The Natural History of Model Organisms: Neurogenomic insights into the behavioral and vocal development of the zebra finch
- Department of Evolution, Ecology, and Behavior, School of Integrative Biology, University of Illinois at Urbana-Champaign, United States
- International Research Center for Neurointelligence, University of Tokyo, Japan
- Department of Biology, Texas A&M University, United States
- Department of Biological Sciences, Macquarie University, Australia
- Article
- Figures and data
- Abstract
- Introduction
- An evolutionary history of the zebra finch
- A model species for the analysis of sex differences in vocal learning and production?
- Differences in captive vs. wild zebra finches and comparisons with northern hemisphere songbirds
- Genes and brains for vocal learning
- The importance of studying female zebra finches
- Conclusions
- Data availability
- References
- Decision letter
- Author response
- Article and author information
- Metrics
Abstract
The zebra finch (Taeniopygia guttata) is a socially monogamous and colonial opportunistic breeder with pronounced sexual differences in singing and plumage coloration. Its natural history has led to it becoming a model species for research into sex differences in vocal communication, as well as behavioral, neural and genomic studies of imitative auditory learning. As scientists tap into the genetic and behavioral diversity of both wild and captive lineages, the zebra finch will continue to inform research into culture, learning, and social bonding, as well as adaptability to a changing climate.
Introduction
The zebra finch Taeniopygia guttata is the most intensively studied species of bird that is maintained in captivity in large numbers despite not being a species bred for its meat or eggs, like the chicken or the quail (reviewed in Zann, 1996). It became popular as a pet bird in the 19th century because it bred well in captivity, and was adopted for scientific study in the third quarter of the 20th century, initially for research into sexual behaviors (Morris, 1954; Immelmann, 1972). Later, the zebra finch was used in studies of the de novo evolution of vocal culture (e.g. Fehér et al., 2009; Diez and MacDougall-Shackleton, 2020), the neuroethology of imitative vocal learning (Terpstra et al., 2004; Vallentin et al., 2016; Yanagihara and Yazaki-Sugiyama, 2019), the neural mechanisms of sensorimotor learning (Mandelblat-Cerf et al., 2014; Okubo et al., 2015; Mackevicius et al., 2020; Sakata and Yazaki-Sugiyama, 2020), and the role of early acoustic experience on the song-based preferences of female mate choice (Riebel and Smallegange, 2003; Chen et al., 2017; Woolley, 2012; see the following video for a mating display in zebra finches: https://www.youtube.com/watch?v=TaC6D1cW1Hs).
Due to the pronounced sexual differences in singing and plumage coloration found in the zebra finch (Figure 1), earlier research quickly focused on when and how males learn to copy and produce a tutor(-like) song (e.g. Eales, 1987; Brainard and Doupe, 2002; Figure 2A), and then eventually on how females learn from their (foster) fathers to prefer particular male vocal displays (Braaten and Reynolds, 1999; Riebel, 2000). This allowed for the characterization and testing of the functions of male song and its female perception in the context of acoustic sexual dimorphism at the behavioral, endocrine, and neurophysiological levels (reviewed in Riebel, 2009; Hauber et al., 2010).
Figure 1

Adult zebra finches in the wild.
Four female and nine male adult zebra finches in the wild in Australia. As the species experiences increasingly extreme climatic fluctuations, future field studies of the zebra finch should also advance our understanding how opportunistically breeding species are able to adapt to accelerating climate change (photo credit: Simon C Griffith).
Figure 2
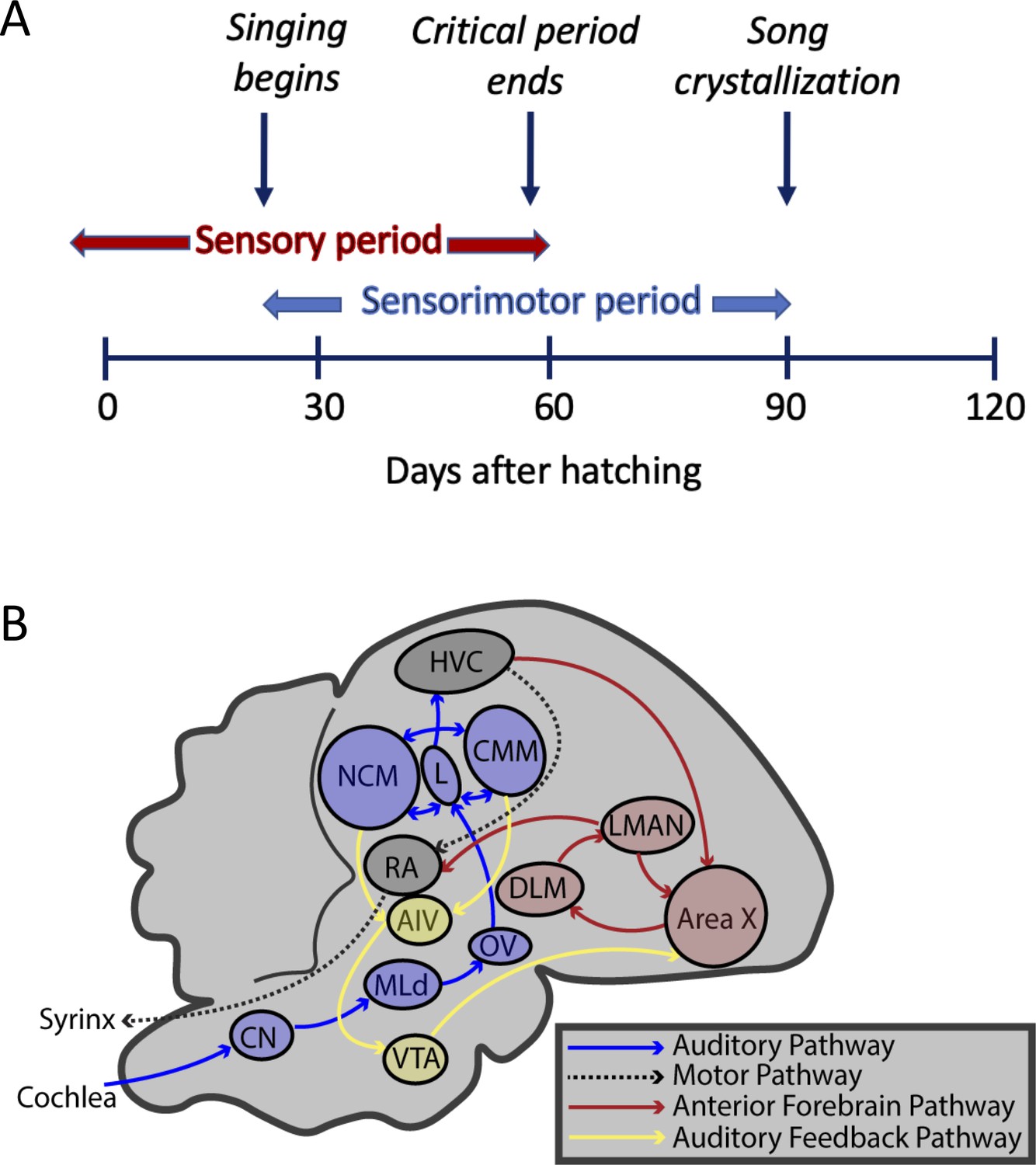
Timeline and brain pathways of auditory and vocal learning in the zebra finch.
(A) Timeline of sensory (auditory learning) and sensory-motor (vocal self-assessment and song-production) critical periods in zebra finch song development. (B) Brain nuclei of male zebra finches for auditory learning (CN: cochlear nucleus; MLd: mesencephalicus lateralis pars dorsalis; OV: nucleus ovoidalis; field L: primary auditory forebrain input area; NCM: caudomedial nidopallium; CMM: caudomedial mesopallium; VTA: ventral tegmental area; and AIV: ventral portion of the intermediate arcopallium), vocal learning (HVC, Area X: basal ganglia; LMAN: lateral magnocellular nucleus of the anterior nidopallium; DLM: nucleus dorsolateralis anterior thalami, pars medialis), and vocal production (HVC, and RA: robust nucleus of the arcopallium).
The zebra finch was the second avian species to have its genome sequenced (Warren et al., 2010), after the domestic fowl (Gallus gallus; International Chicken Genome Sequencing Consortium, 2004). Soon after the appearance of transgenic lines of domestic fowl and the Japanese quail Cortunix japonica (reviewed by Sato and Lansford, 2013), the first generations of transgenic zebra finches become available (e.g. Agate et al., 2009; Abe et al., 2015; Liu et al., 2015). The proven feasibility of genome editing in both developing zebra finches (e.g. Ahmadiantehrani and London, 2017) and adult poultry (reviewed in Woodcock et al., 2017), means that this bird may also be used as both a basic and an applied (i.e., biomedical) model for development and for human health and disease (e.g. Han and Park, 2018; London, 2020).
Studies of zebra finch natural history in Australia have been essential to establish and confirm the rationale for studying this species as a model for acoustic communication (Zann, 1990; Elie et al., 2010), social behavior (McCowan et al., 2015; Brandl et al., 2019a; Brandl et al., 2019b), reproductive physiology (Perfito et al., 2007), life-long pair bonding (Mariette and Griffith, 2012), and adaptations to heat (Cade et al., 1965; Cooper et al., 2020a; Cooper et al., 2020b). Specifically, by understanding the natural history of the zebra finch, research in captivity can capitalize on the manipulation of the behavioral, neuroendocrine, and epigenetic bases of the bird’s phenotype, including conspecific brood parasitism, parent-offspring conflict, and sibling rivalry.
