R-spondin 3 deletion induces Erk phosphorylation to enhance Wnt signaling and promote bone formation in the appendicular skeleton
- School of Dental Medicine, Harvard University, United States
- German Cancer Research Center, DKFZ-ZMBH Alliance, Germany
- Institute of Molecular Biology (IMB), Germany
- Department of Medicine, Harvard Medical School, United States
- Endocrine Unit, Massachusetts General Hospital, United States
Abstract
Activation of Wnt signaling leads to high bone density. The R-spondin family of four secreted glycoproteins (Rspo1-4) amplifies Wnt signaling. In humans, RSPO3 variants are strongly associated with bone density. Here, we investigated the role of Rspo3 in skeletal homeostasis in mice. Using a comprehensive set of mouse genetic and mechanistic studies, we show that in the appendicular skeleton, Rspo3 haplo-insufficiency and Rspo3 targeted deletion in Runx2+ osteoprogenitors lead to an increase in trabecular bone mass, with increased number of osteoblasts and bone formation. In contrast and highlighting the complexity of Wnt signaling in the regulation of skeletal homeostasis, we show that Rspo3 deletion in osteoprogenitors results in the opposite phenotype in the axial skeleton, i.e., low vertebral trabecular bone mass. Mechanistically, Rspo3 deficiency impairs the inhibitory effect of Dkk1 on Wnt signaling activation and bone mass. We demonstrate that Rspo3 deficiency leads to activation of Erk signaling which in turn, stabilizes β-catenin and Wnt signaling activation. Our data demonstrate that Rspo3 haplo-insufficiency/deficiency boosts canonical Wnt signaling by activating Erk signaling, to favor osteoblastogenesis, bone formation, and bone mass.
Editor's evaluation
This seminal paper describes the divergent effects of Rspo3 haploinsufficiency on appendicular versus the axial skeletal in mice and, in doing so, highlights the differential regulation of the Wnt signaling pathway in a tissue-specific manner. Thus, while Rspo3 deficiency in osteoprogenitor cells increases bone mass in the appendicular skeleton, it causes osteopenia of vertebral bone. The study therefore not only identifies RSPO3, variants of which are associated with bone density in people, as a target for enhancing bone density in fracture-prone appendicular sites in the aging population, but also brings forth caution in the interpretation of single-site studies of the skeleton in most general terms.
https://doi.org/10.7554/eLife.84171.sa0Introduction
The Wnt signaling pathway controls cell fate decisions and tissue homeostasis during development and in the adult (Clevers and Nusse, 2012; Nusse and Clevers, 2017). It is also involved in skeletal development and essential for the regulation of bone mass in the adult skeleton (Baron and Kneissel, 2013; Gori and Baron, 2021; Huybrechts et al., 2020). Genetic or therapeutic activation of canonical Wnt signaling is associated with increased bone mass (Baron and Kneissel, 2013) and current therapeutic approaches aim at activating this pathway in patients with osteoporosis or osteogenesis imperfecta for instance (Baron et al., 2020).
Wnt signaling involves a large number of receptors and co-receptors, and of endogenous agonists and antagonists that, together, tightly regulate its activation (Clevers and Nusse, 2012; Nusse and Clevers, 2017; Baron and Kneissel, 2013; Gori and Baron, 2021). Due to this complexity, and even though Wnt signaling has been studied extensively in recent years in bone, several aspects of the mechanisms by which it regulates bone mass remain unclear. Similarly, the specific downstream events regulated by the various components of the Wnt signaling machinery and their interaction with other signaling cascades remain puzzling.
In this context, the fact that several studies have demonstrated that the four Roof plate-specific spondin, R-spondins (Rspo1 to 4), synergize with Wnt ligands to activate Wnt signaling (Gori and Baron, 2021; de Lau et al., 2014; de Lau et al., 2012; Knight and Hankenson, 2014; Lu et al., 2008; Nagano, 2019; Raslan and Yoon, 2019; Shi et al., 2016) raises the question of their potential role in skeletal development and homeostasis. This enhancement of Wnt activity is attributed to the ability of Rspos to prevent the ubiquitination and degradation of the Lrp5/6/Fzd receptor complex via Lgr4-6, closely related orphans of the leucin-rich repeat containing G protein-coupled receptors, and the transmembrane E3 ubiquitin ligases ring finger 43 (Rnf43) and zinc and ring finger 3 (Znrf3), sensitizing cells to Wnt ligands (de Lau et al., 2014; Glinka et al., 2011; Hao et al., 2012; Ruffner et al., 2012; Wang et al., 2013; Xie et al., 2013). Although the role of Lgr receptors in the effects of Rspos is well established, recent reports have shown that Rspo2 and Rspo3 can also enhance Wnt signaling independently of Lgr receptors, possibly by acting as direct antagonist ligands to RNF43 and ZNRF3 (Nagano, 2019; Lebensohn and Rohatgi, 2018; Szenker-Ravi et al., 2018).
Of particular interest is the fact that, in contrast to the many studies that have reported that Rspos co-activate Wnt signaling, studies in Zebrafish have indicated that Rspo3 can also function as a negative regulator of canonical Wnt signaling during dorsoventral and anteroposterior patterning (Rong et al., 2014; Wu et al., 2014). Additionally, it has been shown that Rspos can potentiate non-canonical Wnt cascades, such as the Wnt/PCP signaling (Glinka et al., 2011; Hao et al., 2012) and can function as antagonists of BMPR1 in Xenopus (Lee et al., 2020), two events that could have a negative impact on bone formation and bone mass. Thus, the mechanisms involved in the Rspos/Wnt signaling axis and their influence on skeletal homeostasis appear to be more complex in vivo.
The four Rspos belong to a family of cysteine-rich secreted glycoproteins with high structural similarity and 60% sequence homology (de Lau et al., 2012; Nagano, 2019). Although all Rspos are expressed in bone during development, they have specific and unique functions as reflected by the findings that their deletion leads to different phenotypes (Lu et al., 2008; Shi et al., 2016; Aoki et al., 2007; Neufeld et al., 2012; Ishii et al., 2008; Kazanskaya et al., 2008; Knight et al., 2018; Park et al., 2018; Parma et al., 2006; Wei et al., 2007). Within the Rspo family, Rspo3 is of particular interest for bone because it is highly expressed in skeletal elements during mouse development (Nam et al., 2007; Alhazmi et al., 2021) and several human GWAS studies have shown that RSPO3 SNPs are strongly associated with bone mineral density and risk of fracture (Duncan et al., 2011; Estrada et al., 2012; Lee et al., 2010; Medina-Gomez et al., 2012; Richards et al., 2008; Richards et al., 2012; Nilsson et al., 2021). Not surprisingly, in vitro studies have shown that overexpression of, or treatment with, Rspos can enhance Wnt-ligand mediated osteoblast (OB) differentiation (Lu et al., 2008; Knight et al., 2018; Zhu et al., 2016). However, similar to the Zebrafish and Xenopus studies mentioned above, it has also been reported that Rspo3 may function as a negative regulator of osteoblast differentiation in vitro (Zhang et al., 2017). Thus, taking into account the entire literature raises questions about the true net influence of Rspos, and in particular Rspo3, on skeletal homeostasis. We have recently reported that Rspo3 is expressed in osteoprogenitors in the craniofacial complex in both mice and Zebrafish (Alhazmi et al., 2021). However, Rspo3 disruption in Zebrafish has only mild effects on larval craniofacial cartilage skeleton (Alhazmi et al., 2021) and in mice its haplo-insufficiency does not appear to affect craniofacial development. Nilsson et al. have recently reported that targeted deletion of Rspo3 in osteoblast precursors in mice leads to low bone mass in the axial skeleton (L5) at 12 weeks of age, they examined only the axial skeleton where no changes in cellular and dynamic parameters were observed (Nilsson et al., 2021).
In the current studies, we have further investigated the role of Rspo3 in skeletal homeostasis. We found that if, as recently reported (Nilsson et al., 2021; Nilsson et al., 2022), targeted deletion of Rspo3 in Runx2+ cells decreases bone mass in the axial skeleton, both global Rspo3 haplo-insufficiency and targeted deletion of Rspo3 in Runx2+ osteoblast precursors lead to a marked increase in trabecular bone mass in the appendicular skeleton in both male and female mice, mainly as a result of increased bone formation. Mechanistically, we found that Rspo3 deletion leads to increased Erk phosphorylation, and, similar to increased Rspo3, stabilization of β-catenin and, activation of Wnt signaling, revealing a novel Rspo3/Erk/Wnt signaling axis that contributes to the regulation of skeletal homeostasis.
Results
Rspo3 haplo-insufficiency increases bone formation and trabecular bone mass
Rspo3 is expressed in bone during mouse skeletal development (Nam et al., 2007). We assessed Rspo3 mRNA levels in primary calvarial osteoblasts (cOBs) and found that it is expressed in these cells and that its expression increases significantly during OB differentiation (Figure 1—figure supplement 1a). To determine the physiological role of Rspo3 in skeletal homeostasis, we first used mice in which Rspo3 has been germline-deleted (Kazanskaya et al., 2008). As global deletion of Rspo3 leads to lethality by E9.5 (Aoki et al., 2007; Kazanskaya et al., 2008), before skeletal development, we analyzed mice lacking only one Rspo3 allele (Rspo3+/-) and their WT littermates. Rspo3+/- mice are born at the expected Mendelian ratio and their axial and appendicular skeleton develop normally (Figure 1—figure supplement 1b). Rspo3+/- mice continue to be healthy and do not develop any detectable pathology as they age (up to one year) (data not shown). We confirmed that Rspo3 mRNA levels were significantly decreased (50%) in Rspo3+/- mice in both marrow-depleted long bones and in cultured cOBs compared with WT mice (Figure 1a and Figure 1—figure supplement 1a). Contrary to expectations, two-way ANOVA analysis of the skeletal phenotype at 6, 12, and 18 wk of age (Figure 1b, Table 1, Figure 1—figure supplement 1c, Supplementary file 1), revealed highly significant (P<0.001) anabolic effects of Rspo3 haplo-insufficiency on structural, cellular, and dynamic parameters in both female and male mice. The skeletal phenotype of Rspo3+/- male and female mice is overall characterized by an increase in trabecular bone mass with high bone formation, mineral apposition rate and OB number and surface whereas bone resorption parameters are not affected, as indicated by osteoclast (OC) number and surface as well as eroded surface (ES/BS) (Figure 1b, Table 1, Supplementary file 1).
Figure 1 with 3 supplements see all
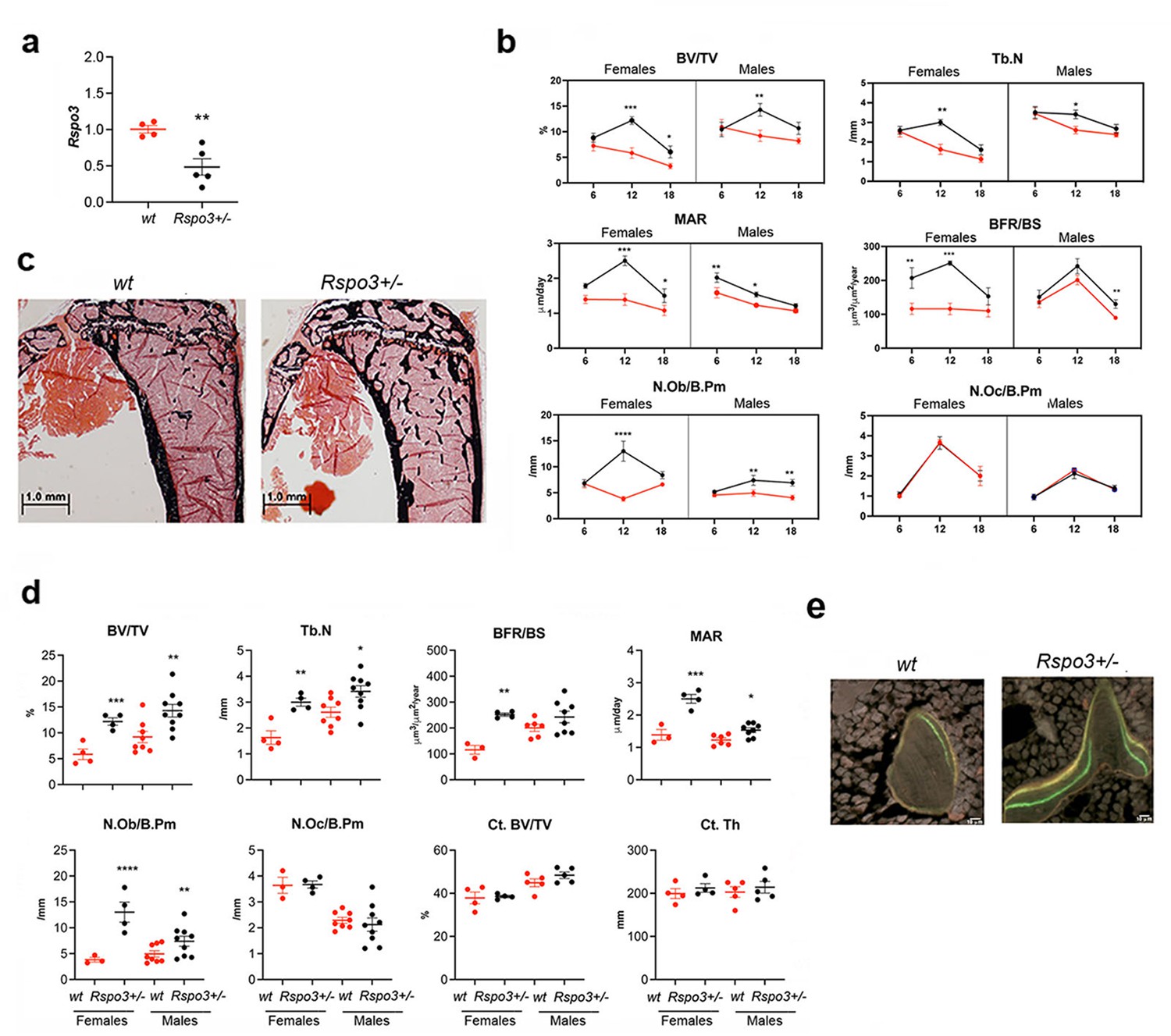
Skeletal phenotype of mice with Rspo3 haplo-insufficiency.
