Discovery and characterization of Hv1-type proton channels in reef-building corals
- Departmento de Fisiología, Facultad of Medicina, Universidad Nacional Autónoma de México, Mexico
- Unidad Académica de Sistemas Arrecifales, Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, Mexico
- EvoDevo Research Group, Unidad Académica de Sistemas Arrecifales, Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, Mexico
- Department of Physiology and Biophysics, School of Medicine, Virginia Commonwealth University, United States
- Departmento of Neurociencia Cognitiva, Instituto de Fisiología Celular, Universidad Nacional Autónoma de México, Mexico
Abstract
Voltage-dependent proton-permeable channels are membrane proteins mediating a number of important physiological functions. Here we report the presence of a gene encoding Hv1 voltage-dependent, proton-permeable channels in two species of reef-building corals. We performed a characterization of their biophysical properties and found that these channels are fast-activating and modulated by the pH gradient in a distinct manner. The biophysical properties of these novel channels make them interesting model systems. We have also developed an allosteric gating model that provides mechanistic insight into the modulation of voltage-dependence by protons. This work also represents the first functional characterization of any ion channel in scleractinian corals. We discuss the implications of the presence of these channels in the membranes of coral cells in the calcification and pH-regulation processes and possible consequences of ocean acidification related to the function of these channels.
Introduction
Scleractinian or stony corals are organisms in the phylum Cnidaria that deposit calcium carbonate (CaCO3) in the form of aragonite to build an exoskeleton. Stony corals are the main calcifying organisms responsible for the construction of coral reefs, which are major ecosystems hosting numerous and diverse organisms. Coral reefs also act as natural barriers from strong ocean currents, waves, and tropical storms, providing coastal protection. This protection centers on the ability of scleractinian corals to produce enough CaCO3. The increase in atmospheric CO2 concentrations as a result of human activity poses threats to coral-reef-building organisms due to rising sea surface temperatures (Hoegh-Guldberg, 1999) and because CO2 is taken up by the ocean, dangerously lowering the pH of the sea water (Caldeira and Wickett, 2003).
It is known that precipitation of the aragonitic form of calcium carbonate is facilitated at elevated pH values, at very low concentrations of protons. Calcification by scleractinian corals is a process that has been shown to be modulated by the pH of the solution in which calcium carbonate is precipitated (Allemand et al., 2011). To this end, corals produce a specialized compartment between the ectoderm and the external substrate or skeleton called calicoblastic compartment, which contains a fluid derived from the surrounding sea water. The composition of this calicoblastic fluid or liquor is strictly regulated by the coral to maintain both an elevated pH, often close to one unit higher than the surrounding sea water, and an increased concentration of Ca2+ and carbonates. The molecular details of pH regulation in the calicoblastic fluid are not understood completely. Involvement of proton pumps has been postulated and is likely to be part of proton transport in corals. Both P-type and V-type hydrogen pumps are present in coral transcriptomes and are known to play roles in the physiology of coral-algal symbiosis (Tresguerres et al., 2017). V-type H+-ATPases have also been shown to be involved in calcification in foraminifera (Toyofuku et al., 2017). If a proton pump is involved in lowering proton concentration in the calicoblastic fluid to maintain high calcification rates, protons will be transported to the cytoplasm of the ectodermal cells that constitute the calicoblastic epithelium, producing a profound acidification of the cytoplasmic pH (pHi). Although measurements of the pHi in corals indicate values of 7.13–7.4 (Venn et al., 2009), it is unknown how coral cells regulate pHi. Thus, an efficient pH-regulatory mechanism is to be expected to be present in corals. We hypothesized that proton channels might be fundamental to this physiological process and also required for calcification in hard corals.
Although a number of studies have delineated the physiological roles of Hv1 voltage-gated proton channels in vertebrate cells (DeCoursey, 2013), less is known about their role in invertebrates. These channels are potential mediators in processes that are critically dependent on proton homeostasis. As an example, they have been shown to be involved in regulating the synthesis of the calcium carbonate skeleton in coccolithophores, calcifying unicellular phytoplankton (Taylor et al., 2011).
The range of voltages over which channel activation occurs is strongly modulated by the transmembrane proton gradient, characterized by ΔpH = pHo-pHi, that is, the difference between the external and internal pH. In the majority of known Hv1 channels, the voltage at which half of the channels are activated, the V0.5 or the apparent threshold for channel opening (VThr), shifts by roughly 40 mV per unit of ΔpH. Thus, the pH gradient strongly biases the voltage-independent free energy of channel activation (Cherny et al., 1995). With few exceptions, channel activation occurs at voltages that are more positive than the reversal potential for protons, implying that protons are always flowing outward under steady-state conditions. The fact that most Hv1s mediate outward currents is the reason these channels are mostly involved in reversing intracellular acidification or producing voltage-dependent cytoplasmic alkalization (Lishko and Kirichok, 2010; DeCoursey, 2013).
Here we report the presence of genes encoding Hv1 channels in two species of reef-building corals. We cloned and characterized the biophysical properties of these channels in an expression system using patch-clamp electrophysiology. The demonstration of the presence of voltage-gated proton channels in corals is an initial step to a deeper understanding of coral calcification and its dysregulation under ocean acidification conditions. We show that some of the coral Hv1’s biophysical properties are different from other known proton channels, and this behavior makes them interesting models to try to understand some basic biophysical mechanisms in these channels. To explain this behavior, we developed a novel activation model to describe voltage- and pH-dependent gating that has general applicability to Hv1 channels.
Results
Ion channels have not been characterized in corals. Here, we have initiated their study by searching the transcriptome of the Indo-Pacific coral Acropora millepora (Moya et al., 2012) for sequences coding for putative voltage-sensing residues present in canonical Hv1 channels with the form RxxRxxRIx, which corresponds to the S4 segment of Hv1 channels and is also found in other voltage-sensitive membrane proteins. Blast searches detected four sequences that seem to correspond to a gene encoding the Hv1 voltage-activated proton-selective ion channel (Ramsey et al., 2006; Sasaki et al., 2006). A. millepora is one of the most widely studied species of scleractinian corals and is well represented in the commercial coral trade (Cleves et al., 2018; Ying et al., 2019). We proceeded to clone this gene from a small specimen of A. millepora obtained from a local aquarium (Reef Services, Mexico City). As indicated in the 'Materials and methods' section, total RNA was extracted from tissue and mRNA was retrotranscribed to obtain complementary DNA (cDNA). We managed to obtain a full-length clone and refer to this sequence as AmHv1 or Hv1-type proton channel of A. millepora.
We were interested in knowing if the same gene is present in a closely related species from the Caribbean Sea. Thus, we used the same primers to clone the Hv1 channel from Acropora palmata, a widespread coral in the same family, and which we call ApHv1. The amino acid sequence is almost identical to AmHv1 (Figure 1—figure supplement 1A); the greatest divergence is found between a few amino acid residues in the C-terminal region. This result suggests that despite the large biogeographic difference, these two genes have not diverged significantly. The ApHv1 sequence also gives rise to fast-activating voltage-gated proton currents (Figure 1—figure supplement 1B).
The most diagnostic feature of the Hv1 protein is the sequence of the fourth transmembrane domain or S4, which contains three charged amino acids in a characteristic triplet repeat. The presence of these repeats in our sequence allowed us to initially identify our clone as an Hv1 channel. However, we decided to compare our sequence to those of several Hv1 orthologs. We selected a list of 130 Hv1 protein sequences that are well curated in the Gene Bank (https://www.ncbi.nlm.nih.gov/), representing several branches of the eukaryotes, from unicellular plants to mammals. As expected, the protein sequence of AmHv1 has similarity to several other Hv1 genes from varied organisms (Figure 1A). The identity varies from 98%, when compared to other putative coral and anemone sequences, to less than 30%, when compared to plant and nematode sequences. In spite of this variability, the putative transmembrane domains of all these proteins show high conservation, and consensus sequence logos can detect the presence of highly conserved individual amino acid sequences that can be considered characteristic of Hv1 channels. Figure 1B compares these transmembrane domain consensus logos with our AmHv1 sequence. It can be gleaned that AmHv1 contains the highly conserved residues that form the voltage-sensing amino acid residues in S4 as well as their acidic pairs present in S2 and S3. The extracellular histidine residues involved in Zn2+ coordination are also present. These results suggest that our sequence is that of a bona fide Hv1 voltage-sensing domain (VSD).