Finally, with Australia experiencing increasingly extreme climatic events and fluctuations, field studies of the zebra finch are also paving the way to understanding how this opportunistically breeding species is adapting to accelerating climate change. For example, recent wild studies have revealed the zebra finch's extensive behavioral and physiological plasticity to withstand extreme temperatures of over 40°C (e.g. Cooper et al., 2020a; Cooper et al., 2020b; Funghi et al., 2019). In turn, studies of captive zebra finches in controlled temperature conditions have already tested the effects of cool vs. hot climates on parental investment (Nord et al., 2010), parent-offspring embryonic communication (Mariette and Buchanan, 2016), offspring development (Wada et al., 2015), tutor choice for song learning (Katsis et al., 2018), adult phenotype (e.g. body size: Andrew et al., 2017), the level of DNA methylation (Sheldon et al., 2020), and the effect of heat waves on sperm (Hurley et al., 2018).
By tapping into the existing genetic and behavioral diversity of wild and captive lineages in zebra finches (e.g. Forstmeier et al., 2007; Knief et al., 2015) to perform comparative avian genomic analyses (Jarvis et al., 2014; Feng et al., 2020), interspecific hybridization studies (Woolley and Sakata, 2019; Wang et al., 2019), and direct genetic manipulations (Liu et al., 2015; London, 2020), the zebra finch shall continue to serve as a focal subject of integrative research into human language-like vocal culture (Hyland Bruno et al., 2021), auditory learning (Theunissen et al., 2004), acoustically-mediated social bonding (Tokarev et al., 2017), and genetic (Balakrishnan et al., 2010) and behavioral (e.g. song) variability (Lansverk et al., 2019; see Box 1).
Box 1.
Outstanding questions in zebra finch research.
Female zebra finches do not sing but have a diverse repertoire of cooperative calls and other social behavioral displays. What is the neurogenomic and ontogenetic basis of this lack of singing in females?
Can gene editing become standard practice in both ontogenetic and adult-onset manipulations of the genomic architecture and gene activational basis of focal zebra finch traits, including imitative song learning and auditory feedback in the maintenance of crystallized song production?
What is the genomic and transcriptomic mechanism of hair-cell regeneration in the songbird inner ear and can it be transferred to human hearing loss treatments?
What is the genomic and physiological basis of aseasonal reproduction in nomadic zebra finches?
An evolutionary history of the zebra finch
The zebra finch is endemic to Australasia, and evolved there as part of the Australian grass finch radiation within the Estrildidae (Olsson and Alström, 2020). The species shares a common ancestor with Poephila finches (long-tailed P. acuticauda; black-throated P. cincta; and masked finch P. personata), diverging around 2.9 million years ago (Singhal et al., 2015). Formerly, the zebra finch was placed in a genus with the double-barred finch (Taeniopygia bichenovii), but in fact these two lineages diverged around 3.5 million years ago (Singhal et al., 2015).
Two subspecies of the zebra finch are recognized, with the continental Australian taxon (T. guttata castanotis) having no clear genetic structure and apparently mating randomly within its breeding population (Balakrishnan and Edwards, 2009). The other subspecies is the Timor zebra finch (T. g. guttata), found to the north of Australia. The genetic divergence between the two lineages suggests that the latter taxon colonized the Lesser Sunda Islands around 1 million years ago and has a reduced diversity and genetic distance driven by founding effects and selection, relative to the continental subspecies (Balakrishnan and Edwards, 2009). The insular subspecies has also been occasionally studied in captivity, and it differs from the continental Australian subspecies in morphological and behavioral traits, including song rate and mate choice (Clayton, 1990; Clayton et al., 1991).
The two subspecies of the zebra finch are physically isolated from one another in the wild, but they can readily hybridize and be back-crossed in captivity to examine a range of questions in classical genetics and functional developmental biology. To date, this approach has seen limited application, with just one study looking at the divergence in gene regulation between the two subspecies (Davidson and Balakrishnan, 2016). Whilst this direction could provide an extremely valuable new research opportunity, a major logistical challenge to overcome will be the capture and export of birds from Indonesia, or the continued maintenance of distinct (non-hybrid) domesticated populations of T. g. guttata in captivity.
A model species for the analysis of sex differences in vocal learning and production?
Zebra finches have a relatively short generation time for altricial birds (those that are underdeveloped at the time of hatching): they become sexually mature at between 90 and 100 days of age in captivity, at which point they are ready to form pair bonds, build nests, and breed (Zann, 1996). They are highly social and can be kept at great densities in shared housing with a relative absence of highly antagonistic behaviors. This is likely to be related to the level of sociality and the highly fluid flock-wide social relationships seen in the wild (McCowan et al., 2015; Brandl et al., 2019a), as individuals congregate around food and water, and nest in close proximity in loose colonies for apparent social benefits (Brandl et al., 2019b).
Provided with sufficient water, nesting sites, and nest materials, and one (or more) mate(s) of the opposite sex, zebra finches can successfully reproduce on a predominantly seed-based diet, simplifying husbandry, even during the nestling stage. Indeed, under a broad range of environmental and social conditions in captivity, when given the infrastructure (e.g. nesting platform or cavity and materials) to breed, most pairs will breed successfully within a short time frame (Griffith et al., 2017), and the life history can be followed across many generations in a relatively short period of time (e.g. Briga et al., 2019).
With a clutch size of between 2 and 9 eggs (mode: 5), and with brood reduction rates that can be less than 30%, each reproductive bout is typically rapid and productive. In the wild, zebra finches pair for life, and partners are found in close proximity during both the breeding and non-breeding periods (Mariette and Griffith, 2012; McCowan et al., 2015). In captivity, this strong pair bond is preceded by rapid pairing, with singletons forming pair bonds within days or weeks when introduced into a new cage or aviary (Rutstein et al., 2007; Campbell et al., 2009). The strength of the pair bond, the high levels of affiliative behaviors, and the relative absence of antagonism between partners also allow zebra finches to be kept in easily monitored single-pair cages, rather than in communal aviaries (Zann, 1996).
However, it was not just ease of breeding in captivity that turned the zebra finch into a popular model for studying the development of sexual dichromatism and vocal dimorphism. Rather, an initial interest in the distinct plumage and the vocal differences between adult female (drabber, non-singing) and male (more colorful, singing) zebra finches resulted in several, now classic, developmental studies. Some of these studies concentrated on the role of early life experience, through chromatic and vocal sexual imprinting, on females choosing attractive males as mates, while others focused on song production and song preference learning by male and female zebra finches (e.g. Clayton, 1987; Eales, 1987). For example, cross-fostering zebra finch chicks with the ‘universal estrildid foster species’, the Bengalese finch (Lonchura striata vars. domestica; Sonnemann and Sjölander, 1977), revealed that both visual and acoustic cues of social parents are learned during early development and used by young zebra finches of both sexes in mate preference following maturity (ten Cate, 1987; Campbell and Hauber, 2009; Verzijden et al., 2012). This occurs through a two-stage process of sexual imprinting (ten Cate, 1985; ten Cate and Voss, 1999).
These ontogenetic, physiological, and behavioral studies since the last quarter of the 20th century (e.g. Price, 1979) have become increasingly coupled with the rapid advances of neuroanatomical and neurophysiological imaging, genome sequencing, and transcriptomic and epigenetic analyses of the neural circuitries of song production in the forebrains of songbirds (reviewed in Mooney, 2009; Mooney, 2014) and song perception (reviewed in Louder et al., 2019). For instance, neurophysiological (Hauber et al., 2013), neuroanatomical (Lauay et al., 2005), immediate-early gene (Tomaszycki et al., 2006), and transcriptomic analyses (Louder et al., 2018) performed on zebra finch females that were reared either in isolation from any male birdsong or in the presence of a different songbird species have confirmed the critical role of early life experience in generating adaptive cognitive-behavioral (Price, 1979), neurogenomic (Louder et al., 2018) and neurophysiological (Moore and Woolley, 2019) responses to conspecific songs. Similarly, the known upregulation of stress responses of formerly pair-bonded, but then separated captive zebra finches (Remage-Healey et al., 2003), is also reported to impact the epigenomic status of similarly treated birds (George et al., 2020).
Despite the earlier prominence of the domestic canary (Serinus canaria) in the neurobiological study of song learning, two other research themes have also benefited significantly from follow-up studies of captive zebra finches. First, adult-onset neurogenesis, accompanying seasonal changes in song behavior, or damage to the underlying neural circuitry, was initially extensively studied in the canary (e.g. Nottebohm, 1981), but with ongoing critical contributions also coming from experiments on zebra finches (e.g. Walton et al., 2012; reviewed in Pytte, 2016). For example, when adult male zebra finches’ RA- (robust nucleus of the arcopallium) and Area X-projecting HVC neurons (Figure 2B) were experimentally ablated, only the RA-projecting neurons were regenerated (Scharff et al., 2000). In turn, a new social environment (e.g. through the exposure to novel aviary mates: Barnea et al., 2006, and/or ongoing auditory experiences: Pytte et al., 2010) may also contribute to the diminished apoptosis of newly generated caudomedial nidopallium (NCM) neurons (Figure 2B) in the forebrains of adults.
Second, hair cell regeneration following a loud noise or antibiotic treatment in both Bengalese (Woolley and Rubel, 2002) and zebra finches (Dooling and Dent, 2001) occurs rapidly, as it does in other, non-oscine birds (Stone and Rubel, 2000) and in some other vertebrate lineages (e.g. fish: Monroe et al., 2015). Research into such auditory system regeneration abilities in birds and other animals had strongly promised, but has thus far evaded, broadly applicable biomedical solutions for curing cell-death based hearing losses in humans (Brigande and Heller, 2009; Menendez et al., 2020).
Differences in captive vs. wild zebra finches and comparisons with northern hemisphere songbirds
Most of the populations of zebra finches in research laboratories around the world have been founded with birds held by aviculturists for over a hundred generations (Zann, 1996; Griffith et al., 2017). These populations have therefore been subject to both direct and indirect forms of natural and artificial selection, as well as founding effects, genetic drift, and inbreeding (Forstmeier et al., 2007; Knief et al., 2015). It has long been known that birds of the domesticated stocks are up to 30% larger in body size than their wild counterparts (Zann, 1996), but reassuringly they appear to be similar with respect to several life history trade-offs, including, for example, slow juvenile feather development and low adult song rates when nestlings are raised in large brood sizes (e.g. Tschirren et al., 2009). Captive birds are also similar to their wild counterparts in respect to the genomic architecture underlying complex traits (Kim et al., 2017; Knief et al., 2016; Knief et al., 2017a), although some caution still needs to be applied, for instance, to known differences in linkage disequilibrium patterns within the genomes of captive and wild populations (Knief et al., 2017b).