(a) Rspo3 expression in marrow depleted long bones, isolated from WT and Rspo3+/- mice (n=4–5). Data show all samples and are the mean ± SEM **p<0.005 by unpaired Student’s t-test. (b) Histomorphometric analysis in 6, 12, and 18 wk males and females (red line = WT, black line = Rspo3+/-). (n=3–9). Data are the mean ± SEM. Two-way ANOVA followed by Fisher’s LSD test. *p<0.05, **p<0.005, ****p<0.0001. (c) Representative images of Von Kossa staining in 12 wk-old WT and Rspo3+/- male tibiae. (n=8-9). Scale bars = 1.0 mm. (d) Histomorphometric analysis of 12 wk-old WT and Rspo3+/- female and male tibiae (n=3–9). Data show all samples and the mean ± SEM. *=p<0.05, **p<0.005 by unpaired Student’s t-test. Red circles = WT, Black circles = Rspo3+/-. (e) Representative images of double labeling in trabecular bone in 12 wk-old WT and Rspo3+/- male tibiae. (n=8-9). Scale bars = 10 μm.
Table 1
Histomorphometric analysis of WT and Rspo3+/- females.
| Parameters | 6 wk | 12 wk | 18 wk | Two way ANOVA | |||||
|---|---|---|---|---|---|---|---|---|---|
| WT(n=6) | Rspo3+/-(n=7) | WT(n=4) | Rspo3+/-(n=4) | WT(n=8) | Rspo3+/-(n=6) | Genotype | Age | Interaction | |
| BV/TV (%) | 7.24±0.98 | 8.8±0.88 | 5.84±1.01 | 12.2±0.72*** | 3.28±0.51 | 6.06±1.1* | <0.0001 | <0.0001 | NS |
| Tb.Th (μm) | 28.3±0.91 | 33.6±1.5* | 35.8±2.54 | 40.7±1.31 | 28.2±1.55 | 36.8±2.7** | 0.0003 | 0.0032 | NS |
| Tb.N (/mm) | 2.53±0.27 | 2.6±0.20 | 1.63±0.26 | 3.00±0.15** | 1.13±0.16 | 1.61±0.25 | 0.0025 | <0.0001 | 0.0429 |
| Tb.Sp (μm) | 397±52.6 | 368±26.4 | 618±86.8 | 297±17.1* | 1050±192 | 690±133 | 0.0378 | 0.0008 | NS |
| MAR (μm/day) | 1.4±0.11 | 1.8±0.06 | 1.39±0.17 | 2.5±0.14*** | 1.08±0.15 | 1.5±0.20* | <0.0001 | 0.0016 | NS |
| MS/BS (%) | 22.2±2.06 | 31.4±4.05* | 23.0±3.25 | 27.8±1.63 | 26.8±1.90 | 27.3±1.55 | 0.0394 | NS | NS |
| BFR/BS (μm3/μm2/year) | 116.2±16.6 | 207±30.4** | 116±16.9 | 251±6.43** | 110±17.6 | 153±25.1 | <0.0001 | NS | NS |
| N.Ob/B.Pm (/mm) | 6.72±0.79 | 6.85±0.64 | 3.83±0.45 | 13±1.94**** | 6.60±0.39 | 8.4±0.69 | <0.0001 | NS | 0.0001 |
| Ob.S/B.Pm (%) | 10.3±1.20 | 10.5±1.44 | 4.89±0.38 | 16.3±2.2**** | 9.66±0.52 | 12.3±0.53 | <0.0001 | NS | 0.0007 |
| OS/BS (%) | 4.68±0.78 | 5.51±1.23 | 2.51±0.30 | 9.17±0.77*** | 4.84±0.66 | 8.3±0.74** | <0.0001 | NS | 0.0263 |
| O.Th (μm) | 3.89±0.30 | 4.64±0.37 | 2.76±0.13 | 4.36±0.09* | 2.78±0.37 | 4.24±0.3** | 0.0003 | NS | NS |
| N.Oc/B.Pm (/mm) | 1.08±0.13 | 0.99±0.11 | 3.64±0.31 | 3.67±0.14 | 2.01±0.26 | 2±0.48 | NS | <0.0001 | NS |
| Oc.S/B.Pm (%) | 2.98±0.40 | 3.07±0.37 | 7.89±0.42 | 8.77±0.55 | 5.85±0.86 | 5.7±1.17 | NS | <0.0001 | NS |
| ES/BS (%) | 4.41±1.06 | 4.05±0.42 | 1.67±0.48 | 2.96±0.43 | 6.85±0.82 | 6.74±1.28 | NS | <0.0001 | NS |
-
Data are expressed as Mean ± SEM. Two way ANOVA followed by Fisher’s LSD post-hoc test. *p<0.05, **=p < 0.005, ***=p < 0.001, ****=p < 0.0001 compared with age-matched WT females.
As expected, Two-way ANOVA also demonstrated an effect of age, affecting primarily the structural parameters (Figure 1b, Table 1 and Supplementary file 1). At 12 wk of age Rspo3 haplo-insufficiency led to a significant increase in trabecular bone mass (BV/TV), trabecular thickness (Tb.Th.), and trabecular number (Tb.N) (Figure 1c–d, Table 1 and Supplementary file 1). Trabecular bone resorption parameters (Oc.S/B.Pm and N.Oc/B.Pm) were not changed in both Rspo3+/- male and female mice. In contrast, Rspo3+/- mice exhibited an increase in trabecular bone formation parameters (BFR/BS) (Figure 1d–e, Table 1 and Supplementary file 1). This increase in BFR was associated with an increase in mineral apposition rate (MAR), indicating a marked increase in the activity of individual OBs in Rspo3+/- mice, in addition to the increase in their numbers (N.Ob/B.pm, Figure 1d, Table 1 and Supplementary file 1). Consistent with these results, the osteoid surface (OS/BS) and the OB surface (Ob.S/B.Pm) were also significantly increased in both sexes (Table 1 and Supplementary file 1). Despite these marked effects on trabecular bone, cortical bone parameters were not significantly changed in Rspo3+/- female and male mice at 12 wk of age (Figure 1d, Supplementary file 2 and Supplementary file 3). In contrast to the observed effects in the tibia, no significant differences in bone mass or cellular activities were noted in the vertebrae (L5) at 12 wk of age (Figure 1—figure supplement 2 and Supplementary file 4), indicating that Rspo3 haplo-insufficiency affects preferentially the trabecular bone compartment in long bones.
Thus, in contrast with the expectation that decreasing the expression of a Wnt signaling potentiator might lead to a decrease in bone mass, our data clearly indicates that in the appendicular skeleton Rspo3 haplo-insufficiency induces an increase in trabecular bone mass due to a significant increase in bone formation, with no changes in bone resorption in both males and females. In agreement with our in vivo observations, we found that, although Rspo3 is expressed in bone marrow macrophage (BMM)-derived OCs (Figure 1—figure supplement 3a), there were no significant differences in the formation of TRAP+ multinucleated cells and in the expression of OC marker genes (Ctsk, Trap, Nfatc1) between BMM cultures from WT and Rspo3+/- in response to M-CSF and RANKL (Figure 1—figure supplement 3b and c). In addition, mix-and-matched co-cultures of cOBs and BMMs from WT or Rspo3+/- mice confirmed that Rspo3 haplo-insufficiency does not affect osteoclastogenesis, whether directly or indirectly (Figure 1—figure supplement 3d).
Rspo3 haplo-insufficiency leads to an increase in bone marrow precursor cells and in their osteoblast potential
Given that the OB number was significantly increased in Rspo3+/- mice compared with WT mice (Figure 1, Table 1 and Supplementary file 1), we determined whether this was associated with an increase in the population of precursor cells. Bone marrow flow cytometry showed that while the total number of bone stromal cells (Lin-CD45-) was not significantly affected by Rspo3 haplo-insufficiency (4047±1245 in WT compared with 5867+2382 in Rspo3+/-, mean ± SEM n=10), the mesenchymal stromal cells (MSC) population (defined here as Lin-CD45-CD31-CD51+Sca-1+) (Schepers et al., 2013) was significantly increased in Rspo3+/- mice compared with WT littermates (p=0.015) (Figure 2a and b). Consistent with these findings and with the observed increase in OB number and bone formation, Rspo3 haplo-insufficiency significantly increased CFU-F (p=0.04), and CFU-OB (p=0.034) formation (Figure 2c). Importantly, while treatment with recombinant Rspo3 did not affect the CFU-F and CFU-OB potential of WT BMSCs, it showed a tendency to decrease CFU-F and rescued significantly, i.e., prevented, the Rspo3 haplo-insufficiency-dependent increase in CFU-OB formation (Figure 2c). These data show that the changes induced by Rspo3 haplo-insufficiency cell-autonomously affect the bone marrow MSC lineage and induce an increase in the pool of progenitor cells with OB potential.
Figure 2
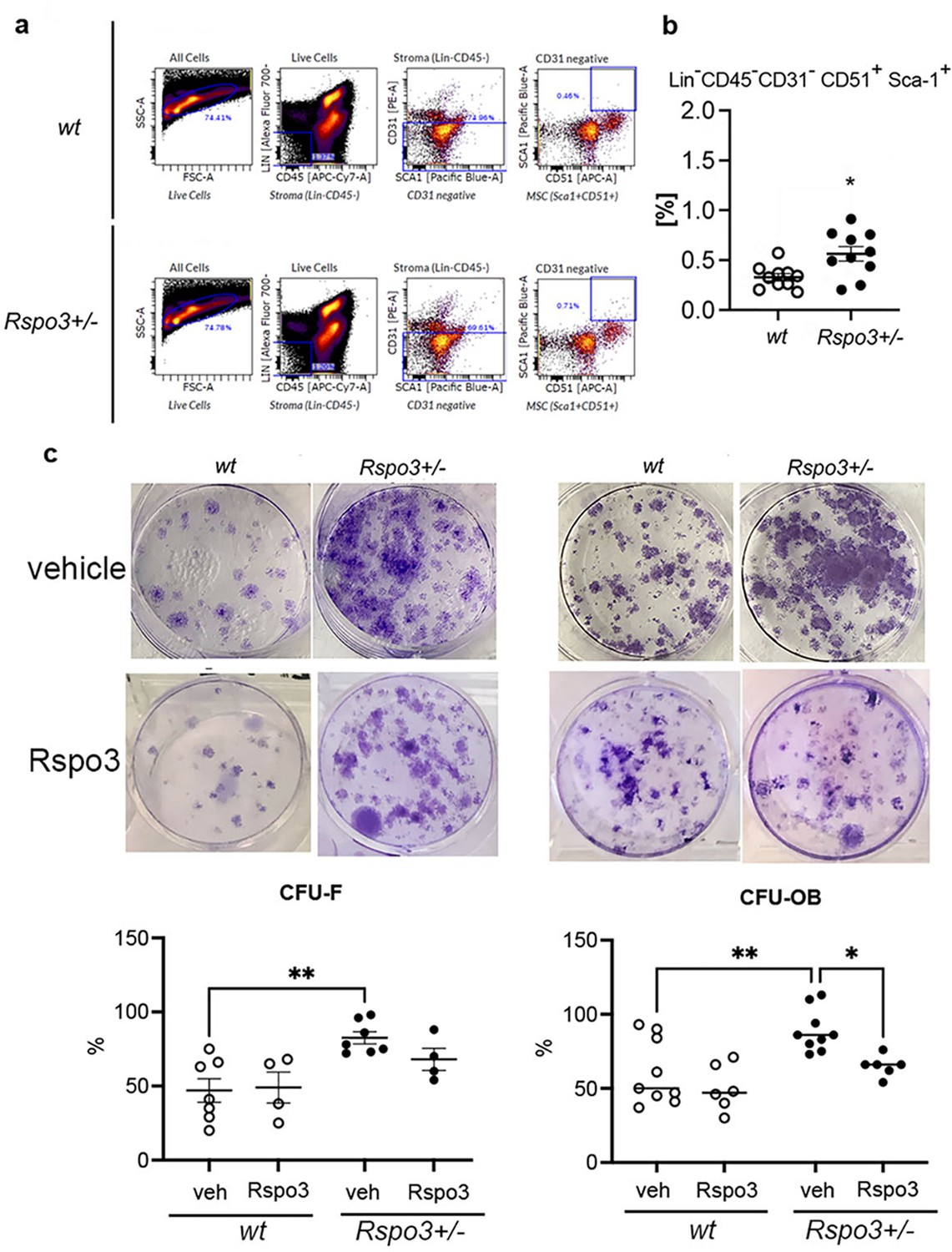
Rspo3 haplo-insufficiency increases the % of osteoprogenitors.
(a) Representative images of Flow citometry analysis. (b) Quantification of the % of Lin-Cd45-Cd31-CD51+Sca+ cells in WT and Rspo3+/- bone marrow. Data show all samples and the ± SEM (n=10) *=p< 0.05 by unpaired Student’s t-test. (c) Representative images of CFU-F and CFU-OB assay and quantification in WT and Rspo3+/- mice treated in the absence and presence of Rspo3. Data show all samples and the mean ± SEM (n=4–9) *=p<0.05, **=p<0.005 by two-way ANOVA followed by Fisher’s LSD test.
