5′-Modifications improve potency and efficacy of DNA donors for precision genome editing
- RNA Therapeutics Institute, University of Massachusetts Medical School, United States
- Department of Pediatrics, Division of Genes and Development, University of Massachusetts Medical School, United States
- Program in Bioinformatics and Integrative Biology, University of Massachusetts Medical School, United States
- Department of Molecular, Cell and Cancer Biology, University of Massachusetts Medical School, United States
- Department of Biochemistry and Molecular Biotechnology, University of Massachusetts Medical School, United States
- Li Weibo Institute for Rare Diseases Research, University of Massachusetts Medical School, United States
- Program in Molecular Medicine, University of Massachusetts Medical School, United States
- Howard Hughes Medical Institute, University of Massachusetts Medical School, United States
Figures
Figure 1 with 1 supplement
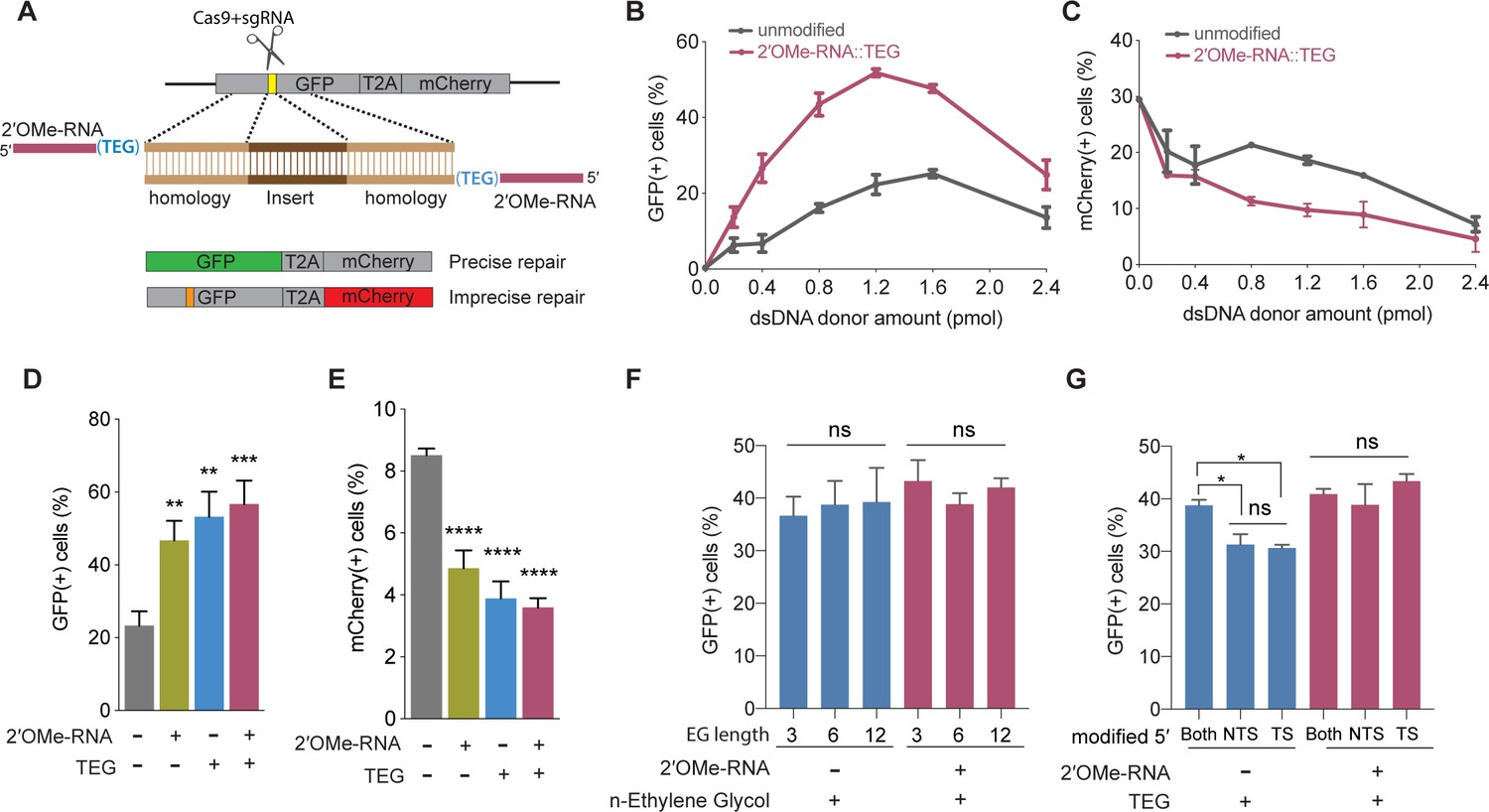
5′-End-modified donors promote homology-directed repair (HDR) in traffic light reporter (TLR) cells.
(A) Schematic showing the TLR assay to quantify HDR efficiencies using unmodified or end-modified double-stranded DNA (dsDNA) donors. Editing efficiencies plotted as percentage of (B) green fluorescent protein (GFP)+ (HDR) and (C) mCherry+ (non-homologous end joining [NHEJ]) HEK293T TLR cells at different amounts of unmodified, 2′-O-methyl (2′OMe)-RNA::triethylene glycol (TEG)-modified dsDNA donors. Editing efficiencies plotted as percentage of (D) GFP+ (HDR) and (E) mCherry+ (NHEJ) HEK293T TLR cells at 1.2 pmol of dsDNA donors indicated. Percentage of GFP+ cells obtained with dsDNA donors modified with various lengths of ethylene glycol (F) and with modifications to only one end or both 5′-ends of the donor. TS, target strand; NTS-,non-target strand (G). Mean ± s.d. for at least three independent replicates are plotted; two replicates for TEG donor in panel G.
Figure 1—figure supplement 1

2′-O-methyl (2′OMe)-RNA at 5′-ends of donors promote homology-directed repair (HDR) in mammalian cells.