The pattern of zebra finches being quite different from many of the species of small passerines that are well studied by researchers in the northern hemisphere may be of greater significance than the differences between captive and wild populations of zebra finches. The zebra finch is an estrildid (Sorenson et al., 2004; Olsson and Alström, 2020), a family that is endemic to the tropics, and found across Africa, Southern Asia, and Australasia – with the whole lineage having evolved far from the ecological and evolutionary pressures of the temperate northern hemisphere. One of the almost ubiquitous characteristics of the estrildid family is the interseasonal strength of the socially monogamous pair-bond and biparental care for the young (Payne, 2010).
Prior breeding experience enhances the success of subsequent breeding bouts by female zebra finches through increased output and shorter times between clutches, even when breeding with a new male in this otherwise lifetime pair-bonded species (Adkins-Regan and Tomaszycki, 2007; Smiley and Adkins-Regan, 2016; Hurley et al., 2020). Relatively high within-pair sexual fidelity and cooperation in nest building, incubation, and provisioning also allow for the directed breeding of known pairs both in large aviaries and in small single-pair cages. Nevertheless, in socially housed groups, both conspecific brood parasitism – inducible by simulated nest predation in captivity (Shaw and Hauber, 2009) and accounting for 5 to 11% of offspring (Griffith et al., 2010) – as well as extra-pair paternity – accounting for around 30% of offspring in aviaries (Forstmeier et al., 2011) – can partially confound social parentage, although extrapair paternity is almost entirely absent in the wild (accounting for ~1% of offspring; Griffith et al., 2010).
A major effort of laboratory-based work on the zebra finch has focused on females’ mate choices (especially with respect to beak color and learned song; Griffith and Buchanan, 2010a). However, despite considerable variance in the reproductive success of individuals even in captive populations (Griffith et al., 2017; Wang et al., 2017), one of the most comprehensive studies examining the consequence of mate choice on fitness found no evidence that either males or females are targeting this variation in individual quality when they choose a partner (Wang et al., 2017). This finding supports the idea that the strength of a partnership is of greater value than the intrinsic quality of the individuals involved.
In this respect, zebra finches may differ from similarly-sized well studied small passerines of the northern hemisphere temperate zone. Since adult zebra finches are likely to live between 3 and 5 years in the wild (Zann, 1996) and can breed continuously throughout the year if conditions are favorable (Griffith et al., 2017), they can potentially accrue considerable experience as part of the sexual-parental partnership. The reproductive benefits of better physiological and behavioral coordination between partners (e.g. Adkins-Regan and Tomaszycki, 2007; Smiley and Adkins-Regan, 2016; Hurley et al., 2020) may outweigh the benefits of frequent and repeated partner switching and genetic infidelity (Griffith, 2019). In turn, the value of the partnership may promote selection for diverse affiliative and cooperative traits, not always seen in the widely studied passerines of the more seasonally constrained northern hemisphere, where most individuals breed just once or twice in a lifetime (Griffith, 2019). Rather, these traits are reminiscent of the long-term cooperative breeding partnerships formed (and the fitness costs paid following divorce or mate loss) by long-lived biparental seabirds (e.g. Ismar et al., 2010).
Indeed, the strength of the pair bond in the wild zebra finch is seen in the expression of acoustic communication throughout the year, and high levels of coordinated duetting between the male and female (Elie et al., 2010). This close, and regular vocal interaction between the members of a pair also perhaps plays a role in individual vocal recognition in this species (Levréro et al., 2009; Elie and Theunissen, 2018; Yu et al., 2020).
Highly coordinated acoustic interactions between female and male partners are a characteristic of the earliest passerine lineages as they had evolved in Australia (Odom et al., 2014). The continuously high level of overall acoustic activity in the zebra finch, which has made it such an attractive model system for neurobiology, sets it apart from many other well studied passerines in the northern hemisphere. This serves to remind us that although most of the laboratory work is conducted in the northern hemisphere, the zebra finch is, in many respects, different from most of the short-lived highly seasonally breeding passerines native to the temperate zone of the northern hemisphere. Indeed, it is important to understand that the species’ adaptations to the highly unpredictable Australian climate and ecology – while making it so easy to maintain and breed in captivity – also set it apart from most other northern hemisphere lineages that could not be used in laboratories to anywhere near the same extent.
Genes and brains for vocal learning
The process through which developing young memorize the acoustic communication signals of adults in humans and songbirds has been a critical research rationale and funding source supporting zebra finch studies. The learning of adult male songs by juveniles is particularly strong during early sensory periods, when embryos (Antonson et al., 2021), nestlings (Rivera et al., 2019), and juveniles (Brainard and Doupe, 2000) likely form a sensory representation of the 'tutor song' (Figure 3). Just as juvenile females develop long-term song-type preferences used for mate choice based on early experiences with their own fathers (Riebel, 2000; Chen et al., 2017), young males also learn and then actively practice to produce songs that match their paternal (tutor) songs (Tchernichovski et al., 2001; Figure 3). Tutors even alter their song structure when singing near young tutees, which influences the song learning process for young zebra finches, analogous to humans changing their speech when speaking to infants (Chen et al., 2016; Carouso-Peck and Goldstein, 2019).
Figure 3
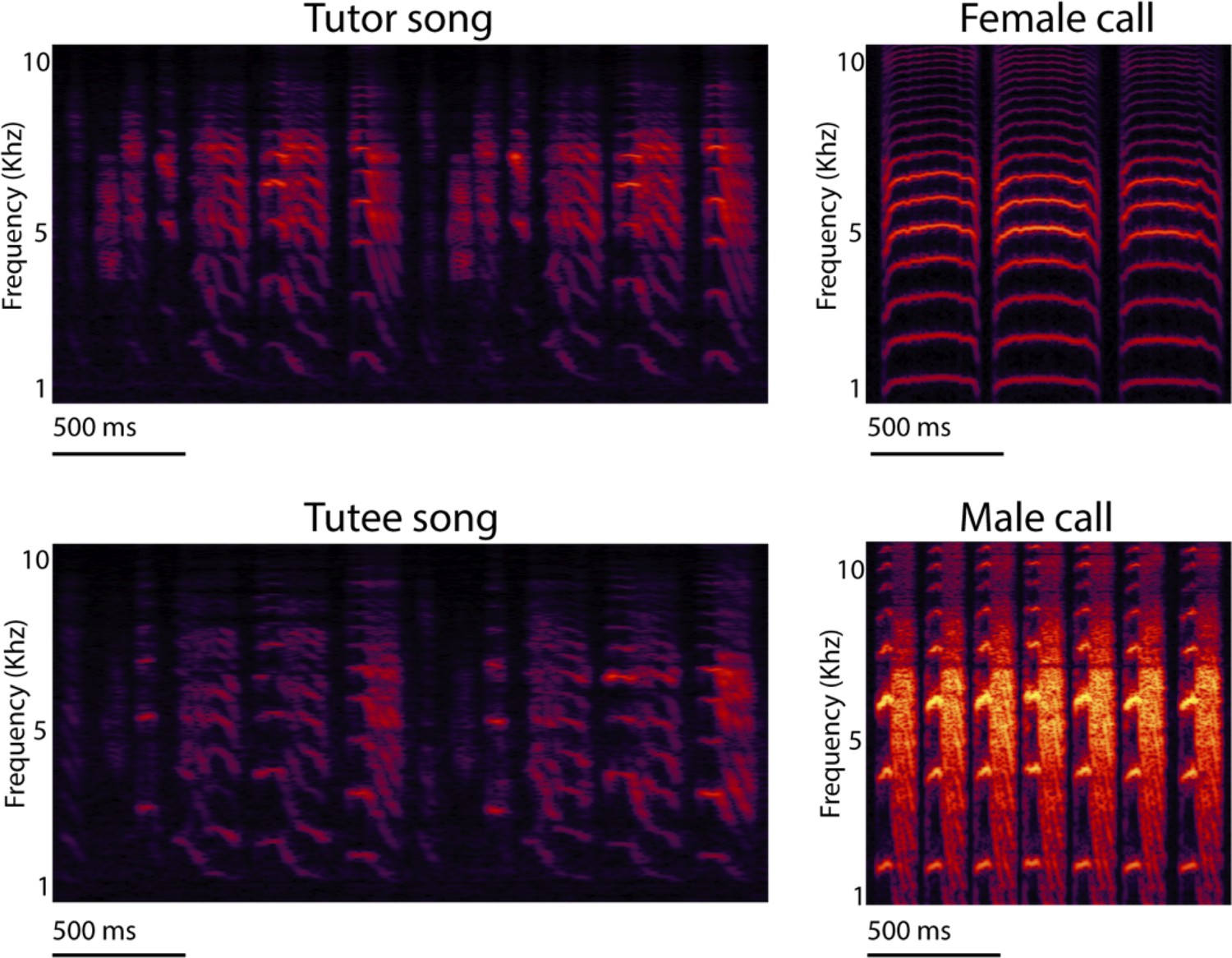
Spectrograms of zebra finch songs and calls.
Spectrogram of tutor and tutee adult male zebra finch songs, and undirected contact calls of adult females and males. Spectrograms represent time (x-axes) and pitch (y-axes) with greater amplitude as increasing brightness. Note the similarity of the tutor (typically social father) and tutee (son) song pair of male zebra finches and the distinct sexual differences of the calls.