Specific deletion of Rspo3 in Runx2+ cells leads to high bone mass in the appendicular skeleton and low bone mass in the axial skeleton
To confirm the cell-autonomous effect of Rspo3 to cells of the OB lineage in vivo, we generated mice with deletion of Rspo3 in the OB lineage (Rspo3-OB-cKO) by crossing Rspo3fl/fl mice with the OB progenitors-specific Runx2-Cre mice (Rauch et al., 2010; Movérare-Skrtic et al., 2014). OB lineage-targeted deletion of Rspo3 (Figure 3a) reproduced the skeletal phenotype seen in the appendicular skeleton of Rspo3+/- mice as indicated by a significant increase in BV/TV (females p=0.017, males p<0.0001), MAR (females p=0.0034, males p<0.0001), BFR/BS (females p=0.0012, males p<0.0001) and N.Ob/B.Pm (p=0.033, males p=0.007) in Rspo3-OB-cKO compared with their control (Rspo3fl) littermates. Once again there were no changes in OC parameters (Supplementary file 5). μCT analysis of the femur, confirmed a significant increase in BV/TV, Tb.N, and Conn.D in both males and females (Figure 3d).
Figure 3 with 1 supplement see all
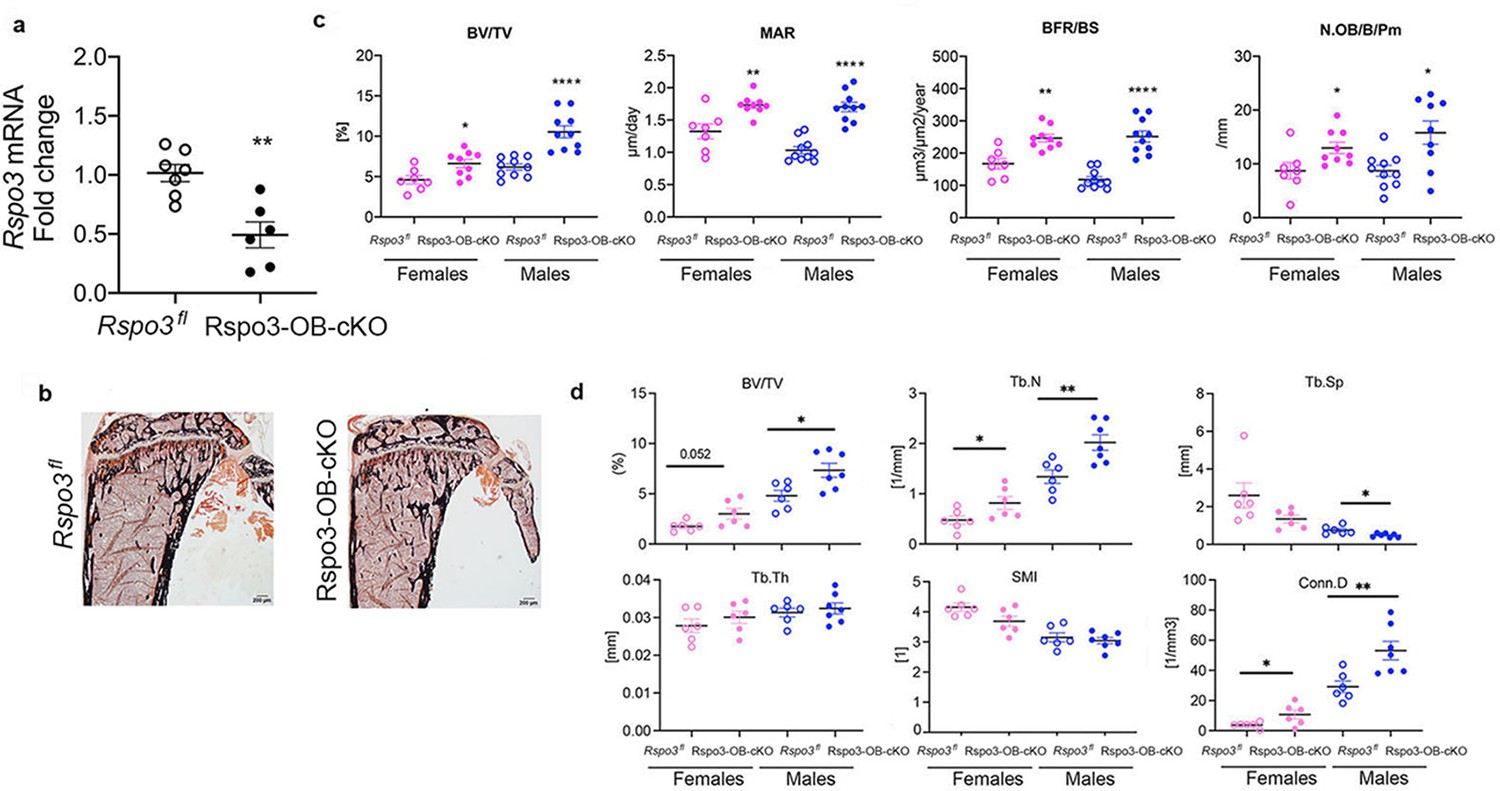
Appendicular skeletal phenotype of mice with Rspo3 targeted deletion in Runx2+ cells (Rspo3-OB-cKO).
(a) Rspo3 expression in marrow depleted long bones, showing deletion efficiency (n=6–7). Data show all samples and are the mean ± SEM. **p=0.0019 by unpaired Student’s t-test. (b) Representative images of Von Kossa staining in 8 wk old Rspo3fl and Rspo3-OB-cKO tibiae. Scale bars = 200 μm (c) BV/TV, MAR, BFR/BS, and N.Ob/BPm by histomorphometric analysis in Rspo3fl and Rspo3-OB-cKO females and males (n=7–10). (d) Structural parameters by μCT analysis Rspo3fl and Rspo3-OB-cKO femur (n=6–7). Data show all samples and the mean ± SEM *=p<0.05, **=p<0.005, ****=p<0.0001 compared with the correspondent Rspo3fl group by unpaired Student’s t-test. Open circles = Rspo3 fl and filled circles = Rspo3-OB-cKO.
Very surprisingly, however, and although the basal expression level of Rspo3 and the efficiency of deletion in long bones and vertebrae were similar (Figure 3a and Figure 4a), L5 μCT analysis showed a significant decrease in BV/TV, Tb.N, Tb.Th, and Conn.D accompanied by a marked increase in Tb.Sp. and SMI in Rspo3-OB-cKO compared with their control (Rspo3fl) male and female littermates (Figure 4b), confirming a recent report (Nilsson et al., 2022). Bone histomorphometry analysis showed that structural parameters were also significantly decreased but only in Rspo3-OB-cKO males compared with the control group (Figure 4c–d and Supplementary file 6). No significant changes were seen in the dynamic parameters. While the N.Ob/B.Pm and Ob.S/BS were not changed, the N.Oc/ B.Pm was significantly increased in Rspo3-OB-cKO males but not females, compared with the control group (Figure 4d and Supplementary file 6), providing a possible explanation for the decreased bone mass, at least in males. Thus, our studies show that targeted deletion of Rspo3 in Runx2+ osteoprogenitors leads to an opposite bone phenotype in the appendicular and the axial skeleton. Excluding any effect of the transgene or the floxed allele on the differential effect of Rspo3 targeted deletion in the axial and appendicular skeleton, μCT analysis of the femur and L5 vertebrae did not show any structural difference in BV/TV, Tb.N, Tb.Th, or Tb.Sp between Runx2-Cre, WT, and Rspo3fl mice (Figure 3 and Figure 3—figure supplement 1).
Figure 4
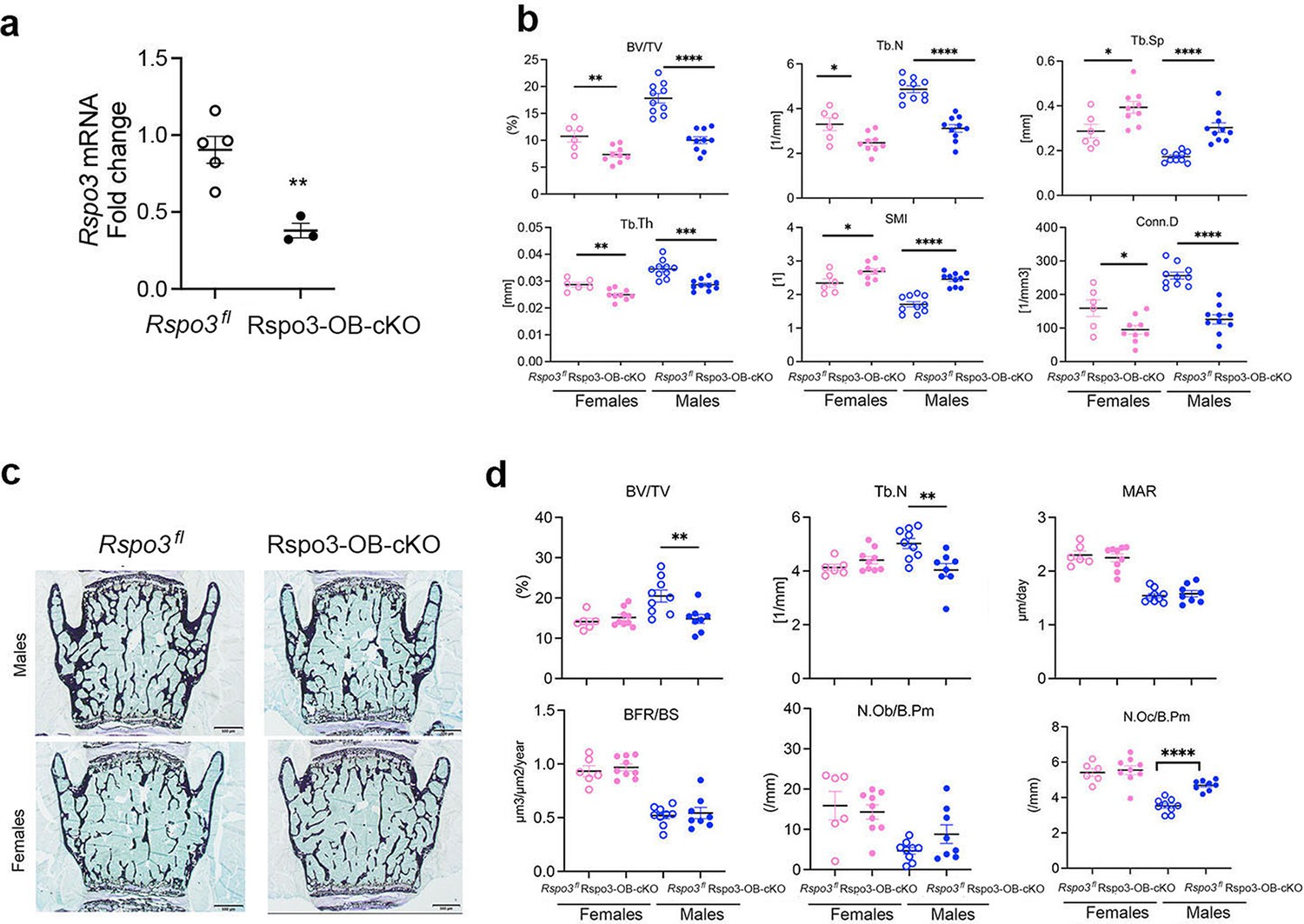
Axial skeletal phenotype of mice with Rspo3 targeted deletion in Runx2+ cells (Rspo3-OB-cKO).
(a) Rspo3 expression in vertebrae, showing deletion efficiency (n=3–5). Data show all samples and are the mean ± SEM **p=0.005 by unpaired Student’s t-test. (b) Structural parameters by μCT analysis Rspo3fl and Rspo3-OB-cKO L5 vertebrae (n=7–10). (c) Representative images of Von Kossa staining in 8 wk old Rspo3fl and Rspo3-OB-cKO L5 vertebrae. (n=7-10). Scale bars = 500 μm (d) BV/TV, Tb.N, MAR, BFR/BS, N.OC/BPm, and N.Ob/BPm by histomorphometric analysis in Rspo3fl and Rspo3-OB-cKO females and males (n=7–10). Data show all samples and the mean ± SEM *=p<0.05, **=p<0.005, ****=p<0.0001 compared with the correspondent Rspo3fl group by unpaired Student’s t-test. Open circles = Rspo3 fl and filled circles = Rspo3-OB-cKO.
Rspo3 haplo-insufficiency and deletion lead toβ-catenin stabilization
Independent of the differences, at least at the 12 wk time point, between axial and appendicular skeletal sites, our results showed a clear increase in OB progenitors in the bone marrow of Rspo3+/- mice (Figure 2) and given that trabecular bone is the main compartment affected by Rspo3 haplo-insufficiency, we then explored how depletion of Rspo3 was affecting Wnt signaling in BMSCs. Surprisingly, but consistent with our observations on bone formation, Rspo3 haplo-insufficiency led to a remarkable increase in the expression of known canonical Wnt target genes Axin2 and Tcf7 and increased active β-catenin levels (Figure 5a and b). In addition, Runx2 expression was also significantly increased in Rspo3+/- BMSCs (Figure 5a). In addition, Rspo3 haplo-insufficiency led to a significant increase in the expression of Axin2 and Dkk1, but not Sost in marrow depleted long bone cortical bone (Figure 5c). Thus, surprisingly but consistent with the bone and OB phenotypes, Rspo3 haplo-insufficiency leads to activation of canonical Wnt signaling.