Editing efficacy plotted as percentage of (A) green fluorescent protein (GFP)+ (HDR) and (B) mCherry+ (non-homologous end joining [NHEJ]) HEK293T traffic light reporter (TLR) cells at different amounts of unmodified, 2′OMe-RNA::triethylene glycol (TEG)-modified and peptide nucleic acid (PNA)::nuclear localization signal (NLS)-annealed double-stranded DNA (dsDNA) donors. Same unmodified controls are used in Figure 1B and C. Addition of PNA (without NLS) to unmodified or end-modified donors does not further improve HDR efficiency in mammalian cells. 0.8 pmol of each type of donor was annealed to PNA (0.1–10 pmol). Editing efficiency was plotted as percentage of (C) GFP+ (HDR) cells and (D) mCherry+ (NHEJ) cells. Percentages were calculated by sorting the cells through flow cytometry (see Materials and methods). PNA::NLS and 2′OMe-RNA::TEG modified donors were mixed at 4:1 molar ratio, heated to 94°C and cooled to anneal PNA to 2′OMe-RNA.
Figure 2 with 1 supplement
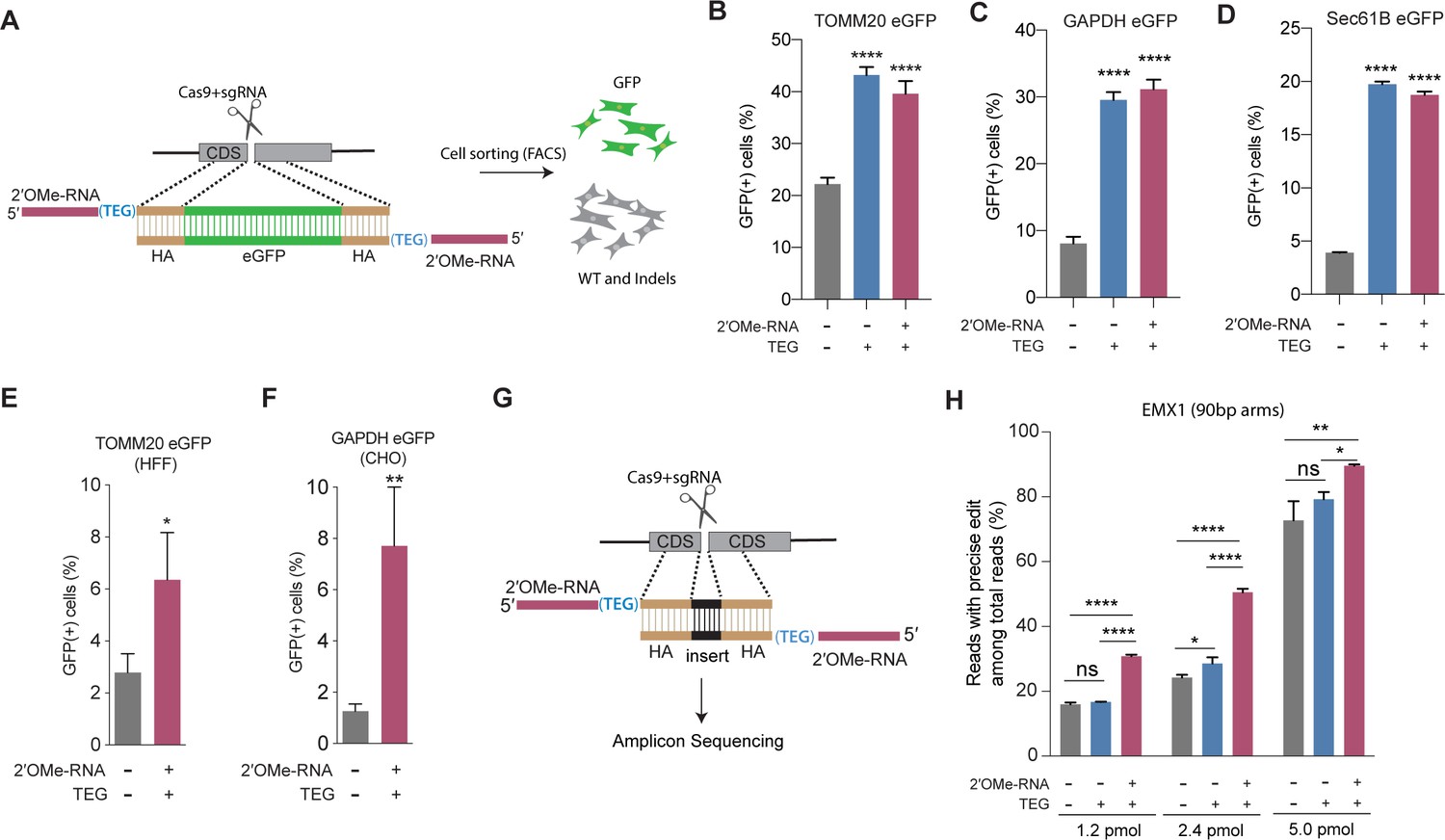
End-modified donors promote homology-directed repair (HDR) at endogenous loci in mammalian cell cultures.
(A) Schematic representation of the 5′-modified donor design for enhanced green fluorescent protein (eGFP) insertion and fluorescence-activated cell sorting (FACS) is shown. Efficacy of eGFP integration at (B) TOMM20 and (C) GAPDH (D) Sec61B loci in HEK293T cells using unmodified, triethylene glycol (TEG) or 2′-O-methyl (2′OMe)-RNA::triethylene glycol (TEG)-modified donors are plotted as percentage of GFP+ cells. Efficacy of eGFP integration at the (E) TOMM20 locus in human foreskin fibroblast (HFF) (747 bp knock-in with ~1 kb homology arms) and (F) Gapdh locus in Chinese hamster ovary (CHO) (1635 bp knock-in with ~800 bp homology arms) cells using double-stranded DNA (dsDNA) (500 ng) donors with and without 2′OMe-RNA::TEG modifications at the 5′-ends. (G) Schematic representation of the dsDNA donor design used for quantification with deep sequencing is shown. (H) Illumina sequencing reads with precise knock-in are plotted for dsDNA donors with 90 bp homology arms at EMX1 locus in HEK293T cells. Mean ± s.d. for at least three independent replicates are plotted. p-Values were calculated using one-way ANOVA and in all cases end-modified donors were compared to unmodified donor unless indicated otherwise (Tukey’s multiple comparisons test; ****p < 0.0001; ***p < 0.001; **p < 0.01; *p < 0.05; ns, not significant).