However, even in the case of strong social environmental impact upon song learning during the sensitive period, the genetic make-up of individuals may contribute to the resulting song preferences and vocal production patterns through gene-by-environment interactions (Mets and Brainard, 2019). Accordingly, in zebra finches, males preferentially learn to sing from song tutors of the same species over those of another species when given equal access (Clayton, 1988), and both song-naïve and cross-fostered females show greater neuronal spike rates in response to unfamiliar conspecific over an unfamiliar third species’ songs (Hauber et al., 2013). Similarly, the species-specific typical pattern of socially learned song structure can culturally evolve across of just a handful of generations in initially naïve zebra finch populations (Fehér et al., 2009; Diez and MacDougall-Shackleton, 2020).
In adulthood, male and female zebra finches can quickly memorize individual vocal characteristics and recognize the identity of others for at least a month without reinforcement (Yu et al., 2020), likely relying on the perception of extremely small differences in calls and songs (Prior et al., 2018). However, experiences with other songs in adulthood do not affect the crystallized songs of males. Given the parallels with language acquisition and speech development in humans, zebra finches have thus long served as an important model for studying the neural mechanisms that control how vocal signals are memorized and copied (Doupe and Kuhl, 1999).
Initial research in the neurobiology of songbirds, primarily with canaries, has revealed the components and plasticity of the neural loops and circuits responsive to learning and producing songs (Figure 2B). Over time, studies of the zebra finch (a species that crystallizes its specific song once and does not deviate from it unless experiencing trauma or training) have become increasingly more instructive in the pursuit of identifying where in the forebrain the auditory memories are stored and how this representation directs both vocal learning in males and mate choice preferences in females (reviewed in Hauber et al., 2010). Accordingly, following the presentation of tape-recorded songs of conspecifics, the expression level of an immediate-early gene, egr-1 (also known as ZENK), which is associated with neural activation, increases within the zebra finch auditory forebrain, as found in other songbird species (Mello et al., 1992; Louder et al., 2016).
Furthermore, neural responses within the NCM, a subregion of the auditory forebrain, are selective for tutor songs (Yanagihara and Yazaki-Sugiyama, 2016) and song-induced expression of neural transcription factors (again, ZENK) also positively correlate with the increased similarity of the bird’s copied song to that of the tutor (Bolhuis et al., 2000), which together suggest that this region may hold the tutor song’s memory. Accordingly, NCM lesions in adult male zebra finches reduce their ability to recognize songs, but not to produce them (Gobes and Bolhuis, 2007). In female zebra finches, on the other hand, behavioral preferences for conspecific versus heterospecific songs can be eliminated by damaging the nearby CMM nucleus (caudomedial mesopallium) (MacDougall-Shackleton et al., 1998).
Overall, the zebra finch remains the best model system to characterize the neural circuitry involved in vocal learning and production, with an often-stated research aim to better understand the capacity of imitative speech learning in humans (e.g. Lipkind et al., 2013). Juvenile male zebra finches mimic the tutor song while females only produce non-learned ‘calls’ (Figure 3). In turn, several regions in the zebra finch brain associated with song production are dramatically larger in male zebra finches, a result of neurons in some of these regions atrophying in females while increasing in size and connections in males (Figure 2B; Konishi and Akutagawa, 1985). Several of these regions selectively respond to the ‘bird’s-own-song’ in anesthetized males (Doupe and Konishi, 1991), which initially suggested a specialized function for this circuit in producing songs; however, the role of such own-song specific auditory responses is no longer clear, as they are gated by behavioral states (Hessler and Doupe, 1999) and much less pronounced in awake birds (Schmidt and Konishi, 1998).
The premotor circuit for song production receives input from auditory nuclei via the HVC, which then projects to the RA, and subsequently connects to the brainstem motor nuclei and syrinx (Figure 2B). This ‘motor pathway’ is crucial during the learning process (Aronov et al., 2008) to generate stereotyped adult songs (Simpson and Vicario, 1990). In turn, while singing, neurons in the HVC that connect to the robust nucleus of the arcopallium (RA) perform time-locked bursts of firing, coincident with precise sequences during the song (Hahnloser et al., 2002). HVC neurons also ontogenetically shift their spike rates to become increasingly sparser while producing the male’s song (Okubo et al., 2015), whereas the spike trains of RA neurons lock into the timing of song’s note identity (Ac and Margoliash, 2008). By altering the local temperature of specific brain nuclei, Long and Fee, 2008 demonstrated that the temporal match between HVC, but not RA, and the song‘s timing pattern is a causal link, as cooling the HVC, but not the RA, slows down the song without affecting its frequency content. This demonstrates how and which elements of this forebrain circuit are critical to controlling the temporal structure of male songs and, in the Bengalese finch, their syntax, too (Zhang et al., 2017). By contrast, the anterior forebrain pathway (AFP), homologous to the mammalian basal ganglia–thalamocortical pathway, is required for vocal learning in juvenile male zebra finches, but not the production of stereotyped adult song (Bottjer et al., 1984). In this pathway, Area X and the lateral magnocellular nucleus of the anterior nidopallium (LMAN) are involved in producing song variability in juvenile birds during vocal learning (Woolley and Kao, 2015; Figure 2B).
Specifically, both theoretical modelling (including in humans) and experimental studies of this pathway (in zebra finches) have pointed to the critical role of vocal motor variability as the substrate upon which trial-and-error learning through reinforcement mechanisms may operate to shape vocal production ontogeny (Dhawale et al., 2017). In turn, the AFP is also involved in auditory-feedback based acoustic correction signaling for the motor pathway, in that inactivation of LMAN in young male zebra finches regresses experimentally induced, recently learned changes in the subjects’ song pitch (Andalman and Fee, 2009). Finally, gene expression patterns, including genes associated with speech in humans such as the transcription factor FOXP2, are highly expressed in the anterior forebrain pathway during sensitive periods for song learning, indicating potential genetic parallels of vocal plasticity in birds and humans (Haesler, 2004; Pfenning et al., 2014).
How the memorized tutor song instructs vocal pathways remains unclear. However, research in the zebra finch points to the involvement of nuclei within and outside of the anterior forebrain pathway. Auditory feedback, in which self-uttered and self-heard vocalizations are compared to a memorized song pattern, is necessary for the development of song in juveniles and the maintenance of song in adult zebra finches (Price, 1979; Nordeen and Nordeen, 1992; Leonardo and Konishi, 1999). Dopaminergic neurons of the ventral tegmental area (VTA) that project to the anterior forebrain pathway through Area X encode perceived errors in song performance from auditory feedback (Gadagkar et al., 2016; Figure 2B). The VTA receives error signals from auditory feedback through the AIV, which receives connections from the auditory forebrain (Kearney et al., 2019). Furthermore, neurons within the auditory forebrain also demonstrate sensitivity to errors in auditory feedback (Keller and Hahnloser, 2009). Such developments, for example regarding error sensitivity, also illustrate how ongoing research and continued breakthroughs in zebra finch neuroscience hold promise to further identify and understand the neural basis of vocal learning and production in general.
Following the widespread use of immediate early gene studies (see above), some of the research efforts aiming to characterize the genes that regulate zebra finch vocal and auditory behaviors, in particular genes related to vocal production in the brain, were based on utilizing DNA microarrays (Wada et al., 2006). Then, in 2010 an international consortium sequenced, assembled, and annotated the first zebra finch genome (Warren et al., 2010), only the second avian genome presented. This effort revealed the sequences of over 17,000 predicted protein-coding genes, as well as many regulatory regions and non-coding RNAs. More importantly, the annotated genome enhanced the next decade’s analyses into identifying the genes and regulatory networks that are involved in social behavior, including genome-wide investigations into vocal learning, such as auditory-experience induced RNA expression (Louder et al., 2018), microRNA expression (Gunaratne et al., 2011), and epigenetically regulated genes associated with developmental song learning (Kelly et al., 2018). Furthermore, the initial genome helped researchers to identify and map the expression patterns of ~650 candidate genes within the brain of zebra finches, resulting in an online atlas database that provides an opportunity to link behavior, neuroanatomy, and molecular function (Lovell et al., 2020).
A recent high quality, second generation genome of the zebra finch, presented as part of the Vertebrate Genomes Project, improves the accuracy of the reference genome assembly and annotation (Rhie et al., 2021). Leveraging recent technological advances, such as long-read sequencing (up to 100 Kbp) and approaches to detect how DNA interacts across genomic loci (up to 100 Mbp), the latest updated zebra finch genome thus resolves numerous regions with repetitive elements and enhanced gene annotation from the first assembly.
In parallel with genomic advances, a suite of new neurobiological techniques available for zebra finches will only continue to increase the ability to understand the development of vocal learning and behavior. Questions regarding the activity of specific neurons can now be tackled using multi-electrode arrays (e.g. Lim et al., 2016; Tanaka et al., 2018) or wireless neurotelemetry (Ma et al., 2020) able to simultaneously record the activity of numerous neurons in awake and freely-behaving birds. Imaging the neural connections between distant brain regions is now also possible with tissue clearing and light-sheet microscopy (Rocha et al., 2019).
The experimental regulation of the expression of candidate genes in targeted areas of the zebra finch brain has also recently become available. Existing or new gene constructs can be inserted into neonatal (hatchling) zebra finches via electroporation-based gene construct delivery to study the genetics of vocal learning as songs are memorized, practiced, and first expressed by young males (Ahmadiantehrani and London, 2017). Similarly, genetically modified constructs of nonpathogenic viruses injected in the brain, such as adeno-associated virus (AAV), are able to drive the expression of certain genes.
Viral constructs were developed to control the expression of FOXP2 (e.g. Heston and White, 2015; Norton et al., 2019), which is expressed in the song control regions within the male zebra finch forebrain and associated with inherited speech and language disorder in humans (Fisher and Scharff, 2009). Viral constructs have also been useful in imaging, such as expressing a genetically encoded calcium indicator (GCaMP6s) for calcium imaging of neuron populations with 2-photon microscopy (Picardo et al., 2016) or the expression of green fluorescent protein (GFP). Recent applications of viral constructs have also enabled researchers to control neurons with light (optogenetics), such as ‘implanting’ artificial song memories into the zebra finch brain (Zhao et al., 2019), or controlling the firing of specific neurons, such as the VTA neurons that project to Area X (Xiao et al., 2018; Kearney et al., 2019). Harnessing these new techniques enables us to tackle how genetic pathways are linked to vocal learning and motor control circuits.