Figure 1 with 2 supplements see all
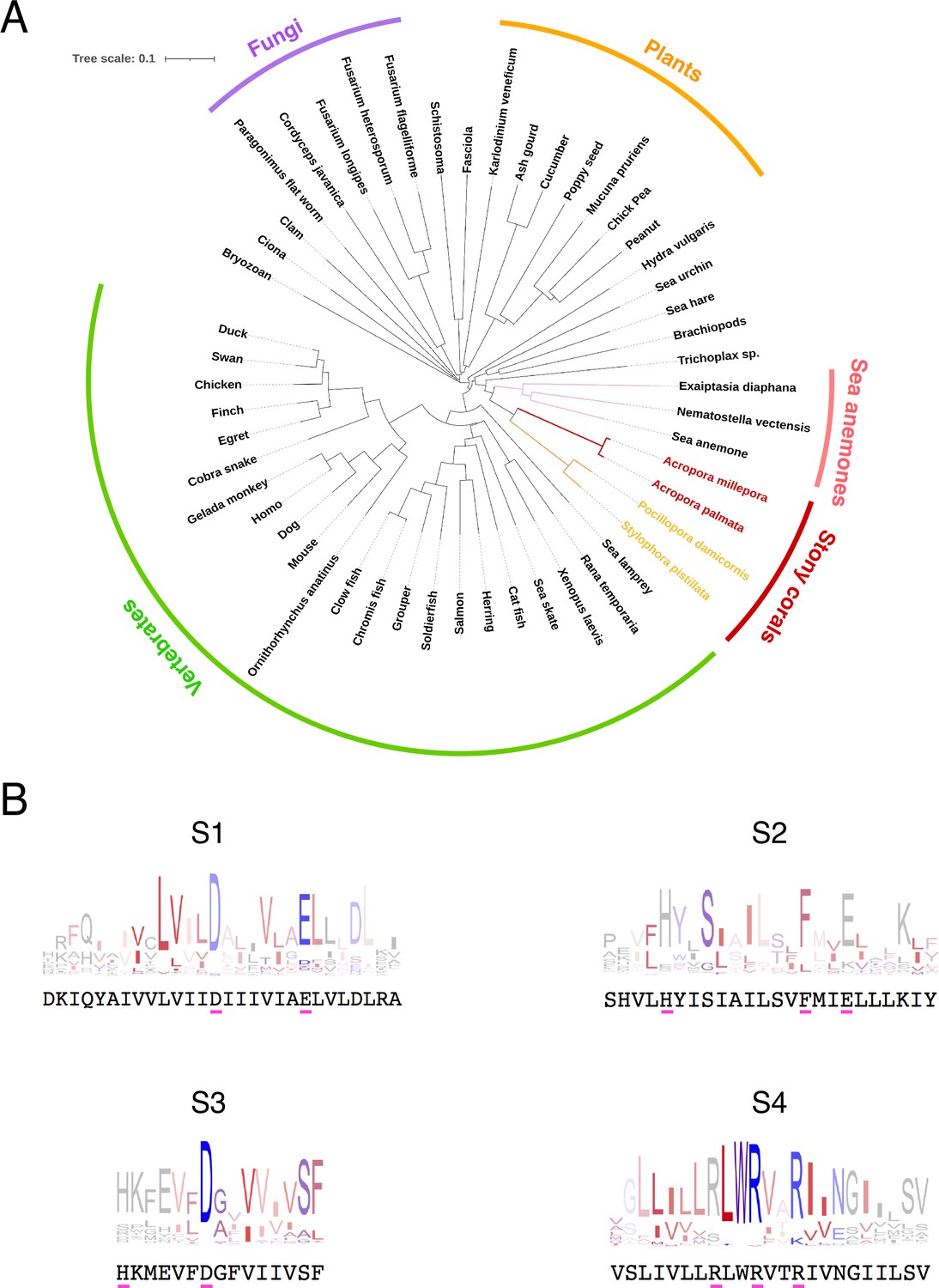
Conservation and phylogenetic relationships of Hv1 channels.
(A) The tree obtained from a multiple sequence alignment from Hv1 channels in CLUSTAL-O. Highlighted in red and yellow are the branches containing coral Hv1 sequences. (B) Consensus logo sequences of transmembrane domains of Hv1 channels. The color code indicates the hydrophobicity of each residue, where blue indicates charged residues, red indicates non-polar residues, and other colors indicate either non-polar or charged residues with less conservation.
-
Figure 1—source code 1
Code for generating the tree in Figure 1.
- https://cdn.elifesciences.org/articles/69248/elife-69248-fig1-code1-v1.xlsx.zip
Apart from canonical voltage-gated channels, several other proteins contain VSDs. Examples are the voltage-sensing phosphatases like VSPs (Iwasaki et al., 2008) and TPTE and TPTE2 (Halaszovich et al., 2012) proteins (transmembrane proteins with tensin homology) and genes like TMEM266. These proteins are relevant to us since some TPTEs have been shown to also mediate proton currents and TMEM266 can be modulated by Zn2+ (Papp et al., 2019). We compared the sequence of AmHv1 with several orthologs of TPTEs and TMEM266. Although there is some similarity within transmembrane domains (Figure 1—figure supplement 2), the overall sequence comparison shows that AmHv1 and these VSD-containing proteins are different.
As mentioned before, we performed a multiple sequence alignment with 130 Hv1 sequences. In Figure 2, we show the detailed sequence alignment of AmHv1 with five of these sequences, which represent some of the best studied Hv1 genes. It can be seen that there is a high degree of identity, especially in the transmembrane domains. The least degree of conservation appears when comparing this sequence to the dinoflagellate Karlodinium veneficum Hv1 channel (Figure 2A). A search of available transcriptomes from several coral species allowed us to detect the presence of sequences that are found in Hv1 channels. This suggests that Hv1 proton channels might be found in many families of scleractinian corals (Figure 2—figure supplement 1), as has also been recently shown (Capasso et al., 2021).
Figure 2 with 1 supplement see all
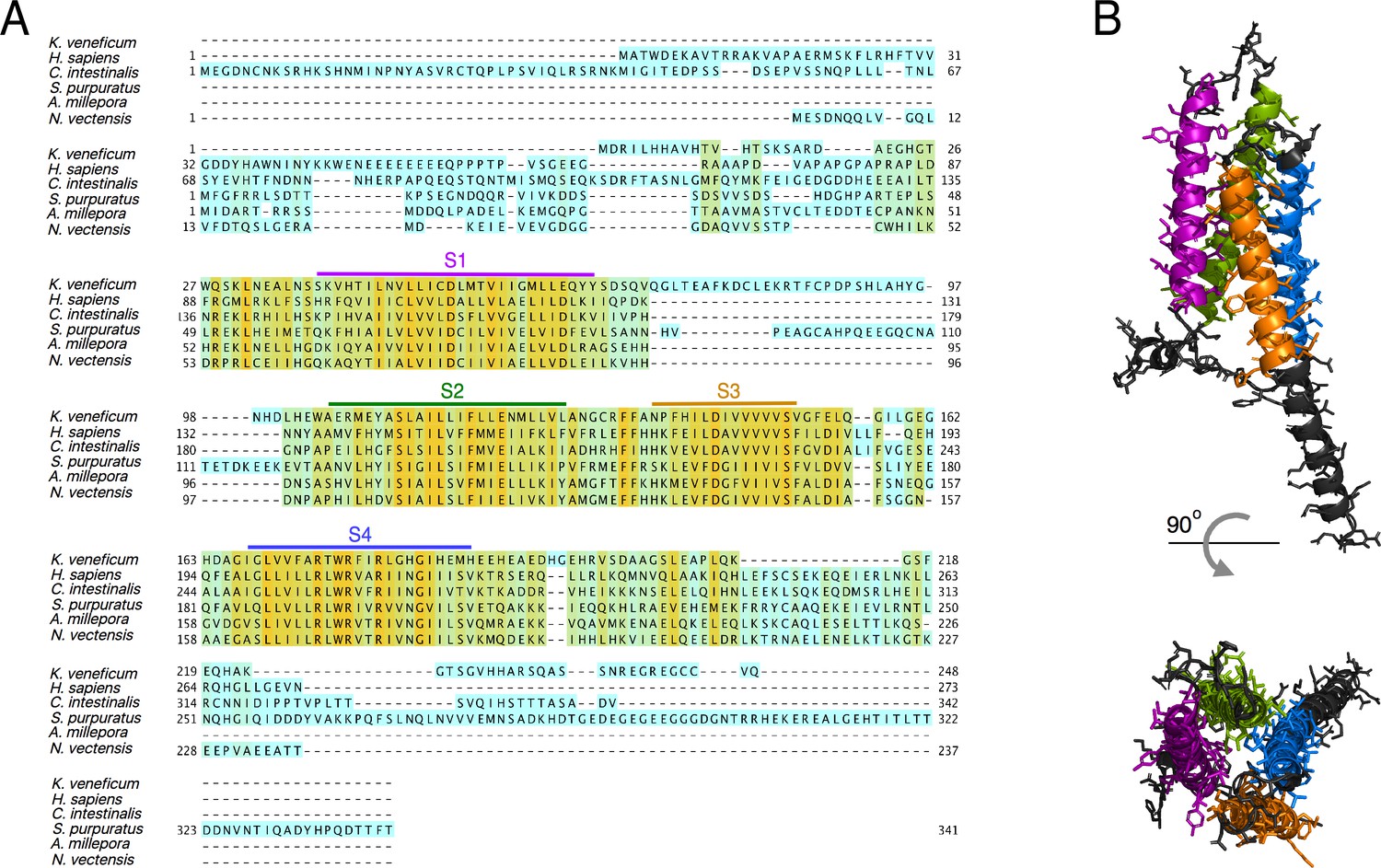
Protein sequence alignment of the AmHv1 channel with selected Hv1s from other organisms.
(A) Amino acid sequence alignment of Acropora millepora Hv1 (AmHv1) with other known Hv1 orthologs provided by the CLUSTAL-O algorithm. The predicted transmembrane domains are shown by the colored horizontal lines and letters. The colors highlighting the sequence indicate sequence identity. Orange indicates identical amino acids, and cyan indicates no identity. (B) Predicted structural topology of AmHv1. Transmembrane domains are colored to correspond with the sequences in (A). The top panel is the view parallel to the membrane while the bottom panel is the view from the top (extracellular) side.
Secondary-structure prediction suggests that AmHv1 is a canonical Hv1 channel formed by a VSD with four transmembrane segments. The protein sequence was used for 3D modeling using the SWISS MODEL server (Waterhouse et al., 2018), which produced models based on the Hv1 chimera structure (Takeshita et al., 2014) and the Kv1.2 potassium channel VSD (Long et al., 2005). This structural model is shown in Figure 2B. The predicted model indicates a shortened N-terminal region, four transmembrane helices, and a long C-terminal helix.
Voltage-gated proton channels from Ciona (Sasaki et al., 2006) and humans (Lee et al., 2008) have been shown to express as dimers in the plasma membrane, and this dimeric form is understood to be the functional unit of these proton channels. The dimer is stabilized by a coiled-coil interaction mediated by an alpha helical C-terminal domain. As shown by the model in Figure 2B, AmHv1 has a long C-terminal helix, which is predicted to engage in a coiled coil (Paircoil2; McDonnell et al., 2006). We calculated the probability per residue to form a coiled coil for all the C-terminal residues, both for human and AmHv1 channels, using the program COILS (Lupas et al., 1991). Figure 3A shows that the coiled-coil probability for AmHv1 C-terminus is at least as high or higher than that for hHv1, an established dimer, strongly suggesting that coral Hv1s might also form dimers.