Figure 2—figure supplement 1
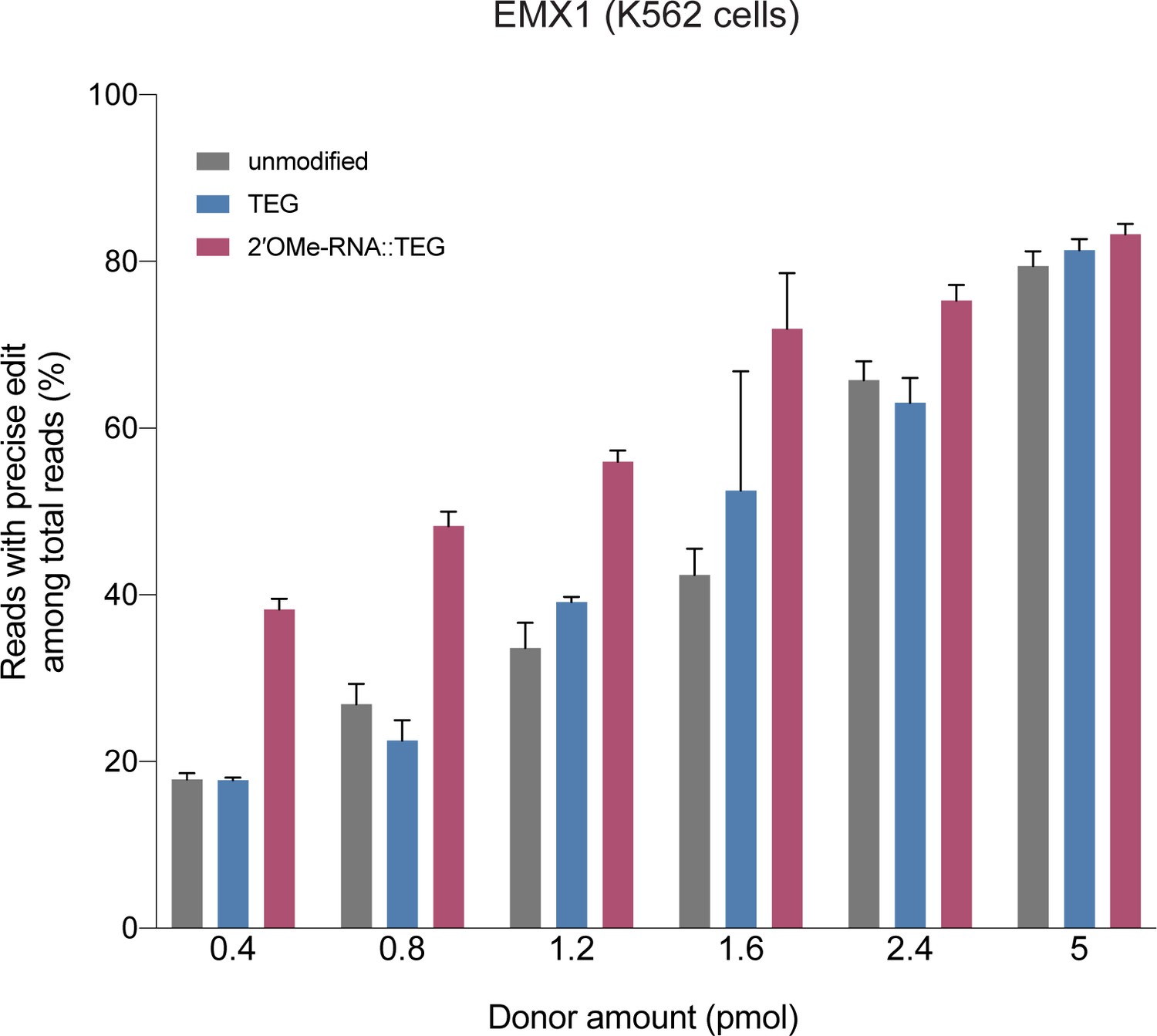
RNA::TEG (triethylene glycol) donors with short (90 bp) homology arms are more potent than unmodified donors at EMX1 locus.
Fraction of precise reads is plotted as percentage of total Illumina reads obtained at various amounts of double-stranded DNA (dsDNA) donors into K562 cells. Cas9 ribonucleoproteins (RNPs) and dsDNA donors were nucleofected into K562 cells and harvested after 3 days.
Figure 3 with 2 supplements

End modifications increase potency of single-stranded oligodeoxynucleotide (ssODN) donors.
(A) Editing efficacy plotted as percentage of green fluorescent protein (GFP+) (precise) HEK293T traffic light reporter (TLR) cells at different amounts of unmodified and 2′-O-methyl (2′OMe)-RNA::triethylene glycol (TEG)-modified long single-stranded DNA (ssDNA) donors (800 nt). (B) Editing efficacy of GFP-to-blue fluorescent protein (BFP) reporter conversion in K562 cells using different amounts of unmodified and 2′OMe-RNA::TEG-modified 66 nt single-stranded oligodeoxynucleotide (ssODN) donors plotted as percentage of BFP+ cells (homology-directed repair [HDR]). (C). Editing efficacy of GFP-to-BFP conversion in K562 cells using 0.5 pmol of ssODN donors modified at the 5′-end alone, the 3′-end alone, or at both the 5′- and 3′-ends, with phosphorothioate (PS), TEG, 2′OMe-RNA, or 2′OMe-RNA::TEG, plotted as percentage of BFP+ cells (precise). Complete figure of panel C is shown, along with other modifications, in Figure 3—figure supplement 2. Mean ± s.d. for at least three independent replicates are plotted. p-Values were calculated using one-way ANOVA and in all cases end-modified donors were compared to unmodified donor unless indicated otherwise (Tukey’s multiple comparisons test; ****p < 0.0001; ***p < 0.001; **p < 0.01; *p < 0.05; ns, not significant).
