µg/mL MOE in LB medium. The concentration of the drug which inhibited growth up to at least 10 hr of treatment was considered as the MIC of the drug against the respective strain.
Figure 3—figure supplement 2
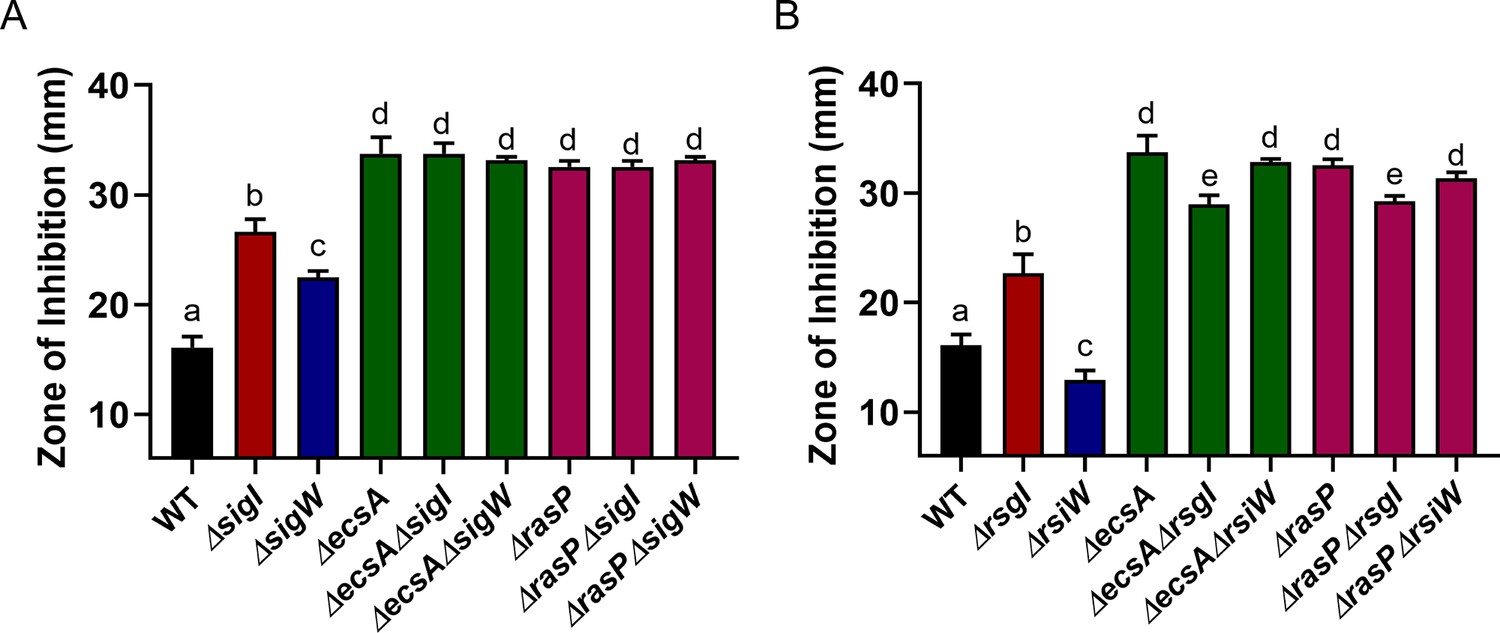
RasP functions primarily through σI to provide resistance against CEF.
Disc diffusion assay for CEF (10 µg) against (A) ΔecsA and ΔrasP in combination with the deletion mutants of ΔsigI and ΔsigW (B) ΔecsA and ΔrasP in combination with the deletion mutants of the anti-sigma factors ΔrsgI and ΔrsiW. P-value cut-off of 0.0001 was used.