However, the utility of the zebra finch as a neurogenetic model laboratory species has been somewhat inhibited by the low success rate in the development of transgenic lines that would enable direct experimental modification of the gene expression patterns in the relevant vocal-production and vocal-perception circuits. This may be due to the unique immune function of oscine birds inhibiting full viral delivery of gene constructs (London, 2020). Nevertheless, the last decade has already seen the successful innovation of lentiviral delivery (e.g. Norton et al., 2019) of, for example, human Huntington’s Disease genes into zebra finch lineages, to causally demonstrate reduced vocal imitation and output consistency as a result of the treatment (Liu et al., 2015). However, to date neither a TALEN nor a CRISPR/Cas9 vector-based gene editing approach has taken off in avian (chicken or songbird) lineages (Woodcock et al., 2017; but see Cooper et al., 2018). With additional research, the zebra finch could be further explored as to which gene delivery and genomic editing methods will be widely and effectively applicable to this species.
The importance of studying female zebra finches
Female zebra finches only slowly and partially assumed a role in some of the earlier behavioral and developmental studies on sexual imprinting (e.g. Collins et al., 1994), but now maintain a co-lead position. This is because mate choice is mutual in this species and females participate in the ever-important initial pair-bonding decisions, as well as in all aspects of collaborative biparental care (Riebel, 2009). As such, females make a critical contribution to the phenotype of their offspring through their investments into eggs, and the care of dependent offspring (Griffith and Buchanan, 2010b). Still, in studying the neurobiological basis of species and mate recognition, and the relevant funding and publications, female-focused research took a secondary role during the earlier decades when much of the work focused on the developing and adult sensory-motor circuitries of the male zebra finch forebrain.
In the last two decades, however, there has been a definite upsurgence of studies focusing on female zebra finches, both from the perspective of the neurosensory-ontogenetic processes of conspecific (Theunissen et al., 2004; Woolley et al., 2010), mate (Lauay et al., 2004; Tokarev et al., 2017), and individual recognition (Vignal et al., 2004; D'Amelio et al., 2017; Yu et al., 2020) by and of females. It is becoming clear that female visual and acoustic displays serve an important role in the development and fine-tuning of male vocalizations during sensitive periods (Benichov et al., 2016; Carouso-Peck and Goldstein, 2019) and that male vocal and/or visual displays serve in the activation of auditory forebrain regions in adult females (Avey et al., 2005; Day et al., 2019).
For example, the reduced volume of the song control system that exists in the female zebra finch brain is likely not at all vestigial (Shaughnessy et al., 2019) and may be even more functional than previously thought, enabling plasticity in the vocal timing of calls in social interactions (Benichov et al., 2016). In turn, female (and male) parental vocal communication with embryos in ovo in the nest have also been discovered to shape not only the functional neurogenomic responses of the embryos themselves (Rivera et al., 2019) but also the acoustic tutor choice of young male zebra finches (Katsis et al., 2018), as well as adult behavioral phenotypes and reproductive success (Mariette and Buchanan, 2016).
Finally, the behavioral, the neurophysiological and gene-activational bases of perceptual learning of conspecific song features appear to be both species-specific in song-naïve (mother-only parent raised) female zebra finches and dependent on early social experience with con- or cross-fostered heterospecific male songs (Hauber et al., 2013; Louder et al., 2018). Some of these latter discoveries in females have been made possible through cross-fostering nestling zebra finches with estrildid finch tutors of other species (e.g. Clayton, 1987). Critically, the results from females have now also been both replicated and advanced in cross-fostered males. Specifically, the extent of heterospecific song learning in males can be directly measured by the altered songs that they produce following experimental manipulation of early song exposure, and compared with the extent of neurophysiological response selectivity for conspecific (innate) vs. heterospecific (learned) tutor songs and their contributory bioacoustic features in the brain (Moore and Woolley, 2019). In turn, cross-fostered males singing the foster species’ song famously show an inability to copy the temporal pattern of heterospecific songs, discovered to be due to a lack of ontogenetic flexibility in the neurons that encode heterospecific song-gap (silent period between song bouts) perception again within field L of the auditory forebrain (Araki et al., 2016).
Conclusions
The zebra finch was not originally brought into the laboratory as a model system, nor championed as such by early research pioneers. From the 1950s onwards, the species has been progressively adopted as a useful focus of study in an increasing set of research fields, largely due to its accessibility and the ease with which it can be held and bred in captivity. In contrast, wild passerine birds have long been the focus of ecological and evolutionary research in the northern hemisphere. When studies of free-living study populations were unable to achieve the necessary manipulative rigor, the zebra finch, found commonly in pet shops throughout Europe and North America, became widely adopted as a surrogate captive experimental model. In parallel with its use in early ethological research, the zebra finch became established as an easier model than the canary for studying the neural basis of song, which in turn saw the former species adopted as a model for genomics, neuroscience, and developmental biology.
The zebra finch has provided great insights into diverse fields in biology and has travelled a long path from its natural habitat in arid Australia. It is important to be mindful that the traits that have contributed to its utility and adoption as ‘the’ avian laboratory model species for basic and biomedical research set it aside from most other avian species. The zebra finch evolved in an austral ecological setting that is profoundly different from those in the many geographic regions where most of this laboratory work takes place.
The zebra finch remains almost uniquely suited as a model system for research and the path ahead is likely to be productive and insightful in established and new areas of research. The late Richard Zann’s excellent monograph of the species (1996), whilst already over two decades old, still provides an excellent overview into the natural history of the species, and is never far from our desks, for the insight that it brings. We encourage future adopters of the zebra finch as a research model to use this book to guide their planning and to help interpret their results. The zebra finch is the most widely researched laboratory songbird in the world because of its uniqueness, and not as a result of any advocacy.
Data availability
No new data were generated in this study.
References
-
Higher temperatures during development reduce body size in the zebra finch in the laboratory and in the wildJournal of Evolutionary Biology 30:2156–2164.https://doi.org/10.1111/jeb.13181
-
The zebra finch genome and avian genomics in the wildEmu - Austral Ornithology 110:233–241.https://doi.org/10.1071/MU09087
-
Auditory feedback in learning and maintenance of vocal behaviourNature Reviews Neuroscience 1:31–40.https://doi.org/10.1038/35036205
-
Water economy and metabolism of two estrildine finchesPhysiological Zoology 38:9–33.https://doi.org/10.1086/physzool.38.1.30152342
-
Context and control: behavioural ecology experiments in the laboratoryAnnales Zoologici Fennici 46:112–123.https://doi.org/10.5735/086.046.0204
-
Geographic variation in zebra finch subspeciesEmu - Austral Ornithology 91:2–11.https://doi.org/10.1071/MU9910002
-
Female mate choice in the zebra finch? The effect of male beak colour and male songBehavioral Ecology and Sociobiology 35:21–25.https://doi.org/10.1007/BF00167055
-
Innovative approaches to genome editing in avian speciesJournal of Animal Science and Biotechnology 9:15.https://doi.org/10.1186/s40104-018-0231-7
-
Physiological responses of wild zebra finches (Taeniopygia guttata) to heatwavesJournal of Experimental Biology 223:jeb225524.https://doi.org/10.1242/jeb.225524
-
Effect of acute exposure to high ambient temperature on the thermal, metabolic and hygric physiology of a small desert birdComparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology 244:110684.https://doi.org/10.1016/j.cbpa.2020.110684
-
Gene regulatory evolution during speciation in a songbirdG3: Genes, Genomes, Genetics 6:1357–1364.https://doi.org/10.1534/g3.116.027946
-
D2 dopamine receptor activation induces female preference for male song in the monogamous zebra finchJournal of Experimental Biology 222:jeb191510.https://doi.org/10.1242/jeb.191510
-
The role of variability in motor learningAnnual Review of Neuroscience 40:479–498.https://doi.org/10.1146/annurev-neuro-072116-031548
-
New studies on hair cell regeneration in birdsAcoustical Science and Technology 22:93–99.https://doi.org/10.1250/ast.22.93
-
Birdsong and human speech: common themes and mechanismsAnnual Review of Neuroscience 22:567–631.https://doi.org/10.1146/annurev.neuro.22.1.567
-
FOXP2 as a molecular window into speech and languageTrends in Genetics 25:166–177.https://doi.org/10.1016/j.tig.2009.03.002
-
Low level of extrapair parentage in wild zebra finchesAnimal Behaviour 79:261–264.https://doi.org/10.1016/j.anbehav.2009.11.031
-
Cooperation and coordination in socially monogamous birds: moving away from a focus on sexual conflictFrontiers in Ecology and Evolution 7:455.https://doi.org/10.3389/fevo.2019.00455
-
The zebra finch: the ultimate Australian supermodelEmu - Austral Ornithology 110:v–0.https://doi.org/10.1071/MUv110n3_ED
-
Maternal effects in the zebra finch: a model mother reviewedEmu - Austral Ornithology 110:251–267.https://doi.org/10.1071/MU10006
-
FoxP2 expression in avian vocal learners and non-learnersJournal of Neuroscience 24:3164–3175.https://doi.org/10.1523/JNEUROSCI.4369-03.2004
-
Primordial germ cell-mediated transgenesis and genome editing in birdsJournal of Animal Science and Biotechnology 9:19.https://doi.org/10.1186/s40104-018-0234-4
-
The functional role and female perception of male song in zebra finchesEmu - Austral Ornithology 110:209–218.https://doi.org/10.1071/MU10003
-
Social context modulates singing-related neural activity in the songbird forebrainNature Neuroscience 2:209–211.https://doi.org/10.1038/6306
-
Behavior-linked FoxP2 regulation enables zebra finch vocal learningJournal of Neuroscience 35:2885–2894.https://doi.org/10.1523/JNEUROSCI.3715-14.2015
-
Reproductive coordination breeds success: the importance of the partnership in avian sperm biologyBehavioral Ecology and Sociobiology 74:3.https://doi.org/10.1007/s00265-019-2782-9
-
Birdsong learning and culture: analogies with human spoken languageAnnual Review of Linguistics 7:449–472.https://doi.org/10.1146/annurev-linguistics-090420-121034
-
ConferenceThe influence of early experience upon the development of social behaviour in estrildine finchesProceedings of the XV International Ornithological Congress. pp. 291–313.