Figure 3
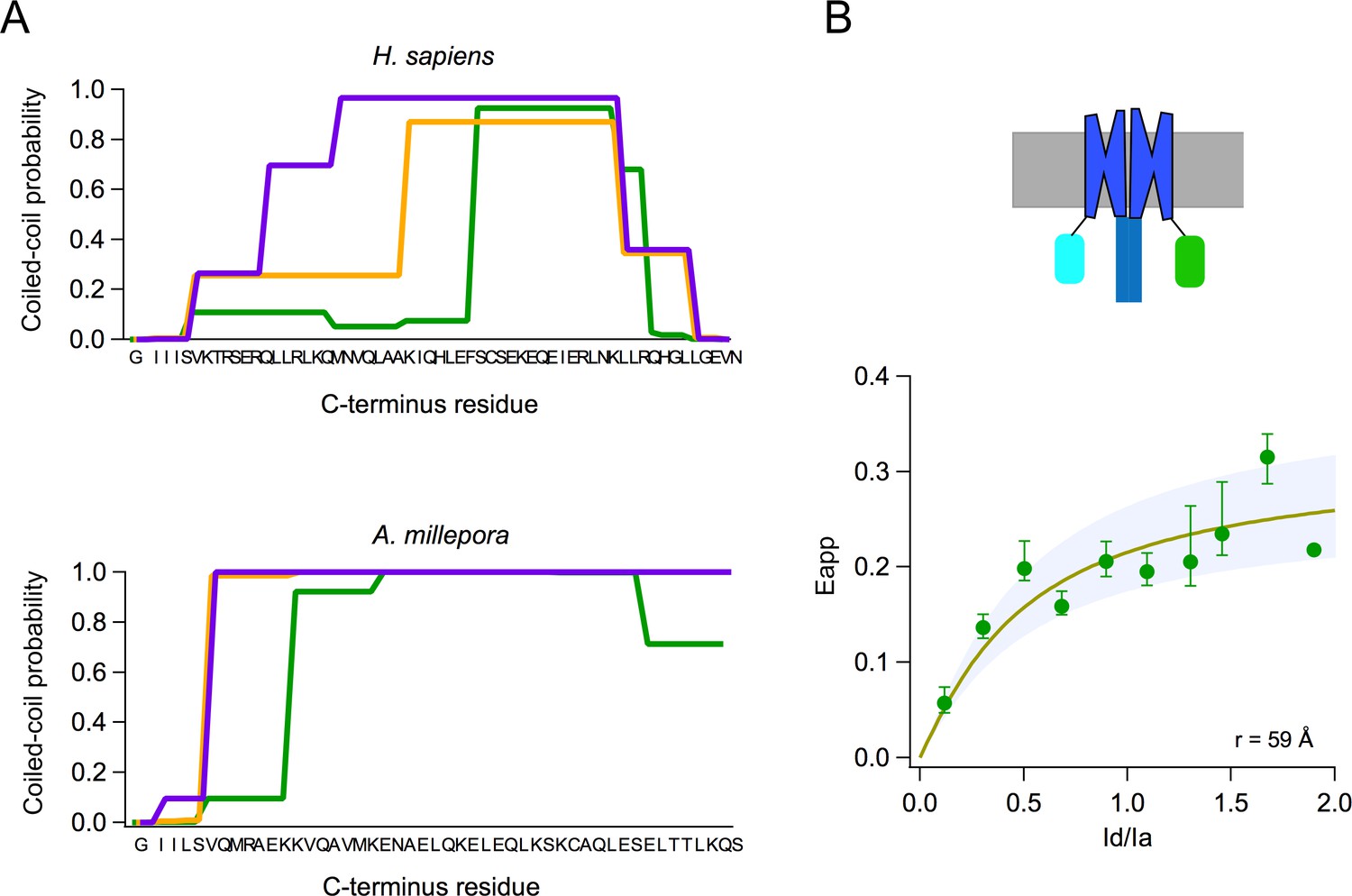
Subunits of the AmHv1 channel associate to form dimers.
(A) Probability of coiled-coil formation per amino acid residue of the C-terminus domain of hHv1 (top) and Acropora millepora Hv1 (AmHv1) (bottom). The different colors correspond to the three seven-residue windows used by the program to calculate the score. The sequence of the C-terminus is shown in the x-axis. (B) FRET measurement of dimer formation. The apparent FRET measured from 134 cells is plotted as a function of the ratio of donor to acceptor fluorescence (Id/Ia). Shown is the average and sem for data in Id/Ia windows of 0.1. The continuous curve is the fit of the data to the prediction of a model that considers random assembly of donor- and acceptor-tagged subunits into a dimer. The separation between the FRET pairs in a dimer is ~60 Å, according to the model. The upper panel depicts a cartoon of the presumed fluorescent protein (FP)-tagged dimer in the membrane.
-
Figure 3—source data 1
Source data for Figure 3.
- https://cdn.elifesciences.org/articles/69248/elife-69248-fig3-data1-v1.xlsx
In order to study the oligomeric state of the coral Hv1, we performed FRET experiments with the AmHv1 channel tagged with fluorescent proteins (FPs) as a FRET pair. Figure 3B shows that there is significant FRET efficiency between FP-tagged subunits, indicating a very close interaction between monomers. The measured apparent FRET efficiency vs the fluorescence intensity ratio can be fitted to a model where the subunits assemble as a dimer. From this fit, we can estimate a distance between fluorophores of ~60 Å, which is compatible with AmHv1 being a dimer, at least in HEK293 cells.
Functional expression of AmHv1 voltage-dependence and kinetics
The cDNA of AmHv1 was cloned in the pcDNA3 expression vector and transfected into HEK293 cells. Under whole-cell conditions, we recorded large voltage-dependent outward currents. Figure 4A shows a family of such currents. The data suggest that these currents were carried mostly by protons, since the reversal potential, measured from a tail current protocol, closely followed the equilibrium potential for protons, as given by the Nernst equation (Figure 4B).
Figure 4

Proton currents mediated by AmHv1 expressed in HEK293 cells.
(A) A typical proton current family elicited by depolarizing pulses from −50 to 60 mV in 10 mV intervals. The duration of the pulses is 500 ms. Linear current components have been subtracted. (B) Reversal potential of currents as a function of the pH gradient. Symbols are individual data and the black horizontal lines are the mean. The dotted line is the expected reversal potential as predicted by the Nernst equation. The inset shows a tail current family from which instantaneous IV curves were extracted to measure the reversal potential. Recordings shown in (A) and (B) were obtained in the whole-cell configuration. (C) Normalized conductance-voltage curve at ∆pH = 1. The purple curve is the fit to Equation 1 with parameters V0.5 = 7.85 mV and q = 2.09 eo. Circles are the mean and error bars are the sem (n = 7). (D) Kinetic parameters of activation. Activation time constant and delay estimated from fits of current traces to Equation 2. Circles are the mean, and the sem is indicated by the shaded areas (n = 6). The voltage-dependence of the delay and tau of activation were estimated from a fit to Equation 3, which appears as the red curve. Parameters are δ(0) = 98.2 ms and qδ = 0.47 eo. The voltage-dependence parameters for tau are τ(0) = 212 ms and qτ = 0.37 eo.
-
Figure 4—source data 1
Source data for Figure 4.
- https://cdn.elifesciences.org/articles/69248/elife-69248-fig4-data1-v1.xlsx
The voltage-dependence of channel gating was estimated from a fit of the normalized conductance vs voltage (G-V) to Equation 1. The steepness of the curve corresponds to an apparent charge of ~2 eo, comparable to other Hv1s under similar recording conditions (Figure 4C).
Interestingly, these channels seem to activate rapidly. This is apparent from the current traces, which approach a steady state within a few hundred ms (Figure 4A), as quantified in Figure 4D. Equation 3 estimates two parameters, an activation time constant (τ) and a delay (δ). Both the time constant and the delay are similarly voltage-dependent at positive potentials. The existence of a delay in the time course implies that activation is a multiple-state process. The delay magnitude is smaller than the time constant at all voltages, which can be interpreted to mean that the rate-limiting step for opening comes late in the activation pathway (Schoppa and Sigworth, 1998).
Comparison to human Hv1 channel properties
Human Hv1 is probably the best characterized of the voltage-gated proton channels (Musset et al., 2008); so we compared some of the properties of AmHv1 with hHv1. AmHv1 channels activate faster than their human counterpart. Figure 5 compares the activation kinetics of these two channels under the same conditions. Steady state is apparently reached sooner after a voltage pulse in AmHv1 (Figure 5A) when compared to hHv1 (Figure 5C). The slower kinetics of the human ortholog is also evidenced in the more sluggish deactivation tail currents (Figure 5C). The range of voltages over which activation happens is also different between the two channels, with the coral Hv1 channel activating at more negative voltages than the human clone (Figure 5E; notice that the proton gradient is such that ΔpH = 1 and is the same for recordings of both channel types). Even though AmHv1 activates at more negative voltages, the activation range is still more positive than the proton reversal potential; thus, coral proton currents activated by depolarization, in the steady state and at least as expressed in HEK293 cells, are always outward.