Figure 5
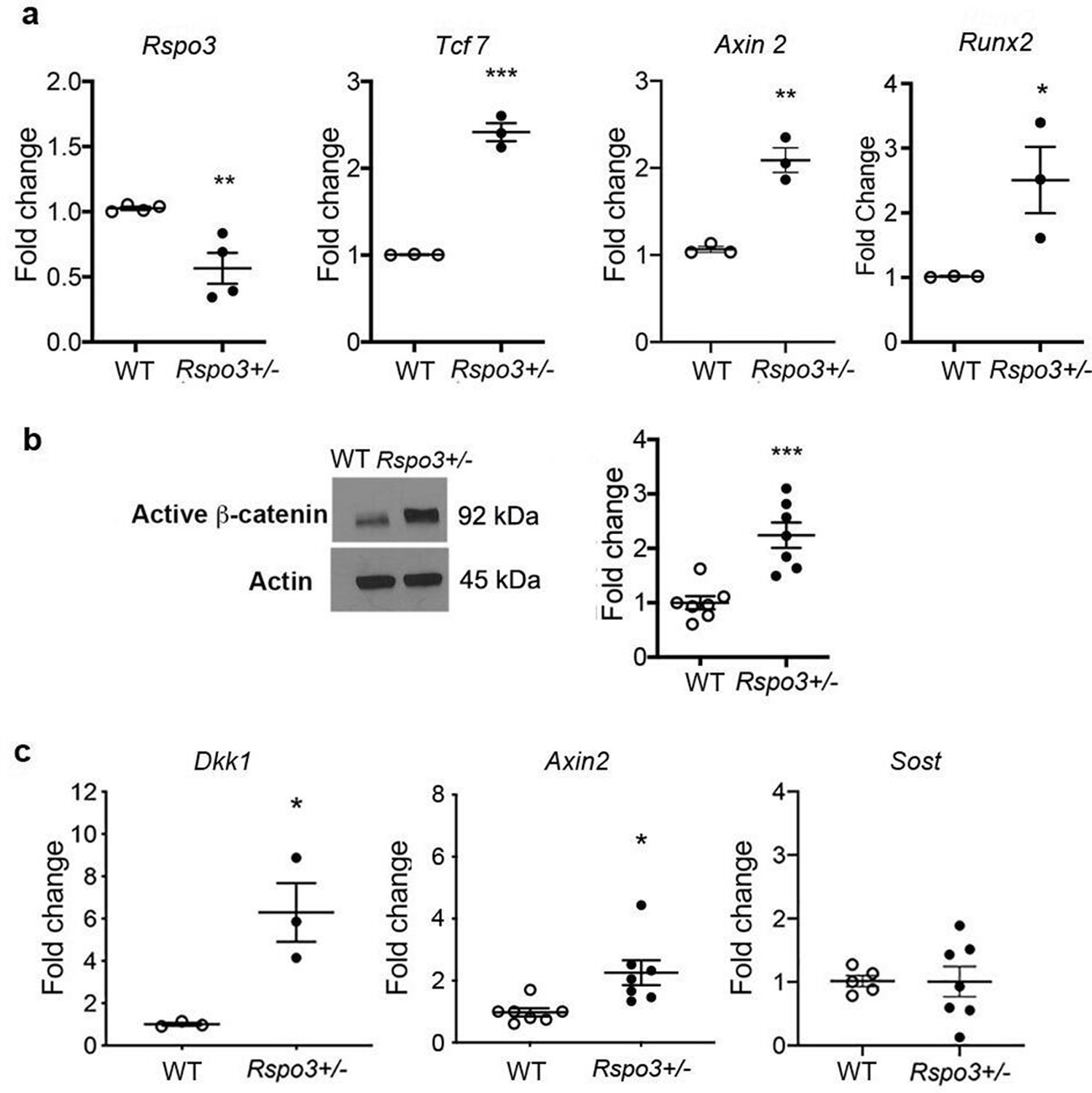
Rspo3 haplo-insufficiency leads to Wnt signaling activation.
(a) Expression of Rspo3, Runx2 and selected Wnt target genes in BMSCs (n=3–4). Data show all samples and the mean ± SEM. (b) Representative images and quantification of active β-catenin by Western analysis in BMSC isolated from WT and Rspo3+/- mice (n=7). (c) Expression of selected Wnt target genes in marrow depleted long bones (n=3–7). Data show all samples and the mean ± SEM. *p<0.05, **p<0.005, ***p<0.0005 by unpaired Student’s t-test.
-
Figure 5—source data 1
Rspo3 haplo-insufficiency leads to Wnt signaling activation.
Representative image active β-catenin by Western analysis in BMSC isolated from WT and Rspo3+/- mice (n=7).
- https://cdn.elifesciences.org/articles/84171/elife-84171-fig5-data1-v2.zip
-
Figure 5—source data 2
Rspo3 haplo-insufficiency leads to Wnt signaling activation.
Representative image of actin by Western analysis in BMSC isolated from WT and Rspo3+/- mice (n=7).
- https://cdn.elifesciences.org/articles/84171/elife-84171-fig5-data2-v2.zip
To exclude any function of the remaining Rspo3 on canonical Wnt signaling in the Rspo3+/- mice and cells, we generated Rspo3-/- mouse embryonic fibroblasts (MEFs) at E9.5, before embryonic lethality (Aoki et al., 2007; Kazanskaya et al., 2008). As shown by several groups (de Lau et al., 2014; de Lau et al., 2012; Raslan and Yoon, 2019; Park et al., 2018), we confirmed that while Rspo3 does not activate Wnt signaling by itself, it potentiates exogenous Wnt3a action in WT MEFs as indicated by the Tcf7/Lef luciferase reporter assay (Figure 6—figure supplement 1a). However, similar to what is observed in Rspo3 haplo-insufficient BMSCs and bone marrow depleted long bones cortical bone, at steady-state Rspo3 deficiency (p<0.0001) (Figure 6—figure supplement 1b) led to a marked increase in the expression of the canonical Wnt target genes Tcf7 (p<0.0001) and Axin2 (p=0.0017) as well as to a significant increase in the levels of phorphorylated Lrp6 (pLrp6), activated β-catenin and Tcf1 (Figure 6a and b). Accordingly, at steady state TOPflash reporter activity was significantly higher in Rspo3 null MEFs compared with WT MEFs (Figure 6a). Upon Wnt3a treatment, as expected, Axin2, and Tcf7 gene expression as well as pLrp6, activated β-catenin levels and TOPflash reporter activity were increased in WT MEFs but were significantly higher in Rspo3 null MEFs (Figure 6a and b). Thus, haplo-insufficiency and absence of Rspo3 in BMSCs and MEFs led to β-catenin stabilization and enhancement of β-catenin-dependent signaling. We then explored the mechanisms by which this might occur.
Figure 6 with 1 supplement see all
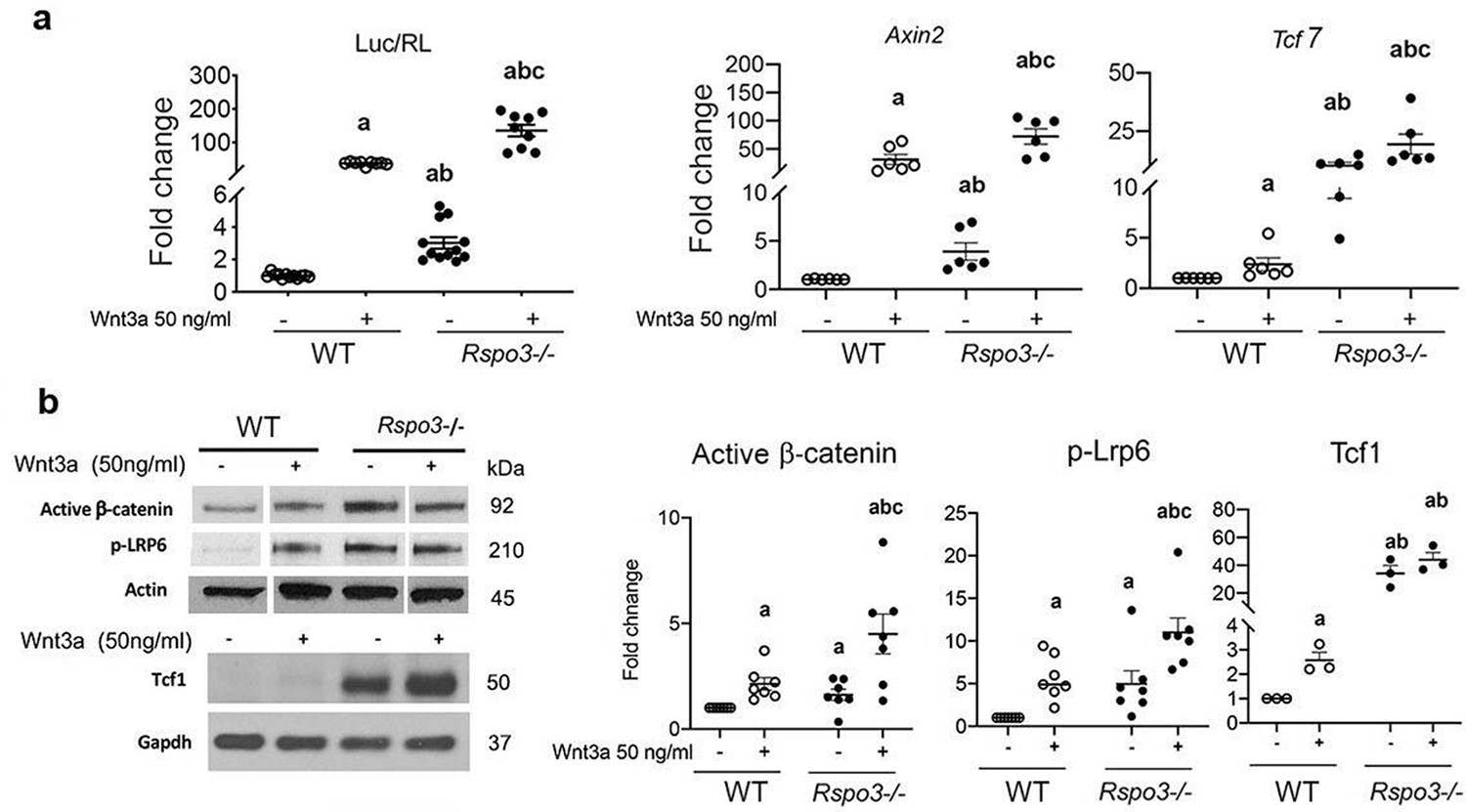
Rspo3 deletion leads to Wnt signaling activation in vitro.
(a) Luciferase assay and Wnt target gene expression in WT and Rspo-/- MEFs treated w/wo Wnt3a (n=6–11). Data show all samples and the mean ± SEM. (b) Representative images and quantification of active β−catenin, pLrp6 and Tcf1 by Western analysis in WT and Rspo-/- MEFs treated w/wo Wnt3a (n=3–7). Data show all samples and the mean ± SEM. a=p<0.05 vs vehicle WT, b=p<0.05 vs Wnt3a–treated WT and c=p<0.05 vs Wnt3a treated Rspo3-/- by two-way ANOVA followed by Fisher’s LSD test.
-
Figure 6—source data 1
Rspo3 deletion leads to Wnt signaling activation in vitro.
Representative image of active β−catenin by Western analysis in WT and Rspo-/- MEFs treated w/wo Wnt3a (n=7).
- https://cdn.elifesciences.org/articles/84171/elife-84171-fig6-data1-v2.zip
-
Figure 6—source data 2
Rspo3 deletion leads to Wnt signaling activation in vitro.
Representative image of pLrp6 by Western analysis in WT and Rspo-/- MEFs treated w/wo Wnt3a (n=7).
- https://cdn.elifesciences.org/articles/84171/elife-84171-fig6-data2-v2.zip
-
Figure 6—source data 3
Rspo3 deletion leads to Wnt signaling activation in vitro.
Representative image of actin by Western analysis in WT and Rspo-/- MEFs treated w/wo Wnt3a (n=7).
- https://cdn.elifesciences.org/articles/84171/elife-84171-fig6-data3-v2.zip
-
Figure 6—source data 4
Rspo3 deletion leads to Wnt signaling activation in vitro.
Representative image of Tcf1 by Western analysis in WT and Rspo-/- MEFs treated w/wo Wnt3a (n=3).
- https://cdn.elifesciences.org/articles/84171/elife-84171-fig6-data4-v2.zip
-
Figure 6—source data 5
Rspo3 deletion leads to Wnt signaling activation in vitro.
Representative image of Gapdh by Western analysis in WT and Rspo-/- MEFs treated w/wo Wnt3a (n=3).
- https://cdn.elifesciences.org/articles/84171/elife-84171-fig6-data5-v2.zip
Rspo3 haplo-insufficiency and deletion impair Dkk1-Wnt inhibitory activity
Activation of canonical Wnt signaling results from changes in endogenous activators and/or inhibitors levels and/or their activity. Interestingly, we found that Wnt3a significantly decreased the expression of Rspo3 in WT MEFs, whereas it is significantly increased by Dkk1 (Figure 7a). Based on our results with Rspo3+/- and Rspo3-/- cells, this raised the possibility that Rspo3 participates in a feedback loop that tones down canonical Wnt activity. As shown in +/-, we found that the ability of Dkk1 to block Wnt3a-dependent activation of canonical Wnt signaling was significantly impaired in the absence of Rspo3: whereas in WT MEFs 50% reduction in the reporter activity was achieved by 50 ng/mL Dkk1, a dose of 400 ng/mL Dkk1 (8 x higher) was needed to obtain the same level of inhibition in Rspo3 null MEFs (Figure 7b). Impairment of Dkk1 efficacy in Rspo3 null MEFs was also confirmed by pLrp6 and β-catenin protein levels (Figure 7c). To determine whether this relationship between Rspo3 levels and Dkk1 efficacy was also happening in vivo, we crossed Rspo3+/- mice with mice expressing high levels of Dkk1 in OBs (Dkk1-Tg mice), which exhibit impaired canonical Wnt signaling and low trabecular bone mass due to decreased bone formation (Li et al., 2006; Guo et al., 2010). Supporting our in vitro data, Rspo3 haplo-insufficiency counteracted the negative effect of OB-targeted overexpression of Dkk1 on trabecular bone mass (Figure 7d and e and Supplementary file 7).