-
Mate replacement entails a fitness cost for a socially monogamous seabirdNaturwissenschaften 97:109–113.https://doi.org/10.1007/s00114-009-0618-6
-
A sex-linked supergene controls sperm morphology and swimming speed in a songbirdNature Ecology & Evolution 1:1168–1176.https://doi.org/10.1038/s41559-017-0235-2
-
A sex-chromosome inversion causes strong overdominance for sperm traits that affect siring successNature Ecology & Evolution 1:1177–1184.https://doi.org/10.1038/s41559-017-0236-1
-
The variability of song variability in zebra finch (Taeniopygia guttata) populationsRoyal Society Open Science 6:190273.https://doi.org/10.1098/rsos.190273
-
Begging calls support offspring individual identity and recognition by zebra finch parentsComptes Rendus Biologies 332:579–589.https://doi.org/10.1016/j.crvi.2009.02.006
-
Human mutant huntingtin disrupts vocal learning in transgenic songbirdsNature Neuroscience 18:1617–1622.https://doi.org/10.1038/nn.4133
-
Gene manipulation to test links between genome, brain and behavior in developing songbirds: a test caseJournal of Experimental Biology 223:jeb206516.https://doi.org/10.1242/jeb.206516
-
Neural mechanisms of auditory species recognition in birdsBiological Reviews 94:1619–1635.https://doi.org/10.1111/brv.12518
-
ZEBrA: Zebra Finch Expression Brain Atlas—a resource for comparative molecular neuroanatomy and brain evolution studiesJournal of Comparative Neurology 528:2099–2131.https://doi.org/10.1002/cne.24879
-
Neurotelemetry reveals putative predictive activity in HVC during call-based vocal communications in zebra finchesThe Journal of Neuroscience 40:6219–6227.https://doi.org/10.1523/JNEUROSCI.2664-19.2020
-
The size and composition of social groups in the wild zebra finchEmu - Austral Ornithology 115:191–198.https://doi.org/10.1071/MU14059
-
Sensory hair cell death and regeneration in fishesFrontiers in Cellular Neuroscience 9:131.https://doi.org/10.3389/fncel.2015.00131
-
Neural mechanisms for learned birdsongLearning & Memory 16:655–669.https://doi.org/10.1101/lm.1065209
-
Auditory–vocal mirroring in songbirdsPhilosophical Transactions of the Royal Society B: Biological Sciences 369:20130179.https://doi.org/10.1098/rstb.2013.0179
-
Emergent tuning for learned vocalizations in auditory cortexNature Neuroscience 22:1469–1476.https://doi.org/10.1038/s41593-019-0458-4
-
Auditory feedback is necessary for the maintenance of stereotyped song in adult zebra finchesBehavioral and Neural Biology 57:58–66.https://doi.org/10.1016/0163-1047(92)90757-U
-
Female song is widespread and ancestral in songbirdsNature Communications 5:3379.https://doi.org/10.1038/ncomms4379
-
A comprehensive phylogeny and taxonomic evaluation of the waxbills (Aves: estrildidae)Molecular Phylogenetics and Evolution 146:106757.https://doi.org/10.1016/j.ympev.2020.106757
-
BookFamily Estrildidae (waxbills)In: del Hoyo J, Elliott A, Christie D. A, editors. Handbook of the Birds of the World, 15. Barcelona: Lynx Edicions. pp. 234–377.
-
Developmental determinants of structure in zebra finch songJournal of Comparative and Physiological Psychology 93:260–277.https://doi.org/10.1037/h0077553
-
Deafening decreases neuronal incorporation in the zebra finch caudomedial nidopallium (NCM)Behavioural Brain Research 211:141–147.https://doi.org/10.1016/j.bbr.2010.03.029
-
Adult neurogenesis in the songbird: region-specific contributions of new neurons to behavioral plasticity and stabilityBrain, Behavior and Evolution 87:191–204.https://doi.org/10.1159/000447048
-
Song and female mate choice in zebra finches: a reviewAdvances in the Study of Behavior 40:197–238.https://doi.org/10.1016/S0065-3454(09)40006-8
-
Does zebra finch (Taeniopygia guttata) preference for the (familiar) father's song generalize to the songs of unfamiliar brothers?Journal of Comparative Psychology 117:61–66.https://doi.org/10.1037/0735-7036.117.1.61
-
Evaluating mate choice in the zebra finchAnimal Behaviour 74:1277–1284.https://doi.org/10.1016/j.anbehav.2007.02.022
-
BookNeural circuits underlying vocal learning in songbirdsIn: Sakata J. T, Woolley S. C, Fay R. R, Popper A. N, editors. The Neuroethology of Birdsong. New York: Springer. pp. 29–63.https://doi.org/10.1007/978-3-030-34683-6_2
-
Transgenesis and imaging in birds, and available transgenic reporter linesDevelopment, Growth & Differentiation 55:406–421.https://doi.org/10.1111/dgd.12058
-
Gating of auditory responses in the vocal control system of awake songbirdsNature Neuroscience 1:513–518.https://doi.org/10.1038/2232
-
Female zebra finches do not sing yet share neural pathways necessary for singing in malesJournal of Comparative Neurology 527:843–855.https://doi.org/10.1002/cne.24569
-
Experimental support for the role of nest predation in the evolution of brood parasitismJournal of Evolutionary Biology 22:1354–1358.https://doi.org/10.1111/j.1420-9101.2009.01745.x
-
Brain pathways for learned and unlearned vocalizations differ in zebra finchesThe Journal of Neuroscience 10:1541–1556.https://doi.org/10.1523/JNEUROSCI.10-05-01541.1990
-
Relationship between prolactin, reproductive experience, and parental care in a biparental songbird, the zebra finch (Taeniopygia guttata)General and Comparative Endocrinology 232:17–24.https://doi.org/10.1016/j.ygcen.2015.11.012
-
Effects of cross-fostering on the sexual imprinting of the female zebra finch Taeniopygia guttataZeitschrift Für Tierpsychologie 45:337–348.https://doi.org/10.1111/j.1439-0310.1977.tb02024.x
-
Clade-limited colonization in brood parasitic finches (Vidua spp.)Systematic Biology 53:140–153.https://doi.org/10.1080/10635150490265021
-
On sex differences in sexual imprintingAnimal Behaviour 33:1310–1317.https://doi.org/10.1016/S0003-3472(85)80191-3
-
Sexual imprinting and evolutionary processes in birds: a reassessmentAdvances in the Study of Behavior 28:1–31.https://doi.org/10.1016/S0065-3454(08)60214-4
-
An analysis of the neural representation of birdsong memoryJournal of Neuroscience 24:4971–4977.https://doi.org/10.1523/JNEUROSCI.0570-04.2004
-
Song selectivity in the song system and in the auditory forebrainAnnals of the New York Academy of Sciences 1016:222–245.https://doi.org/10.1196/annals.1298.023
-
Short- and long-term consequences of early developmental conditions: a case study on wild and domesticated zebra finchesJournal of Evolutionary Biology 22:387–395.https://doi.org/10.1111/j.1420-9101.2008.01656.x
-
The impact of learning on sexual selection and speciationTrends in Ecology & Evolution 27:511–519.https://doi.org/10.1016/j.tree.2012.05.007
-
The zebra finch paradox: song is little changed, but number of neurons doublesJournal of Neuroscience 32:761–774.https://doi.org/10.1523/JNEUROSCI.3434-11.2012
-
Gene editing in birds takes flightMammalian Genome 28:315–323.https://doi.org/10.1007/s00335-017-9701-z
-
Developmental experience alters information coding in auditory midbrain and forebrain neuronsDevelopmental Neurobiology 70:235–252.https://doi.org/10.1002/dneu.20783
-
Early experience shapes vocal neural coding and perception in songbirdsDevelopmental Psychobiology 54:612–631.https://doi.org/10.1002/dev.21014
-
Vocal memory and learning in adult Bengalese finches with regenerated hair cellsThe Journal of Neuroscience 22:7774–7787.https://doi.org/10.1523/JNEUROSCI.22-17-07774.2002
-
Mechanisms of species diversity in birdsong learningPLOS Biology 17:e3000555.https://doi.org/10.1371/journal.pbio.3000555
-
BookThe Zebra Finch: A Synthesis of Laboratory and Field StudiesOxford: Oxford University Press.https://doi.org/10.5860/choice.34-5095
-
Temperature manipulation in songbird brain Implicates the premotor nucleus HVC in birdsong syntaxThe Journal of Neuroscience 37:2600–2611.https://doi.org/10.1523/JNEUROSCI.1827-16.2017
Decision letter
-
Peter RodgersSenior Editor; eLife, United Kingdom
-
Helena Pérez ValleReviewing Editor; eLife, United Kingdom
-
Leslie PhillmoreReviewer; Dalhousie University, Canada
-
Mimi KaoReviewer; Tufts University, United States
In the interests of transparency, eLife publishes the most substantive revision requests and the accompanying author responses.
Thank you for submitting your article "The Natural History of Model Organisms: Neurogenomic insights into behavioral and vocal development of the zebra finch" to eLife for consideration as a Feature Article. Your article has been reviewed by three peer reviewers and the evaluation has been overseen by two editors from the eLife Features Team (Helena Pérez Valle and Peter Rodgers). The following individual involved in review of your submission has agreed to reveal their identity: Leslie Phillmore (Reviewer #3).