Figure 4 with 1 supplement
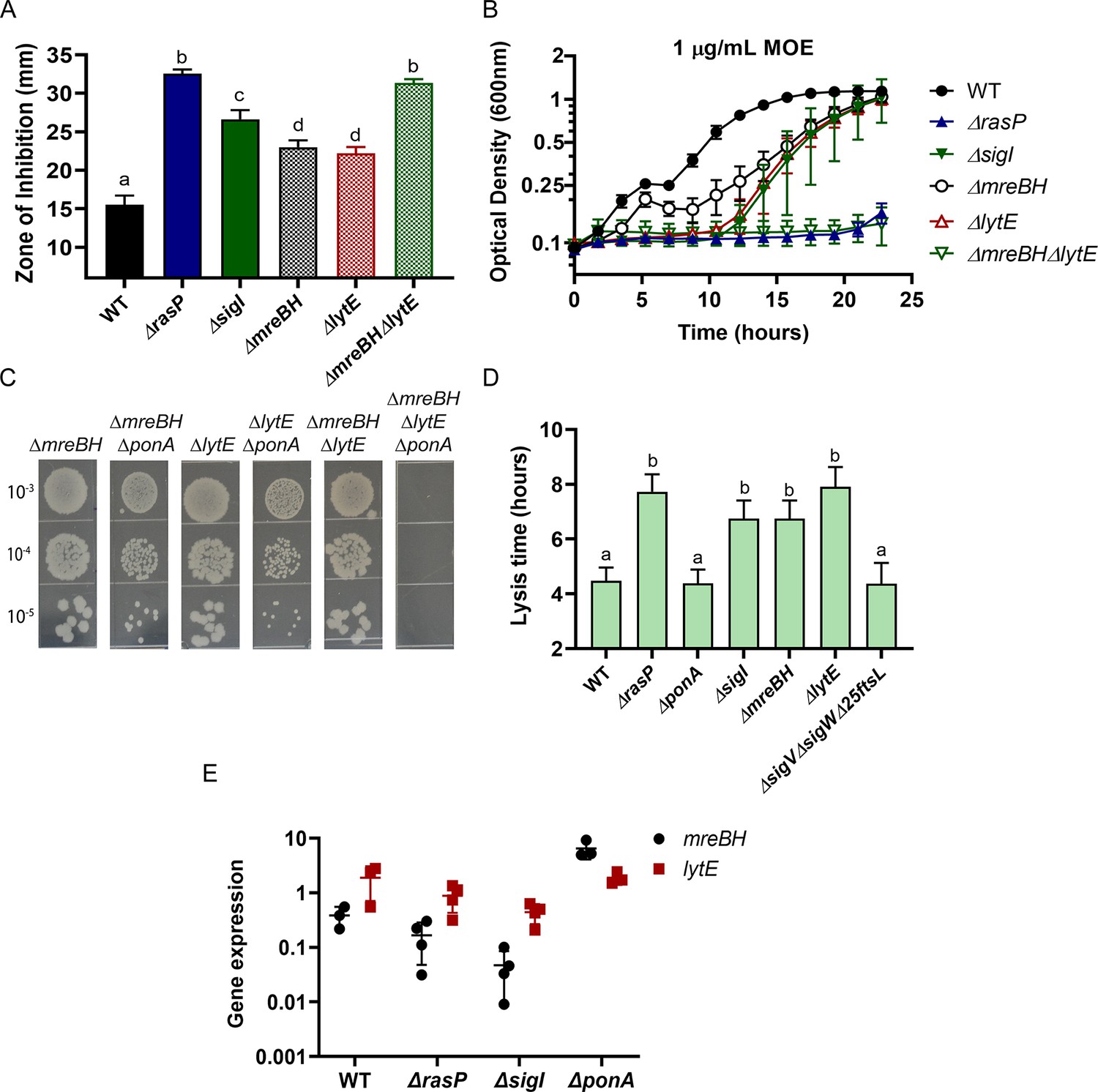
σI functions by increasing expression of mreBH and lytE.
(A) CEF (10 µg) sensitivity (disc diffusion assay) of ΔmreBH, ΔlytE and ΔmreBHΔlytE strains. Significance was determined with a P-value cut-off of <0.0001. (B) Growth kinetics of the mutants in LB medium with 1 µg/mL MOE. (C) Plating efficiency of the ΔmreBH, ΔlytE, and ΔmreBHΔlytE mutants alone and in combination with ΔponA. (D) The autolytic potential of the cells (WT, ΔponA, ΔrasP, ΔsigI, ΔmreBH, ΔlytE and ΔsigVΔsigWΔ25ftsL) measured by the time taken to reach 50% of initial cell density on treatment with sodium azide. P-value cut-off of <0.0001 was used. (E) Gene expression values (2-Δct) of mreBH and lytE normalized to gyrA plotted on log10 scale for WT, ΔrasP, ΔsigI and ΔponA strains.
-
Figure 4—source data 1
Data of zone of inhibition, MOE growth kinetics, lysis time and gene expression.