Figure 7
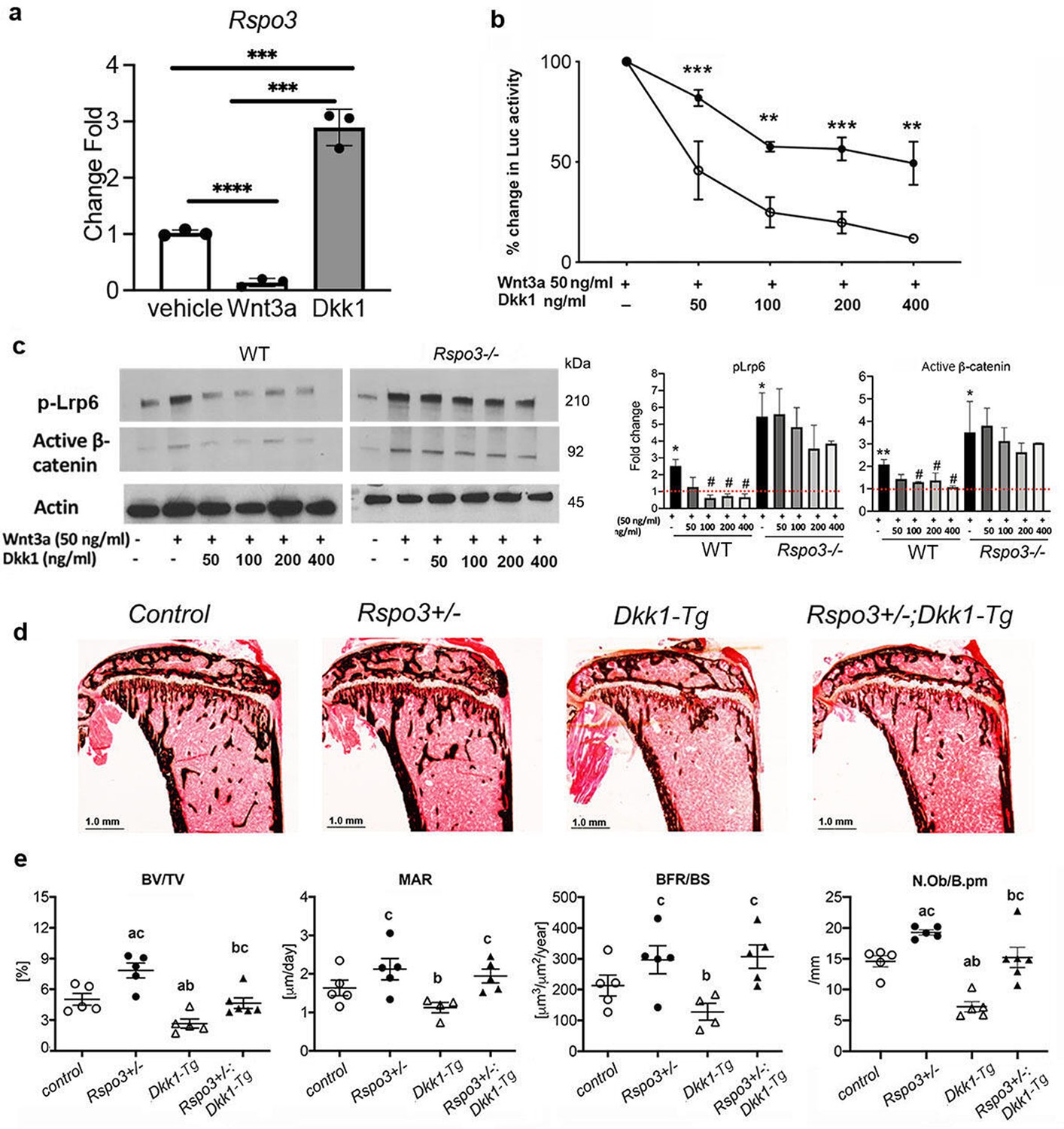
Rspo3 deletion/ haplo-insufficiency impairs Dkk1 efficacy.
(a) Regulation of Rspo3 by Wnt3a and Dkk1 in WT MEFs (n=3) Data are the mean ± SEM. ***p<0.0005, ****p<0.0001, by Student’s t-test. (b) Luciferase assay in WT and Rspo-/- MEFs treated w/wo Wnt3a and increasing doses of Dkk1 (n=3–4). Data are the mean ± SEM **p<0.005, ***<p< 0.0005 compared with vehicle same genotype by unpaired Student’s t-test. (c) Representative images and quantification of active β-catenin and pLrp6 by Western analysis in WT and Rspo3-/- MEFs treated w/wo Wnt3a and increasing doses of Dkk1 (n=3). Data are the mean ± SEM *p<0.05, **p<0.005 vs WT vehicle, # p<0.05, vs Wnt3a–treated same genotype by unpaired Student’s t-test. (d) Representative images of Von Kossa staining in 6 wk old female mice (n=5-6). Scale bars = 1.0 mm. (e) BV/TV, MAR, BFR/BS, and N.Ob./B.pm by histomorphometric analysis in females (n=5–6). Data show all samples and the mean ± SEM a=p<0.05 compared with control mice, b=p < 0.05 compared with Rspo3+/- mice, c=p<0.05 compared with Dkk1-Tg mice by two-way ANOVA followed by Fisher’s LSD test.
-
Figure 7—source data 1
Rspo3 deletion/ haplo-insufficiency impairs Dkk1 efficacy.
Representative image of active β-catenin and pLrp6 by Western analysis in WT and Rspo3-/- MEFs treated w/wo Wnt3a and increasing doses of Dkk1 (n=3).
- https://cdn.elifesciences.org/articles/84171/elife-84171-fig7-data1-v2.zip
-
Figure 7—source data 2
Rspo3 deletion/ haplo-insufficiency impairs Dkk1 efficacy.
- https://cdn.elifesciences.org/articles/84171/elife-84171-fig7-data2-v2.zip
-
Figure 7—source data 3
Rspo3 deletion/ haplo-insufficiency impairs Dkk1 efficacy.
Representative image of actin by Western analysis in WT and Rspo3-/- MEFs treated w/wo Wnt3a and increasing doses of Dkk1 (n=3).
- https://cdn.elifesciences.org/articles/84171/elife-84171-fig7-data3-v2.zip
Rspo3 deletion enhances Erk signaling to increase pLrp6 and stabilize β-catenin
Since both Rspo3 haplo-insufficiency and its deletion led toβ-catenin stabilization, we asked whether Rspo3 might regulate other signaling pathways which in turn can stabilize β-catenin, stimulating osteoblastogenesis and counteracting Dkk1 efficacy. It has been proposed that Rspo3 binding to Lgr4 inhibits Erk phosphorylation (pErk) (Lee et al., 2020; Zhang et al., 2017; Zhou et al., 2007). We, therefore, investigated whether Erk signaling, known to activate Wnt signaling and to regulate OB differentiation and bone mass (Červenka et al., 2011; Gortazar et al., 2013; Kim et al., 2019; Krejci et al., 2012), was affected by the absence of Rspo3 in MEFs. Rspo3 deficiency led to a clear and significant increase in pErk basal levels (Figure 8a). Inhibition of pErk by the specific Erk inhibitor U0126, in Rspo3-/- cells led to a significant decrease in active β-catenin levels in both steady state and Wnt3a-stimulated cultures. A similar effect was also seen for the levels of pLrp6 (Figure 8a). Confirming these findings, the increase in the expression of the canonical Wnt signaling target genes Tcf7 and Axin2 in Rspo3 null MEFs was also partially rescued by blocking Erk signaling (Figure 8b). In contrast, the Erk inhibitor U0126 significantly decreased both the basal and the Wnt3a-dependent increase in pErk in WT cells but failed to affect significantly active β-catenin levels. Thus, the stabilization of β-catenin seen in the absence of Rspo3 is due, at least in part, to activation of the Erk pathway.
Figure 8
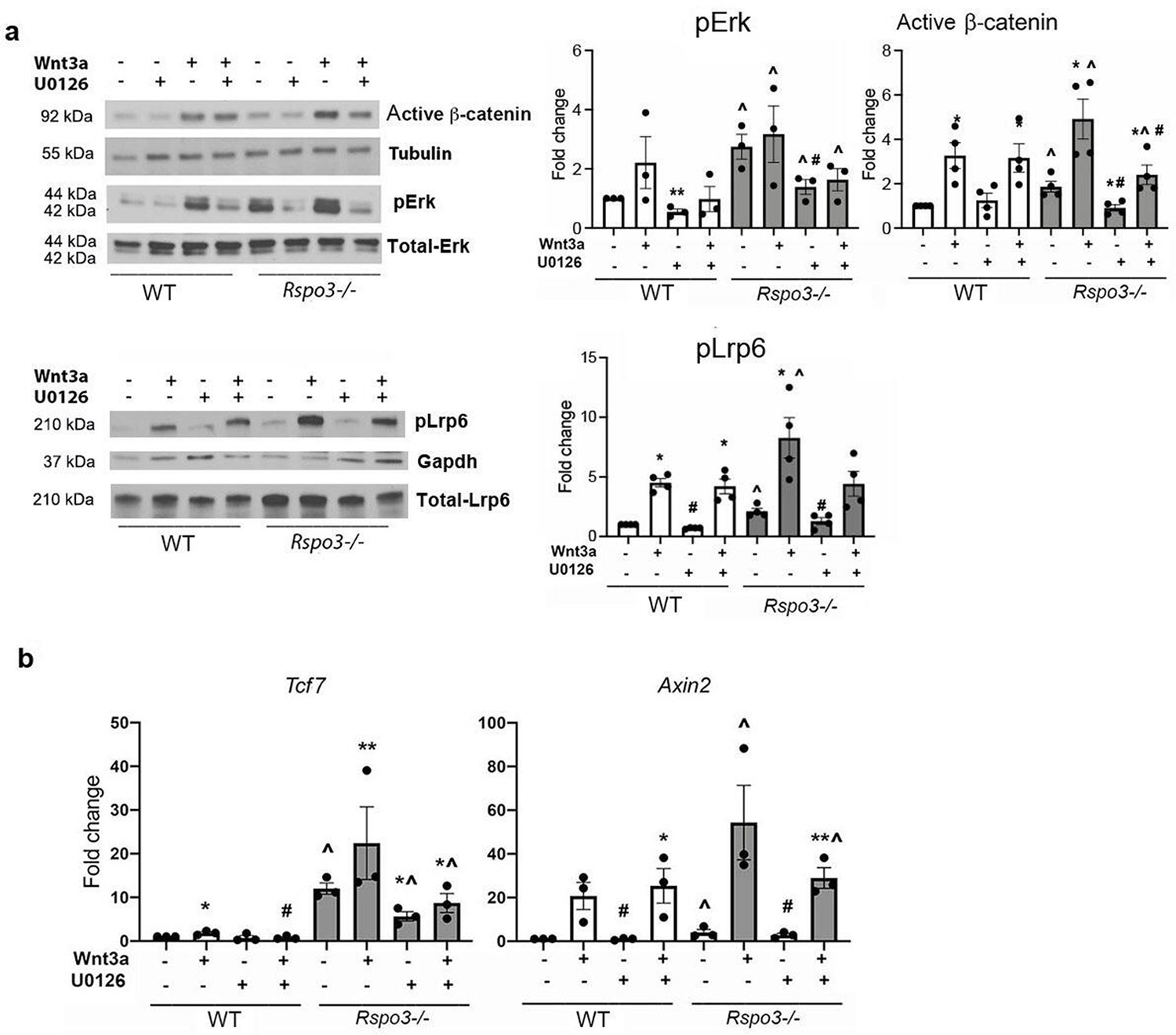
Erk signaling is involved in the Wnt signaling activation seen in the absence of Rspo3.
(a) Representative images and quantification of pERK, active β-catenin and pLrp6 levels by western analysis in WT and Rspo3-/- MEFs treated w/wo w/wo Wnt3a and U0126. (b) Expression of Wnt target genes in WT and Rspo3-/- MEFs treated w/wo Wnt3a and U0126. Data show all samples and the mean ± SEM (n=3–4) *p<0.05, **p<0.005 vs vehicle of the same genotype, ^=p<0.05 vs WT vehicle and # p<0.05 vs Wnt3a–treated same genotype by unpaired Student’s t-test.
-
Figure 8—source data 1
Erk signaling is involved in the Wnt signaling activation seen in the absence of Rspo3.
Representative image of active β-catenin in WT and Rspo3-/- MEFs treated w/wo w/wo Wnt3a and U0126 (n=3–4).
- https://cdn.elifesciences.org/articles/84171/elife-84171-fig8-data1-v2.zip
-
Figure 8—source data 2
Erk signaling is involved in the Wnt signaling activation seen in the absence of Rspo3.
Representative image of tubulin levels by western analysis in WT and Rspo3-/- MEFs treated w/wo w/wo Wnt3a and U0126 (n=3–4).
- https://cdn.elifesciences.org/articles/84171/elife-84171-fig8-data2-v2.zip
-
Figure 8—source data 3
Erk signaling is involved in the Wnt signaling activation seen in the absence of Rspo3.
Representative image of pERK levels by western analysis in WT and Rspo3-/- MEFs treated w/wo w/wo Wnt3a and U0126 (n=3–4).
- https://cdn.elifesciences.org/articles/84171/elife-84171-fig8-data3-v2.zip
-
Figure 8—source data 4
Erk signaling is involved in the Wnt signaling activation seen in the absence of Rspo3.
Representative image of total ERK levels by western analysis in WT and Rspo3-/- MEFs treated w/wo w/wo Wnt3a and U0126 (n=3–4).
- https://cdn.elifesciences.org/articles/84171/elife-84171-fig8-data4-v2.zip
-
Figure 8—source data 5
Erk signaling is involved in the Wnt signaling activation seen in the absence of Rspo3.
Representative image of pLrp6 levels by western analysis in WT and Rspo3-/- MEFs treated w/wo w/wo Wnt3a and U0126 (n=3–4).