Figure 5
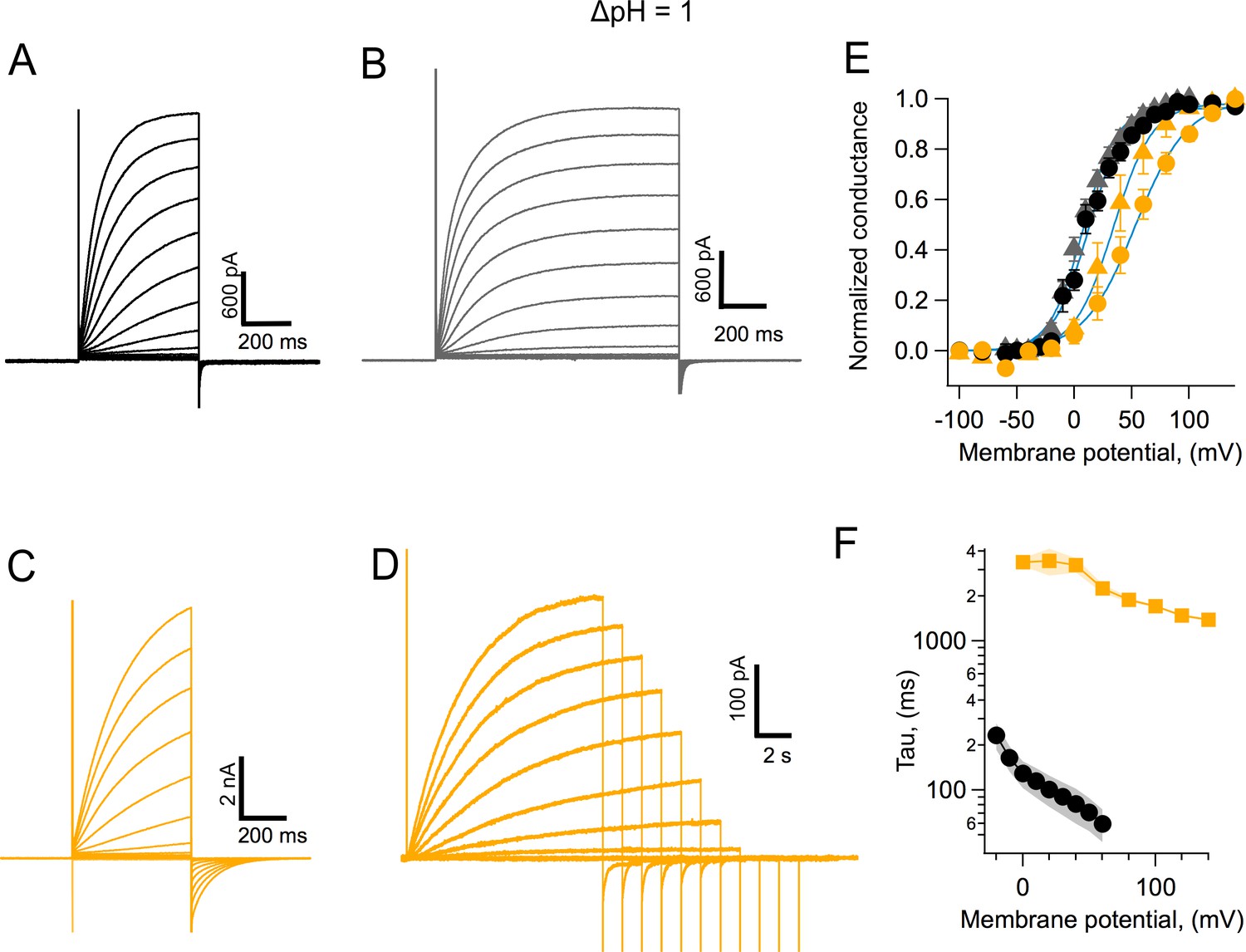
Coral Hv1 channels are faster and activate more readily than their human counterpart.
(A) Acropora millepora Hv1 (AmHv1) currents in response to voltage-clamp pulses from −100 to 120 mV and of 500 ms duration. (B) AmHv1 currents in response to the same voltage-clamp pulses as in (A) but of a longer duration of 800 ms. (C) Currents through hHv1 channels in response to voltage-clamp pulses from −100 to 120 mV of 500 ms duration, compared with (A). (D) hHv1 currents recorded with long pulses designed to reach the steady state. Pulses were shortened in duration as depolarizations became larger, in an effort to reduce intracellular proton depletion. Pulses are from −100 to 140 mV. Recordings shown in (A), (B), (C), and (D) were obtained in the whole-cell configuration. (E) Comparison of the conductance-voltage relationship for both channels for short- (circles) and long (triangles)-duration pulses. Black symbols are the mean G/Gmax for AmHv1 and yellow symbols for hHv1. The error bars are the sem (n = 3, for short pulses, both channels, and n = 4, for long pulses, both channels). The continuous blue curves are fits to Equation 1. The fitted parameters are AmHv1, short pulses, q = 1.62 eo, V0.5 = 12.2 mV; AmHv1, long pulses, q = 1.7 eo, V0.5 = 7.7 mV; hHv1, short pulses, q = 1.11 eo, V0.5 = 53.1 mV; hHv1, long pulses, q = 1.47 eo, V0.5 = 34.1 mV. (F) The activation time constant estimated from fits of currents elicited by long pulses to Equation 2. Squares are the mean for hHv1 and circles, for AmHv1. The shaded areas are the sem (n = 4, for both channels).
-
Figure 5—source data 1
Source data for Figure 5.
- https://cdn.elifesciences.org/articles/69248/elife-69248-fig5-data1-v1.xlsx
In order to better estimate both kinetics and activation, we performed experiments with longer pulse durations. This is especially important for the very slow activation of the human channel. The resulting currents are shown in Figure 5B and D. The normalized conductance for these currents that are closer to steady state are shown in Figure 5E by triangles. These G-V curves are shifted to more negative voltages than the G-V from shorter pulses, as expected. The faster kinetics of AmHv1 is clearly evidenced when the time constant of activation, τ, estimated using fits of the activation time course to Equation 2, is compared for coral and human Hv1 channels. AmHv1 is more than tenfold faster at 0 mV and over a range of positive voltages (Figure 5F).
Effects of the pH gradient on gating
Both native and cloned voltage-gated proton channels are characteristically modulated by the pH gradient (Cherny et al., 1995; Sasaki et al., 2006; Ramsey et al., 2006). We carried out experiments to investigate the modulation of the coral Hv1 channels by different pH gradients. We first recorded whole-cell currents at various ΔpH values and estimated the voltage-dependence of the conductance. These G-V curves were fitted to Equation 2 to obtain the voltage of half activation, V0.5, and apparent gating charge, q, which determines the steepness of the fit. As is the case with other Hv1 channels, the V0.5 shifts to negative voltages when ΔpH is greater than 0 and to positive voltages when ΔpH is less than 0 (Figure 6A). When we plot the V0.5 as a function of ΔpH, the relationship seems to be mostly linear over the range of ΔpH −1 to 2. This relationship is somewhat steeper than the generally observed −40 mV/ΔpH (Figure 6B). We tried to obtain recordings over an extended range of ΔpH values. To this end, we performed inside-out recordings in which the composition of solutions can be better controlled, tends to be more stable, and the size of currents is smaller. However, recordings were unstable at extreme pH values, and we only managed to reliably extend the data to a ΔpH value of −2. Figure 6C shows the summary of the inside-out recordings. We have plotted both the V0.5 and the threshold voltage, VThr. To obtain this last parameter, we fitted the exponential rise of the G-V curve to a function of the form
Figure 6 with 2 supplements see all
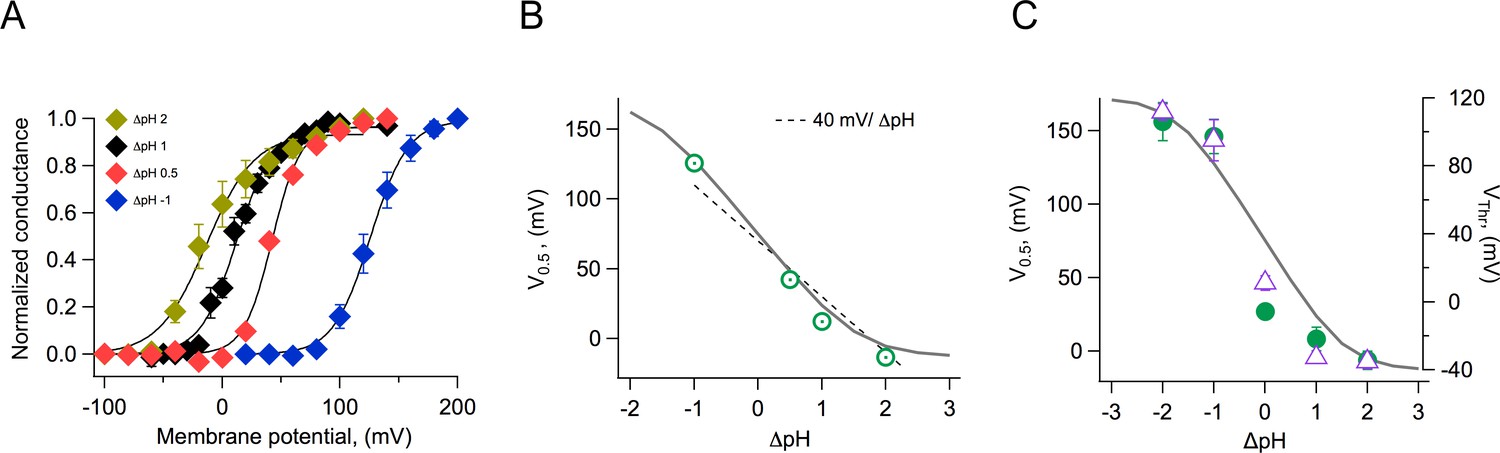
Modulation of channel activation by the pH gradient.
(A) Conductance vs voltage relationships obtained at the indicated ΔpH values, from whole-cell recordings of Acropora millepora Hv1 (AmHv1) proton currents. Continuous lines are fits to Equation 1. (B) The parameter V0.5 was obtained from the fits in (A) and is displayed as a function of ΔpH. The dotted line is the 40 mV/ΔpH linear relationship. The continuous gray curve is the prediction of the allosteric model (Figure 7). (C) Parameters V0.5 (green circles) and VThr (purple triangles) obtained from a different set of inside-out current recordings. Data are mean ± sem. The continuous gray curve is the same prediction of the allosteric model (Figure 7) that is shown in panel (B). The model parameters used to generate the theoretical curve are E = 5x105, D = 105, C = 0.0002, Kv(0) = 0.00005, qg = 1.0 eo, pKo = 3.4, and pKi = 7.
-
Figure 6—source data 1
Source data for Figure 6.
- https://cdn.elifesciences.org/articles/69248/elife-69248-fig6-data1-v1.xlsx
VThr was calculated as the voltage at which the fit reaches 10% of the maximum conductance. The parameter VThr should be less sensitive than V0.5 to the possible change in the proton gradient that can occur with large currents. It is clear from these data that at extreme values, the dependence of V0.5 or VThr on ΔpH deviates from a simple linear relationship and instead appears to saturate at extreme values of ΔpH.