- https://cdn.elifesciences.org/articles/57902/elife-57902-fig4-data1-v1.xlsx
Figure 4—figure supplement 1
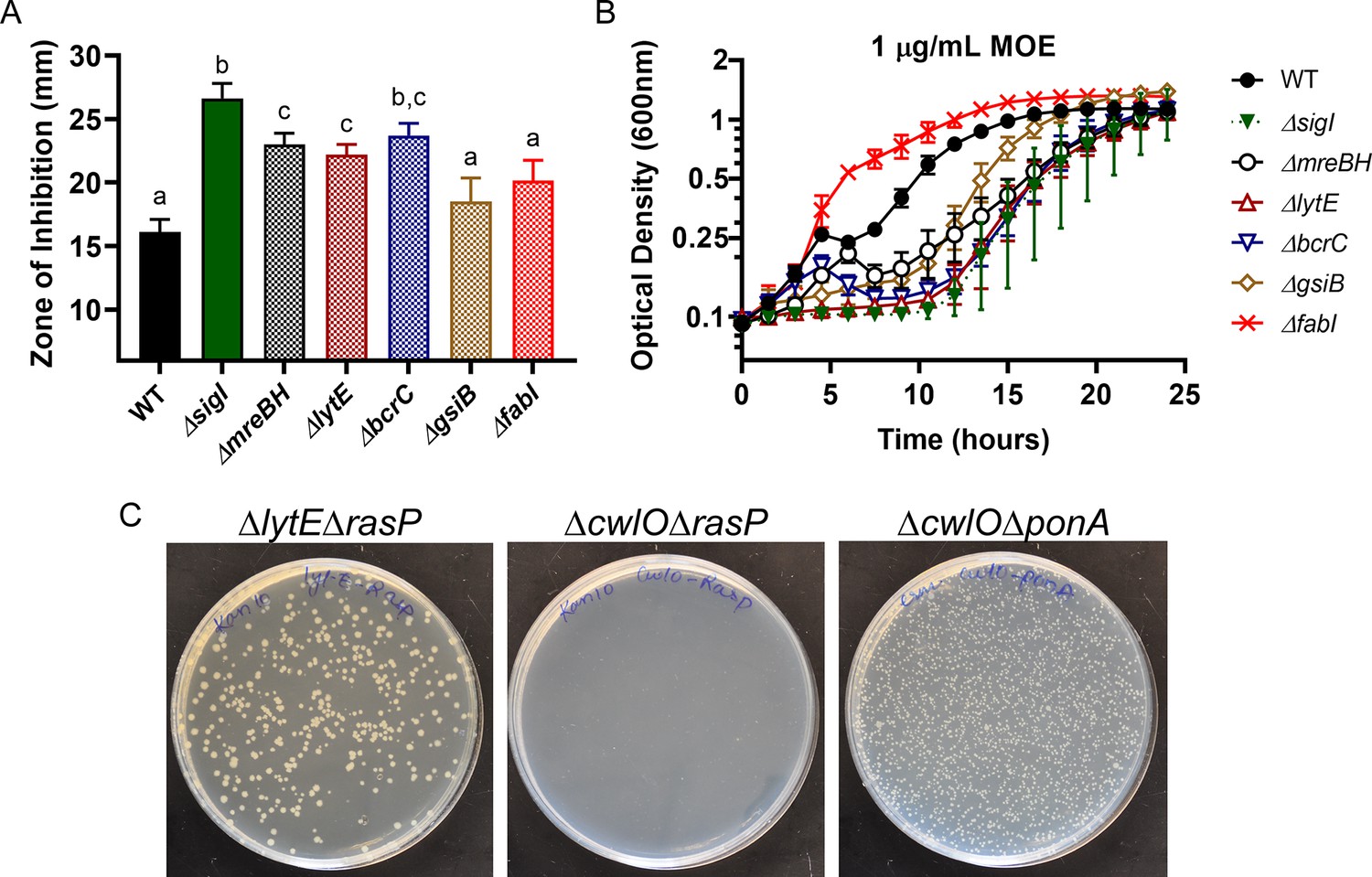
σI regulates the expression of mreBH and lytE to support elongasome function.
The importance of the σI regulon genes (mreBH, lytE, gsiB, fabI, bcrC) in the absence of ponA determined by the (A) Disc diffusion assay for CEF (10 µg). P-value cut-off of 0.0001 was used. (B) Growth kinetics in the presence of LB medium supplemented with 1 µg/mL MOE. (C) Transformation images of rasP::kan gDNA in ΔlytE and ΔcwlO background. ponA::erm transformation carried out as a control in ΔcwlO background. It validates that the transformation efficiency of the ΔcwlO strain was not compromised.