- https://cdn.elifesciences.org/articles/84171/elife-84171-fig8-data5-v2.zip
-
Figure 8—source data 6
Erk signaling is involved in the Wnt signaling activation seen in the absence of Rspo3.
Representative image of total Lrp6 levels by western analysis in WT and Rspo3-/- MEFs treated w/wo w/wo Wnt3a and U0126 (n=3–4).
- https://cdn.elifesciences.org/articles/84171/elife-84171-fig8-data6-v2.zip
-
Figure 8—source data 7
Erk signaling is involved in the Wnt signaling activation seen in the absence of Rspo3.
Representative image of Gapdh levels by western analysis in WT and Rspo3-/- MEFs treated w/wo w/wo Wnt3a and U0126 (n=3–4).
- https://cdn.elifesciences.org/articles/84171/elife-84171-fig8-data7-v2.zip
Consistent with these findings, the increase in CFU-F and CFU-OB formation seen in Rspo3+/- BMSCs was significantly decreased by blocking Erk signaling (Figure 9). Not surprisingly, knowing the role of Erk signaling activation in osteoblastogenesis, a significant decrease in CFU-OB, but not in CFU-F, formation was also seen in WT cells treated with U0126 (Figure 9).
Figure 9
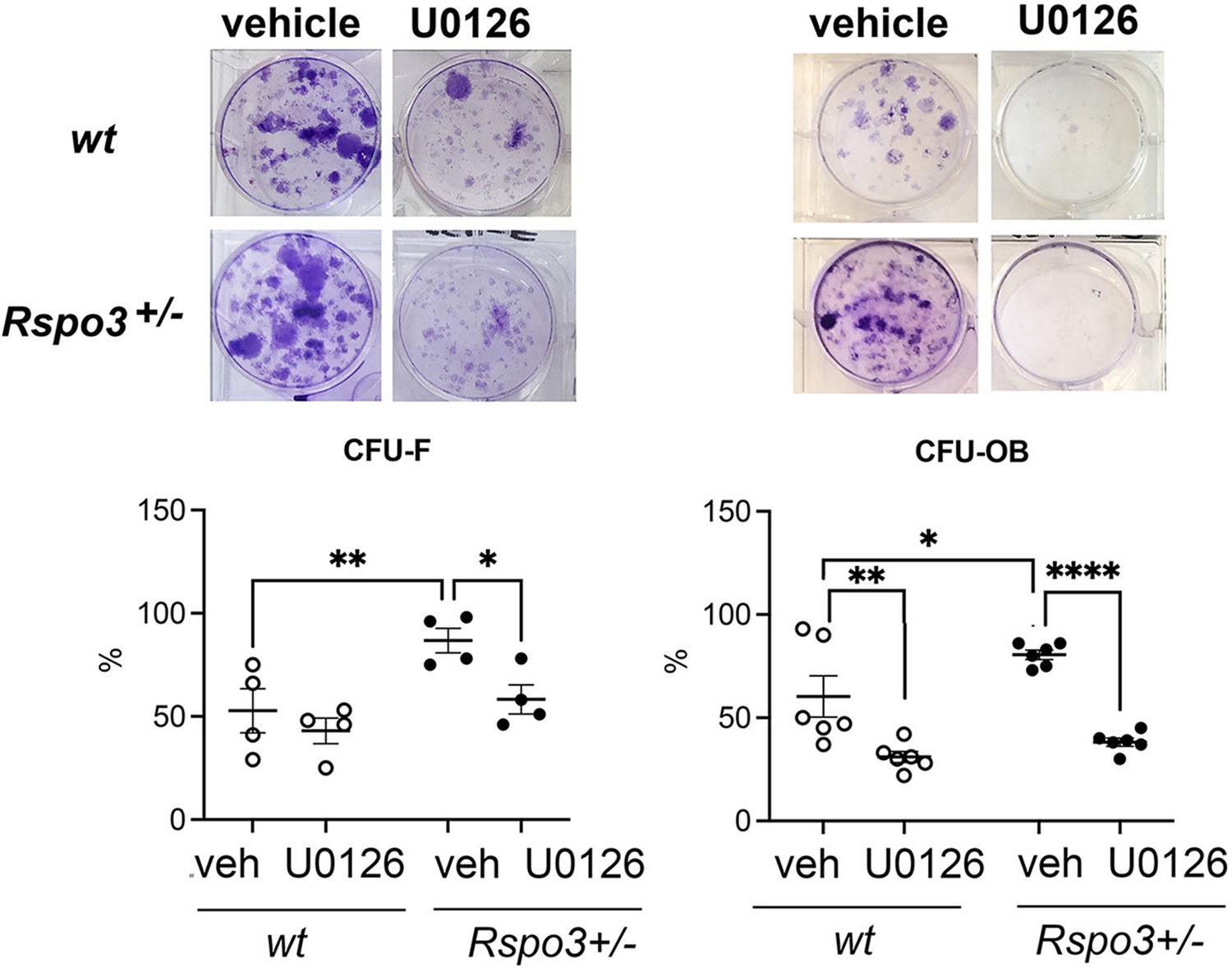
Effect of Erk signaling inhibition of CFU assays.
Representative images of CFU-F and CFU-OB and quantification in WT and Rspo3+/- mice treated with and w/o U0126. Data show all samples and the mean ± SEM (n=3) *=p<0.05, **=p<0.005, ****=p<0.0001 two-way ANOVA followed by Fisher’s LSD test.
Discussion
Wnt signaling is central to skeletal development and homeostasis in health and disease (Baron and Kneissel, 2013). Understanding the biological mechanisms by which this signal operates is, therefore, of both scientific and clinical interest. R-Spondins, classically considered as positive modulators of Wnt signaling, play an important role in normal development of several tissues and organs, including bone, and are implicated in human diseases (de Lau et al., 2012; Knight and Hankenson, 2014; Nagano, 2019; Raslan and Yoon, 2019; Shi et al., 2016). Among the 4 R-Spondins, GWAS studies in humans have shown that RSPO3 might be specifically involved in bone homeostasis due to the strong association between RSPO3 common variants and bone mineral density and fracture rate (Duncan et al., 2011; Estrada et al., 2012; Medina-Gomez et al., 2012; Richards et al., 2008; Richards et al., 2012; Nilsson et al., 2022). Our results demonstrate, through several independent lines of genetic in vivo and in vitro experiments, that, counter-intuitively, decreasing Rspo3 levels globally or specifically in Runx2+ OB precursors leads to increased trabecular bone formation and high bone mass in the appendicular skeleton, mainly driven by increased number of OB progenitors and OBs as well as an increase in their bone forming activity. Unexpectedly though, and as recently reported by Nilsson et al., 2021, we also found that while the axial skeleton is only moderately affected in the haplo-insufficient mice, vertebrae are affected in the opposite manner, that is with a decrease in trabecular bone mass, after OB precursor-targeted deletion of Rspo3. Although it is not possible to identify the mechanism(s) responsible for these differential responses of the skeleton, it is worth mentioning here that axial and appendicular skeleton have different embryonic origin (paraxial mesoderm and later plate mesoderm, respectively) (Berendsen and Olsen, 2015) and that a new population of skeletal stem cells has been recently identified in the vertebrae (vSSC) that is absent from the long bones (Ay et al., 2022), raising the possibility that distinct populations of stem cells and non-stem progenitors in the vertebrae and in the long bones might respond differentially to stimuli. These findings underline the complexity of Wnt signaling regulation of bone homeostasis and support the hypothesis that different bone microenvironments, their embryonic origin, their differential exposure and response to mechanical cues, and the differential expression levels of receptors and co-receptors as well as of agonists and antagonists that balance Wnt signaling activity, might induce specific and distinct skeletal responses. Nevertheless, our findings show that skeletal phenotypes should always be investigated in both the appendicular and axial skeleton, as these two regions may respond differently. In contrast, Nilsson et al. performed their studies only in vertebrae. Furthermore, this is a trabecular bone-rich skeletal site, but the DEXA studies that led to the identification of a link between RSPO3 variants and bone fragility in humans were performed in the distal forearm, a cortical-rich appendicular skeletal site (Nilsson et al., 2021; Nilsson et al., 2022). Furthermore, both groups found that cortical bone is not significantly affected by Rspo3 haplo-insufficiency or deletion in mice, confirming that trabecular and cortical bone are differentially regulated (Movérare-Skrtic et al., 2014; Kiper et al., 2016) and further underlining the biologic complexity of Wnt signaling regulation of bone homeostasis.
Thus, our studies reveal a novel and unexpected role of Rspo3 in the Wnt signaling machinery and bone homeostasis. Given the unexpected nature of our observations it is important to summarize here the multiple independent in vivo and in vitro experiments that unequivocally support the novel concept that decreasing the levels of Rspo3 can, at least in vitro and in the cellular environment of the appendicular skeleton’s trabecular bone, result in the activation of canonical Wnt signaling, increasing osteoblast differentiation and bone formation, and thereby bone mass: 1/Global haplo-insufficient or Runx2-Cre driven depletion of Rspo3 increased bone mass, OB numbers, and BFR in mice long bones; 2/Ex vivo, haplo-insufficient BMSCs exhibited increased OB differentiation and canonical Wnt signaling; 3/Rspo3 null MEFs also exhibited an enhanced Wnt signaling activity; 4/Basal levels of Erk phosphorylation were high in these cells and inhibition of Erk phosphorylation prevented the activation of Wnt signaling; 5/Rspo3 haplo-insufficiency rescued partially the inhibition of Wnt signaling induced by Dkk1 in vitro and in vivo.
In vitro studies have demonstrated that overexpression of and treatment with Rspo1 or Rspo2 enhance Wnt ligand-mediated OB differentiation (Knight and Hankenson, 2014; Lu et al., 2008; Knight et al., 2018). The literature and our own in vitro studies confirm that Rspo3 can be, as expected, a co-activator of canonical Wnt signaling in the Topflash assay, potentiating Wnt3a-dependent activation of canonical Wnt signaling in cellular assays (Yoon and Lee, 2012 and Figure 6—figure supplement 1). However, the expression of canonical Wnt target genes and the levels of pLrp6, activated β-catenin and Tcf1 were also markedly increased in Rspo3 haplo-insufficient BMSCs and in Rspo3-null MEFs, independent of the addition of Wnt3a to the assays, indicating that, counter-intuitively, the decrease or absence of Rspo3 activates mechanisms that favor β-catenin-dependent signaling. This is in contrast with the data reported by Nilsson et al. who show a decrease in Dkk1 and Sost, but this was done in the vertebrae of Runx2-CreRspo3fl/fl mice, where BFR is in fact decreased, or a decrease in Tcf7 and Lef1 expression in calvarial OBs (cortical-derived cells Azzolin et al., 2014) treated with and without Tamoxifen to induce Rspo3 deletion in vitro (Nilsson et al., 2021), when cortical bone is in fact not affected. Although we have not analyzed mRNA expression in the vertebrae or calvaria, our extensive set of data (Topflash, Q-RTPCR, Western blot analysis for several Wnt signaling downstream proteins) using different cells (BMSC and MEFs) and long bones clearly show an activation of Wnt signaling with Rspo3 haploinsufficiency or deletion.
Another new finding from our studies is that Rspo3 expression is strongly repressed by Wnt3a and increased by Dkk1. This, together with the results discussed above, suggests that Rspo3 may provide a negative feedback-loop helping to balance canonical Wnt activity. Dkk1 efficacy in blocking Wnt3a-dependent activation of canonical Wnt signaling is significantly impaired in the absence of Rspo3 and Rspo3 haplo-insufficiency antagonizes the inhibition of bone formation induced by OB-targeted expression of Dkk1, confirming in vivo that Rspo3 haplo-insufficiency counteracts the OB-Dkk1 dependent function. Our results show that this is the result of intracellular changes in alternative pathways regulated by Rspo3. Indeed, we found that the Erk signaling pathway is activated and the basal level of Lrp6 phosphorylation enhanced by Rspo3 depletion. Thus, the activation of the Wnt signaling pathway when Rspo3 is expressed at low levels results from intra-cellular changes, distal to the receptor complexes. Our data and that of others (Lee et al., 2020) suggest that there are alternate Rspo3-mediated signaling mechanisms, separate from the Fzd/Lrp/β-catenin Wnt pathway, including the Wnt/PCP signaling (Glinka et al., 2011; Hao et al., 2012) and that these events can in turn regulate Wnt signaling intra-cellularly, such that β-catenin can be stabilized independent of the proximal activation of the canonical Wnt signaling machinery through changes in other signaling pathways (Baron and Kneissel, 2013; Krejci et al., 2012; Azzolin et al., 2014).
Thus, deletion of Rspo3 enhances Erk signaling which, in turn, stabilizes β−catenin independent of the canonical Wnt signaling receptor complex. In turn, this has a positive effect on OB differentiation (Červenka et al., 2011; Kim et al., 2019; Krejci et al., 2012). Our finding that the increase in Wnt signaling activation and the OB potential of MSCs seen in the absence of Rspo3 is abrogated by blocking Erk signaling confirms that the activation of Erk signaling associated with Rspo3 deficiency is responsible, at least in part, for the observed Wnt signaling activation.
Supporting our findings, in vitro studies have shown that Rspo3 silencing activates Erk signaling downstream of Lgr4, leading to increased OB differentiation of human adipose-derived stem cells (Zhang et al., 2017). This study did not however establish a connection between Erk and Wnt signaling. Although Lgrs function as receptors for Rspos, and Rspos/Lgrs interactions enhance Wnt signaling by inducing the clearance of Rnf43 and Znrf3 (Ruffner et al., 2012; Wang et al., 2013; de Lau et al., 2011), there is also strong evidence that Rspos/Lgrs interaction can activate distinct signaling cascades that can affect bone, including the cAMP/PKA/Creb signaling pathway in Lgr4 null mice (Luo et al., 2009) and the Erk signaling cascade (Zhang et al., 2017; Xu et al., 2016; Lin et al., 2019; Vieira et al., 2015).