Figure 3—figure supplement 1
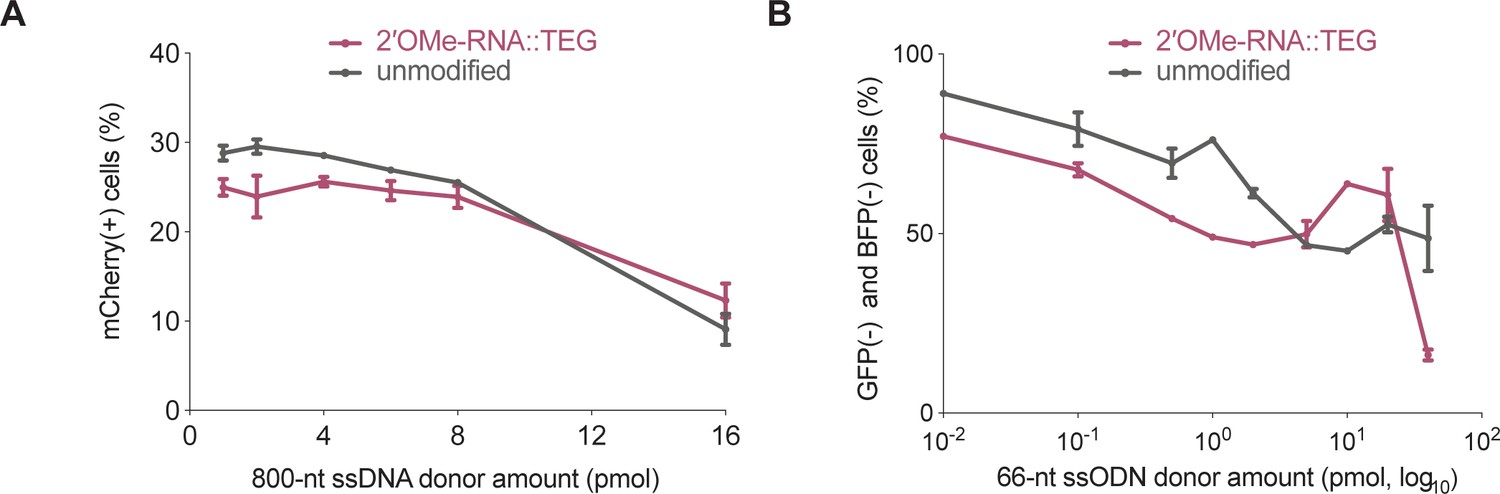
2′-O-methyl (2′OMe)-RNA::triethylene glycol (TEG) modification of single-stranded DNA (ssDNA) donors results in reduced imprecise editing.
(A) Imprecise editing efficiency plotted as percentage of mCherry(+)HEK293T traffic light reporter (TLR) cells at different amounts of unmodified or TEG::2′OMe-RNA-modified ssDNA donor. Fraction of cells expressing green fluorescent protein (GFP) is plotted in Figure 3A. (B) Imprecise editing plotted as percentage of GFP(-) and blue fluorescent protein (BFP)(-) cells in GFP-to-BFP reporter K562 cells using different amounts of unmodified and TEG::2′OMe-RNA-modifed single-stranded oligodeoxynucleotide (ssODN) donors. Fraction of cells expressing BFP is plotted in Figure 3B.
Figure 3—figure supplement 2
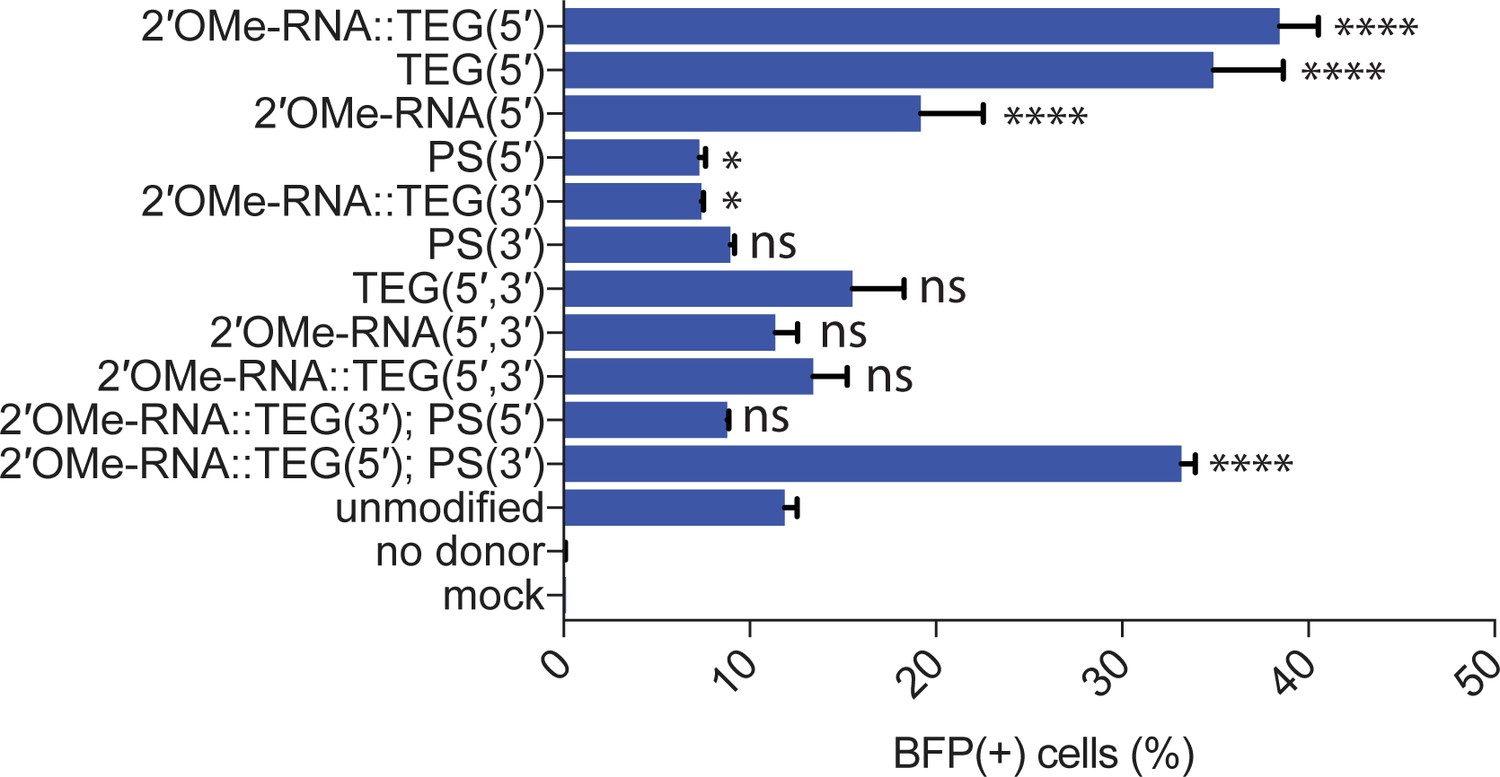
Effects of terminal and non-terminal modifications of single-stranded oligodeoxynucleotide (ssODN) donors on homology-directed repair (HDR) efficacy.