The reviewers and editors have discussed the reviews and we have drafted this decision letter to help you prepare a revised submission.
Summary:
This article reviews the history of the zebra finch as a species of choice to study different aspects of developmental neurobiology and vocal communication. However, the article is lacking references to important works, and should also discuss the roles of the descending motor pathway in relation to the anterior forebrain pathway for song learning and adult song production.
Essential revisions:
1. Please include references to the work from labs that have substantially contributed to the fields of neuroscience and behaviour using zebra finches, including the Fee lab (MIT), the Goldberg lab (Cornell), the Sakata lab (McGill). Also please ensure that the article includes citations to the key works of other labs that are already cited, including the Jarvis lab (Rockefeller), the Long lab (NYU), the Brainard lab (UCSF), the Tchernikovski lab (CUNY) and the Theunissen lab (Berkeley). Please add up to 20 references to address this point. Additionally, please consider the following:
a) Please cite Desmond Morris (1954 and 1957) along with Immelmann when talking about the reproductive behaviour of zebra finches (line 35), and try to find a reference for Immelmann that is not an abstract for a conference.
b) Please cite Katharina Riebel (2000) "Early exposure leads to repeatable preferences for male song in female zebra finches", or any of her later reviews, in addition to Braaten and Reynolds (1999) (line 40).
c) Please check whether Day, 2019 is the correct citation in line 48; and whether work by the White, Mooney, Roberts or Scharff labs would be more appropriate when talking about genome editing.
d) Please check whether there are references missing in line 51 after "illness".
e) Please add references for genetic diversity in line 73 (e.g. "Genetic variation and differentiation in captive and wild zebra finches (Taeniopygia guttata)", from the Forstmeier lab).
f) Please cite reviews on line 140, since the literature is extensive. "Sexual imprinting and evolutionary processes in birds: a reassessment", Ten Cate and Vos (1999), and "The impact of learning on sexual selection and speciation", Verzijden et al. (2012).
g) In line 145, Louder et al. covers auditory but not song production, please find an appropriate reference to cover both.
h) In line 320, please add a reference to Scharff's work on FoxP2
i) In line 334/35, please provide a reference for "the unique immune function of oscine birds inhibiting full viral delivery of gene constructs".
2. Please add between 500 and 1000 words describing the roles of the descending motor pathway vis-à-vis the anterior forebrain pathway for song learning and adult song production; and the use of the zebra finch to study auditory perception of both biologically relevant stimuli (i.e. conspecific vocalizations) and others (e.g. human speech patterns).
3. Figure 2B should include spectrograms of at least a female call (see line 272), perhaps both a male and female call, as well as the male song.
4. The authors' work currently makes up about a third of the references, please check whether these references are all necessary and whether it might be possible to replace some of them with references to other important works in the field.
[Editors' note: further revisions were suggested prior to acceptance, as described below.]
Thank you for resubmitting your work entitled "Natural History of Model Organisms: Neural and genomic insights into behavioral and vocal development of the zebra finch" for further consideration by eLife. Your revised article has been reviewed by a new peer reviewer (Mimi Kao), and the evaluation was overseen by Helena Pérez Valle as the Assistant Features Editor and Peter Rodgers as the Features Editor.
The manuscript has been improved but there are some remaining issues that need to be addressed, as outlined below.
Summary:
This article reviews the history of the zebra finch as a species of choice to study different aspects of developmental neurobiology and vocal communication, and particularly as a system for investigating sex differences in vocal communication. The revised manuscript is much improved, but it would still benefit of additional revisions to references, and including a discussion of studies of auditory perception using cross-fostered birds to examine innate preferences versus experience-dependent changes in auditory responses.
Essential revisions:
1. Please make the following modifications to references:
a) In line 37, add references to the work of the Fee and Yazaki-Sugiyama labs. These would be useful for neural mechanisms underlying sensorimotor learning (e.g. Okubo et al., 2015; Mackevicius, Happ, and Fee, 2020; Yanagihara and Yazaki-Sugiyama, 2019).
b) In lines 38-39, the reference to Chen et al., 2017 does not seem to be appropriate, since that study focuses on vocal learning, not female preference. Please remove this reference and replace it with another that examines influences on female song preferences (e.g. Riebel, 2000).
c) In lines 63 and/or 73, please add a reference to Cade et al., 1965, Water economy and metabolism of two estrildine finches, Physiological Zoology.
d) In lines 88-89, please add references to papers by Mets and Brainard (2018, 2019) that investigate the interaction of genes and social and/or auditory experience on vocal
learning and performance. Please add a short discussion about gene-environment interactions that shape song learning based on the findings of these papers in the section "Brains and genes".
e) In line 159 please add a reference to Price, PH (1979) Developmental determinants of structure in zebra finch song. Journal of Comparative and Physiological Psychology.
f) Please check the references in lines 258-260. While several papers have shown that both calls and songs have strong individual signatures that can be used for individual recognition, the references cited did not specifically examine "high levels of coordinated duetting between the male and female".
g) In lines 323-327, please add a reference to Aronov et al. (already cited in the article) in regard to the critical role of HVC for generating stereotyped, learned vocalization in lines 323-327.
h) In lines 330-331, please add references for the idea that the AFP generates song variability in juvenile birds (Olveczky et al., 2005, 2011; or a review article, e.g. Mooney, 2009; Woolley and Kao, 2015; or Dhawale, Smith and Olveczky 2017), and discuss these findings.
i) Please add references to studies showing that LMAN also sends an instructive signal that can bias song and drive systematic changes in mean pitch, mean duration and syllable transition probabilities (Turner and Brainard, 2007; Andalman and Fee, 2009; Ali et al., 2013, Charlesworth et al., 2012), and discuss these findings.
j) In lines 341-344, please add a reference for deafening experiments in juvenile birds, such as Price, 1979.
k) In lines 349-351, it is unclear why the Ma et al., 2020 reference is included, since the paragraph discusses mechanisms by which evaluation of song performance using auditory feedback can drive changes in song. Ma et al., 2020 report activity prior to calls by conspecifics. While predictive activity may be important for learning or maintaining song, please make clear what the activity is predicting (timing of upcoming calls? Expected auditory feedback given the motor commands?) and how it relates to learning the tutor song model.
l) In line 401, please add Xiao and Roberts, Neuron, 2018 as a reference, in addition to Kearney et al., 2019.
m) In lines 441-444, please add a reference to Shaughnessey et al., JCN, 2018 when mentioning the song system in females as that paper argues against the idea that females possess only vestiges of the song control network and suggests that the forebrain network may have a different function in females.
2. Given the manuscript's emphasis on vocal imprinting, please include a brief discussion of studies of auditory perception using cross-fostered birds to examine innate preferences versus experience-dependent changes in auditory responses (e.g. Araki et al., 2017; Moore and Woolley, 2019).
3. In lines 187-189, please modify your description of hair cell regeneration as "a specialized avian feat". Sensory hair cell regeneration after antibiotic treatment has been shown in other vertebrates, including fish (in the inner ear and lateral line; reviewed by Monroe et al., 2015, Front Cell Neurosci), as well as invertebrates (e.g. frogs).
4. Please revise the section "Brains and genes for vocal learning" to either omit mention of selective auditory responses, or expand this section for clarity (lines 319-321). While many studies have shown that neurons in both the motor pathway and the anterior forebrain pathway respond selectively to playback of bird's own song in anesthetized birds, the role of such auditory responses is not entirely clear. Auditory responses in HVC and in LMAN and Area X are gated by behavioral state and are much less pronounced in awake birds (Schmidt and Konishi , 1998; Hessler and Doupe, 1999).
5. Please expand the discussion of the role of HVC in coding the timing of song (lines 323-327). For example, HVC neurons that project to RA fire once per motif while RA neurons may fire multiple bursts at specific times in song (Yu and Margoliash, 1996). Similarly, Long and Fee, 2008 is referenced, but it would help the reader to explain how the cooling experiments help to establish that HVC codes for time in song.
https://doi.org/10.7554/eLife.61849.sa1Author response
Summary:
This article reviews the history of the zebra finch as a species of choice to study different aspects of developmental neurobiology and vocal communication. However, the article is lacking references to important works, and should also discuss the roles of the descending motor pathway in relation to the anterior forebrain pathway for song learning and adult song production.
We have now included a fully-cited section on the role of the motor pathway in mediating juvenile song learning and adult song production.
Essential revisions:
1. Please include references to the work from labs that have substantially contributed to the fields of neuroscience and behaviour using zebra finches, including the Fee lab (MIT), the Goldberg lab (Cornell), the Sakata lab (McGill).
Dozens of previously unmentioned labs’ references and discussion of these works are now added.
Also please ensure that the article includes citations to the key works of other labs that are already cited, including the Jarvis lab (Rockefeller), the Long lab (NYU), the Brainard lab (UCSF), the Tchernikovski lab (CUNY) and the Theunissen lab (Berkeley). Please add up to 20 references to address this point.
Additional references (65 new ones) and discussion of these works are now also added to the manuscript.
Additionally, please consider the following:
a) Please cite Desmond Morris (1954 and 1957) along with Immelmann when talking about the reproductive behaviour of zebra finches (line 35), and try to find a reference for Immelmann that is not an abstract for a conference.
Thank you. Done (Morris 1954) but Immelmann 1971 is a full article in the Proceedings of the 15th IOC, it’s not just a conference abstract.
b) Please cite Katharina Riebel (2000) "Early exposure leads to repeatable preferences for male song in female zebra finches", or any of her later reviews, in addition to Braaten and Reynolds (1999) (line 40).
The Riebel 2000 paper and a Riebel-lab 2003 paper are now included.
c) Please check whether Day, 2019 is the correct citation in line 48; and whether work by the White, Mooney, Roberts or Scharff labs would be more appropriate when talking about genome editing.