Based on our observations, we propose that Rspo3 has a dual mode of action to regulate canonical Wnt signaling and bone formation. This duality is based on the regulation of two distinct signaling cascades and their crosstalk: Rspo3 functions via both the Lgr/Rnf43/Znrf3 and the Lgr/Erk axes, and while activation of the Lgr/Rnf43/Znrf3 axis boosts Wnt signaling strength by the membrane clearance of Rnf43/Znrf3 and subsequent stabilization of Fzd receptors, binding of Rspo3 to Lgr impairs Erk signaling, preventing Erk signaling activation and further stabilization of β−catenin (Figure 10). Thus, haplo-insufficiency and deletion of Rspo3 would dampen Wnt signaling at the cell surface by preventing the Rnf43/Znrf3 effects while intracellularly enhancing pLrp6 and β−catenin stabilization, via Erk phosphorylation, overcompensating the decrease in Rspo3-dependent Lrp5/6 receptors-dependent Wnt activation in OBs and their progenitors. Because activation of the Lgr/Rnf43/Znrf3 cascade is not exclusively dependent on Rspo3, deletion of Rspo3 would only hinder canonical Wnt signaling partially. In contrast, lack of Rspo3 promotes the Lgr/Erk cascade, to not only enhance β−catenin stabilization (Figure 10) but also regulate OB differentiation and bone formation. This model also explains the observed loss of Dkk1 efficacy in inhibiting Wnt signaling: the Erk-dependent stabilization of β−catenin being independent of Wnt receptor activation, Dkk1, which binds to the LRP5/6 receptors, cannot dampen the activation of downstream events as they are independent of the LRP5/6-Fzd receptor complex.
Figure 10
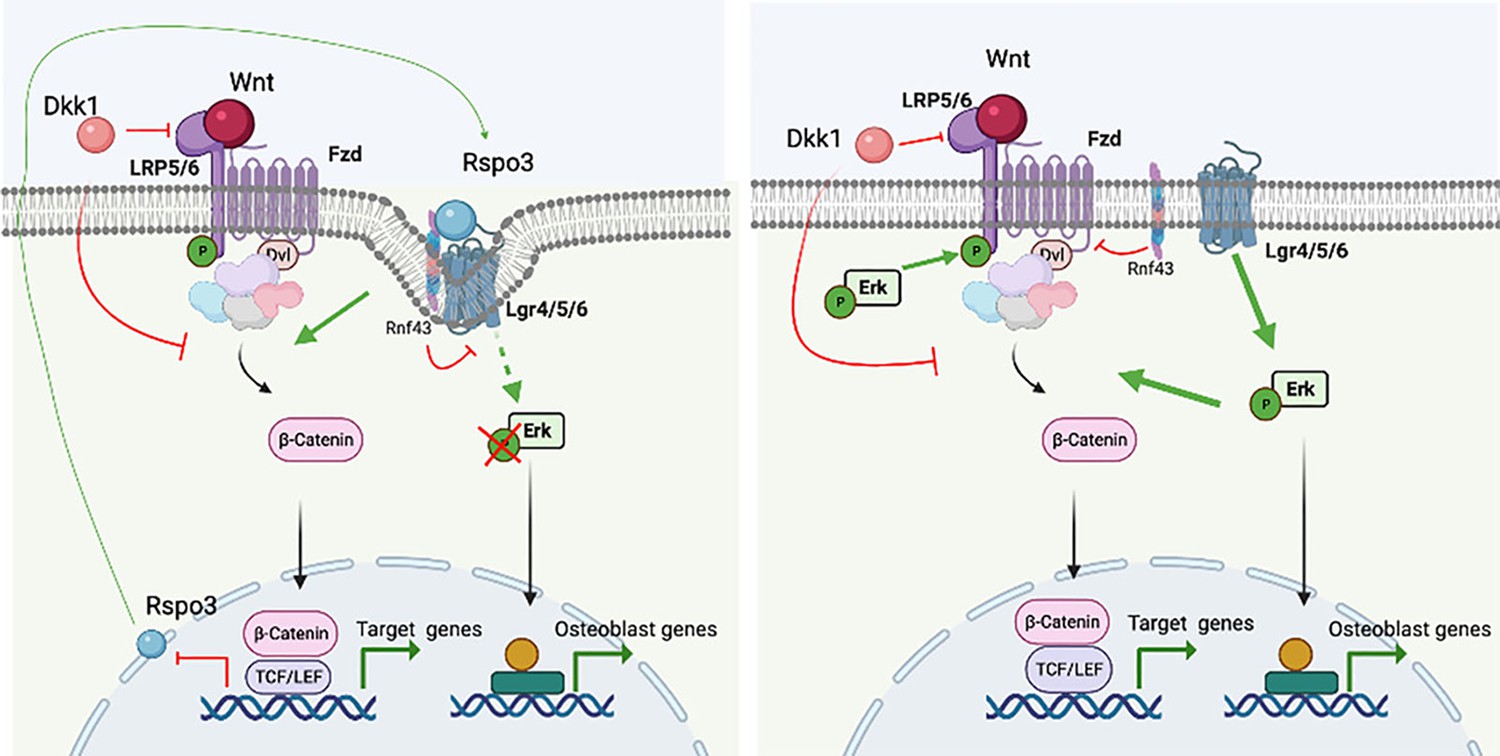
Proposed model.
Rspo3 has a dual mode of action to regulate canonical Wnt signaling and thereby bone formation. This duality is based on the regulation of two distinct signaling cascades and their crosstalk: Rspo3 functions via both the Lgr/Rnf43/Znrf3 and the Lgr/Erk axes. In the presence of Rspo3, the Rspo3/Lgr/Rnf43/Znrf3 axis boosts Wnt signaling strengths by the membrane clearance of Rnf43/Znrf3 and subsequent stabilization of Fzd receptors. In addition, binding of Rspo3 to Lgr impairs Erk signaling likely due to the membrane clearance of the Lgr/Rnf43/Znrf3 receptors, preventing Erk signaling activation. Deletion of Rspo3 would dampen Wnt signaling at the cell surface by preventing the Rnf43/Znrf3 effects while promoting Erk activation downstream of Lgr receptors in turn enhancing Lrp5/6 phosphorylation and β−catenin stabilization intracellularly, which has a more potent effect and overcompensates the decrease in Rspo3-dependent proximal Wnt activation in osteoblasts and their progenitors. Figure created with Biorender.
Supporting the fact that Rspo3 can also regulate Wnt-independent pathways, a recent study has suggested that Rspo3 acts as an antagonist to BMPR1A, inhibiting BMP signaling during development (Lee et al., 2020). Thus, our observations may be due, at least in part, to changes (activation) in BMP signaling, which in turn could lead to the observed increase in pErk (Zhou et al., 2007). Although this remains a possibility, it seems unlikely. First, in contrast to our observations here, BMP activation in the adult skeleton has been linked to activation of non-canonical Wnt signaling, increased Sost expression, and bone resorption (Kiper et al., 2016; Kamiya et al., 2016). Second, several studies have shown that activation of BMP signaling in the osteoblast lineage has a negative impact on bone formation and bone mass (Kamiya et al., 2010; Kamiya et al., 2008; Ko et al., 2017; Shi et al., 2018).
In conclusion, our studies indicate that Rspo3 is required for skeletal homeostasis and show for the first time that axial and appendicular skeleton are differentially regulated by this Wnt signaling regulator. The molecular basis for this differential regulation has not been elucidated but might include different embryologic origin, differences in local micro-environment, or differences in mechanical loading, for instance. Nevertheless, our in vitro studies suggest that Rspo3 regulates bone formation through its interaction not only with the Wnt receptor machinery, as a positive co-activator, but also with other signaling pathways that affect β−catenin stability independent of the receptor complexes. Consequently, its deletion removes a co-activator of Wnt signaling, potentially decreasing bone formation, but also promotes Erk signaling activation, increasing β−catenin stability sufficiently to enhance bone formation and increase bone mass in the appendicular skeleton. Furthermore, because Rspo3 depletion increases Dkk1 and Dkk1 increases Rspo3 expression, this study also reveals a novel Rspo3-dependent negative feedback Wnt signaling regulatory loop. These findings have important implications for understanding the pleiotropic functions of Rspos and Wnt signaling in skeletal homeostasis and the mechanisms that regulate bone mass.
Materials and methods
Biological variables and reproducibility
Request a detailed protocolTo conduct the proposed studies, we insured strict application of scientific methods that supports robust and unbiased design, analysis, interpretation, and reporting of results, and sufficient information for all studies undertaken. In vivo analysis was performed in males and females, at different ages, and in the axial and appendicular skeleton. Rspo3+/- mice and Rspo3-OB-cKO mice with targeted deletion of Rspo3 in osteoprogenitors (Runx2-Cre) were investigated. Given that our studies with Rspo3 mutant mice do not identify sex differences, and the osteoblast specific Dkk1 transgenic (Dkk1-Tg) mice did not show sex differences (Guo et al., 2010), studies including crossing with the two strains were performed in female mice. To avoid bias, data were collected in a blinded fashion, in that the observer was unaware of the experimental groups and more than one individual performed key studies. In vivo studies were performed with n=4–10 mice per genotype. We based this number on a priori calculations for power to detect differences in the primary outcome skeletal phenotype by histomorphometric analysis. We found that based on three independent methods for measuring the skeletal phenotype 4–10 mice per group provide enough power (80%–90%) to find statistical significance at p<0.05. Ex vivo and in vitro studies involved at least three biological replicates per group/treatment.
Animals
Rspo3+/- and Rspo3fl mice were provided by Dr. Christof Niehrs (DKFZ-ZMBH Alliance, Germany) and were previously described (Kazanskaya et al., 2008). The osteoblast specific Dkk1 transgenic (Dkk1-Tg) mice, expressing high levels of Dkk1 in osteoblasts, were generously provided by Dr. Guo and Dr. Kronenberg (Massachusetts General Hospital, MA, USA) (Guo et al., 2010). Runx2-Cre mice were provided by Dr. Tuckermann (Ulm University, Ulm, Germany) (Rauch et al., 2010). All experiments were performed with age- and sex-matched littermates. All animals are in the C57BL/6 background and were housed in the Harvard Center for Comparative Medicine and all experimental procedures were approved by the Harvard University Institutional Animal Care and Use Committee. The protocol number associated with the ethical approval of the animal work is IS1045.
Skeletal phenotype
Request a detailed protocolFor bone histomorphometric analysis of Rspo3+/- mice, 6, 12, or 18 wk-old mice were injected with 20 mg/kg of calcein and 40 mg/kg of demeclocycline (Sigma Aldrich, St. Louis, MO, USA) 6, 8, or 9 days, respectively and 2 days prior to the sacrifice. For mice with Rspo3 deletion in Runx2+ cells, 8-wk-old mice were injected with 20 mg/kg of calcein and 40 mg/kg of demeclocycline, 8 and 2 days prior to sacrifice and for the experiments with the Dkk1-Tg mice, 6-wk-old mice were injected 6 and 2 days prior to sacrifice. Bone histomorphometric analysis was performed within the proximal tibia under 200× magnification in a 0.9 mm high and 1.3-mm-wide region that was 200 μm away from the growth plate. For vertebra analysis, data were obtained under 200× magnification in a 1.3 mm × 1.8 mm region away from the growth plate to avoid including primary spongiosa. Consecutive sections of the proximal tibia and frontal sections of the vertebral body (4 µm thickness) were stained with von Kossa and Toluidine blue for the analysis of cellular parameters and osteoid. Bone sections were viewed with a Nikon E800 microscope equipped with Olympus DP71 digital camera (RRID:SCR_020326). Images were captured using Olympus CellSens software (RRID:SCR_014551). The OsteoMeasure analyzing software (Osteometrics) was used to generate and calculate the data. Structural parameters bone volume fraction (BV/TV), trabecular thickness (Tb.Th), trabecular number (Tb.N), and trabecular separation (Tb.Sp) were obtained by calculating the average of two different measurements from consecutive sections. The structural, dynamic, and cellular parameters were presented according to the standardized nomenclature (Dempster et al., 2013).
A high-resolution desktop micro-tomographic imaging system (µCT40, Scanco Medical AG, RRID:SCR_017119, Brüttisellen, Switzerland) was used to assess trabecular bone architecture and mineral density in the L5 vertebral body and femur and cortical bone architecture in the diaphysis of the femur or the tibia. Scans were acquired using a 10 µm3 isotropic voxel size, 70 kVP, 114 mAs, and 200ms integration time. μCT scanning and analysis were performed according to recommended guidelines (Bouxsein et al., 2010). Trabecular bone within this region was segmented from soft tissue using a threshold of 400 mg HA/cm3. The Scanco trabecular morphology evaluation script was used to measure trabecular bone volume fraction (Tb.BV/TV, %), trabecular bone mineral density (Tb.BMD, mg HA/cm3), Tb.N (mm–1), Tb.Th (μm), Tb.Sp (μm), structural model index (SMI), and connectivity density (Conn.D, mm–3). Cortical bone was assessed in 500 μm long regions (50 transverse slices) at the femoral mid-diaphysis and tibial diaphysis (2 mm superior to the distal tibiofibular junction). Images within the cortical region of interest were segmented using a threshold of 700 mgHA/cm3 and then the Scanco midshaft evaluation script was used to measure total cross-sectional area Total area (Tt.Ar, mm2), cortical bone area (Ct.Ar, mm2), medullary area (Ma.Ar, mm2), bone area fraction (Ct.Ar/Tt.Ar, %), cortical tissue mineral density (Ct.TMD, mgHA/cm3), cortical thickness (Ct.Th, mm).