Editing efficacy of green-to-blue fluorescent protein (GFP-to-BFP) conversion in K562 cells using 0.5 pmol of ssODN donors modified at the 5′-end alone, the 3′-end alone, or at both the 5′- and 3′-ends, with phosphorothioate (PS), triethylene glycol (TEG), 2′-O-methyl (2′OMe)-RNA, or 2′OMe-RNA::TEG, plotted as percentage of BFP(+) cells (HDR). This figure consists of the data shown in Figure 3C along with other controls and modifications to the donors. Note that the PS modification is at the 5′- or 3′-internal linkages while TEG modifications are appended to the 5′- or 3′-terminus. All data points represent a mean of at least three independent replicates and all error bars represent standard deviation. p-Values were calculated using one-way ANOVA and in all cases end-modified donors were compared to the unmodified donor (Tukey’s multiple comparisons test; ****p < 0.0001; ***p < 0.001; **p < 0.01; *p < 0.05; ns, not significant).
Figure 4 with 1 supplement
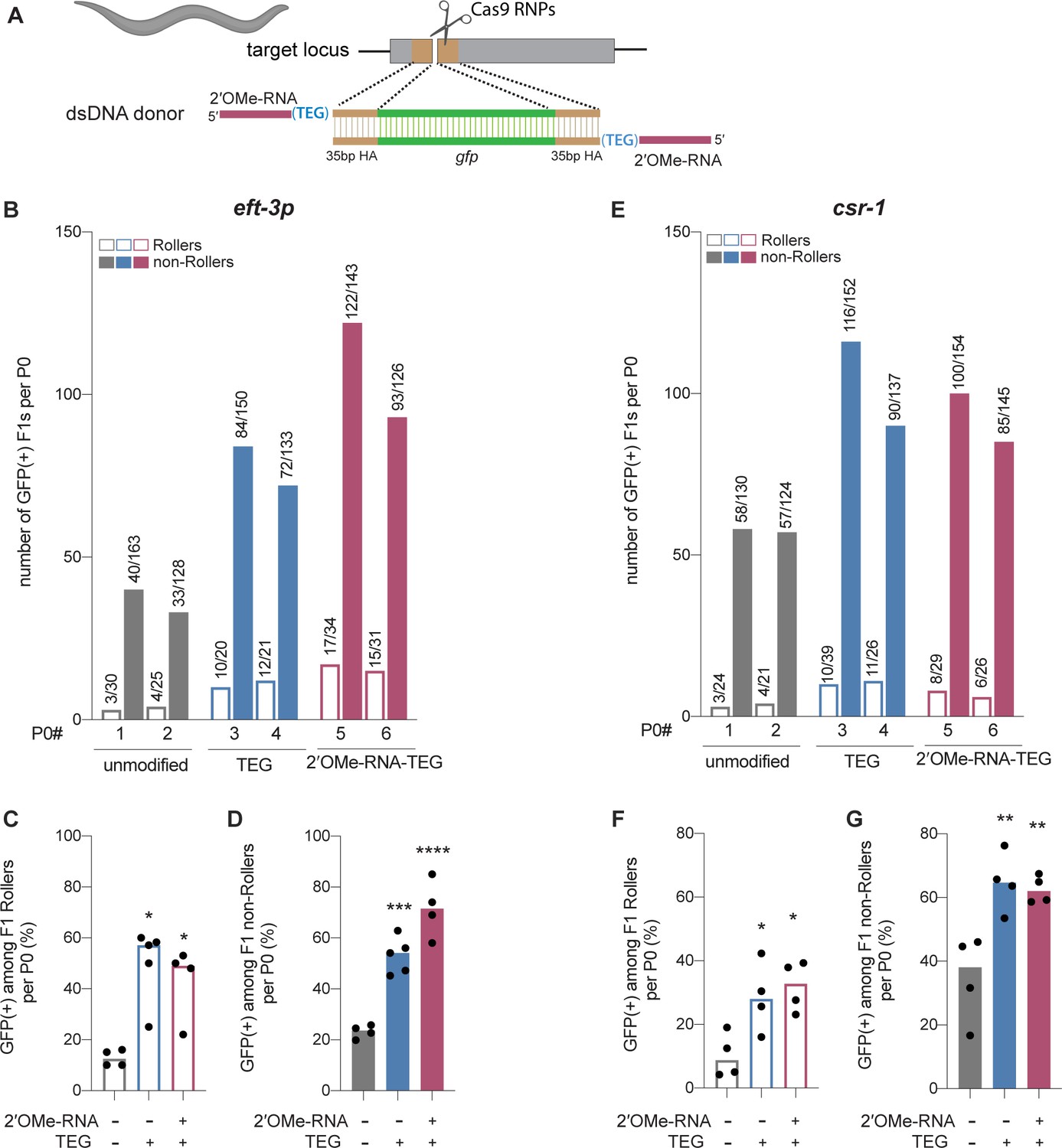
Modified donors promote precise editing in Caenorhabditis elegans.