Figure 5

MreBH and LytE function cooperatively to increase elongasome function.
(A) CEF (10 µg) sensitivity (disc diffusion assay) of the ΔmreBH, ΔlytE, and ΔmreBHΔlytE strains with and complementation by ectopic expression of genes from the leaky promoter, Pspac(hy), or (for the ΔmreBHΔlytE strain) expression of mreBH from a xylose inducible promoter (Pxyl) and lytE from the Pspac(hy). P-value cut-off of <0.0001 was used. (B) CEF sensitivity (as for panel A) for ΔrasP and ΔsigI mutants with ectopic expression of mreBH from Pspac(hy) in the presence and absence of lytE. P-value cut-off of <0.0001 was used. Cell length (C) and width (D) of WT, ΔponA, ΔrasP, ΔsigI, ΔmreBHΔlytE, and ΔmreBH and ΔlytE strains was determined using at least 100 cells for each strain. P-value cut-off of <0.005 was used.
-
Figure 5—source data 1
Data of zone of inhibition and cell size measurements.
- https://cdn.elifesciences.org/articles/57902/elife-57902-fig5-data1-v1.xlsx
Figure 6
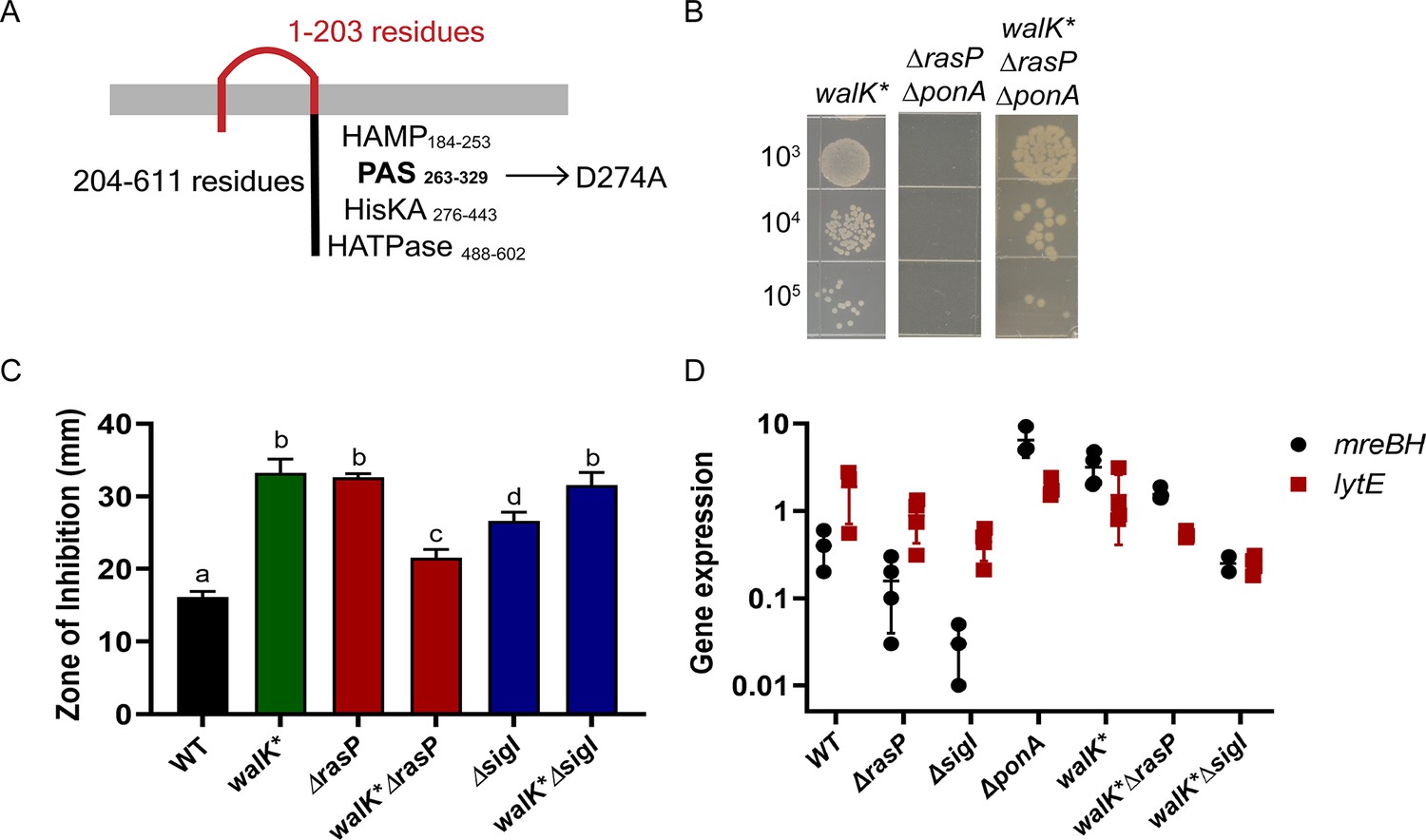
A walK* suppressor mutation elevates mreBH transcription.