Allosteric model of voltage- and pH-dependent gating
Currently, there is only one quantitative model that has been used to explain ΔpH gating of Hv1 channels (Cherny et al., 1995). However, this model is euristic and does not provide mechanistic insight into the process of proton modulation of the voltage dependence of proton-permeable channels. In order to explain the modulation of the range of activation by the proton gradient, parameterized by the V0.5, we developed a structurally inspired allosteric model of voltage and proton activation. As many voltage-sensing domains, Hv1 has two water-occupied cavities exposed to the extracellular and intracellular media (Ramsey et al., 2010; Islas and Sigworth, 2001; Ahern and Horn, 2005). Recent evidence suggests that these cavities function as proton-binding sites through networks of electrostatic interactions (De La Rosa et al., 2018). In our model, we propose that these two proton-binding sites, one intracellular and one extracellular, allosterically modulate the movement of the voltage-sensing S4 segment and, thus, channel activation in opposite ways. The extracellular site is postulated as inhibitory, while the intracellular site is excitatory, facilitating voltage sensor movement. As a first approximation, we employed a simplified allosteric formalism based on a Monod-Wyman-Changeux (MWC) style model (Horrigan and Aldrich, 2002; Changeux, 2012). As a simplifying assumption, in this model we assume that the voltage sensor moves in a single voltage-dependent activation step. We assume the external and internal proton-binding sites have simple protonation given by a single pKa value. These sites operate as two allosteric modules and are coupled to the voltage sensor according to coupling factors C and D, respectively. These binding sites in turn interact with each other through the coupling factor E. The modular representations of the model are illustrated in Figure 7A, while the full model depicting all open and closed states with all permissible transitions and the corresponding equilibrium constants for each transition is shown in Figure 7B. Full details of equations derived from these schemes are given in supplementary data.
Figure 7

Gating scheme I.
(A) Modular representation of a simple Monod-Wyman-Changeux (MWC) model; the channel opening transition is voltage-dependent, with equilibrium constant K(V). Bo and Bi are the unbound states of the extracellular and intracellular proton-binding sites, respectively, and BoHo and BiHi are the proton-bound states of these binding sites. Qo and Qi are equilibrium constants that depend on the pKa of each of these binding states. C, D, and E are the coupling constants between each of the indicated modules. (B) All the individual states implied in (A) are depicted, along with proton-binding states and the appropriate equilibrium constants. C, closed states, O, open states. OHx, OHxHx and CHx, CHxHx are single or double proton-occupied states, where x can be o for outside or i for inside-facing binding sites.
This allosteric model represents the first attempt at producing a quantitative mechanistic understanding of the interaction of the voltage sensor and protons in Hv1 channels.
From the data shown in Figure 6C, it can be seen that the model is capable of reproducing the very steep dependence of V0.5 on ΔpH and importantly, the saturation of this relationship at extreme values. Some Hv1 channels from other organisms show a linear dependence of gating over a large range of ΔpH values, while others show a reduced dependence and even saturation over some range of ΔpH (Thomas et al., 2018). Our model can explain these different behaviors as different channels having distinct values of pKas for the internal or external sites, differences in coupling factors, or differences in the voltage-dependent parameters (Figure 6—figure supplements 1 and 2).
Block by Zn2+
The best-characterized blocker of proton channels is the divalent ion zinc (Cherny et al., 2020; De La Rosa et al., 2018; Qiu et al., 2016). We performed experiments to determine if AmHv1 channels are also inhibited by zinc. We found that indeed, extracellular application of zinc in outside-out patches produced inhibition of the channels, reflected in reduced current amplitudes (Figure 8A). Figure 8B shows average current-voltage (I-V) relationships in the absence and presence of 10 μM external zinc. It can be seen that the fraction of current blocked is not the same at every voltage, indicating that this inhibition might be voltage-dependent. The fraction of blocked channels was calculated and is plotted at each voltage along with the I-V curves (Figure 8B). It can be clearly seen that inhibition by Zn2+ is voltage-dependent. A simple mechanism for voltage-dependent blockage was proposed by Woodhull, 1973. This model postulates that a charged blocker molecule interacts with a binding site in the target molecule that is located within the electric field. Fitting the data according to this model, and given that zinc is a divalent ion, its apparent binding site is located at a fraction δ = 0.2 of the membrane electric field from the extracellular side (Figure 8B).
Figure 8
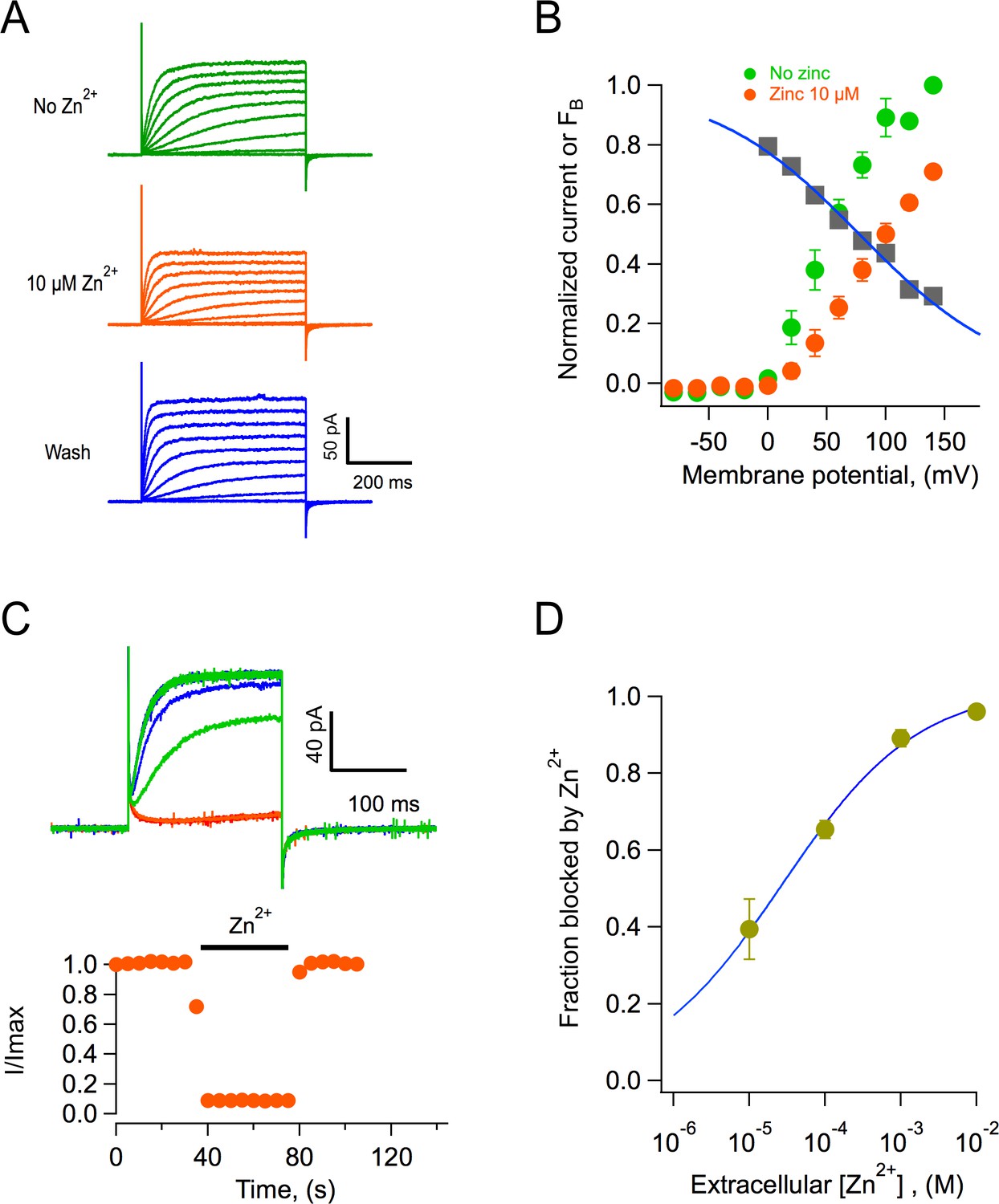
Block of AmHv1 channels by extracellular zinc.
(A) Acropora millepora Hv1 (AmHv1)-mediated currents from an outside-out patch in the absence (top) and presence of 10 μM zinc (middle) and after washing of zinc (bottom). The scale bars apply to the three current families. Voltage pulses lasted 500 ms and were from −80 to 140 mV in 20 mV steps. The ΔpH = 1 with pHi = 6 and pHo = 7. (B) Normalized current-voltage relationships before and in the presence of 10 μM zinc from four patches as in (A). The gray squares are the ratio Izinc(V)/I(V), which gives the voltage-dependence of the blocking reaction. The blue curve is the fit to the Woodhull equation , where FB is the fraction of the current blocked, δ is the fraction of the electric field where the blocker binds, z is the valence of the blocker, V0.5 is the potential where half of the current is blocked, KB is Boltzmann’s constant, and T is the temperature in Kelvin. The fitting parameters are δ = 0.19, V0.5 = 77.6 mV. (C) The effect of zinc is fast. Application of 1 mM zinc to an outside-out patch produces almost instantaneous block of ~90% of the current. The effect also washes off quickly upon removal of zinc. Trace colors are as in (A). Voltage pulse was 100 mV applied every 5 s. (D) Dose-response curve of zinc block of AmHv1 obtained at 100 mV. The continuous curve is a fit of the data to Equation 4 with apparent KD = 27.4 μM and n = 0.48.
-
Figure 8—source data 1
Source data for Figure 8.
- https://cdn.elifesciences.org/articles/69248/elife-69248-fig8-data1-v1.xlsx
Zinc blockage proceeds very fast. At 1 mM, the channels are blocked almost instantaneously, and the inhibition washes off very fast as well (Figure 8C). Finally, we report the dose-response curve (Figure 8D). The inhibition dose-response curve can be fit by a Hill equation (Equation 4) with a slope factor of nearly 0.5 and an apparent dissociation constant, KD, of 27 μM.
Discussion
A few ion transport mechanisms in reef-building corals have been described, but up to now, no ion channels have been characterized from any scleractinian species. Here we have shown that voltage-gated proton-permeable channels formed by the Hv1 protein are present in corals. In particular, we have cloned these channels from two species of the genus Acropora, A. millepora and A. palmata. It is interesting that the protein sequence of these proteins shows a very high degree of conservation, suggesting that, even when the two species are found in different oceans, they haven’t had time to diverge substantially or alternatively; selective pressures on these channels are very similar in both species. The presence of Hv1 sequences in many other species of corals from disparate clades suggests that Hv1 plays an important role in coral physiology.