(A) Schematic showing end-modified double-stranded DNA (dsDNA) donors (25 ng/µl) with short (~35 bp) homology arms to insert gfp tag. (B) Number of green fluorescent protein (GFP) expressing animals among entire F1 brood of two representative P0 animals for each donor type are plotted for eft-3p reporter locus. Fraction of F1 animals expressing GFP among (C) Roller and (D) non-Roller cohorts are plotted as percentage for eft-3p locus. Similarly, (E) number of GFP expressing animals among two representative broods, fraction of F1 animals expressing GFP among (F) Roller and (G) non-Roller cohorts are plotted for csr-1 locus. Open bars (Rollers) and closed bars represent (non-Rollers) median. Number of GFP expressing animals among total number of animals scored per cohort are shown above the bars. n ≥ 4 broods for each donor condition. p-Values were calculated using one-way ANOVA and in all cases end-modified donors were compared to unmodified donors (Tukey’s multiple comparisons test; ****p < 0.0001; ***p < 0.001; **p < 0.01; *p < 0.05; ns, not significant).
Figure 4—figure supplement 1
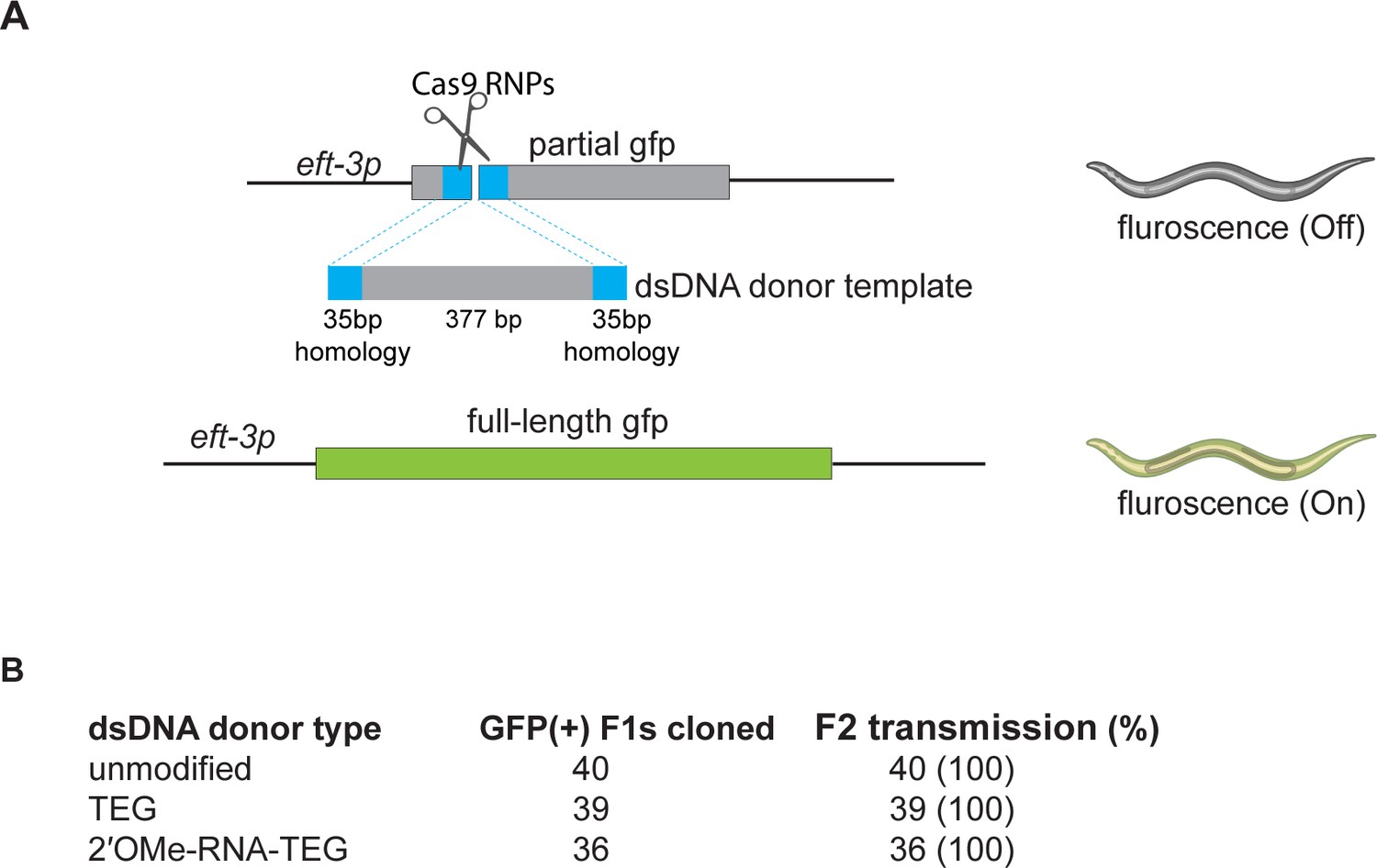
Precise insertions are germline transmitted in Caenorhabditis elegans.
(A) Schematic representation of the eft3p-gfp (partial) locus edited with double-stranded DNA (dsDNA) donors with or without end modifications in C. elegans. Precisely edited animals express green fluorescent protein (GFP) signal ubiquitously. (B) GFP-positive F1 animals were cloned, and their progeny (F2s) were scored for GFP expression. Number of F1s that produced GFP expressing F2 in a Mendelian fashion are shown under F2 transmission column.