(A) The D274 residue of WalK is part of a PAS-domain associated Zn-binding motif. (B) A walK* mutation rescues growth of the ΔrasPΔponA strain as monitored by a spot dilution assay. (C) CEF (10 µg) resistance (disc diffusion assay) of ΔrasP and ΔsigI and the respective double mutants of walK*ΔrasP and walK*sigI. A P-value cut-off of <0.0001 was used. (D) The effect of walK* on the expression profile of mreBH and lytE genes, alone and in combination with ΔrasP and ΔsigI. The gene expression values (2-Δct) were normalized with the house-keeping gene gyrA and then plotted on a log10 scale.
-
Figure 6—source data 1
Data of zone of inhibition and gene expression.
- https://cdn.elifesciences.org/articles/57902/elife-57902-fig6-data1-v1.xlsx
Figure 7
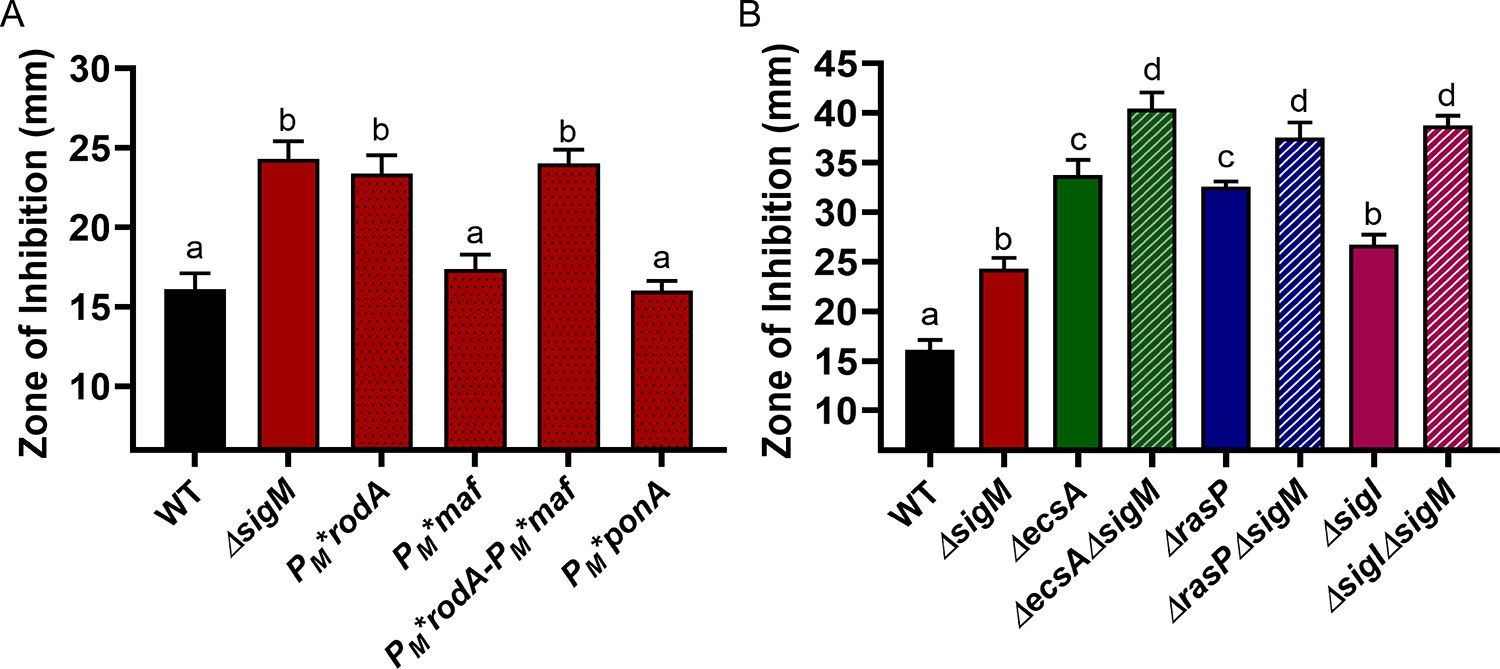
σM contributes additively with σI to CEF resistance by increasing expression of rodA.