Hv1 channels are formed by a protein fold that is structurally equivalent to the VSDs of canonical voltage-gated channels (Sasaki et al., 2006; Ramsey et al., 2006). The VSD is formed by a bundle of four antiparallel alpha helices (Takeshita et al., 2014). In some species, it has been shown that Hv1 channels are dimeric (Lee et al., 2008; Mony et al., 2020; Lee et al., 2008). Accordingly, we have also shown here that the AmHv1 is a dimer. Our FRET results are consistent with the high propensity to form a coiled coil shown by its C-terminal domain.
Hv1 channels are different from canonical voltage-gated channels in that both voltage sensing and permeation are mediated through a single protein domain. Voltage sensing is thought to occur through the interaction of charged amino acid side chains with the electric field, leading to the outward movement of the fourth domain or S4, in a similar fashion to other voltage-sensing domains (Carmona et al., 2018; De La Rosa and Ramsey, 2018). This outward movement of the S4 is coupled to protons moving through the VSD in a manner that is not completely understood (Randolph et al., 2016). Most proton-permeable channels seem to have evolved to extrude protons from the cell, and toward this end, their voltage dependence is tightly modulated by the proton gradient between extracellular and intracellular solutions (Cherny et al., 1995).
Our electrophysiology experiments show that these coral channels give rise to proton currents when expressed in HEK293 cells and that they retain the functional characteristics that have been shown to define the class in other species, such as very high selectivity for protons, activation by voltage, and modulation of this activation by the proton gradient. The new channels reported here activate faster than the human Hv1 channel. It has been known that different orthologs of Hv1 activate with varying kinetics. For example, sea urchin, dinoflagellate, and recently, fungal Hv1 channels activate rapidly, while most mammalian counterparts have slow activation rates (Musset et al., 2008; Smith et al., 2011; Zhao and Tombola, 2021). A comparative study suggests that two amino acids in the S3 transmembrane segment are important determinants of kinetic differences between sea urchin and mouse Hv1 (Sakata et al., 2016). The authors suggest that the time course of activation is slow in channels containing a histidine and a phenylalanine at positions 164 and 166, respectively (mouse sequence numbering). The AmHv1 has a histidine at an equivalent position, 132, and a methionine at 134. It is possible that this last amino acid in AmHv1 confers most of the fast kinetics phenotype. A separate work has shown that a lack of the amino-terminal segment in human sperm Hv1 also produced fast-activating channels (Berger et al., 2017). Interestingly, the Acropora channels have a shorter amino-terminal sequence, which could also contribute to their fast kinetics.
One of the most interesting characteristics found in these new proton channels is their modulation by the proton gradient. As opposed to other Hv1 channels, we can observe a trend toward saturation of the V0.5 for activation as a function of ΔpH at extreme values of this variable. A tendency toward saturation of the V0.5-ΔpH relationship has been observed in mutants of the hHv1 channel (Cherny et al., 2015) or at negative values of ΔpH for a snail Hv1 (Thomas et al., 2018), but it seems it can be fully appreciated in AmHv1. Since our model explains the observation of saturation of voltage gating at extreme values of ΔpH as a consequence of the existence of two saturable sites for proton binding, we attribute this behavior to the large separation of pKa values for the extracellular and intracellular proton-binding sites. Thus, channels that do not show saturation might have sites with well-separated and differing pKa values. Evolutionary fine tuning of these pKa values will produce channels with pH activation in ranges adapted to their physiological function.
It is important to point out that the external and internal proton-binding sites can be part of the voltage sensor itself. We envision these sites as being formed not by distinct protonatable amino acids but by a network of electrostatic interactions between amino acids in S4 and other transmembrane segments. In this context, the pKa values of the sites in our model do not reflect pKas of individual amino acids but of the whole proton-binding site. Recent works by Carmona et al., 2021 and Schladt and Berger, 2020 suggest that voltage sensing is directly responsible for pH gating. These authors suggest that the energy of the pH gradient is part of the free energy needed for voltage-sensor movement. This purely energetic statement is encompassed in our model as the coupling factors between voltage-sensor movement and proton binding, which are in fact energetic factors.
In our model, the strength of allosteric coupling of these sites and the voltage sensor determines if saturation is observed over a short or extended range of ΔpH values and the range of values of V0.5 that a particular channel can visit. Coupling also determines the value of the slope of the ΔpH vs V0.5 curve. Values larger or smaller than the typical ~40 mV/pH are obtained as a consequence of strong or weak coupling between binding and voltage-sensor movement, respectively (Figure 6—figure supplement 2). Our model should provide a framework to better understand the gating mechanisms in future work.
It is clear that more complicated models, with a larger number of voltage-dependent and -independent steps (Villalba-Galea, 2014) and coupling to protonation sites, should be the next step to improve data fitting and explore voltage- and proton-dependent kinetics. In particular, these types of models can help explain mutagenesis experiments exploring the nature of the protonation sites. For example, Villalba-Galea, 2014 proposed the existence of a voltage-independent step in Hv1 gating, and this can be easily incorporated in the model proposed here, as is the fact that hHv1 and AmHv1 channels are dimers. The present form of the allosteric model has a single subunit. Since gating in dimeric channels seems to be cooperative, allosteric models have the advantage that cooperativity between subunits can be handled naturally by including coupling between two voltage sensors.
Hv1 proton channels seem fundamental in handling fluctuations in intracellular pH and take part in several well-characterized physiological processes that depend on proton concentration changes, such as intracellular pH regulation, sperm flagellum beating, reactive oxygen species production and bacterial killing in immune cells, initiation of bioluminescence in single-celled algae, and so on Castillo et al., 2015.
What is the function of voltage-gated proton channels in corals? The deposition of a CaCO3 exoskeleton is one of the main defining characteristics of scleractinians; however, the ionic transport mechanisms involved in this process are mostly unknown. In order for aragonite precipitation to occur favorably, the pH of the calicoblastic fluid, right next to the skeleton, is maintained at high levels, between 8.5 and 9 and above the pH of sea water (Le Goff et al., 2017). It has been posited that corals control this pH via vectorial transport of protons to the gastrodermal cavity (Jokiel, 2013). Since proton transport away from the site of calcification would incur a drastically lower intracellular pH in the cells of the aboral region, we propose that, given their ability to rapidly regulate the intracellular pH (De-la-Rosa et al., 2016), Hv1 proton channels contribute by transporting protons from the cells. Thus, these proton channels could be a major component of the mechanisms of intracellular pH regulation in corals. Given that the activation range of Hv1 is controlled by the pH gradient, a large intracellular acidification would facilitate opening of these channels at the resting potential of cells, which is presumably negative.
The finding that coral Hv1 channels retain their sensitivity to Zn2+ opens the possibility of using this ion as a pharmacological tool to study the role of proton channels in pH homeostasis. It is interesting that a recent report has shown detrimental effects of zinc supplementation on coral growth (Tijssen et al., 2017), a result that could be explained by zinc inhibition of Hv1.
The physiological role of Hv1 channels in corals might be essential in the response of these organisms to ocean acidification. Although further research is needed to understand the cellular and subcellular localization of these channels, we theorize that as the pH of sea water acidifies, gating of Hv1 should require stronger depolarization, thus hindering its capacity to transport protons from the cell. This will contribute to a diminished calcification rate and less aragonite saturation of the CaCO3 skeleton. It would be interesting and important to study the effects of acidification on Hv1 physiology and pH regulation in corals in vivo. Essentially nothing is known about the electrophysiological properties of coral cells. This report represents the first time that an ion channel has been cloned and characterized in any coral and should open a new avenue of research, such as uncovering the cellular and possible subcellular localization of these channels and carefully measuring their physiological role in vivo.
Materials and methods
Identification of Hv1 sequences and cloning
Request a detailed protocolBlast searches of the transcriptome of the Indo-Pacific coral A. millepora (Moya et al., 2012) detected four sequences that we identified as belonging to a putative proton-permeable channel. The GenBank accession numbers for these are XM_015907823.1, XM_015907824.1, XM_029346499.1, and XM_029346498.1. We designed two pairs of oligonucleotides to amplify two of these sequences (Table 1). Total RNA was extracted from the tissue obtained from a fragment of A. millepora acquired from a local salt-water aquarium provider (Reefservices, Mexico City). RNA was extracted by dipping the whole fragment for 2 min in 5 ml of solution D (4 M guanidinium thiocyanate, 25 mM sodium citrate, 5% sarkosyl, and 0.1 M 2-mercaptoethanol). After incubation, the tissue was removed by gently pipetting the solution for 2 min. At this point, the calcareous skeleton was removed and RNA extraction continued according to Chomczynski and Sacchi, 1987. Total RNA (1 µg) from A. millepora was used for reverse transcription polymerase chain reaction (RT-PCR), employing oligo dT and SuperScripII reverse transcriptase (Invitrogen, USA). cDNA obtained from RT-PCR was used in three PCRs using oligos (1) AcHv1Nter5´ and 3´, (2) AcHv1Cter5´ and 3´, and (3) AcHv1Nter5´ and AcHv1Cter3´ (Table 1). The Platinium Pfx DNA polymerase (Invitrogen) was used for amplification according to the manufacturer’s instructions. 1 µl of Taq DNA polymerase (Invitrogen, USA) was used for 10 min at 72°C to add a poly-A tail at 5’ and 3’ ends and facilitate cloning into the pGEM-T vector.
Table 1
Oligonucleotides used to clone amino- and carboxy-terminal partial sequences of AmHv1 from total reverse-transcribed mRNA from A. millepora.
| Oligo name | Sequence |
|---|---|
| AcHvNt5´ | ATGATTGATGCAAGAACCAGACGATCGAGCATGGATGAT |
| AcHvNt3´ | TGATCCTGCTCTCAAGTCAAGAACCAACTCAGCAATGAC |
| AcHvCt5´ | ATGGGATTCACATTTTCAAGCACAAATGGAGGTGTTT |
| AcHvCt3´ | TCAGCTTTGTTTTAATGTTGTCAATTCAGACTCCAACTG |
The PCR 3 gave rise to a full open reading frame (ORF) containing AmHv1. New oligos, AcHv1Nter5´ and AcHv1Cter3´, containing restriction sites Kpn1 and Not1, respectively, were used to re-amplify the ORF in pGEM-T and subclone it into pcDNA3.1 for heterologous expression.
