Isoform-specific mutation in Dystonin-b gene causes late-onset protein aggregate myopathy and cardiomyopathy
- Division of Neurobiology and Anatomy, Graduate School of Medical and Dental Sciences, Niigata University, Japan
- Transdisciplinary Research Programs, Niigata University, Japan
- Department of Physiology, School of Dentistry, Iwate Medical University, Japan
- Medical AI Center, School of Medicine, Niigata University, Japan
- Department of Nursing, Niigata College of Nursing, Japan
- Electron Microscope Laboratory, Keio University, Japan
- Department of Neuromuscular Research, National Institute of Neuroscience, National Center of Neurology and Psychiatry, Japan
- Division of Microscopic Anatomy, Graduate School of Medical and Dental Sciences, Niigata University, Japan
- Center for Coordination of Research Facilities, Niigata University, Japan
Peer review process
This article was accepted for publication as part of eLife's original publishing model.
History
- Version of Record published
- Accepted
- Preprint posted
- Received
Decision letter
-
Christopher CardozoReviewing Editor; Icahn School of Medicine at Mount Sinai, United States
-
Mone ZaidiSenior Editor; Icahn School of Medicine at Mount Sinai, United States
-
Mark M RichReviewer; Wright State University, United States
Our editorial process produces two outputs: (i) public reviews designed to be posted alongside the preprint for the benefit of readers; (ii) feedback on the manuscript for the authors, including requests for revisions, shown below. We also include an acceptance summary that explains what the editors found interesting or important about the work.
Decision letter after peer review:
Thank you for submitting your article "Isoform-specific mutation in Dystonin-b gene causes late-onset protein aggregate myopathy and cardiomyopathy" for consideration by eLife. Your article has been reviewed by 3 peer reviewers, and the evaluation has been overseen by a Reviewing Editor and Mone Zaidi as the Senior Editor. The following individual involved in the review of your submission has agreed to reveal their identity: Mark M Rich (Reviewer #1).
The reviewers have discussed their reviews with one another, and the Reviewing Editor has drafted this to help you prepare a revised submission.
Essential revisions:
1. Figure 1E. If possible, please provide data on what percentage of mutant mice showed kyphosis. Please also provide individual hind limb muscle weight normalized with body weight.
2. Please quantify histologic outcomes shown in Figures1C, 1D and 4.
3. Please quantify the amount of fibrosis in Figures3C and 3D.
4. Please show data on desmin aggregates as supplemental material.
5. Please add quantification of mitochondrial content.
6. Please add independent verification of selected gene expression changes by qRT-PCR.
7. Please quantify myofiber CSA as recommended by Reviewer #3.
8. Please carefully consider the remaining comments from the reviewers.
Reviewer #1 (Recommendations for the authors):
Given that the majority of the paper is about muscle pathology, I was surprised that a few of the images of wild-type muscle cross-sections used for immunostaining were not of the quality I would expect. Two examples:
– In 3A the wild-type muscle has a great deal of space between fibers. That is an artifact. This is not a problem for interpretation per se, but it raises the question of whether that is the best wild-type section the authors have.
– In Figure 4B, the WT staining of actin does not show great striations. The staining of desmin in WT muscle in 4B versus 4F is very different. It is much nicer in 4F.
The discussion highlights the problem with the significance of the findings presented. It is a list of findings, but there are no big-picture discussion points. I believe this is because the significance of the findings and underlying mechanisms remain unknown.
Reviewer #2 (Recommendations for the authors):
A few new experiments and quantification of some of the histological data should improve the impact of the manuscript. Authors should also consider rigorously studying alternation in mitochondrial content and function in skeletal and cardiac muscle of mutant mice.
1) Histological alteration in Figures 1C, 1D, and Figure 4 should be quantified.
2) The amount of fibrosis in WT and mutant mice (Figure 3C, 3D) should be quantified.
3) What is the difference in Figure 4A top right and bottom left panel pictures?
4) Expression of a few mRNAs found to be altered in RNA-Seq experiment should be independently verified by performing QRT-PCR.
Reviewer #3 (Recommendations for the authors):
I would strongly recommend that authors perform a skeletal muscle regeneration study by cardiotoxin to see how muscle regeneration is affected by mutant Dst-b. This experiment can also highlight whether or not Dst-b mutant muscle stem cells have any defect in their ability and their contribution to the formation of new muscle fibers post injury.
https://doi.org/10.7554/eLife.78419.sa1Author response
Essential revisions:
1. Figure 1E. If possible, please provide data on what percentage of mutant mice showed kyphosis. Please also provide individual hind limb muscle weight normalized with body weight.
Thank you for your suggestions. The kyphosis was observed in some (more than one third of) Dst-b mutant mice as shown in the author response image 1. MRI or CT imaging of the skeleton is necessary to accurately diagnose kyphosis, however, the imaging was not performed in this paper. Therefore, we would like not to provide data on what percentage of mutant mice showed kyphosis.
We weighed the soleus of hind limb and demonstrated the data (lines 132-135).
Author response image 1

2. Please quantify histologic outcomes shown in Figures1C, 1D and 4.
As suggested, we quantified the histological data and demonstrated in Figures 2D-G and Figure 4B. Quantification data confirmed that neuropathy do not occur in Dst-bE2610Ter/E2610Ter mice and that desmin accumulates in the mutant mice.
3. Please quantify the amount of fibrosis in Figures3C and 3D.
According to the comment, we quantified the amount of fibrosis and demonstrate the data in Figure 3E.
4. Please show data on desmin aggregates as supplemental material.
We presented data on desmin aggregates in the cardiomyocytes of Dst-bE2610Ter/E2610Ter mice (Figure 4—figure supplement 1).