CEF (10 µg) sensitivity (disc diffusion assay) for (A) WT, ΔsigM and promoter mutants of PM*rodA, PM*maf (which controls expression of mreBCD), PM*rodA-PM*maf and PM*ponA and (B) WT and ΔsigM mutants, alone and in combination with ΔecsA, ΔrasP and ΔsigI. P-value cut-off of <0.0001 was used for both the graphs.
-
Figure 7—source data 1
Data of zone of inhibition.
- https://cdn.elifesciences.org/articles/57902/elife-57902-fig7-data1-v1.xlsx
Figure 8
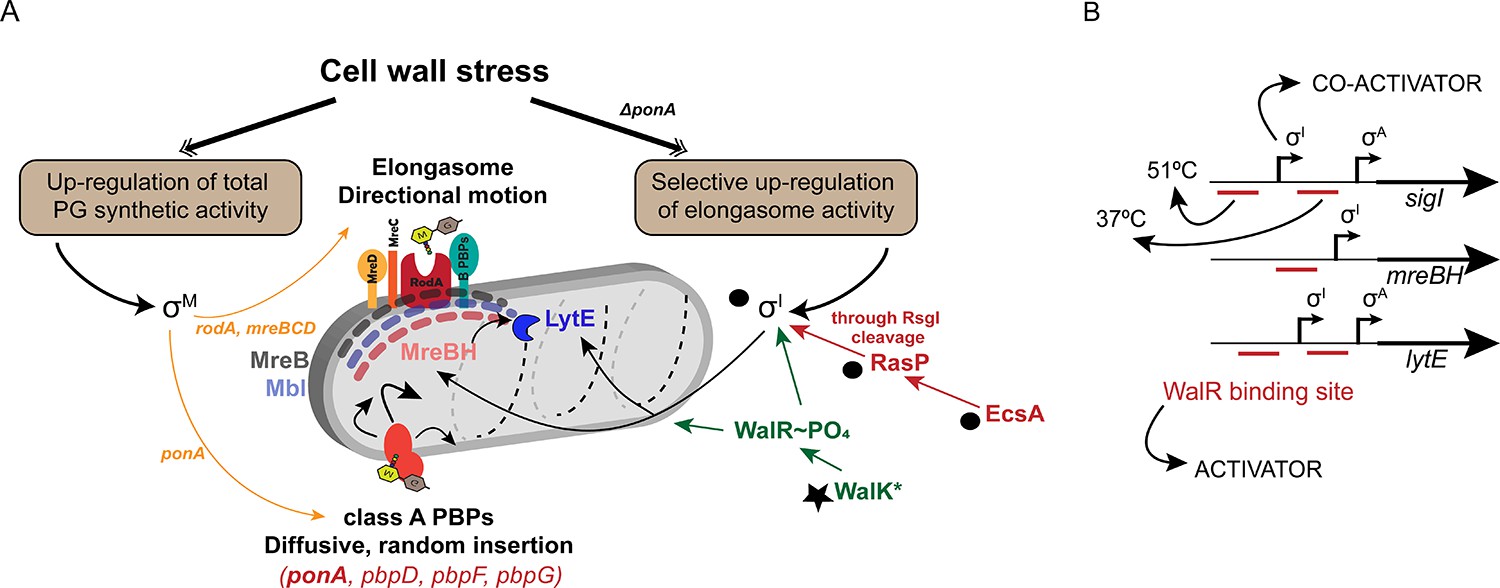
σI co-ordinates with WalKR to regulate elongasome function, and complements the σM dependent stress response.