The Hv1 channel from A. palmata was cloned from a fragment of an adult specimen collected from the Limones Reef off of Puerto Morelos, Mexico. RNA extraction from small coral pieces was carried out by flash freezing in liquid nitrogen and grinding the frozen tissue. All other cloning procedures were as for A. millepora. All clones and DNA constructs were confirmed by automatic sequencing at the Molecular Biology Facility of the Instituto de Fisiología Celular at UNAM.
Heterologous expression of AmHv1
Request a detailed protocolAll electrophysiological and FRET experiments were carried out in HEK293 cells heterologously expressing the specified clone or DNA construct. HEK293 cells were acquired from ATCC (Gaithersburg, MD, USA) and were found to be free of mycoplasma infection using a PCR-based detection kit (Sigma-Aldrich, USA). Cells were grown on 100 mm culture dishes with 10 ml of Dulbecco’s Modified Eagle Medium (DMEM, Invitrogen) containing 10% fetal bovine serum (FBS) (Invitrogen, USA) and 100 units/ml-100 μg/ml of penicillin-streptomycin (Invitrogen, USA), incubated at 37°C in an incubator with 5.2% CO2 atmosphere. When cells reached 90% confluence, the medium was removed, and the cells were treated with 1 ml of 0.05% trypsin-ethylenediaminetetraacetic acid (EDTA) (Invitrogen, USA) for 5 min. Subsequently, 1 ml of DMEM with 10% FBS was added. The cells were mechanically dislodged and reseeded in 35 mm culture dishes over 5x5 mm coverslips for electrophysiology or in 35 mm glass bottom dishes for FRET experiments. In both cases, 2 ml of complete medium was used. Cells at 70% confluence were transfected with pcDNA3.1-AmHv1 prepared from a plasmid midiprep, using jetPEI transfection reagent (Polyplus Transfection, France). For patch-clamp experiments, pEGFP-N1 (BD Biosciences Clontech, USA) was cotransfected with the channel DNA to visualize successfully transfected cells via their green fluorescence. Electrophysiological recordings were done 1 or 2 days after transfection.
FRET measurement of stoichiometry
Request a detailed protocolIn order to measure the stoichiometry of subunit interaction employing FRET, we constructed fusion proteins between AmHv1 and mCerulean and mCitrine FPs, to be used as the donor and acceptor, respectively. The FPs were fused to the N-terminus of the channel in order to disrupt as little as possible the C-terminus-mediated interaction. These constructs were transfected into HEK293 cells as described above. The apparent FRET efficiency between FP-containing constructs, Eapp, was measured via sensitized emission of the acceptor, employing the spectral-FRET method (De-la-Rosa et al., 2013; Zheng et al., 2002). Fluorescence was measured in a home-modified TE-2000U inverted epifluorescense microscope (Nikon, Japan). The excitation light source was an Argon Ion laser (Spectra-Physics, Germany) mainly producing light at 458, 488, and 514 nm; the laser beam was focused and then collimated using a 3 mm ball lens and a 50-mm focal length planoconvex lens. Collimated light is steered with a mirror and then is focused into the objective back focal plane by a 300-mm focal length achromatic lens.
Cells were imaged with a Nikon 60x oil immersion objective (numerical aperture 1.4). The detection arm of the microscope is coupled to a spectrograph (Acton Instruments, USA) and an EM-CCD camera (Ixon Ultra, Andor, Ireland) controlled by Micromanager software (Edelstein et al., 2014). Excitation was achieved with appropriate excitation filters (Chroma, Vermont, USA) for mCerulean (458 nm) and mCitrine (488 nm). The emission filter was a long-pass filter in order to collect the full emission spectrum of the FRET pair. Apparent FRET efficiency is plotted as a function of the fluorescence intensity ratio (Idonor/Iacceptor). This relationship can be fitted with models of subunit association with a fixed stoichiometry, according to De-la-Rosa et al., 2013.
Electrophysiology
Request a detailed protocolAll chemicals for solutions were acquired from Sigma-Aldrich (Mexico). Proton current recordings were made from HEK293 cells expressing pCDNA3.1-AmHv1 in the inside-out, whole-cell, and outside-out configurations of the patch-clamp recording technique. For whole-cell and inside-out recordings, the extracellular solution (bath and pipette, respectively) was (in mM) 80 tetramethylammonium and methanesulfonic acid (TMA-HMESO3), 100 buffer ((2-(N-morpholino)ethanesulfonic acid (MES): pH 5.5, 6.0, and 6.5; 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES): pH 7.0, 7.5), 2 CaCl2, 2 MgCl2, and pH-adjusted N-methyl-d-glucamine/tetramethylammonium hydroxide (NMDG/TMA-OH) and HCl). The intracellular solution (pipette and bath, respectively) was (in mM) 80 TMA-HMESO3, 100 buffer (MES: pH 5.5, 6.0, and 6.5; HEPES: pH 7.0, 7.5), 1 ethylene glycol tetraacetic acid (EGTA), and pH-adjusted NMDG/TMA-OH and HCl.
Macroscopic currents were low-pass filtered at 2.5 kHz, sampled at 20 kHz with an Axopatch 200B amplifier (Axon Instruments, USA) using an Instrutech 1800 AD/DA board (HEKA Elektronik, Germany) or an EPC-10 amplifier (HEKA Elektronik, Germany). Acquisition control and initial analysis were done with PatchMaster software. Pipettes for recording were pulled from borosilicate glass capillaries (Sutter Instrument, USA) and fire-polished to a resistance of 4–7 MΩ when filled with recording solution for inside- and outside-out recordings and 1–3 MΩ for the whole-cell recording. The bath (intracellular) solutions in inside-out patches were changed using a custom-built rapid solution changer. For whole-cell recordings, all the bath solution was exchanged to manipulate pH. In some recordings, linear current components were subtracted using a p/4 subtraction protocol.
Conditions for recording zinc effects
Request a detailed protocolThe effect of zinc was evaluated in outside-out patches at a ΔpH of 1. The bath solution composition was (in mM) 100 TMA-HMESO3, 100 HEPES, 8 HCl, 2 CaCl2, 2 MgCl2, and the indicated concentration of ZnCl2. The pipette solution was (in mM) 100 TMA-MESO3, 100 MES, 8 HCl, 10 EGTA, and 2 MgCl2. Both solutions were adjusted to pH 7 and pH 6, respectively, with TMA-OH/HCl. Patches were placed in front of a perfusion tube that was gravity-fed with the appropriate solution. Tubes were changed with a home-built rapid perfusion system.
Data analysis
Request a detailed protocolConductance, G, was calculated from I-V relations assuming ohmic instantaneous currents, according to
The normalized G-V relations were fit to a Boltzmann function according to Equation 1,
Here, V0.5 is the voltage at which G/Gmax = 0.5, q is the apparent gating charge (in elementary charges, eo), KB is the Boltzmann constant, and T is the temperature in Kelvin (22°C).
The time constant of activation was estimated via a fit of the second half of currents to the equation
(2)
where Iss is the amplitude of the current at steady state, δ is the delay of the exponential with respect to the start of the voltage pulse, and τ is the time constant, both with units of ms. The voltage-dependence of δ and τ was estimated from a fit to equation
(3)
where i stands for δ or τ and k(0) is the value of either parameter at 0 mV.
Currents in the presence of zinc were normalized to the current before application of the ion to obtain a normalized fraction of current blocked as FB = 1-I/Imax. The zinc dose-response curve was fitted to Hill’s equation in the form
(4)
where KD is the apparent dissociation constant, [Zn2+]o is the extracellular zinc concentration, and nH is the Hill coefficient.
Data availability
All data generated or analyzed during this study are included in the manuscript and supporting files. We have provided an Excel file with source data used for figures.
-
NCBI GenBankID MZ029046. Acropora millepora Hv1 proton-channel nucleotide sequence. MZ029047 - Acropora palmata Hv1 proton-channel nucleotide sequence.
-
NCBI GenBankID MZ029047. Acropora palmata Hv1 proton-channel nucleotide sequence.
-
NCBI ProteinID XP_029202331.1. Title voltage-gated hydrogen channel 1-like [Acropora millepora].
-
NCBI ProteinID XP_022795192.1. voltage-gated hydrogen channel 1-like [Stylophora pistillata].
-
NCBI ProteinID XP_027057117.1. voltage-gated hydrogen channel 1-like [Pocillopora damicornis].
-
NCBI ProteinID XP_031564162.1. voltage-gated hydrogen channel 1-like [Actinia tenebrosa].
-
NCBI ProteinID XP_001626501.1. voltage-gated hydrogen channel 1 [Nematostella vectensis].
-
NCBI ProteinID XP_012554112.1. PREDICTED: voltage-gated hydrogen channel 1-like [Hydra vulgaris].
-
NCBI ProteinID NP_001119779.1. voltage-gated proton channel [Strongylocentrotus purpuratus].
-
NCBI ProteinID NP_001071937.1. voltage-gated hydrogen channel 1 [Ciona intestinalis].
-
NCBI ProteinID XP_032803138.1. voltage-gated hydrogen channel 1 [Petromyzon marinus].
-
NCBI ProteinID XP_040202566.1. voltage-gated hydrogen channel 1 [Rana temporaria].
-
NCBI ProteinID ETE71598.1. Voltage-gated hydrogen channel 1, partial [Ophiophagus hannah].
-
NCBI ProteinID XP_030847861.1. voltage-gated hydrogen channel 1-like isoform X1 [Strongylocentrotus purpuratus].
-
NCBI ProteinID EOA95241.1. Voltage-gated hydrogen channel 1, partial [Anas platyrhynchos].
-
NCBI ProteinID NP_001088875.1. voltage-gated hydrogen channel 1 [Xenopus laevis].
-
NCBI ProteinID KXJ27230.1. Voltage-gated hydrogen channel 1 [Exaiptasia diaphana].
-
NCBI ProteinID XP_035421542.1. voltage-gated hydrogen channel 1 [Cygnus atratus].
-
NCBI ProteinID NP_001025834.1. voltage-gated hydrogen channel 1 [Gallus gallus].
-
NCBI ProteinID XP_028914661.1. voltage-gated hydrogen channel 1 [Ornithorhynchus anatinus].
-
NCBI ProteinID XP_025257726.1. voltage-gated hydrogen channel 1 [Theropithecus gelada].
-
NCBI ProteinID AAH07277.1. HVCN1 protein [Homo sapiens].