5. Please add quantification of mitochondrial content.
To address this issue, we quantified muscle fibers with mitochondrial accumulations (Figure 5B). Quantitative data confirmed that mitochondria accumulate in the myofibers of soleus in Dst-bE2610Ter/E2610Ter mice.
6. Please add independent verification of selected gene expression changes by qRT-PCR.
The reliability of RNA-seq was validated by real time-PCR of 13 genes (seven up-regulated genes and six down-regulated genes). The data was demonstrated in Figure 8—figure supplement 1C.
7. Please quantify myofiber CSA as recommended by Reviewer #3.
According to the comment, we quantified distribution of cross-sectional area (CSA) in the soleus. As shown in Figure 3C, small-caliber myofibers are abundant in Dst-bE2610Ter/E2610Ter mice.
8. Please carefully consider the remaining comments from the reviewers.
The remaining comments were also carefully considered and incorporated into this revised version as much as possible.
Reviewer #1 (Recommendations for the authors):
Given that the majority of the paper is about muscle pathology, I was surprised that a few of the images of wild-type muscle cross-sections used for immunostaining were not of the quality I would expect. Two examples:
– In 3A the wild-type muscle has a great deal of space between fibers. That is an artifact. This is not a problem for interpretation per se, but it raises the question of whether that is the best wild-type section the authors have.
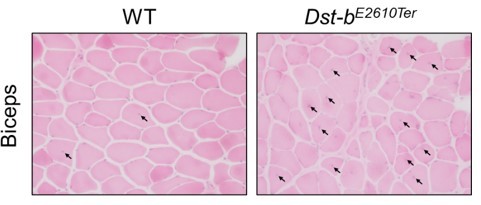
Thank you very much for the comments. In this study, we used paraffin sections, which are versatile and usually retain tissue morphology in great detail. As reviewer#1 point out, space between myofibers is artifact which is pronounced in paraffin sections than fresh frozen sections. To address the concern, we present images of muscle fibers of fresh frozen sections from WT and Dst-bE2610Ter/E2610Ter mice as Author response image 2. We believe that the use of paraffin sections does not affect the conclusions, because a large number of centrally nucleated fibers (CNFs) were observed both in paraffin sections and fresh frozen sections.
Author response image 2
– In Figure 4B, the WT staining of actin does not show great striations. The staining of desmin in WT muscle in 4B versus 4F is very different. It is much nicer in 4F.
Thank you very much for the pointing out. We replaced the images of Figure 4B with new image.
The discussion highlights the problem with the significance of the findings presented. It is a list of findings, but there are no big-picture discussion points. I believe this is because the significance of the findings and underlying mechanisms remain unknown.
Thank you very much for the critical comments. As pointed out, the mechanisms of Dst-b mutation-induced myopathy remain unknown. However, RNA-seq analysis provided molecular insights into pathophysiological mechanisms of cardiomyopathy in Dst-b mutant mice. For example, many genes responsible for unfolded protein response are affected (Figure 8C, Hspa1l and Hspb1 in Figure 8—figure supplement 1C), which is similar to several animal models of myofibrillar myopathy (Winter et al., 2014; Fang et al., J Clin Invest, 2017). Moreover, we found nuclear inclusions in Dst-b mutant cardiomyocytes as a novel pathological hallmark. In the future, we would like investigate detailed molecular mechanisms underlying formation of protein aggregates.
The biological significance of this study would have been more obvious if we could have found MFM patients with DST-b mutations. However, we have not found them so far. We hope that this report will provide an opportunity to find MFM patients with DST-b mutations.
Reviewer #2 (Recommendations for the authors):
A few new experiments and quantification of some of the histological data should improve the impact of the manuscript. Authors should also consider rigorously studying alternation in mitochondrial content and function in skeletal and cardiac muscle of mutant mice.
1) Histological alteration in Figures 1C, 1D, and Figure 4 should be quantified.
As suggested, we quantified the histological data and demonstrated in Figures 2D-G and Figure 4B. Quantification data confirmed that neuropathy do not occur in Dst-bE2610Ter/E2610Ter mice and that desmin accumulates in the mutant mice.
2) The amount of fibrosis in WT and mutant mice (Figure 3C, 3D) should be quantified.
According to the comment, we quantified the amount of fibrosis and demonstrate the data in Figure 3E.
3) What is the difference in Figure 4A top right and bottom left panel pictures?
Thank you very much for the question. Figure 4A top right and bottom left are both soleus of Dst-b mutant mice. We would like to show desmin aggregation underneath the sarcolemma in the top right picture and desmin aggregates in the sarcoplasmic region in the bottom left picture.
4) Expression of a few mRNAs found to be altered in RNA-Seq experiment should be independently verified by performing QRT-PCR.
According to the comment, the reliability of RNA-seq was validated by real time-PCR of 13 genes (seven up-regulated genes and six down-regulated genes). The data was demonstrated in Figure 8—figure supplement 1C.
Reviewer #3 (Recommendations for the authors):
I would strongly recommend that authors perform a skeletal muscle regeneration study by cardiotoxin to see how muscle regeneration is affected by mutant Dst-b. This experiment can also highlight whether or not Dst-b mutant muscle stem cells have any defect in their ability and their contribution to the formation of new muscle fibers post injury.
We agree that muscle regeneration study is very interesting subject. We would like to investigate on it in the future study.
https://doi.org/10.7554/eLife.78419.sa2Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Isoform-specific mutation in Dystonin-b gene causes late-onset protein aggregate myopathy and cardiomyopathy
eLife 11:e78419.
https://doi.org/10.7554/eLife.78419




{kind=link}
{kind=link}