(A) PG synthesis potential is dictated by the activity of the elongasome in collaboration with aPBPs. Cell wall stress activates σM (left), which up-regulates both pathways. In the absence of aPBPs, cells up-regulate elongasome activity through σI, which increases expression of genes (mreBH and lytE) important for elongasome function. Synthetic lethal relationships are shown here between deletion of ponA and genes in the σI pathway (black circles). Bypass of synthetic lethality can be compensated by a gain of function mutation in walK (star). (B) The promoter regions of sigI, mreBH and lytE are shown, depicting the binding sites of WalR and σI as annotated before (Huang et al., 2013). σI and WalR act as activators for the expression of sigI and lytE from the σA promoter. The downstream WalR binding site is important for expression of sigI and lytE at 37°C whereas the upstream binding site is crucial for the heat induction of these genes at 51°C.
Tables
Table 1
Minimum inhibitory concentration (MIC) of various strains for moenomycin in µg/mL.
| Strains | Moenomycin MIC (µg/mL) |
|---|---|
| WT | 1.6 |
| ΔecsA | 0.4 |
| ΔrasP | 0.4 |
| ΔponA | >1.6 |
| ΔsigW | 1.6 |
| ΔsigV | 1.6 |
| ΔsigI | 0.4 |
| Δ25ftsL | 1.6 |
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Strain, strain background (Bacillus subtilis, strain 168) | WT | Lab stock | B. subtilis 168 | (see Materials and methods) |
| Recombinant DNA reagent | This study | E. coli with pMarA1 | (see Materials and methods) | |
| Recombinant DNA reagent | HB20725 | This study | 168 pMarA1 | (see Materials and methods) |
| Recombinant DNA reagent | HB20738 | This study | pbpDFG null; ponA::erm;pMarA | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | Δ4 Class A PBP | This study | ponA::erm; pbpDFG::null | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ponA::erm Pspank*-ponA | This study | ycgO::Pspank*-ponA; ponA::erm | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | pbpDFG ponA::erm Pspank*-ponA | This study | pbpDFG::null; ycgO::Pspank*-ponA; ponA::erm | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ecsA ponA::erm Pspank*-ponA | This study | ecsA::null; ycgO::Pspank*-ponA; ponA::erm | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | pbpDFG ecsA-ponA::erm Pspank*-ponA | This study | ecsA::null;pbpDFG::null; ycgO::Pspank*-ponA; ponA::erm | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ytxG ponA::erm Pspank*-ponA | This study | ytxG::null; ycgO::Pspank*-ponA; ponA::erm | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | pbpDFG ytxG ponA::erm Pspank*-ponA | This study | ytxG::null;pbpDFG::null;ycgO::Pspank*-ponA; ponA::erm | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔecsA | This study | ecsA::kan | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔrasP | This study | rasP::kan | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔponA | This study | ponA::erm | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔecsAΔponA | This study | ecsA::null;ponA::erm | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔrasPΔponA | This study | rasP::null;ponA::erm | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔecsAΔrasP | This study | ecsA::null;rasP::erm | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔecsA Pspac(hy)-ecsA | This study | amyE::Pspac(hy)-ecsA; ecsA::erm | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔecsA Pspac(hy)-ecsAecsB | This study | amyE::Pspac(hy)-ecsAB; ecsA::erm | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔrasP Pspac(hy)-rasP | This study | amyE::Pspac(hy)-rasP; rasP::erm | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔsigW | This study | sigW::null | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔsigV | This study | sigV::null | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔsigI | This study | sigI::null | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | Δ25ftsL | This study | Made using CRISPR to remove the 2-26th AAs of FtsL so it is no longer a target of RasP | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔsigVΔsigW Δ25ftsLΔsigI | This study | sigV::null;sigW::null; Δ25ftsL;sigI::kan | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔsigIΔsigW | This study | sigI::null;sigW::kan | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔsigVΔsigW Δ25ftsL | This study | sigV::null;sigW::null; Δ25ftsL | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔsigIΔponA Pspac(hy)-sigI | This study | sigI::null; amyE::Pspac(hy)-sigI; ponA::erm | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔecsAΔsigI | This study | sigI::null;ecsA::kan | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔecsAΔsigW | This study | sigW::null;ecsA::kan | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔrasPΔsigI | This study | sigI::null;rasP::kan | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔrasPΔsigW | This study | sigW::null;rasP::kan | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔrsgI | This study | rsgI::null | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔrsiW | This study | rsiW::mls | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔecsAΔrsgI | This study | rsgI::null;ecsA::kan | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔecsAΔrsiW | This study | rsiW::mls;ecsA::kan | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔrasPΔrsgI | This study | rsgI::null;rasP::kan | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔrasPΔrsiW | This study | rsiW::mls;rasP::kan | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔsigM | This study | sigM::null | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔecsAΔsigM | This study | sigM::null;ecsA::kan | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔrasPΔsigM | This study | sigM::null;rasP::kan | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔsigIΔsigM | This study | sigM::null;sigI::kan | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | Pm*rodA | Zhao et al., 2019 | WT 168 transformed with CRISPR plasmid to remove Pm of rodA | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | Pm* maf | Zhao et al., 2019 | WT 168 transformed wth pMUTIN to introduce maf-Pm*(TGTT) | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | Pm*rodA Pm*murG | This study | Pm*murG transformed with CRISPR plasmid to remove Pm of ProdA | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | Pm*ponA | This study | WT168 transformed with CRISPR plasmid to remove Pm of ponA | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔmreBH | This study | mreBH::null | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔlytE | This study | lytE::null | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔgsiB | This study | gsiB::spec | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔfabI | This study | fabI::null | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔbcrC | This study | bcrC::null | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔmreBHΔlytE | This study | mreBH::null;lytE::null | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔmreBHΔponA | This study | mreBH::null;ponA::erm | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔlytEΔponA | This study | lytE::null;ponA::erm | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔmreBHΔlytE ΔponA | This study | mreBH::null;lytE::null; ponA::erm | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔmreBH Pspac(hy)-mreBH | This study | mreBH::null; amyE::Pspac(hy)-mreBH | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔlytE Pspac(hy)-lytE | This study | lytE::null; amyE::Pspac(hy)-lytE | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔmreBHΔlytE Pxyl-mreBH | This study | mreBH::null;lytE::null; lacA::Pxyl-mreBH | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔmreBHΔlytE Pxyl-mreBH Pspac(hy)-lytE | This study | lytE::null; amyE::Pspac(hy)-lytE; lacA::Pxyl-mreBH; mreBH::kan | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔrasPΔmreBH Pspac(hy)-mreBH | This study | mreBH::null; amyE::Pspac(hy)-mreBH; rasP::kan | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔrasPΔmreBH ΔlytE Pspac(hy)-mreBH | This study | mreBH::null;lytE::null; amyE::Pspac(hy)-mreBH; rasP::kan | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔsigIΔmreBH Pspac(hy)-mreBH | This study | mreBH::null; amyE::Pspac(hy)-mreBH; sigI::kan | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | ΔsigIΔmreBHΔlytE Pspac(hy)-mreBH | This study | mreBH::null;lytE::null; amyE::Pspac(hy)-mreBH; sigI::kan | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | walK* | This study | WalKD274A, constructed using CRISPR | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | walK*ΔrasP | This study | WalKD274A;rasP::kan | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | walK*ΔsigI | This study | WalKD274A;sigI::kan | (see Materials and methods) |
| Strain, strain background (Bacillus subtilis, strain 168) | walK*ΔrasPΔponA | This study | WalKD274A;rasP::kan; ponA::erm | (see Materials and methods) |
| Recombinant DNA reagent | pMarA | Le Breton et al., 2006 | a plasmid harboring the mariner-Himar1 transposase | |
| Recombinant DNA reagent | pMarA1 | Modified pMarA to introduce MmeI sites | ||
| Recombinant DNA reagent | pDR244 | BGSC (ECE274) | To remove the kan/erm cassette from BKE strains | |
| Recombinant DNA reagent | pAM012 | Meeske et al., 2015 | For Pspank*-ponA constructs | |
| Recombinant DNA reagent | pPL82 | For Pspac(hy) constructs at amyE locus | ||
| Recombinant DNA reagent | pBS2EXylRPxylA | BGSC (ECE741) | For Pxyl constructs at lacA locus |
Additional files
-
Supplementary file 1
List of primers used in this study.
- https://cdn.elifesciences.org/articles/57902/elife-57902-supp1-v1.docx
-
Transparent reporting form
- https://cdn.elifesciences.org/articles/57902/elife-57902-transrepform-v1.docx
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
A regulatory pathway that selectively up-regulates elongasome function in the absence of class A PBPs
eLife 9:e57902.
https://doi.org/10.7554/eLife.57902




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}