Figure 5 with 3 supplements
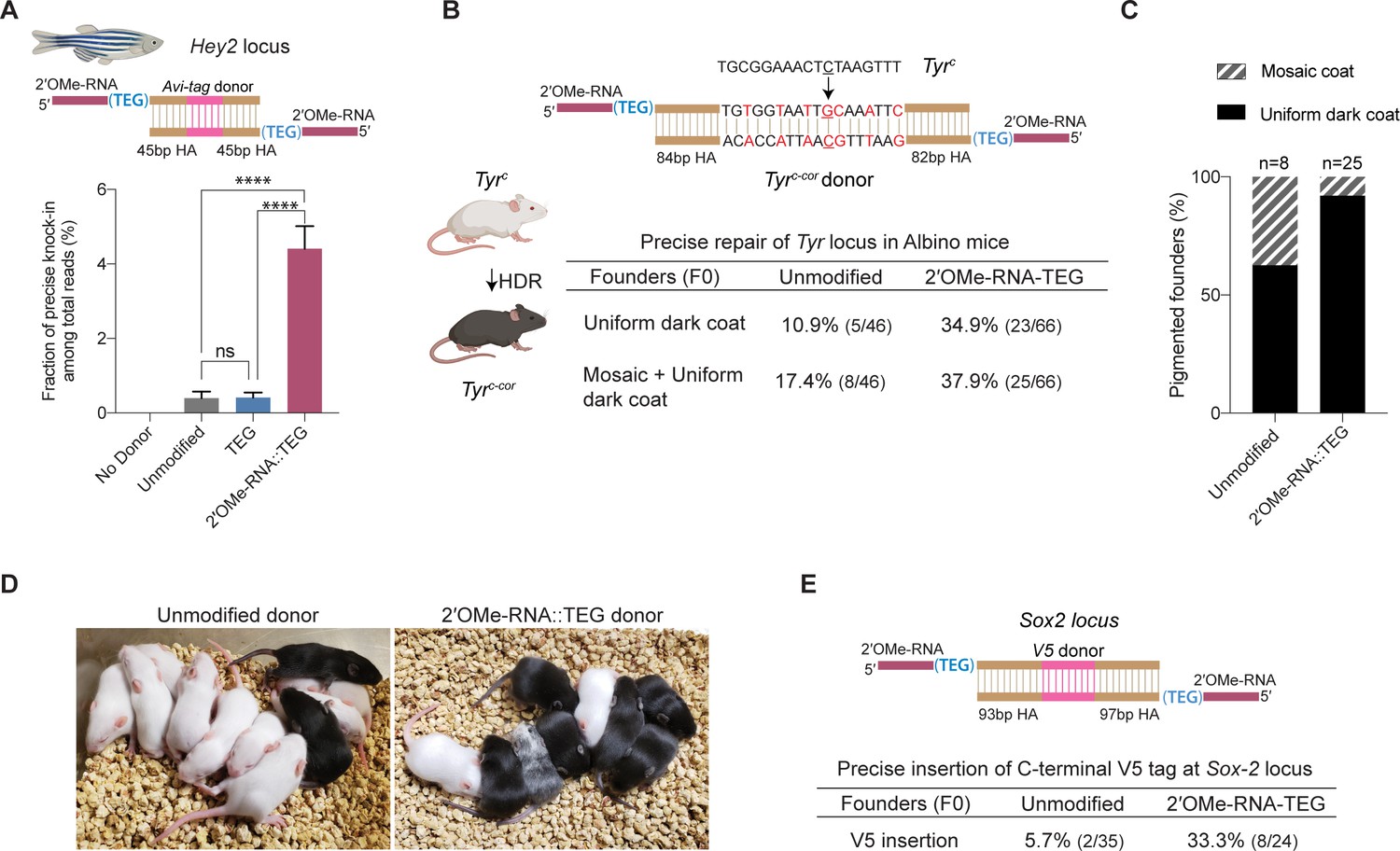
2′-O-methyl (2′-OMe)-RNA-triethylene glycol (TEG) donors promote precise editing in vertebrate zygotes.
(A) Unmodified, TEG, and 2′-OMe-RNA::TEG-modified double-stranded DNA (dsDNA) donors were injected into zebrafish zygotes. dsDNA donor design to knock-in Avi-tag is shown on the top and the fraction of Illumina reads containing precise knock-in are plotted as percentages. Mean ± s.d. for at least three independent replicates (two for unmodified donors) are plotted (B). Design of the dsDNA donors injected into mouse zygotes to precisely convert the coat color of albino mice (TyrC) to pigmented (TyrC-Cor) by editing C to G (underscored) along with six silent mutations (in red) is shown. Percentages of F0 founder mice with black coat are shown. (C) Percentages of animals among homology-directed repair (HDR)-positive F0s that have uniform dark coat or mosaic coat color are plotted for unmodified and 5′-modified donors. (D) Representative pictures of 10-day- old F0 mice with pigmented (HDR) or white (wild-type [WT] or indel) coat color are shown. One mosaic mouse (third from left) can be seen among the pups obtained with end-modified donor. (E) Donor design to knock-in V5 tag at the C-terminus of Sox2 is shown on the top. Percentage of founder animals containing perfect V5 insertion at Sox2 locus are shown for each donor type. HA, homology arms. p-Values were calculated using one-way ANOVA (Tukey’s multiple comparisons test; ****p < 0.0001; ***p < 0.001; **p < 0.01; *p < 0.05; ns, not significant).
Figure 5—figure supplement 1

Indel efficiencies in zebrafish zygotes.
(A) Fraction of reads with indels among total reads obtained in experiments with either no donor, unmodified, or end-modified donors are plotted as percentage. Precise repair (homology-directed repair [HDR]) efficiencies are shown in Figure 5A.
Figure 5—figure supplement 2

5′-Modified donors improve targeted editing efficiency at the Tyr locus in albino mice.
(A) Microinjection information and homology-directed repair (HDR) efficiencies obtained for unmodified and 2′-O-methyl (2′OMe)-RNA::triethylene glycol (TEG)-modified donors. (B) Germline transmission of the edited Tyrc-cor allele was confirmed by crossing some of the pigmented F0 mice to Swiss-Webster mice (Tyrc) and phenotyping their F1 progeny. (C) Representative images of F1 litters obtained from crosses for germline transmission tests.
Figure 5—figure supplement 3

5′-Modified donors improve targeted editing efficiency at the Sox2 locus in mouse zygotes.