-
NCBI ProteinID XP_038292573.1. voltage-gated hydrogen channel 1 isoform X1 [Canis lupus familiaris].
-
NCBI ProteinID NP_001035954.1. voltage-gated hydrogen channel 1 [Mus musculus].
-
NCBI ProteinID XP_005424087.1. voltage-gated hydrogen channel 1 [Mus musculus].
-
NCBI ProteinID XP_009633183.1. voltage-gated hydrogen channel 1 [Egretta garzetta].
-
NCBI ProteinID XP_032899181.1. voltage-gated hydrogen channel 1 [Amblyraja radiata].
-
NCBI ProteinID RGP61076.1. voltage-gated hydrogen channel 1 [Fusarium longipes].
-
NCBI ProteinID RFN53390.1. voltage-gated hydrogen channel 1 [Fusarium flagelliforme].
-
NCBI ProteinID KAF5660113.1. voltage-gated hydrogen channel 1 [Fusarium heterosporum].
-
NCBI ProteinID TQW00298.1. voltage-gated hydrogen channel 1 [Cordyceps javanica].
-
NCBI ProteinID XP_005100666.1. voltage-gated hydrogen channel 1 [Aplysia californica].
-
NCBI ProteinID THD25470.1. Voltage-gated hydrogen channel 1 [Fasciola hepatica].
-
NCBI ProteinID KAF6036357.1. HVCN1 [Bugula neritina].
-
NCBI ProteinID CAC5426376.1. HVCN1 [Mytilus coruscus].
-
NCBI ProteinID KAF5400532.1. Voltage-gated hydrogen channel 1 [Paragonimus heterotremus].
-
NCBI ProteinID TNN21174.1. Voltage-gated hydrogen channel 1 [Schistosoma japonicum].
-
NCBI ProteinID XP_012568882.1. voltage-gated hydrogen channel 1 [Cicer arietinum].
-
NCBI ProteinID XP_011656484.2. voltage-gated hydrogen channel 1 [Cucumis sativus].
-
NCBI ProteinID RDX63547.1. Voltage-gated hydrogen channel 1, partial [Mucuna pruriens].
-
NCBI ProteinID XP_026460796.1. voltage-gated hydrogen channel 1 [Papaver somniferum].
-
NCBI ProteinID QHO09623.1. Voltage-gated hydrogen channel [Arachis hypogaea].
-
NCBI ProteinID XP_038886538.1. voltage-gated hydrogen channel 1 [Benincasa hispida].
-
NCBI ProteinID RDD43770.1. Voltage-gated hydrogen channel 1 [Trichoplax sp. H2].
-
NCBI ProteinID XP_035634051.1. voltage-gated hydrogen channel 1-like [Oncorhynchus keta].
-
NCBI ProteinID XP_023152539.1. voltage-gated hydrogen channel 1 [Amphiprion ocellaris].
-
NCBI ProteinID XP_028830549.1. voltage-gated hydrogen channel 1 [Denticeps clupeoides].
-
NCBI ProteinID XP_033488479.1. voltage-gated hydrogen channel 1 [Epinephelus lanceolatus].
-
NCBI ProteinID XP_029918050.1. voltage-gated hydrogen channel 1 [Myripristis murdjan].
-
NCBI ProteinID XP_022070642.1. voltage-gated hydrogen channel 1 [Acanthochromis polyacanthus].
-
NCBI ProteinID XP_027031192.1. voltage-gated hydrogen channel 1-like [Tachysurus fulvidraco].
-
NCBI ProteinID AAH32672.1. HVCN1 protein [Homo sapiens].
-
NCBI ProteinID AEQ59286.1. voltage-gated proton channel kHv1 [Karlodinium veneficum].
-
NCBI ProteinID XP_013413952.1. voltage-gated hydrogen channel 1 [Lingula anatina].
References
-
BookCoral calcification, cells to reefsIn: Dubinsky Z, Stambler N, editors. Coral Reefs Ecosyst. Transit. Springer. pp. 119–150.https://doi.org/10.1007/978-94-007-0114-4_9
-
Post-translational cleavage of Hv1 in human sperm tunes pH- and voltage-dependent gatingThe Journal of Physiology 595:1533–1546.https://doi.org/10.1113/JP273189
-
Allostery and the Monod-Wyman-Changeux model after 50 yearsAnnual Review of Biophysics 41:103–133.https://doi.org/10.1146/annurev-biophys-050511-102222
-
The voltage-activated hydrogen ion conductance in rat alveolar epithelial cells is determined by the pH gradientJournal of General Physiology 105:861–896.https://doi.org/10.1085/jgp.105.6.861
-
Tryptophan 207 is crucial to the unique properties of the human voltage-gated proton channel, hHV1Journal of General Physiology 146:343–356.https://doi.org/10.1085/jgp.201511456
-
Engineered high-affinity zinc binding site reveals gating configurations of a human proton channelJournal of General Physiology 152:202012664.https://doi.org/10.1085/jgp.202012664
-
Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extractionAnalytical Biochemistry 162:156–159.https://doi.org/10.1016/0003-2697(87)90021-2
-
Coupling between an electrostatic network and the Zn2+ binding site modulates Hv1 activationJournal of General Physiology 150:863–881.https://doi.org/10.1085/jgp.201711822
-
Gating currents in the Hv1 proton channelBiophysical Journal 114:2844–2854.https://doi.org/10.1016/j.bpj.2018.04.049
-
Coarse architecture of the transient receptor potential vanilloid 1 (TRPV1) ion channel determined by fluorescence resonance energy transferJournal of Biological Chemistry 288:29506–29517.https://doi.org/10.1074/jbc.M113.479618
-
Currents through Hv1 channels deplete protons in their vicinityThe Journal of General Physiology 147:127–136.https://doi.org/10.1085/jgp.201511496
-
Advanced methods of microscope control using μmanager softwareJournal of Biological Methods 1:e10.https://doi.org/10.14440/jbm.2014.36
-
A human phospholipid phosphatase activated by a transmembrane control moduleJournal of Lipid Research 53:2266–2274.https://doi.org/10.1194/jlr.M026021
-
Climate change, coral bleaching and the future of the world’s coral reefsMarine & Freshwater Research 50:839–866.https://doi.org/10.1071/MF99078
-
Coupling between voltage sensor activation, Ca2+ binding and channel opening in large conductance (BK) potassium channelsJournal of General Physiology 120:267–305.https://doi.org/10.1085/jgp.20028605
-
Electrostatics and the gating pore of shaker potassium channelsJournal of General Physiology 117:69–90.https://doi.org/10.1085/jgp.117.1.69
-
Coral reef calcification: carbonate, bicarbonate and proton flux under conditions of increasing ocean acidificationProceedings of the Royal Society B: Biological Sciences 280:20130031.https://doi.org/10.1098/rspb.2013.0031
-
The role of Hv1 and CatSper channels in sperm activationThe Journal of Physiology 588:4667–4672.https://doi.org/10.1113/jphysiol.2010.194142
-
Detailed comparison of expressed and native voltage-gated proton channel currentsThe Journal of Physiology 586:2477–2486.https://doi.org/10.1113/jphysiol.2007.149427
-
An aqueous H+ permeation pathway in the voltage-gated proton channel Hv1Nature Structural & Molecular Biology 17:869–875.https://doi.org/10.1038/nsmb.1826
-
Comparison between mouse and sea urchin orthologs of voltage-gated proton channel suggests role of S3 segment in activation gatingBiochimica Et Biophysica Acta (BBA) - Biomembranes 1858:2972–2983.https://doi.org/10.1016/j.bbamem.2016.09.008
-
Activation of shaker potassium channels. I. characterization of voltage-dependent transitionsThe Journal of General Physiology 111:271–294.https://doi.org/10.1085/jgp.111.2.271
-
X-ray crystal structure of voltage-gated proton channelNature Structural & Molecular Biology 21:352–357.https://doi.org/10.1038/nsmb.2783
-
Exotic properties of a voltage-gated proton channel from the snail Helisoma trivolvisJournal of General Physiology 150:835–850.https://doi.org/10.1085/jgp.201711967
-
Proton pumping accompanies calcification in foraminiferaNature Communications 8:1–6.https://doi.org/10.1038/ncomms14145
-
BookCell biology of reef-building corals: ion transport, acid/base regulation, and energy metabolismIn: Weihrauch D, editors. Acid-Base Balance Nitrogen Excretion Invertebr. Springer. pp. 193–218.https://doi.org/10.1007/978-3-319-39617-0_7
-
Hv1 proton channel opening is preceded by a voltage-independent transitionBiophysical Journal 107:1564–1572.https://doi.org/10.1016/j.bpj.2014.08.017
-
SWISS-MODEL: homology modelling of protein structures and complexesNucleic Acids Research 46:W296–W303.https://doi.org/10.1093/nar/gky427
-
Ionic blockage of sodium channels in nerveJournal of General Physiology 61:687–708.https://doi.org/10.1085/jgp.61.6.687
-
The Whole-Genome sequence of the coral Acropora milleporaGenome Biology and Evolution 11:1374–1379.https://doi.org/10.1093/gbe/evz077
Article and author information
Author details
Miguel A Cervantes-Rocha

Esteban Suárez-Delgado
Anastazia T Banaszak
Funding
Universidad Nacional Autónoma de México (IN215621)
- Leon D Islas
Universidad Nacional Autónoma de México (IN200720)
- Tamara Rosenbaum
Universidad Nacional Autónoma de México (IN247765)
- Anastazia T Banaszak
Consejo Nacional de Ciencia y Tecnología (2)
- Ernesto Maldonado
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
We would like to thank Itzel A Llorente for the excellent technical assistance. We also thank Gerardo Coello from IFC-UNAM for help with analysis of coral transcriptomes. This work was funded in part by grant no. IN215621 from DGAPA-PAPIIT-UNAM to LDI, grant no. 247765 to ATB, and grant no. IN200720 to TR. EM was funded by Conacyt-Fronteras en la Ciencia Grant No. 2.
Copyright
© 2021, Rangel-Yescas et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 1,342
- views
-
- 217
- downloads
-
- 16
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 16
- citations for umbrella DOI https://doi.org/10.7554/eLife.69248
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Discovery and characterization of Hv1-type proton channels in reef-building corals
eLife 10:e69248.
https://doi.org/10.7554/eLife.69248




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}