Flow cytometry
Request a detailed protocolBone marrow was analyzed by flow cytometry as previously described (Schepers et al., 2013). Briefly, bone marrow cells were flushed from femurs and tibiae of 6–8 wk old WT or Rspo3+/- mice and washed with Hank’s Balanced Salt Solution (HBSS). Residual bone samples were further digested in 3 mg/ml type I collagenase (Worthington Biochemical Corp., Lakewood, NJ 08701, USA) for 1 hr at 37 °C and released cells were mixed with flushed bone marrow cells. Cells were stained with LIVE/DEAD Fixable Aqua Dead Cell Stain Kit (Thermo Fisher Scientific, Waltham, MA, USA), AF700-anti-lineage (RRID:AB_2715571), PE-anti-CD31 (RRID:AB_2572182), PB-anti-Sca-1 (RRID:AB_2143237), anti- Cy7APC-CD45 (RRID:AB_2860726), and biotin-anti-CD51 (RRID:AB_313073) with streptavidin-APC antibodies (BioLegend, San Diego, CA, USA). Cells were analyzed on a BD FACS ARIAII (RRID:SCR_018091) upon exclusion of dead cells.
Bone marrow stromal cells and calvarial osteoblasts
Request a detailed protocolBone marrow cells were flushed from femurs and tibiae of 6–8 wk-old WT or Rspo3+/- mice and cultured in DMEM supplemented with 10% fetal bovine serum (FBS) and 1% penicillin (100 U/ml) and streptomycin (100 µg/ml) for 3 days (GIBCO, Thermo Fisher Scientific, Waltham, MA, USA). Adherent MSCs were counted, re-plated onto at a 5,000 /cm2 density, and RNA or protein isolated 3 days later. For colony forming unit assays, flushed bone marrow stromal cells were plated (3X106/6 wells) for CFU- (Fibroblast) F and CFU-OB assays. Cells were treated either with recombinant mouse Rspo3 (100 ng/ml) (all from R&D system, Minneapolis, MN, USA) or U0126 (10 μM) (Selleckchem, Houston TX, USA). CFU-F was detected by staining with 0.2% crystal violet in 2% ethanol for 1 hr after 10 days in culture and CFU-OB was detected by alkaline phosphatase activity with Napthol AS-MX, n,n-dimethylformamide and Fast Blue RR salt (Sigma Aldrich, St. Louis, MO, USA) after 12 days in culture with OB differentiation medium: DMEM supplemented with 10%FBS, 1% penicillin (100 U/ml) and streptomycin (100 µg/ml) (GIBCO, Thermo Fisher Scientific, Waltham, MA, USA), 5 μg/ml ascorbic acid and 10 mM β-glycerolphospahte (Sigma-Aldrich, St. Louis, MO, USA). Calvarial OBs were isolated from 1 to 3-day-old pups via serial enzymatic digestions and cultured as previously reported (Chen et al., 2019).
Mouse embryonic fibroblasts (MEFs) primary culture
Request a detailed protocolTo obtain WT and Rspo3-/- MEFs, Rspo3+/- males and females were crossed, and the morning of vaginal plug detection was defined as embryonic day (E) 0.5. At E9.5, whole embryos were isolated, washed in PBS, minced in 0.05% trypsin (GIBCO, Thermo Fisher Scientific, Waltham, MA, USA) followed by incubation at 37 C for 10 min. After incubation samples were pipetted to obtain single cell suspension and cells cultured in DMEM supplemented with 10% FBS and 1% penicillin/streptomycin. All the experiments were performed in passage 4–6 of WT or Rspo3-/- MEFs. Cells were treated either 50 or 200 ng/ml of recombinant human Wnt3a, recombinant human Dkk1 (50–400 ng/ml), recombinant human and/or mouse Rspo3 (100 ng/ml) (all from R&D system, Minneapolis, MN, USA) or U0126 (10 μM) (Selleckchem, Houston TX, USA). For TOPflash luciferase reporter assay, cells were transiently co-transfected with 400 ng TOPflash-luc reporter plasmid (RRID:Addgene_12456) and 10 ng control pCMV-Renilla-luciferase (RRID:Addgene_45968, Promega, Madison WI, USA) using Lipofectamine 2000 (Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA) according to the manufacturers protocol. Cells were subjected to serum starvation in DMEM containing 1% FBS overnight. Cells were subsequently treated with recombinant human Wnt3a in the presence and absence of increasing concentration of Dkk1 for 24 hr followed by luciferase assay using the Dual-Glo Assay system (Promega, Madison WI, USA) according to the manufacturers protocol. Data were normalized by Renilla-firefly activity and presented as fold change compared with control group.
Osteoclast primary culture and mix-matched co-cultures
Request a detailed protocolMurine bone marrow macrophages were isolated from bone marrow flushed tibiae and femurs of WT and Rspo3+/- mice at 6–8 wk-old as described previously (Chen et al., 2019). Briefly, cells were cultured in complete α-MEM with 30 ng/ml macrophage colony-stimulating factor M-CSF (R&D system, Minneapolis, MN, USA) in suspension culture dish to which stromal cells and lymphoid cells cannot adhere, at 37 °C for 2–3 days. For osteoclast generation, cells were cultured in 30 ng/ml M-CSF and 10 ng/ml RANKL (R&D systems, Minneapolis, MN, USA). For co-culture experiments, mouse calvarial osteoblasts were isolated from newborn WT and Rspo3+/- as previously reported (Movérare-Skrtic et al., 2014; Chen et al., 2019) and seeded in 96-well plates (2.000 cells/well) in complete osteogenic α-MEM containing 100 nM Vitamin D3 and 1 µM prostaglandin E2 (Enzo Life Science, Farmingdale, NY, USA). After 3 days, 10,000 BMM from WT and Rspo3+/- mice at 6–8 week-old mice were added per well and cocultured for 9 days in complete osteogenic α-MEM. Tartrate-resistant acid phosphatase (TRAP) staining was performed to evaluate the number of osteoclasts according to the manufacture’s protocol (Sigma-Aldrich, St. Louis, MO, USA).
Western Blot analysis
Request a detailed protocolFive μg of total proteins were resolved by SDS-PAGE under reducing conditions. Immunodetection was performed with antibodies specific to: Active β-catenin, phosphorylated (p) Lrp6, p-Erk, total Erk, Tcf1, Lrp6, and Tubulin [(CST8814, RRID:AB_11127203) (CST2568, RRID:AB_2139327) (CST9101, RRID:AB_331646), (CST9102, RRID:AB_330744), (CST2203, RRID:AB_2199302), (CST3395, RRID:AB_1950408), (CST2125, RRID:AB_2619646) Cell Signaling, Beverly, MA, USA] GAPDH and Actin [(SC32233, RRID:AB_627678) and (SC47778, RRID:AB_626632) Santa Cruz, Santa Cruz, CA, USA]. Immunoreactivities were assessed using ECL plus kit following the manufacture’s protocol (Perkin Elmer, Waltham, MA, USA). Quantification was performed using Image J (RRID:SCR_003070) Protein levels were normalized to the levels of housekeeping protein or total protein in within the same sample.
Quantitative-real time PCR
Request a detailed protocolTotal RNA was isolated from cells using the RNeasy Mini Kit (Qiagen Germantown, MD, USA) according to the manufacturer’s protocols. Total RNA from cortical bone of WT and Rspo3+/- mice was extracted using Trizol reagent (Invitrogen) followed by RNeasy Micro Kit (Qiagen Germantown, MD, USA) according to the manufacturer’s protocols. cDNA was synthesized using iScript cDNA synthesis kit (BIO-RAD. Hercules, CA, USA) and quantitative real time PCR performed. mRNA levels encoding each gene of interest were normalized for β2M or actin mRNA in the same sample and the relative expression of the genes of interest was determined using the formula of Livak and Schmittgen, 2001. Data are presented as fold change relative to WT cells or animals.
Statistical analysis
Request a detailed protocolData are expressed as the mean ± SEM. All experiments include at least three biological replicates and were done in duplicate or triplicate. Values represent the number of biological replicates. Statistical analysis was conducted using unpaired two-tail Student’s t-test, or two-way ANOVA followed by post-hoc test for multiple comparisons. GraphPad PRISM 9 (RRID:SCR_002798) was also used for statistical analysis. A two-sided p-value of <0.05 was considered as the threshold for statistical significance.
Data availability
All data generated or analysed during this study are included in the manuscript and supporting file.
References
-
R-spondin3 is required for mouse placental developmentDevelopmental Biology 301:218–226.https://doi.org/10.1016/j.ydbio.2006.08.018
-
ConferenceDiscovery of a vertebral stem cell driving spine metastasisJournal of Bone and Mineral Research, 2022 Annual Meeting of the American Society for Bone and Mineral Research.
-
BookSclerostin inhibition in the treatment of osteoporosisIn: Leder BZ, Wein MN, editors. Osteoporosis Pathophysiology and Clinical Management. Springer. pp. 6–19.https://doi.org/10.1007/978-3-319-69287-6_19
-
Guidelines for assessment of bone microstructure in rodents using micro-computed tomographyJournal of Bone and Mineral Research 25:1468–1486.https://doi.org/10.1002/jbmr.141
-
Mitogen-activated protein kinases promote WNT/beta-catenin signaling via phosphorylation of LRP6Molecular and Cellular Biology 31:179–189.https://doi.org/10.1128/MCB.00550-10
-
The R-spondin/lgr5/rnf43 module: regulator of Wnt signal strengthGenes & Development 28:305–316.https://doi.org/10.1101/gad.235473.113
-
Crosstalk between caveolin-1/extracellular signal-regulated kinase (ERK) and β-catenin survival pathways in osteocyte mechanotransductionThe Journal of Biological Chemistry 288:8168–8175.https://doi.org/10.1074/jbc.M112.437921
-
Wnt signaling and bone: lessons from skeletal dysplasias and disordersFrontiers in Endocrinology 11:165.https://doi.org/10.3389/fendo.2020.00165
-
Mutations in R-spondin 4 (RSPO4) underlie inherited anonychiaThe Journal of Investigative Dermatology 128:867–870.https://doi.org/10.1038/sj.jid.5701078
-
Wnt inhibitors DKK1 and SOST are downstream targets of BMP signaling through the type IA receptor (BMPRIA) in osteoblastsJournal of Bone and Mineral Research 25:200–210.https://doi.org/10.1359/jbmr.090806
-
The ERK MAPK pathway is essential for skeletal development and homeostasisInternational Journal of Molecular Sciences 20:E1803.https://doi.org/10.3390/ijms20081803
-
Cortical-bone fragility -- insights from sfrp4 deficiency in py’e's diseaseThe New England Journal of Medicine 374:2553–2562.https://doi.org/10.1056/NEJMoa1509342
-
R-spondin signaling as a pivotal regulator of tissue development and homeostasisThe Japanese Dental Science Review 55:80–87.https://doi.org/10.1016/j.jdsr.2019.03.001
-
Dynamic expression of R-spondin family genes in mouse developmentGene Expression Patterns 7:306–312.https://doi.org/10.1016/j.modgep.2006.08.006
-
Estradiol and RSPO3 regulate vertebral trabecular bone mass independent of each otherAmerican Journal of Physiology. Endocrinology and Metabolism 322:E211–E218.https://doi.org/10.1152/ajpendo.00383.2021
-
Differential activities and mechanisms of the four R-Spondins in potentiating Wnt/β-catenin signalingThe Journal of Biological Chemistry 293:9759–9769.https://doi.org/10.1074/jbc.RA118.002743
-
R-spondin1 is essential in sex determination, skin differentiation and malignancyNature Genetics 38:1304–1309.https://doi.org/10.1038/ng1907
-
R-spondins: multi-mode WNT signaling regulators in adult stem cellsThe International Journal of Biochemistry & Cell Biology 106:26–34.https://doi.org/10.1016/j.biocel.2018.11.005
-
Genetics of osteoporosis from genome-wide association studies: advances and challengesNature Reviews. Genetics 13:576–588.https://doi.org/10.1038/nrg3228
-
The role of R-spondins and their receptors in bone metabolismProgress in Biophysics and Molecular Biology 122:93–100.https://doi.org/10.1016/j.pbiomolbio.2016.05.012
-
Structural basis for R-spondin recognition by LGR4/5/6 receptorsGenes & Development 27:1339–1344.https://doi.org/10.1101/gad.219360.113
-
R-spondin1 is a high affinity ligand for LRP6 and induces LRP6 phosphorylation and beta-catenin signalingThe Journal of Biological Chemistry 282:15903–15911.https://doi.org/10.1074/jbc.M701927200
-
RSPO2-LGR5 signaling has tumour-suppressive activity in colorectal cancerNature Communications 5:3149.https://doi.org/10.1038/ncomms4149
-
Cellular signaling and biological functions of R-SpondinsCellular Signalling 24:369–377.https://doi.org/10.1016/j.cellsig.2011.09.023
-
ERK signaling is a central regulator for BMP-4 dependent capillary sproutingCardiovascular Research 76:390–399.https://doi.org/10.1016/j.cardiores.2007.08.003
Article and author information
Author details
Funding
National Institute of Arthritis and Musculoskeletal and Skin Diseases (R01AR064724)
- Roland Baron
National Institute of Dental and Craniofacial Research (R01DE029615)
- Francesca Gori
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
This work was supported by NIH-NIAMS R01AR064724 to RB and in part by NIH-NIDCR R01DE029615 to FG.
Ethics
This study was performed in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. All experiments were performed with age- and sex-matched littermates. All animals are in the C57BL/6 background and were housed in the Harvard Center for Comparative Medicine and all experimental procedures were approved by the Harvard University Institutional Animal Care and Use Committee. The protocol number associated with the ethical approval of the animal work is IS1045.
Copyright
© 2022, Nagano et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 1,147
- views
-
- 218
- downloads
-
- 14
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 14
- citations for umbrella DOI https://doi.org/10.7554/eLife.84171
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
R-spondin 3 deletion induces Erk phosphorylation to enhance Wnt signaling and promote bone formation in the appendicular skeleton
eLife 11:e84171.
https://doi.org/10.7554/eLife.84171




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}