(A) Microinjection information and homology-directed repair (HDR) efficiencies obtained using unmodified and 2′-O-methyl (2′OMe)-RNA::triethylene glycol (TEG)-modified donors are shown. (B) Germline transmission rates of the Sox2::V5 allele was confirmed by crossing the HDR-positive F0 mice with wild-type (WT) mice and genotyping the F1 pups. (C). Sanger sequencing trace of Sox2::V5 allele in F1 mice.
Figure 6
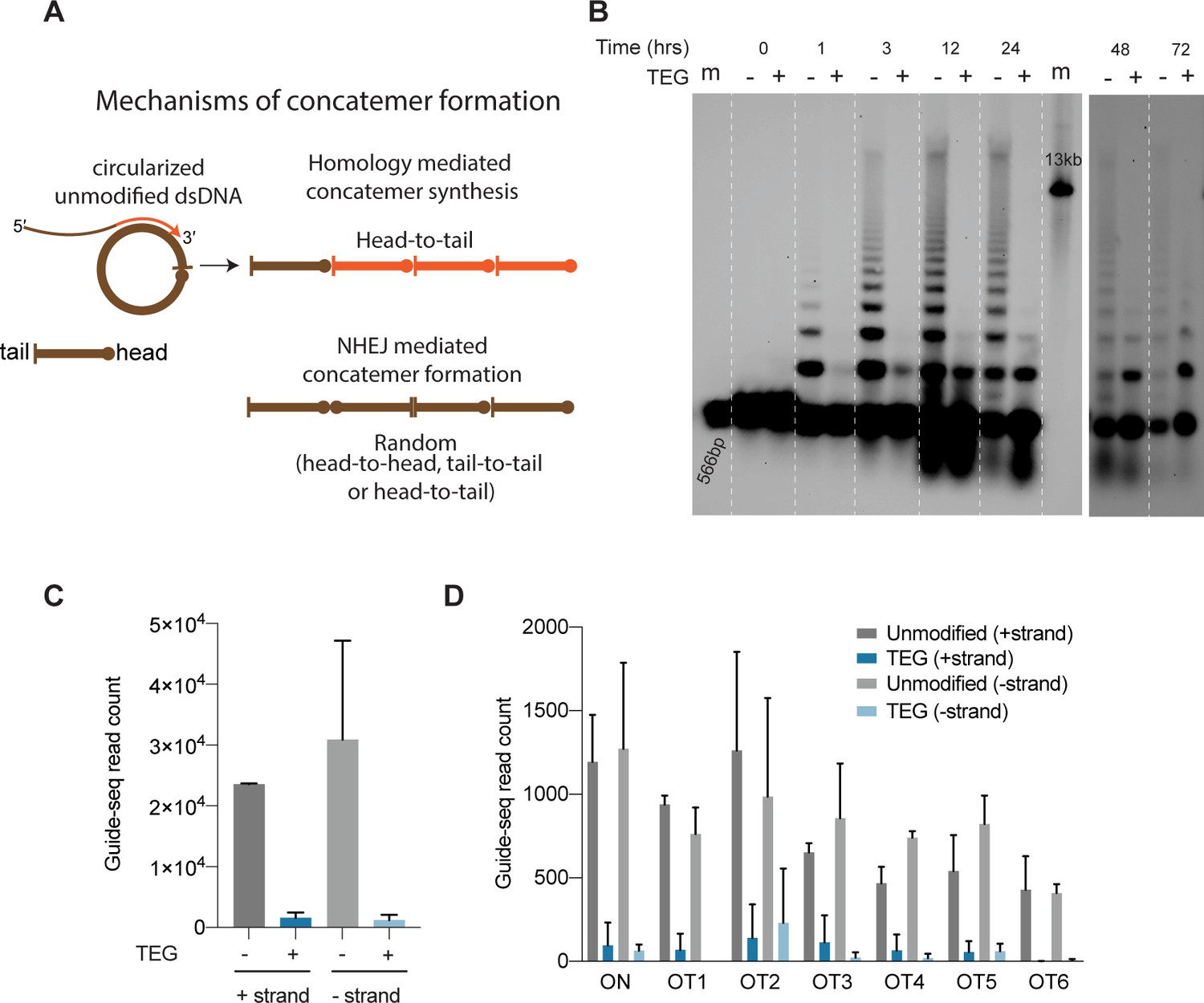
5′-Modifications suppress donor ligation reactions.
(A) Model for mechanisms of concatemer formation for unmodified donors is shown. (B) Southern blot of unmodified and triethylene glycol (TEG)-modified double-stranded DNA (dsDNA) (566 bp) nucleofected into HEK293T cells and collected at indicated time points. Concatemerization of unmodified DNA is visualized as ladders; 566 bp DNA and 13 kb long DNA are used as size markers (m). Number of GUIDE-seq reads with unmodified and TEG-modified short dsDNA (34 bp) integration for (C) whole genome and (D) on-target (ARHGEF9) and six previously validated off-target loci are plotted. Data from two biological replicates is shown.
-
Figure 6—source data 1
Uncropped full blot image of Figure 6B (0–24 hr).
Area in the box is shown in the main figure.
- https://cdn.elifesciences.org/articles/72216/elife-72216-fig6-data1-v2.zip
-
Figure 6—source data 2
Uncropped full blot image of Figure 6B (48–72 hr).
- https://cdn.elifesciences.org/articles/72216/elife-72216-fig6-data2-v2.zip
Additional files
-
Supplementary file 1
Sequences of guide RNA spacers.
- https://cdn.elifesciences.org/articles/72216/elife-72216-supp1-v2.docx
-
Supplementary file 2
Sequences of oligos.
- https://cdn.elifesciences.org/articles/72216/elife-72216-supp2-v2.docx
-
Supplementary file 3
Flow cytometry analysis to determine the percentage precise and imprecise genome editing events.
- https://cdn.elifesciences.org/articles/72216/elife-72216-supp3-v2.zip
-
Transparent reporting form
- https://cdn.elifesciences.org/articles/72216/elife-72216-transrepform1-v2.docx
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
5′-Modifications improve potency and efficacy of DNA donors for precision genome editing
eLife 10:e72216.
https://doi.org/10.7554/eLife.72216




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}