Thank you. We have removed the incorrect Day et al. reference here and added additional avian genome editing review instead.
d) Please check whether there are references missing in line 51 after "illness".
Reference (Han and Park 2018, London 2020) is added.
e) Please add references for genetic diversity in line 73 (e.g. "Genetic variation and differentiation in captive and wild zebra finches (Taeniopygia guttata)", from the Forstmeier lab).
New Forstmeier lab references are added.
f) Please cite reviews on line 140, since the literature is extensive. "Sexual imprinting and evolutionary processes in birds: a reassessment", Ten Cate and Vos (1999), and "The impact of learning on sexual selection and speciation", Verzijden et al. (2012).
Two review papers are now also added as citations.
g) In line 145, Louder et al. covers auditory but not song production, please find an appropriate reference to cover both.
We are now citing an additional reference (Mooney 2009) to cover song production.
h) In line 320, please add a reference to Scharff's work on FoxP2
The Scharff TiG review/reference to FoxP2 is added as well as other Scharff lab references.
i) In line 334/35, please provide a reference for "the unique immune function of oscine birds inhibiting full viral delivery of gene constructs".
Reference of London 2020 is added.
2. Please add between 500 and 1000 words describing the roles of the descending motor pathway vis-à-vis the anterior forebrain pathway for song learning and adult song production; and the use of the zebra finch to study auditory perception of both biologically relevant stimuli (i.e. conspecific vocalizations) and others (e.g. human speech patterns).
We have now added discussion of both of these themes to the manuscript.
3. Figure 2B should include spectrograms of at least a female call (see line 272), perhaps both a male and female call, as well as the male song.
Call spectrograms are also added and turned into a new figure.
4. The authors' work currently makes up about a third of the references, please check whether these references are all necessary and whether it might be possible to replace some of them with references to other important works in the field.
We have added 65 new citations to the manuscript, only 59 of which were non-self citations, especially with respect to the neurobiological, developmental, and behavioral contents of the paper and so the proportion of self-citations is now much diminished (25% instead of the earlier 35%). However, we still note that almost exclusively the only field work conducted on wild zebra finches over the past 15 years has been through the lab of Prof. Simon Griffith, our senior author. The main focus of this body of work (of which we have only cited a small fraction) has been to provide a relevant behavioral and ecological context to inform the vast amount of captive work on this species.
[Editors' note: further revisions were suggested prior to acceptance, as described below.]
Summary:
This article reviews the history of the zebra finch as a species of choice to study different aspects of developmental neurobiology and vocal communication, and particularly as a system for investigating sex differences in vocal communication. The revised manuscript is much improved, but it would still benefit of additional revisions to references, and including a discussion of studies of auditory perception using cross-fostered birds to examine innate preferences versus experience-dependent changes in auditory responses.
We now include more discussion of cross-fostering as a method to address innate vs. experience-dependent changes in auditory responses (something that I also did my postdoc research on!)
Essential revisions:
1. Please make the following modifications to references:
a) In line 37, add references to the work of the Fee and Yazaki-Sugiyama labs. These would be useful for neural mechanisms underlying sensorimotor learning (e.g. Okubo et al., 2015; Mackevicius, Happ, and Fee, 2020; Yanagihara and Yazaki-Sugiyama, 2019).
We have now added the content and the three references to the text.
b) In lines 38-39, the reference to Chen et al., 2017 does not seem to be appropriate, since that study focuses on vocal learning, not female preference. Please remove this reference and replace it with another that examines influences on female song preferences (e.g. Riebel, 2000).
This is not correct as Chen et al. 2017 does focus on experience-dependence in females’ song preferences for male song variants. We, therefore, kept this reference here: Chen Y, Clark O, Woolley SC (2017) Courtship song preferences in female zebra finches are shaped by developmental auditory experience. Proceedings of the Royal Society of London B 284: 20170054.
c) In lines 63 and/or 73, please add a reference to Cade et al., 1965, Water economy and metabolism of two estrildine finches, Physiological Zoology.
We have now added this reference to the first instance.
d) In lines 88-89, please add references to papers by Mets and Brainard (2018, 2019) that investigate the interaction of genes and social and/or auditory experience on vocal
learning and performance. Please add a short discussion about gene-environment interactions that shape song learning based on the findings of these papers in the section "Brains and genes".
Thank you, we agree that G X E interactions are essential in this section and we added the more recent citation, of these two on Bengalese finches, from eLife to support our updated content for the requested new sentences regarding Gene-by-Environment interactions in song learning in zebra finches, too.
e) In line 159 please add a reference to Price, PH (1979) Developmental determinants of structure in zebra finch song. Journal of Comparative and Physiological Psychology.
We have now incorporated this reference twice at around this section.
f) Please check the references in lines 258-260. While several papers have shown that both calls and songs have strong individual signatures that can be used for individual recognition, the references cited did not specifically examine "high levels of coordinated duetting between the male and female".
Thank you. We have now rephrased this section and included our duetting reference here, too (Elie et al. 2010).
g) In lines 323-327, please add a reference to Aronov et al. (already cited in the article) in regard to the critical role of HVC for generating stereotyped, learned vocalization in lines 323-327.
We have now added this reference here, too.
h) In lines 330-331, please add references for the idea that the AFP generates song variability in juvenile birds (Olveczky et al., 2005, 2011; or a review article, e.g. Mooney, 2009; Woolley and Kao, 2015; or Dhawale, Smith and Olveczky 2017), and discuss these findings.
Thank you, we really needed references here. We have now cited two of these reviews and discuss briefly trial-and-error learning specifically in this section.
i) Please add references to studies showing that LMAN also sends an instructive signal that can bias song and drive systematic changes in mean pitch, mean duration and syllable transition probabilities (Turner and Brainard, 2007; Andalman and Fee, 2009; Ali et al., 2013, Charlesworth et al., 2012), and discuss these findings.
We have now added to our description and discussion of the role of LMAN and AFP in general, using the Andalman and Fee citation.
j) In lines 341-344, please add a reference for deafening experiments in juvenile birds, such as Price, 1979.
We have now added Price 1979 here, too.
k) In lines 349-351, it is unclear why the Ma et al., 2020 reference is included, since the paragraph discusses mechanisms by which evaluation of song performance using auditory feedback can drive changes in song. Ma et al., 2020 report activity prior to calls by conspecifics. While predictive activity may be important for learning or maintaining song, please make clear what the activity is predicting (timing of upcoming calls? Expected auditory feedback given the motor commands?) and how it relates to learning the tutor song model.
We have deleted the Ma et al. 2020 reference and the tangential text associated with it in this paragraph.
l) In line 401, please add Xiao and Roberts, Neuron, 2018 as a reference, in addition to Kearney et al., 2019.
We have now added this prior article here, too.
m) In lines 441-444, please add a reference to Shaughnessey et al., JCN, 2018 when mentioning the song system in females as that paper argues against the idea that females possess only vestiges of the song control network and suggests that the forebrain network may have a different function in females.
Thank you for this important reference, we now rephrased this sentence to be able to cite this article.
2. Given the manuscript's emphasis on vocal imprinting, please include a brief discussion of studies of auditory perception using cross-fostered birds to examine innate preferences versus experience-dependent changes in auditory responses (e.g. Araki et al., 2017; Moore and Woolley, 2019).
We now contrast female vs. male specific findings using these references in the paragraph just prior to the Conclusions.
3. In lines 187-189, please modify your description of hair cell regeneration as "a specialized avian feat". Sensory hair cell regeneration after antibiotic treatment has been shown in other vertebrates, including fish (in the inner ear and lateral line; reviewed by Monroe et al., 2015, Front Cell Neurosci), as well as invertebrates (e.g. frogs).
Thank you; we have corrected this flaw and have now referred to two additional citations reviewing (including the one suggested) this topic in non-birds, too.
4. Please revise the section "Brains and genes for vocal learning" to either omit mention of selective auditory responses, or expand this section for clarity (lines 319-321). While many studies have shown that neurons in both the motor pathway and the anterior forebrain pathway respond selectively to playback of bird's own song in anesthetized birds, the role of such auditory responses is not entirely clear. Auditory responses in HVC and in LMAN and Area X are gated by behavioral state and are much less pronounced in awake birds (Schmidt and Konishi , 1998; Hessler and Doupe, 1999).
We have now rephrased this section, and adopted the referee’s suggestion for modifying the text and for the two additional citations included.
5. Please expand the discussion of the role of HVC in coding the timing of song (lines 323-327). For example, HVC neurons that project to RA fire once per motif while RA neurons may fire multiple bursts at specific times in song (Yu and Margoliash, 1996). Similarly, Long and Fee, 2008 is referenced, but it would help the reader to explain how the cooling experiments help to establish that HVC codes for time in song.
Thank you for these suggestions; we have now included new text and the new citation (plus one more) regarding HVC, RA, and temperature-dependent effects on song production in this section.
https://doi.org/10.7554/eLife.61849.sa2Article and author information
Author details
Funding
National Science Foundation (IOS1456524)
- Mark E Hauber
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
We thank our many colleagues working with zebra finches for productive discussions, including the editors and the referees of eLife. M Rivera and O Tchernichovski kindly provided song and call files of adult zebra finches. We dedicate our article to the memories of the late Richard Zann and Mark Konishi. FUNDING Financial support was provided by the Harley Jones Van Cleave Professorship and the National Science Foundation IOS # 1456524 (to MIML and MEH).
Publication history
- Received:
- Accepted:
- Accepted Manuscript published:
- Version of Record published:
Copyright
© 2021, Hauber et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 2,968
- views
-
- 270
- downloads
-
- 22
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 22
- citations for umbrella DOI https://doi.org/10.7554/eLife.61849
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
The Natural History of Model Organisms: Neurogenomic insights into the behavioral and vocal development of the zebra finch
eLife 10:e61849.
https://doi.org/10.7554/eLife.61849





{kind=link}
{kind=link}
{kind=link}